转录调控ppt课件
合集下载
基因组学课件73基因表达的转录后调控

CHAPTER 03
基因表达的转录后调控实例
肿瘤细胞中的转录后调控
肿瘤细胞中经常出现基因表达的异常,这些异常可以通过转录后调控机制来调节。例如,某些肿瘤细胞中会出现miRNA的异 常表达,这些miRNA可以调控肿瘤相关基因的表达,从而影响肿瘤的生长和扩散。
表观遗传修饰也是肿瘤细胞中常见的转录后调控机制。DNA甲基化和组蛋白乙酰化等修饰可以影响基因的表达,从而影响肿 瘤细胞的生长和分化。
基因组学课件73基因 表达的转录后调控
目 录
• 基因表达的转录后调控概述 • 基因表达的转录后调控方式 • 基因表达的转录后调控实例 • 基因表达的转录后调控研究方法与技术 • 基因表达的转录后调控展望与挑战
CHAPTER 01
基因表达的转录后调控概述
转录后调控的定义与重要性
转录后调控定义
转录后调控是指在转录起始后,对基因表达的调控过程,包括对mRNA的加工 、修饰、转运、稳定性等方面的调控。
CHAPTER 05
基因表达的转录后调控展望 与挑战
转录后调控在疾病治疗中的应用前景
01
精准医疗
通过转录后调控机制,实现对特 定疾病的精准治疗,提高治疗效 果和减少副作用。
药物研发
02
03
个体化治疗
利用转录后调控机制,开发新型 药物,针对特定疾病进行靶向治 疗。
根据个体基因表达差异,通过转 录后调控手段,制定个性化的治 疗方案,提高治疗效果。
感谢您的观看
测序技术
测序技术是基因表达分析的重要手段 ,通过直接测定基因序列,可以深入 了解基因的表达和调控机制。
高通量测序技术如Illumina、PacBio 等,能够快速、准确地测定全基因组 的基因表达情况,为转录后调控研究 提供了强大的技术支持。
原核生物的转录与调控.ppt

例如:lac基因簇(乳糖操纵子中的三个结构基因)
原核生物操纵子中的全部结构基因从同一个启动子开 始-转半录乳成糖一苷个分多解顺酶反子的mRNA分子。
-半乳糖苷透性酶
Plac
硫代半乳糖苷乙酰转移酶
二、转录水平的调控-操纵子
3. 操纵基因:是原核生物的操纵子中调节蛋白的结合 位点,为控制结构基因转录的DNA序列。
和终止的调控。
机体可以在基因表达过程的任何阶段进行调控,一般以 转录水平上的调控为主。
二、转录水平的调控-操纵子
1. 操纵子
操纵子是原核生物基因结构、表达和调控的基本形式。 一个操纵子包括一个上游的调控区和一个以上的结构基 因组成。 调控区包含启动子和操纵基因两部分。该区控制连锁在 一起的多个基因的转录。
4. 调节基因:仅指参与其他基因表达调控的RNA和蛋 白质的编码基因。
调节基因编码的调节蛋白通过与DNA上的操纵基因结 合而控制结构基因的转录,是基因表达调控的关键。
二、转录水平的调控-操纵子
5. 反式作用因子与顺式作用元件
基因表达的产物(蛋白质或RNA)从合成的场所扩散到目 标场所而发挥作用的过程称为反式作用(trans-acting), 此基因表达产物被称为反式作用因子(trans-acting factor) 。
反式作用因子通常为蛋白质或RNA,其特征为可以从合成 地扩散到目标场所发挥作用。
顺式作用元件(cis-acting factor)是指对结构基因表达 有调节活性的DNA序列,其活性只影响与其自身同处在 一个DNA分子上的基因。顺式作用元件通常不编码蛋白 质,多位于基因旁侧或内含子中。
顺式作用(cis-acting)的概念用于任一不转变为任何其 他形式的DNA序列,它只在原位发挥DNA序列的作用, 仅影响与其物理上相连的DNA序列的活性。
《转录水平的调控》课件

转录因子在转录过程中的作用机制
激活机制
转录因子通过与DNA上的特异序 列结合,促进RNA聚合酶的招募 ,从而激活基因转录。
抑制机制
转录因子通过与DNA上的特异序 列结合,阻止RNA聚合酶的招募 ,从而抑制基因转录。
共激活剂和共抑制
因子
一些转录因子可以招募共激活剂 或共抑制因子,进一步增强或减 弱其调控作用。
转录因子在疾病中的调控作用
肿瘤发生和发展
