包涵体蛋白的纯化和复性
包涵体纯化蛋白复性的方法操作流程

包涵体纯化蛋白复性的方法操作流程返回:包涵体复性常见问题分析与解决包涵体的纯化和复性总结包涵体折叠复性的方法应具备以下几个特点:较高的活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度的蛋白;折叠复性方法易于放大等。
蛋白复性的过程通常分为以下几个步骤:包涵体的洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体的洗涤通常选用浓度较低的变性剂,如2M尿素在50mM Tris,pH7。
0—8。
5,1mM EDTA中洗涤包涵体。
此外可以用温和去垢剂TritonX—100洗涤去除膜碎片和膜蛋白。
同时,因为去垢剂的洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度的尿素或高浓度的NaCl,使包涵体的纯度达到50%以上。
包涵体的溶解变性剂如尿素、盐酸胍,主要是通过离子间的相互作用,打断包涵体蛋白分子间的各种化学键,使多肽延伸.盐酸胍是较尿素强的变性剂,它能溶解尿素不溶的包涵体.SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内的疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸的蛋白质中分离出的包涵体,通常含有一些链间形成的二硫键和链内的非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用的二硫化物置换试剂所取代。
二硫键的形成和断裂是可逆的,直到最有利的蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素较盐酸胍慢而弱,溶解度为用尿素溶解具有不电离,呈中性,成(8-10M)70-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解的包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质的共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀对蛋白质离子交换色谱有干扰包涵体的复性复性是指通过去除变性剂使目标蛋白从完全伸展的变性状态恢复到正常的折叠结构,同时去除还原剂确保二硫键正常形成。
(整理)包涵体的分离纯化.

包涵体的纯化和复性总结(二)关于包涵体的纯化是一个令人头疼的问题,包涵体的复性已经成为生物制药的瓶颈,关于包涵体的处理一般包括这么几步:菌体的破碎、包涵体的洗涤、溶解、复性以及纯化,内容比较庞杂一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物内脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
包涵体蛋白的分离纯化

包涵体蛋白的分离纯化赵玲0743085096 包涵体是外源基因在原核细胞中表达时,尤其在大肠杆菌中高效表达时,形成的由膜包裹的高密度、不溶性蛋白质颗粒,在显微镜下观察时为高折射区,与胞质中其他成分有明显区别。
包涵体形成是比较复杂的,与胞质内蛋白质生成速率有关,新生成的多肽浓度较高,无充足的时间进行折叠,从而形成非结晶、无定形的蛋白质的聚集体;此外,包涵体的形成还被认为与宿主菌的培养条件,如培养基成分、温度、pH 值、离子强度等因素有关。
细胞中的生物学活性蛋白质常以可融性或分子复合物的形式存在,功能性的蛋白质总是折叠成特定的三维结构型。
包涵体内的蛋白是非折叠状态的聚集体,不具有生物学活性,因此要获得具有生物学活性的蛋白质必须将包涵体溶解,释放出其中的蛋白质,并进行蛋白质的复性。
包涵体的主要成分就是表达产物,其可占据集体蛋白的40%~95%,此外,还含有宿主菌的外膜蛋白、RNA聚合酶、RNA、DNA、脂类及糖类物质,所以分离包涵体后,还要采用适当的方法(如色谱法)进行重组蛋白质的纯化。
1. 包涵体的形成重组蛋白不论在原核细胞还是真核细胞中表达时,都可形成包涵体。
通常所说的包涵体是指重组蛋白在大肠杆菌中高效表达时形成的无活性蛋白聚集体,一般含有50%以上重组蛋白,其余为核糖体组分、RNA聚合酶,外膜蛋白等杂蛋白,以及质粒DNA、RNA片断、脂质、肽聚糖、脂多糖等成分]。
由于包涵体在相差显微镜下为黑色斑点, 所以也称为折射体。
包涵体形成的原因主要有以下几点: ⑴蛋白合成速度太快,以致于没有足够的时间进行折叠。
蛋白折叠的动力学模型表明:蛋白质天然构象形成的速率取决于肽链的合成速率、折叠速率和聚集速率几个因素。
中间体正确折叠是分子内的一级反应,而中间体的聚集是发生在分子间的二级或高级反应,因此,折叠中间体的浓度对聚集反应影响非常大];⑵重组蛋白是大肠杆菌的异源蛋白,由于缺少真核生物的翻译后修饰系统(如糖基化等) ,致使中间体大量积累,容易形成包涵体;⑶培养条件不佳和重组蛋白所处的环境也可导致包涵体形成,如发酵温度高,胞内pH 接近蛋白的等电点等;⑷二硫键在蛋白折叠中有重要作用,而大肠杆菌胞内的还原环境不利于二硫键的形成;⑸包涵体不溶可能由于分子间无活性的β2片层含量高于天然结构或盐沉淀蛋白。
包涵体蛋白质复性-纯化