一些转录因子在肿瘤发生和发展过程中发挥重要作用,如MYC、FOXM1等。这些转录因子的异常表达可以导致肿瘤 细胞的增殖、侵袭和转移。
免疫系统调控
一些转录因子在免疫系统的发育和功能中发挥重要作用,如NF-κB、IRF等。这些转录因子的异常表达可以导致免疫 系统紊乱,增加疾病易感性。
在转录过程中,RNA聚合酶识别DNA上的启动子 02 序列,并开始合成RNA链。
转录过程中,DNA双链结构中的一条链作为模板 03 ,合成RNA链。
转录的步骤
起始
RNA聚合酶结合到DNA上的启动子序列, 并开始合成RNA链。
延长
RNA聚合酶沿着DNA模板链不断向前移动,同时合 成RNA链。
终止
RNA聚合酶到达DNA上的终止子序列,停 止合成RNA链,并从DNA上释放出来。
表观遗传学主要包括DNA甲基化、组蛋白修饰、染色质重塑和非编码 RNA等机制。
表观遗传学调控在细胞分化、胚胎发育、肿瘤发生等多种生物学过程中发 挥重要作用。
DNA甲基化在转录水平调控中的作用
DNA甲基化是指在 DNA序列中,CpG位 点的胞嘧啶被甲基所
修饰的一种形式。
DNA甲基化可以影响 转录因子与DNA的结 合,从而调控基因的
02
基因组学课件73基因表达的转录后调控
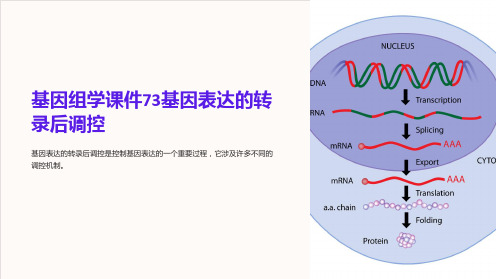
基因组学课件73基因表达的转 录后调控
基因表达的转录后调控是控制基因表达的一个重要过程,它涉及许多不同的 调控机制。
转录后调控的定义和概述
定义
转录后调控是指 转录后 对 RNA 的 进一步调控作用,进而影响基因表达。
概述
它对格局和强度进行调整,以保证基因表达适应不同的细胞状态和环境的需要。
重要性
转录后调控在许多生物学过程中都起着至关重要的作用,尤其是在细胞分化和发育过程中。
转录后调控的机制和过程
1
mRNA降解
通过降解mRNA来调整基因表达水平,这是一个快速的机制。
2
RNA剪接
将RNA中的一个或多个内含子切割掉或不保留,产生不同的变异体。
3
RNA编辑
通过改变RNA序列中的碱基,以改变RNA翻译成蛋白质的方式,从而影响蛋白质 的功能。
miRNA的调节作用
miRNA的结构
未来转录后调控的研究方向
单细胞研究
发展单细胞技术,研究基因表 达在不同类型细胞中的转录后 调控机制。
技术革新
研究新的技术平台来更好地了 解转录后调控机制,例如: Riboseq,Proteomics
物种之间的比较
比较特定物种之间的转录后调 控差异,揭示物种进化的基础, 为生物学研究提供更多的细节。
3 影响因素
转录因子的表达水平和表 达模式可以在细胞分化和 特定环境下发生变化。
转录后修饰的调节作用
1
RNA剪接后修饰
包括带电氨基酸的碳酸酐化、磷Байду номын сангаас化、去甲基化等,它们可以增强或降低RNA的 稳定性。
2
蛋白质的翻译后修饰
一些修饰会影响蛋白质的配置或功能,从而影响基因表达。
基因表达的转录后调控是控制基因表达的一个重要过程,它涉及许多不同的 调控机制。
转录后调控的定义和概述
定义
转录后调控是指 转录后 对 RNA 的 进一步调控作用,进而影响基因表达。
概述
它对格局和强度进行调整,以保证基因表达适应不同的细胞状态和环境的需要。
重要性
转录后调控在许多生物学过程中都起着至关重要的作用,尤其是在细胞分化和发育过程中。
转录后调控的机制和过程
1
mRNA降解
通过降解mRNA来调整基因表达水平,这是一个快速的机制。
2
RNA剪接
将RNA中的一个或多个内含子切割掉或不保留,产生不同的变异体。
3
RNA编辑
通过改变RNA序列中的碱基,以改变RNA翻译成蛋白质的方式,从而影响蛋白质 的功能。
miRNA的调节作用
miRNA的结构
未来转录后调控的研究方向
单细胞研究
发展单细胞技术,研究基因表 达在不同类型细胞中的转录后 调控机制。
技术革新
研究新的技术平台来更好地了 解转录后调控机制,例如: Riboseq,Proteomics
物种之间的比较
比较特定物种之间的转录后调 控差异,揭示物种进化的基础, 为生物学研究提供更多的细节。
3 影响因素
转录因子的表达水平和表 达模式可以在细胞分化和 特定环境下发生变化。
转录后修饰的调节作用
1
RNA剪接后修饰
包括带电氨基酸的碳酸酐化、磷Байду номын сангаас化、去甲基化等,它们可以增强或降低RNA的 稳定性。
2
蛋白质的翻译后修饰
一些修饰会影响蛋白质的配置或功能,从而影响基因表达。
第八章 真核基因转录调控(共84张PPT)

(一) 基因表达是基因经过转录、翻译、产生有生物活 性的蛋白质的整个过程。同原核生物一样,转录 依然是真核生物基因表达调控的主要环节。
但真核基因转录发生在细胞核(线粒体基因的转录 在线粒体内),翻译则多在胞浆,两个过程是分开 的,因此其调控增加了更多的环节和复杂性,转 录后的调控占有了更多的分量。
• (二)真核基因的转录与染色质的结构变化相 关。
❖ ④增强子的作用机理虽然还不明确,但与其他顺 式调控元件一样,必须与特定的蛋白质结合后才 能发挥增强转录的作用。
❖ 增强子一般具有组织或细胞特异性,许多增强子只在 某些细胞或组织中表现活性,是由这些细胞或组织中 具有的特异性蛋白质因子所决定的。
• ①影响模板附近的DNA双螺旋结构,导 致DNA双螺旋弯折或在反式因子的参与 下,以蛋白质之间的相互作用为媒介形 成增强子与启动子之间“成环”连接,活 化基因转录;
• 实验表明这段序列甲基化可使其后的基因不能转录,甲基 化可能阻碍转录因子与DNA特定部位的结合从而影响转录
。
• 如果用基因打靶的方法除去主要的DNA甲基化酶,小鼠的 胚胎就不能正常发育而死亡,可见DNA的甲基化对基因表 达调控是重要的。
• 由此可见,染色质中的基因转录前先要有一个被激活的过程, 但目前对激活机制还缺乏认识。
• 真核启动子一般包括转录起始点及其上游约100- 200bp序列,包含有若干具有独立功能的DNA序列元 件,每个元件约长7-30bp。
• 最常见的哺乳类RNA聚合酶Ⅱ启动子中的元件
哺乳类RNA聚合酶Ⅱ启动子中的元件序列
元件名称 共同序列
名称
结合的蛋白因子 分子量 结合DNA长度
TATAbox TATAAAA
• 三、真核基因转录水平的调控
但真核基因转录发生在细胞核(线粒体基因的转录 在线粒体内),翻译则多在胞浆,两个过程是分开 的,因此其调控增加了更多的环节和复杂性,转 录后的调控占有了更多的分量。
• (二)真核基因的转录与染色质的结构变化相 关。
❖ ④增强子的作用机理虽然还不明确,但与其他顺 式调控元件一样,必须与特定的蛋白质结合后才 能发挥增强转录的作用。
❖ 增强子一般具有组织或细胞特异性,许多增强子只在 某些细胞或组织中表现活性,是由这些细胞或组织中 具有的特异性蛋白质因子所决定的。
• ①影响模板附近的DNA双螺旋结构,导 致DNA双螺旋弯折或在反式因子的参与 下,以蛋白质之间的相互作用为媒介形 成增强子与启动子之间“成环”连接,活 化基因转录;
• 实验表明这段序列甲基化可使其后的基因不能转录,甲基 化可能阻碍转录因子与DNA特定部位的结合从而影响转录
。
• 如果用基因打靶的方法除去主要的DNA甲基化酶,小鼠的 胚胎就不能正常发育而死亡,可见DNA的甲基化对基因表 达调控是重要的。
• 由此可见,染色质中的基因转录前先要有一个被激活的过程, 但目前对激活机制还缺乏认识。
• 真核启动子一般包括转录起始点及其上游约100- 200bp序列,包含有若干具有独立功能的DNA序列元 件,每个元件约长7-30bp。
• 最常见的哺乳类RNA聚合酶Ⅱ启动子中的元件
哺乳类RNA聚合酶Ⅱ启动子中的元件序列
元件名称 共同序列
名称
结合的蛋白因子 分子量 结合DNA长度
TATAbox TATAAAA
• 三、真核基因转录水平的调控
真核生物基因转录水平的反式调控ppt

从功能上讲,植物Myb蛋白作为转录因子参与诸如
类黄酮类化合物的生物合成、毛状体发育、分生组织增 殖、GA应答以及干旱胁迫响应等信号转导过程。而编码 植物Myb蛋白的myb相关基因 是一个很大的基因家族, 因此也能调节发育进程以及参与对外界胁迫应答相关基 因的表达。
15
• 有些转录因子同时包含特异与非特异的DNA结合域, 而且后者对于目标基因的转录激活是必需的;