蛋白质折叠的三态模型 (伸展态)U→(中间态)I→(自然态)N ↓ A(包涵体)
从中间体转变为天然态的过程比较缓慢。当 溶液中离子强度或变性剂浓度很低,又无其它 辅助手段存在时,聚集趋势占主导地位,导致 蛋白质的自发复性效率极低。
复性目的
复性:通过缓慢去除变性剂(避免折叠中 间体重新聚集)使目标蛋白从变性的完全伸展 状态(?)恢复到正常的折叠结构,同时去除 还原剂使二硫键正常形成。一般在尿素浓度 4M左右时复性过程开始,到2M左右时结束。
1.可溶性蛋白在细胞内容易受到蛋白酶的攻击,包涵体 表达可以避免蛋白酶对外源蛋白的降解。 2.降低了胞内外源蛋白的浓度,有利于表达量的提高。 有时甚至可以达到细胞总蛋白含量的30%
3.包涵体中杂蛋白含量较低,且只需要简单的低速离心
就可以与可溶性蛋白分离,有利于分离纯化。
4.包涵体表达的蛋白质没有活性,细胞破碎和以后复性
包涵体:在某些生长条件下,基因工程菌能
积累某种特殊的生物大分子,它们致密地集聚 在细胞内,或被膜包裹或形成无膜裸露结构, 这种水不溶性的结构称为包涵体(Inclusion B odies,IB)。
包涵体的组成及特性
一般含有50%以上的重组蛋白,其余为核
糖体元件、RNA聚合酶、外膜蛋白等,环状或
缺口的质粒DNA,以及脂体、脂多糖等,大小
Market Proportion by Host Cells
(biopharmaceutical proteins or ”Pharmateins”)
Yeast: 28%
E. coli: 54%
Animal cell: 21%
E. coli inclusion body route
occupies about a half
(整理)包涵体变性、复性及纯化