29
阴阳因子-1(Yin Yang-1,YY1)是一类新的转录因 子,它通过与其它蛋白发生互作从而具有转录激活和阻 遏功能,同时YY1也是核基质结合蛋白,其结合基序的 多样性可能是其功能多样性的基础。
由于YY1的结合基序核心是 CCAT、ACAT、TCAT,
其旁侧序列也具有很大的异质性,其结合基序的可变性 很大程度上允许YY1结合和影响一大批基因的转录,而 结合位点的可变性也使 YY1因子结合基序与大量其它因 子的结合基序互相靠近或重叠,从而使转录调控更加严
同,大多数bZIP类因子的亮氨酸拉链通常包含有4个或5
个7单位重复,而在拟南芥的ATB2中则有9个。 另外寡聚化结构域以外区域的差异也会影响到其功
能。
25
细胞核定位信号 真核生物转录因子是选择性的进入细胞核的,因此 转录因子也包含以富含精氨酸和赖氨酸核心肽为特征的
NLS,而碱性核心区的功能也受旁侧残基的影响。转录
• VP1是一种植物基因(包括小麦的Em基因)的转录因
子,含有一个弱的非特异性的DNA结合域-BR2(B2) 和一个序列特异的结合域BR3(B3),能够识别Sph元
件(CATGCATG),而该元件存在于Em以及其它一些
植物基因中。 • 有意思的是,VP1是通过BR2而非BR3来激活Em表达
基因转录表达调控PPT精品文档44页

The Rho transcription terminator
第二节 真核生物基因转录
真核生物具有三种RNA聚合酶;与原核生物RNA聚合酶需要σ因子来启动特定染 色体区段转录不同,真核生物通常需要多种不同起始因子来协助RNA聚合酶在特 定启动子序列有效起始转录,这些起始因子统称为普通转录因子(general transcription factors,GTFs). 在体外条件下, GTFs与RNA聚合酶足以启动特定DNA模板 的转录;而在胞内条件下,还需要其它蛋白因子如mediator复合物,DNA结合调 控蛋白及chromatin-modifying enzymes来启动促进转录。
Rho非依赖型终止子含有两段特征性序列元素, 第一段为长度20bp的 倒转重复序列; 第二段为紧随其后的富含A:T区(7-8对). 终止子本身对 转录无任何影响, 只有在其被聚合酶转录后会造成转录终止. 被转录 的倒转重复序列区段可以通过自身碱基配对形成发夹结构(Hairpin), 可以对转录三维复合物产生一种张力而容易引起复合物的解体; 当 此种茎环结构与富含A:U区相邻时, 由于A:U配对间较弱的作用力, 可以最终促使上游形成的茎环结构使转录复合物解体而终止转录.
PolⅡ core promoter
二、转录前复合物的形成
普通转录因子可以协助RNA聚合酶Ⅱ结合到启动子并协助实现从闭合复合物向
开放复合物的转化;同时还协助聚合酶脱离启动子顺利进入延伸阶段。把结合在 启动子上准备起始转录的一整套GTFs及RNA聚合酶Ⅱ称为前起始复合物(preinitiation complex). 前起始复合物的形成位点是核心启动子的TATA元素。GTFs中的TFⅡD首先通过 TBP亚基结合到TATA序列上而形成一个其它GTFs与RNA聚合酶Ⅱ对启动子结合 的平台。体外实验结果显示,其它GTFs与RNA聚合酶Ⅱ按照一定的顺序在启动子 上完成组装。
第五章转录ppt课件

一、E.coli RNA聚合酶
E.coli RNA聚合酶由5个亚基组成(5个 多肽链),即α2ββ′σ
αββ′σ四个亚基的分子量分别为36.5KDa、 150 KDa、160KDa和82 KDa,整个酶 分子的分子量为465KDa
分别是基因rpoA、rpoB、rpoC和rpoD的
产物
RNA聚合酶Ⅲ也存在于核质中,其功 能是合成tRNA和5S rRNA以及转录 Alu序列
在细胞质中也能发现一些RNA聚合酶 Ⅲ,它是从细胞核中渗漏出来的。
三种主要的RNA聚合酶的分子量都在 500 KDa左右(14S-15S),每种酶分子 含有两个大亚基和4~8个小亚基,每 个小亚基的分子量为10 KDa-90KDa
这些名称最早是依据它们从DEAE-纤维素柱上洗脱的 先后顺序而定出来的。后来发现不同生物的三种RNA聚 合酶的洗脱顺序并不相同,因而改用三种不同的RNA聚 合酶对于α-鹅膏蕈碱(α-amanitine)的敏感性不同来 进行区别。RNA聚合酶I基本不受α-鹅膏蕈碱的抑制, 在大于10-3 mol/L时才表现出轻微的抑制作用;RNA聚 合酶Ⅱ对于α-鹅膏蕈碱最为敏感,在10-9-10-8mol/L 浓度下就会被抑制;RNA聚合酶Ⅲ的敏感性介于RNA聚 合酶Ⅰ和Ⅱ之间,在10-5-10-4 mol/L时表现抑制作用。
与RNA聚合酶相结合的一个很小的蛋白质(MW= 10KDa),叫做ω亚基,其功能尚不清楚,有人认 为ω亚基对于RNA聚合酶的结构和功能没有太大的 影响。
原核生物的 RNA聚合酶
亚基 分子量
36512
150618
155613
70263
功能
决定哪些基因被转录 催化功能
大学生化课件-15.转录与基因表达调控

2.空间特异性spatial specificity
多细胞生物的个体在某一生长发育阶段,同一 基因在不同组织器官表达不同;
在个体生长、发育全过程,一种基因产物在个 体的不同组织或器官表达,即在个体不同的空间出 现。
基因表达伴随时间或阶段顺序所表达出的这种 空间分布差异,由细胞在器官的分布决定,
又称细胞特异性(cell specificity) 或组织特异性(tissue specificity)。
种类
Ⅰ
Ⅱ 最活跃的酶
Ⅲ
转录产物 存在部位
45S-rRNA 核仁
hnRNA 核质
5S-rRNA tRNA、snRNA
核仁外
对鹅膏蕈碱
的反应
耐受
极敏感
中度敏感
线粒体中与原核类似。 原核生物转录过程RNApol完成;真核生物需要一些蛋白
质的参与。
三.转录过程(起始、延长、终止)
(1)起始 a. (E.coli)RNApol全酶结合到DNA启动子
转录在胞核;翻译在胞浆。
5. 转录后修饰、加工 转录后剪接、修饰较原核复杂
第二节 基因转录的调节
转录水平的基因表达调控是调控环节中最重要的。 一、原核生物转录水平调节-- 操纵子学说 操纵子operon:原核基因组中由几个功能相关的结构基
因及其调控区组成一个基因表达的协同单位。 结构:
结构基因:编码几个功能相关的酶 调控区:启动子:RNA聚合酶结合部位
2.基因组数量 不同生物的基因组含有不同数量的基因。 细菌的基因组约4千个基因;多细胞生物基因达数万个, 人类基因组含3万~4万个基因。
在某一特定时期,基因组中只有一部分基因处于表达状态 大肠杆菌通常约5%的基因处于高水平转录活性状态,
[课件]第四讲 转录后调控及信号传导通路PPT共87页PPT
![[课件]第四讲 转录后调控及信号传导通路PPT共87页PPT](https://img.taocdn.com/s3/m/f8359cadd0f34693daef5ef7ba0d4a7302766cb3.png)
33、如果惧怕前面跌宕的山岩,生命 就永远 只能是 死水一 潭。 34、当你眼泪忍住要流出来的时候 ,睁大 眼睛, 千万别 眨眼!你会看到 世界由 清晰变 模糊的 全过程 ,心会 在你泪 水落下 的那一 刻变得 清澈明 晰。盐 。注定 要融化 的,也 许是用 眼泪的 方式。
35、不要以为自己成功一次就可以了 ,也不 要以为 过去的 光荣可 以被永 远肯定 。
44、卓越的人一大优点是:在不利与艰 难的遭遇里百折不饶。——贝多芬
45、自己的饭量自己知道。——苏联
41、学问是异常珍贵的东西,从任何源泉吸 收都不可耻。——阿卜·日·法拉兹
42、只有在人群中间,才能认识自 己。——德国
43、重复别人所说的话,只需要教育; 而要挑战别人所说的话,则需要头脑。—— 玛丽·佩蒂博恩·普尔
[课件]第四讲 转录后调控及 信号传导通路PPT
31、别人笑我太疯癫,我笑他人看不 穿。(名 言网) 32、我不想听失意者的哭泣,抱怨者 的牢骚 ,这是 羊群中 的瘟疫 ,我不 能被它 传染。 我要尽 量避免 绝望, 辛勤耕 耘,忍 受苦楚 。我一 试再试 ,争取 每天的 成功, 避免以 失败收 常在别 人停滞 不前时 ,我继 续拼搏 。