一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物内脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
这是标准配方:裂解液:50mM Tris-HCl(pH8.5~9.0),2mM EDTA,100mM NaCl,0.5%Triton X-100,1mg/ml溶菌酶。
包涵体纯化方法及包涵体蛋白制备
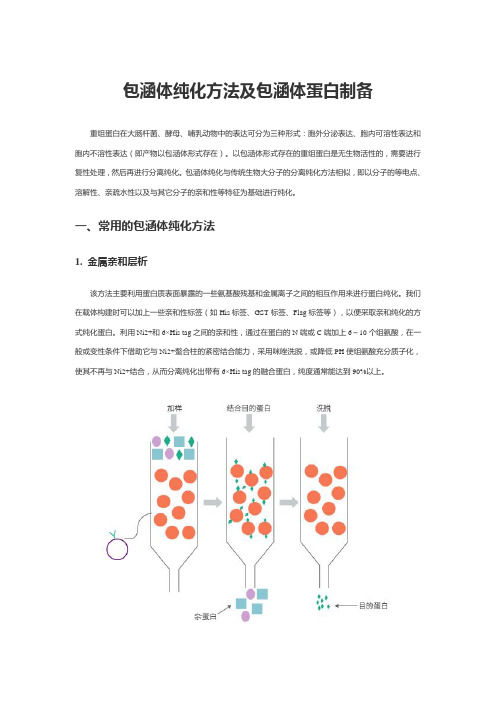
包涵体纯化方法及包涵体蛋白制备重组蛋白在大肠杆菌、酵母、哺乳动物中的表达可分为三种形式:胞外分泌表达、胞内可溶性表达和胞内不溶性表达(即产物以包涵体形式存在)。
以包涵体形式存在的重组蛋白是无生物活性的,需要进行复性处理,然后再进行分离纯化。
包涵体纯化与传统生物大分子的分离纯化方法相似,即以分子的等电点、溶解性、亲疏水性以及与其它分子的亲和性等特征为基础进行纯化。
一、常用的包涵体纯化方法1. 金属亲和层析该方法主要利用蛋白质表面暴露的一些氨基酸残基和金属离子之间的相互作用来进行蛋白纯化。
我们在载体构建时可以加上一些亲和性标签(如His标签、GST标签、Flag标签等),以便采取亲和纯化的方式纯化蛋白。
利用Ni2+和6×His tag之间的亲和性,通过在蛋白的N端或C端加上6~10个组氨酸,在一般或变性条件下借助它与Ni2+螯合柱的紧密结合能力,采用咪唑洗脱,或降低PH使组氨酸充分质子化,使其不再与Ni2+结合,从而分离纯化出带有6×His tag的融合蛋白,纯度通常能达到90%以上。
2.离子交换层析离子交换层析是根据在一定pH条件下,蛋白质所带电荷不同而进行的分离的方法。
常用的离子交换剂有羧甲基纤维素(阳离子交换剂,弱酸型)和二乙基氨基乙基纤维素(阴离子交换剂,弱碱型)。
阴离子交换基质结合带有负电荷的蛋白质,所以这类蛋白质被留在柱子上,需要通过提高洗脱液中的盐浓度等措施将其洗脱下来。
根据蛋白质结合能力不同,洗脱的速度会存在差异,通常结合较弱的蛋白质先被洗脱。
反之,阳离子交换基质会结合带有正电荷的蛋白质,可以通过提高洗脱液的PH或增加洗脱液中的盐浓度将蛋白洗脱下来。
3.凝胶过滤层析凝胶过滤层析通常又称为分子筛方法,主要是根据蛋白质的大小和形状达到分离和纯化的目的。
一般是大分子先流出来,小分子后流出来。
此方法的优点在于层析所用的凝胶属于惰性载体,不带电荷并且吸附力弱,可较广的温度范围内进行。
蛋白质、包涵体复性

目录一、脲和盐酸胍在包涵体蛋白质纯化中的作用二、包涵体变复性三、包涵体洗涤纯化——7~10四、包涵体提出、纯化和复性一、二、包涵体变复性包涵体是指细菌表达的蛋白在细胞内凝集,形成无活性的固体颗粒。
一般含有50%以上的重组蛋白,其余为核糖体元件、RNA聚合酶、内毒素、外膜蛋白ompC、ompF和ompA等,环状或缺口的质粒DNA,以及脂体、脂多糖等。
基本信息中文名称包涵体变复性复性方法稀释复性原因基因工程菌的表达产率过高包涵体变性破菌洗涤溶解目录1包涵体2包涵体变性3包涵体复性包涵体是指细菌表达的蛋白在细胞内凝集,形成无活性的固体颗粒。
一般含有50%以上的重组蛋白,其余为核糖体元件、RNA聚合酶、内毒素、外膜蛋白ompC、ompF和ompA等,环状或缺口的质粒DNA,以及脂体、脂多糖等,大小为0.5-1μm,具有很高的密度(约1.3mg/mL),无定形,呈非水溶性,只溶于变性剂如尿素、盐酸胍等。
NMR等新技术的应用表明包涵体具有一定量的二级结构,他们可能在复性的启动阶段中具有一定的作用。
包涵体的形成原因主要因为在重组蛋白的表达过程中缺乏某些蛋白质折叠的辅助因子,或环境不适,无法形成正确的次级键等原因形成的。
1.基因工程菌的表达产率过高,超过了细菌正常的代谢水平,由于细菌的δ因子的蛋白水解能力达到饱和,使之表达产物积累起来。
研究发现在低表达时很少形成包涵体,表达量越高越容易形成包涵体。
原因可能是合成速度太快,以至于没有足够的时间进行折叠,二硫键不能正确的配对,过多的蛋白间的非特异性结合,蛋白质无法达到足够的溶解度等。
2.重组蛋白的氨基酸组成:一般说含硫氨基酸越多越易形成包涵体,而脯氨酸的含量明显与包涵体的形成呈正相关。
3.重组蛋白所处的环境:发酵温度高或胞内pH接近蛋白的等电点时容易形成包涵体。
4.重组蛋白是大肠杆菌的异源蛋白,由于缺乏真核生物中翻译后修饰所需酶类和辅助因子,如折叠酶和分子伴侣等,致使中间体大量积累,容易形成包涵体沉淀。
包涵体蛋白的纯化及复性讨论专贴