35、不要以为自己成功一次就可以了 ,也不 要以为 过去的 光荣可 以被永 远肯定 。
44、卓越的人一大优点是:在不利与艰 难的遭遇里百折不饶。——贝多芬
45、自己的饭量自己知道。——苏联
41、学问是异常珍贵的东西,从任何源泉吸 收都不可耻。——阿卜·日·法拉兹
42、只有在人群中间,才能认识自 己。——德国
43、重复别人所说的话,只需要教育; 而要挑战别人所说的话,则需要头脑。—— 玛丽·佩蒂博恩·普尔
[课件]第四讲 转录后调控及 信号传导通路PPT
31、别人笑我太疯癫,我笑他人看不 穿。(名 言网) 32、我不想听失意者的哭泣,抱怨者 的牢骚 ,这是 羊群中 的瘟疫 ,我不 能被它 传染。 我要尽 量避免 绝望, 辛勤耕 耘,忍 受苦楚 。我一 试再试 ,争取 每天的 成功, 避免以 失败收 常在别 人停滞 不前时 ,我继 续拼搏 。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
协调一致,共同表达。
四、基因表达调控的生物学意义
(一)适应环境变化、维持细胞增殖、分化 (二)维持个体生长、发育
五、基因转录激活调节基本要素
(一) 特异DNA序列 (二) 调节蛋白 (三) RNA聚合酶
(一)特异DNA序列
1. 原核生物的特异DNA序列
2. 原核生物的基因表达与调控是通过操纵子机制 实现的。 操纵子 是由功能上相关联的多个编码序列(2个以上)及 其上游的调控序列(包括操纵序列、启动序列和调 节序列)等成簇串联在一起,构成的一个转录协调 单位。
活性的特异DNA序列,称为顺式作用元件。 顺式作用元件能够被转录调节蛋白特异识别和
结合,从而影响基因表达活性。
顺式作用元件又分
启动子 增强子 沉默子
(1)启动子 是RNA聚合酶结合位点及其周围的一组转录调控
组件(包括转录起始点以及典型的TATA盒)。 (2)增强子
是增强启动子转录活性的DNA序列,并决定组织 特异性表达。 (3)沉默子
组成性基因表达 管家基因较少受环境因素的影响,在个体发育的任
一阶段都能在大多数细胞中持续表达。
(二)诱导和阻遏表达
诱导表达 在特定环境信号刺激下,基因表现为开放或 增强,表达产物增加。
阻遏表达 在特定环境信号刺激下,基因被抑制,从而 使表达产物减少。
(三)协调表达
协调表达 在一定机制控制下,功能相关的一组基因,
1. 原核生物的调节蛋白(3类) 特异因子
--
24
(4)增强子所含核苷酸序列大多为重复序列, 其内部含有的核心序列,对于它进入到另一宿 主之后重新产生增强子效应至关重要;
(5)增强子一般都具有组织和细胞特异性;
(6)增强子在DNA双链中没有5′与3′固定的方向 性;
--
25
(7)增强子可远离转录起始点,通常在 1~4 kb (个别情况可达30 kb)外起作用;
(8)增强子的活性与其在DNA双螺旋结构中的 空间方向性有关。另外,许多增强子还受到外 部信号的调控,如金属硫蛋白基因的增强子就 可对环境中的锌、镉浓度作出反应。
--
26
3、静止子
类似增强子但起负调控作用的顺式元件。静止 子与反式作用因子(蛋白质)结合后,使正调 控系统失去作用。
--
27
(二)调节蛋白
--
23
(1)它能通过启动子大幅度地增加同一条DNA链上靶 基因转录的频率,一般能增加 10~200倍,有的甚至 可达千倍。
(2)增强子的作用对同源或异源的基因同样有效,如把 SV40 的增强子连接到兔β-珠蛋白的基因上,可使转录 强度增大100倍;
(3)增强子的位置可在基因5′上游、基因内或其3′下游 的序列中,而其作用与所在基因旁侧部位的方向似无 关系,因为无论正向还是反向,它都具有增强效应;
操纵子
调节序列 启动序列 操纵序列 编码序列
I
CAP
P
O
Z YA
表达 阻遏蛋白
RNA酶聚合 结合部位
阻转遏录蛋白 结合部位
多顺反子 mRNA
编码序列
规定蛋白质结构,又称结构基因;
多顺反子mRNA
由多个结构基因串联在一起,受同一个启动序列调控,转录
生成一个mRNA, 翻译生成多个蛋白质,称此为多顺反子mRNA.