包涵体蛋白的纯化及复性讨论专贴!zbbnet wrote:最近实验遇到了大麻烦,说出来大伙帮忙想想法。
实验所用菌株为BL21,载体pMAL-p2X,与MBP融合表达,目的蛋白有三对二硫键。
最初的设计思想是想通过MBP的信号肽实现可分泌表达,但实验发现主要是包涵体表达。
菌体超声破碎后12000rpm离心,所获得的沉淀不能很好的贴在离心管底,而是处于部分溶解状态,对沉淀再作如下处理:1、2M尿素洗涤,pH8.5,12000rpm离心,获得的上清不是澄清透明而是略显混浊,这次的沉淀能够牢固的贴在管底。
2、4M尿素洗涤,pH9.0,12000rpm离心,获得的上清不是澄清透明而是略显混浊,这次的沉淀为半溶状态的冻状物。
3、8M尿素溶解上述2中的沉淀,几乎不见不溶的颗粒,但溶液还是略显混浊。
上述三溶液取样电泳,均见较多的目的蛋白。
对3中的溶液取样调pH至12溶液依然混浊。
0.22um过滤,无法去除混浊物。
考虑到混浊物是多聚体,样品中加入1%SDS和0.2%β-Me,都无法改善。
做上述处理,主要考虑到混浊物是目的蛋白,因为目的蛋白设计了6HIS但不能成功的结合Ni柱。
主要想请教这样几个问题:1、有没有那位战友也碰上我上述的问题,这种问题是如何解决的?2、我所描述的包涵体的状态说明了什么问题?3、对与蛋白溶液混浊有什么好的处理方法?暂不讨论分泌表达的问题。
回答:1. 你的情况很正常,根据三次沉淀处理情况,应该可以肯定你的蛋白在8MUrea中是完全可以裂解的。
并且该蛋白如若作复性应该可溶性较佳。
但根据我个人的经验。
如果MBP融合蛋白还是以包含体表达的话是很没有意思的,建议作单独表达,除非单独表达不行。
否则即使完全复性,目的蛋白活性也大打折扣。
另可优化表达条件,有可能可容表达的,不要放弃,你的破菌上清也可以跑胶看看有没有目的蛋白。
2. 说明蛋白蔬水性不强。
3。
有些蛋白裂解后有浑浊非常正常,尤其是MBP融合的,即使做到可容也会有白乎乎浑浊的现象,只要不影响使用就行,没必要在意。
包涵体的纯化和复性情况总结

板块一、大肠杆菌(这里讨论的大肠杆菌为非分泌到培养基中的重组蛋白,是否有重组蛋白分泌到培养基中的工程菌我没有见过。
)一、关于菌体的量大肠杆菌表达的基因工程蛋白是纯化人员最方便获得的原料,对纯化工艺开发来说几乎没有原料方面的限制。
常看到有战友用个几毫升的菌液去做纯化,对此我十分不解,同样要做,为什么不多做点呢?很少的菌体会给纯化带来一些难以估计的问题,工艺的重复性和放大往往出现问题。
因此,要做个好工艺就多发酵表达一些菌体吧。
我做纯化时,初始工艺摸索用的菌体量一般为10g左右。
二、关于是否包涵体表达包涵体的定义我就不讲了。
我要讲的是,一个基因工程蛋白是否是包涵体表达的说法本身就不完全准确。
至于包涵体在电镜下的晶体颗粒表现等等对我们纯化来说毫无意义,我相信做纯化工艺的人没有谁去看这个电镜,也不关心。
我们判断的依据只是SDS-PAGE,目的蛋白在破菌沉淀中,我们就认为是包涵体表达,但这是一个似是而非的结论。
看着没问题,实际上是有毛病的。
关键在于你用的是什么破菌缓冲液!有些蛋白在用缓冲液A破菌时是在破菌沉淀中,而用缓冲液B破菌时却在破菌上清中。
缓冲液A和B的差别可能只是pH上相差1-2个单位。
那么它是包涵体表达还是可溶上清表达呢?说这个问题主要在于有些战友往往非常在意他的目标蛋白是否包涵体表达。
甚至还有包涵体表达就用专门的包涵体蛋白纯化方法等等。
我们应该关心的是目标蛋白在什么缓冲体系下是可溶的,在什么缓冲体系下是不溶的!不要让包涵体这个概念给你误导。
三、关于表达量我们常常在发表的文章上看到,我这个工程菌的目标蛋白的表达量达到菌体总蛋白的30%、50%等等。
我要说都是文章的作者在忽悠。
不知道他们是如何定量的,用的最多的大概就是SDS-PAGE的扫描分析吧。
且不说一个SDS-PAGE不能表现出所有的菌体蛋白,电泳的染色方法、染色脱色强度、照片的曝光强度、扫描分析时的条带选择等等无不对这个百分比影响巨大。
在公司的QC部门做的对此应该最有体验,20%的条带要它变成30%又有何难?我的观点是对待表达量的描述不可定量,只能定性。
包涵体纯化蛋白复性的方法操作流程

包涵体纯化蛋白复性的方法操作流程返回:包涵体复性常见问题分析与解决包涵体的纯化与复性总结包涵体折叠复性的方法应具备以下几个特点:较高的活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度的蛋白;折叠复性方法易于放大等。
蛋白复性的过程通常分为以下几个步骤:包涵体的洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体的洗涤通常选用浓度较低的变性剂,如2M尿素在50mM Tris,pH7、0-8、5,1mM EDTA中洗涤包涵体。
此外可以用温与去垢剂TritonX-100洗涤去除膜碎片与膜蛋白。
同时,因为去垢剂的洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度的尿素或高浓度的NaCl,使包涵体的纯度达到50%以上。
包涵体的溶解变性剂如尿素、盐酸胍,主要就是通过离子间的相互作用,打断包涵体蛋白分子间的各种化学键,使多肽延伸。
盐酸胍就是较尿素强的变性剂,它能溶解尿素不溶的包涵体。
SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内的疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸的蛋白质中分离出的包涵体,通常含有一些链间形成的二硫键与链内的非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用的二硫化物置换试剂所取代。
二硫键的形成与断裂就是可逆的,直到最有利的蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素较盐酸胍慢而弱,溶解度为用尿素溶解具有不电离,呈中性,成(8-10M)70-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解的包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质的共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀对蛋白质离子交换色谱有干扰包涵体的复性复性就是指通过去除变性剂使目标蛋白从完全伸展的变性状态恢复到正常的折叠结构,同时去除还原剂确保二硫键正常形成。
包涵体的纯化和复性总结--最全的前人经验

包涵体的纯化和复性总结二、包涵体的洗涤1、包涵体的洗涤问题通常的洗涤方法一般是洗不干净的,我以前是这么做的,先把包涵体用6M盐酸胍溶解充分,过滤除去未溶解的物质,注意留样跑电泳,然后用水稀释到4M,离心把沉淀和上清分别跑电泳,如此类推可以一直稀释到合适的浓度,你可以找到一个合适去除杂质的办法,其实这就是梯度沉淀的方法,我觉得比通常的直接洗脱效果好。
包涵体一般难溶解,所以你要注意未溶解的部分,你可以跑电泳对比,因为有时候难溶解的就是你的目标蛋白,所以每次处理都要把上清和沉淀跑电泳对比,免得把目标蛋白弄丢了。
此外刚处理完的包涵体好溶解。
冷冻后难溶解,溶解也需要长点时间,也需要大量的溶剂。
如果说是不少不溶解的不是你要的,那就不用管了。
2、如何得到比较纯的包涵体对于包涵体的纯化,包涵体的前处理是很重要的。
包涵体中主要含有重组蛋白,但也含有一些细菌成分,如一些外膜蛋白、质粒DNA和其它杂质。
洗涤常用1%以下的中性去垢剂,如Tween、Triton、Lubel和NP40等加EDTA 和还原剂2-巯基苏糖醇(DTT)、β-巯基乙醇等反复多次进行,因去垢剂洗涤能力随溶液离子强度升高而加强,在洗涤包涵体时可加50 mM NaCL。
这样提取的包涵体纯度至少可达50%以上,而且可保持元结构。
也可用低浓度的盐酸胍或尿素/中性去垢剂/EDTA/还原剂等洗去包涵体表面吸附的大部分不溶性杂蛋白。
洗涤液pH以与工程菌生理条件相近为宜,使用的还原剂为0.1-5mM。
EDTA为0.1-0.3 mM。
去垢剂如Triton X-100、脱氧胆酸盐和低浓度的变性剂如尿素充分洗涤去除杂质,这一步很重要,因为大肠杆菌外膜蛋白Omp T(37 KDa)在4-8mol/L尿素中具有蛋白水解酶活性,在包涵体的溶解和复性过程中可导致重组蛋白质的降解。
对于尿素和盐酸胍的选择:尿素和盐酸胍属中强度变性剂,易经透析和超滤除去。
它们对包涵体氢键有较强的可逆性变性作用,所需浓度尿素8-10M,盐酸胍6-8M。
6HIS 包涵体蛋白纯化方法