Байду номын сангаас
二、基因表达的特异性
(一)时间特异性
往往与细胞或个体的特定分化、发育阶段相适应, 又称阶段特异性。
(二)空间特异性
由细胞在各组织器官的分布差异所决定的,故又 称为细胞特异性或组织特异性。
三、基因表达的方式
(一)组成性表达
管家基因 在生命全过程都是必需的、且在一个生物个体的几
乎所有细胞中持续表达的基因。
乳糖操纵子
P 启动基因:RNA聚合酶的结合位点
O 操纵基因:阻遏物的结合位点,当阻遏物附着在操纵
基因上时,结构基因无法转录。
I: 调节基因(阻遏基因):编码阻遏物
没有乳糖的时候,阻遏基因编码产生的阻遏蛋白 结合在操纵基因序列上,使得RNA聚合酶无法对
下游的结构基因进行转录
异乳糖
原核生物的共有序列
--
19
RNA聚合酶Ⅱ启动子结构
(1)TATA框(TATA frame):其一致顺序为 TATAA(T)AA(T)。TATA框中心在-30附近,相 当于原核的-10序列(pribnow box)。
对大多数真核生物来说,RNA聚合酶与 TATA框牢固结合之后才能开始转录。 TATA框 的左右富含G┇C 序列,这就有利于该框与 RNA聚合酶形成开放性启动子复合物。
能够对基因转录起阻遏作用的DNA片段,属于 负性调控元件。
1、启动子的结构和功能
启动子 与原核启动子的含义相同,是指RNA聚 合酶结合并起动转录的DNA序列 。 但真核同启动子间不像原核那样有明显共同 一致的序列。而且单靠RNA聚合酶难以结合 DNA而起动转录,而是需要多种蛋白质因子的 相互协调作用。
原核生物的启动序列(RNA聚合酶结合位点),在距离转录起始 点-10区和-35区往往含有一些重要的保守序列(共有序列)。
-10区:含TATAAT序列,又称Pribnow盒。 -35区:含TTGACA序列。
RNA聚合酶结合部位 决定转录起始点
2. 真核生物的特异DNA序列 3. 真核生物基因组中含有可以调控自身基因表达
--
20
(2)CAAT框(CAAT frame):位置在-75附近, 一致序列为GG C(T)CAATCT。CAAT框可能 控制着转录起始的频率。
(3)GC框
在-90bp左右的GGGCGG序列称为GC框。
--
21
--
22
2、增强子的结构和功能
❖ 增强子(enhancer):又称为远上游序列(far upstream sequence) 。它是远距离调节启动子 以增加转录速率的DNA序列,其增强作用与序 列的方向无关,与它在基因的上下游位置无关。 增强子有强烈的细胞类型选择,即不同细胞类 型,增强作用不同。
基因表达调控
(Regulation of Gene Expression)
本章主要内容 ❖ 基本概念与原理
❖ 原核生物基因转录调控 ❖ 真核生物基因表达调控
一、基因表达的概念
基因表达 是指生物体基因组种结构基因所携带的遗传信息 经过转录及翻译等一系列过程,合成特定的蛋白质, 进而发挥其特定生物学功能的全过程。 基因表达产物 各种RNA(tRNA、mRNA和rRNA)以及蛋白质、 多肽。
四、基因表达调控的生物学意义
(一)适应环境变化、维持细胞增殖、分化 (二)维持个体生长、发育
五、基因转录激活调节基本要素
(一) 特异DNA序列 (二) 调节蛋白 (三) RNA聚合酶
(一)特异DNA序列
1. 原核生物的特异DNA序列
2. 原核生物的基因表达与调控是通过操纵子机制 实现的。 操纵子 是由功能上相关联的多个编码序列(2个以上)及 其上游的调控序列(包括操纵序列、启动序列和调 节序列)等成簇串联在一起,构成的一个转录协调 单位。
活性的特异DNA序列,称为顺式作用元件。 顺式作用元件能够被转录调节蛋白特异识别和
结合,从而影响基因表达活性。
顺式作用元件又分
启动子 增强子 沉默子
(1)启动子 是RNA聚合酶结合位点及其周围的一组转录调控
组件(包括转录起始点以及典型的TATA盒)。 (2)增强子
是增强启动子转录活性的DNA序列,并决定组织 特异性表达。 (3)沉默子
组成性基因表达 管家基因较少受环境因素的影响,在个体发育的任
一阶段都能在大多数细胞中持续表达。
(二)诱导和阻遏表达
诱导表达 在特定环境信号刺激下,基因表现为开放或 增强,表达产物增加。
阻遏表达 在特定环境信号刺激下,基因被抑制,从而 使表达产物减少。
(三)协调表达
协调表达 在一定机制控制下,功能相关的一组基因,
1. 原核生物的调节蛋白(3类) 特异因子
--
24
(4)增强子所含核苷酸序列大多为重复序列, 其内部含有的核心序列,对于它进入到另一宿 主之后重新产生增强子效应至关重要;
(5)增强子一般都具有组织和细胞特异性;
(6)增强子在DNA双链中没有5′与3′固定的方向 性;
--
25
(7)增强子可远离转录起始点,通常在 1~4 kb (个别情况可达30 kb)外起作用;
(8)增强子的活性与其在DNA双螺旋结构中的 空间方向性有关。另外,许多增强子还受到外 部信号的调控,如金属硫蛋白基因的增强子就 可对环境中的锌、镉浓度作出反应。
--
26
3、静止子
类似增强子但起负调控作用的顺式元件。静止 子与反式作用因子(蛋白质)结合后,使正调 控系统失去作用。
--
27
(二)调节蛋白
--
23
(1)它能通过启动子大幅度地增加同一条DNA链上靶 基因转录的频率,一般能增加 10~200倍,有的甚至 可达千倍。
(2)增强子的作用对同源或异源的基因同样有效,如把 SV40 的增强子连接到兔β-珠蛋白的基因上,可使转录 强度增大100倍;
(3)增强子的位置可在基因5′上游、基因内或其3′下游 的序列中,而其作用与所在基因旁侧部位的方向似无 关系,因为无论正向还是反向,它都具有增强效应;
操纵子
调节序列 启动序列 操纵序列 编码序列
I
CAP
P
O
Z YA
表达 阻遏蛋白
RNA酶聚合 结合部位
阻转遏录蛋白 结合部位
多顺反子 mRNA
编码序列
规定蛋白质结构,又称结构基因;
多顺反子mRNA
由多个结构基因串联在一起,受同一个启动序列调控,转录
生成一个mRNA, 翻译生成多个蛋白质,称此为多顺反子mRNA.
Байду номын сангаас
二、基因表达的特异性
(一)时间特异性
往往与细胞或个体的特定分化、发育阶段相适应, 又称阶段特异性。
(二)空间特异性
由细胞在各组织器官的分布差异所决定的,故又 称为细胞特异性或组织特异性。
三、基因表达的方式
(一)组成性表达
管家基因 在生命全过程都是必需的、且在一个生物个体的几
乎所有细胞中持续表达的基因。
乳糖操纵子
P 启动基因:RNA聚合酶的结合位点
O 操纵基因:阻遏物的结合位点,当阻遏物附着在操纵
基因上时,结构基因无法转录。
I: 调节基因(阻遏基因):编码阻遏物
没有乳糖的时候,阻遏基因编码产生的阻遏蛋白 结合在操纵基因序列上,使得RNA聚合酶无法对
下游的结构基因进行转录
异乳糖
原核生物的共有序列
--
19
RNA聚合酶Ⅱ启动子结构
(1)TATA框(TATA frame):其一致顺序为 TATAA(T)AA(T)。TATA框中心在-30附近,相 当于原核的-10序列(pribnow box)。
对大多数真核生物来说,RNA聚合酶与 TATA框牢固结合之后才能开始转录。 TATA框 的左右富含G┇C 序列,这就有利于该框与 RNA聚合酶形成开放性启动子复合物。
能够对基因转录起阻遏作用的DNA片段,属于 负性调控元件。
1、启动子的结构和功能
启动子 与原核启动子的含义相同,是指RNA聚 合酶结合并起动转录的DNA序列 。 但真核同启动子间不像原核那样有明显共同 一致的序列。而且单靠RNA聚合酶难以结合 DNA而起动转录,而是需要多种蛋白质因子的 相互协调作用。
原核生物的启动序列(RNA聚合酶结合位点),在距离转录起始 点-10区和-35区往往含有一些重要的保守序列(共有序列)。
-10区:含TATAAT序列,又称Pribnow盒。 -35区:含TTGACA序列。
RNA聚合酶结合部位 决定转录起始点
2. 真核生物的特异DNA序列 3. 真核生物基因组中含有可以调控自身基因表达
--
20
(2)CAAT框(CAAT frame):位置在-75附近, 一致序列为GG C(T)CAATCT。CAAT框可能 控制着转录起始的频率。
(3)GC框
在-90bp左右的GGGCGG序列称为GC框。
--
21
--
22
2、增强子的结构和功能
❖ 增强子(enhancer):又称为远上游序列(far upstream sequence) 。它是远距离调节启动子 以增加转录速率的DNA序列,其增强作用与序 列的方向无关,与它在基因的上下游位置无关。 增强子有强烈的细胞类型选择,即不同细胞类 型,增强作用不同。
基因表达调控
(Regulation of Gene Expression)
本章主要内容 ❖ 基本概念与原理
❖ 原核生物基因转录调控 ❖ 真核生物基因表达调控
一、基因表达的概念
基因表达 是指生物体基因组种结构基因所携带的遗传信息 经过转录及翻译等一系列过程,合成特定的蛋白质, 进而发挥其特定生物学功能的全过程。 基因表达产物 各种RNA(tRNA、mRNA和rRNA)以及蛋白质、 多肽。