6HIS 包涵体蛋白纯化—复性方法
1.摇500ml菌至OD小于0.5时加IPTG在37℃诱导表达3小时。
2.离心,将沉淀置于-20℃冻融。
(可无)
3.用8M尿素重溶沉淀:先用0.5ml tris-cl buffer 将沉淀吹打成悬浮状,再加入20ml 8M尿素,摇匀,室温于脱色摇床上摇20分钟,充分溶解沉淀。
4.12000rpm,15℃以上离心15分钟。
5.挂柱:取上清与平衡好的镍柱柱料混合,室温于脱色摇床上摇20分钟。
6.WASH:ph8.0尿素(8M)十倍体积
7.洗脱:PH5.0
8.选浓度OD在1.0以上的来透析,透析之前加1mMDTT,两种buffer:
IB solubilization(10×):caps(3-[Cyclohexylamino]-1-propanesulfonic acid) 500mM PH11.0 N-lauroylsarcosine(30%)10×
Sometimes: 3M NDSB(10×)
9.先加到4M 尿素in Tris 50mM,并加DTT到1mM按PH8.5
10. 降至2M, 成分同上,并加150mMNaCl
11. 降到1M 同10
12. 最后透析液:Tris 50mM
NaCl 150mM
DTT 1mM
PH8.2
13.再透析去掉DTT。
包涵体变性、复性及纯化

一、菌体的裂解1、怎样裂解细菌?细胞的破碎方法1.高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3体积,盖紧筒盖,将调速器先拨至最慢处,开动开关后,逐步加速至所需速度。
此法适用于动物内脏组织、植物肉质种子等。
2.玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。
3.超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料,用大肠杆菌制备各种酶,常选用50-100毫克菌体/毫升浓度,在1KG至10KG 频率下处理10-15分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施,时间以及超声间歇时间、超声时间可以自己调整,超声完全了菌液应该变清亮,如果不放心可以在显微镜下观察。
对超声波及热敏感的蛋白和核酸应慎用。
4.反复冻融法:将细胞在-20度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。
5.化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好,我用的浓度一般为1mg/ml。
无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以抑制那些活性中心需要有疏基的蛋白水解酶的活性,加入苯甲磺酰氟化物(PMSF)也能清除蛋白水解酶活力,但不是全部,而且应该在破碎的同时多加几次;另外,还可通过选择pH、温度或离子强度等,使这些条件都要适合于目的物质的提取。
这是标准配方:裂解液:50mM Tris-HCl(pH8.5~9.0), 2mM EDTA, 100mM NaCl, 0.5% Triton X-100, 1mg /ml溶菌酶。
包涵体纯化蛋白复性的方法操作流程

包涵体纯化蛋白复性得方法操作流程返回:包涵体复性常见问题分析与解决包涵体得纯化与复性总结包涵体折叠复性得方法应具备以下几个特点:较高得活性蛋白质回收率;复性产物易于与错误折叠蛋白质分离;折叠复性后能够得到较高浓度得蛋白;折叠复性方法易于放大等。
蛋白复性得过程通常分为以下几个步骤:包涵体得洗涤包涵体在溶解之前需要进行洗涤,包涵体中主要含有重组蛋白,但也有一些杂质,如一些外膜蛋白、质粒DNA等,这些杂质会与包涵体粘连在一起,可以通过洗涤去除大多数杂质,但无法将杂蛋白去除干净。
包涵体得洗涤通常选用浓度较低得变性剂,如2M尿素在50mM Tris,pH7。
0-8。
5,1mMEDTA中洗涤包涵体。
此外可以用温与去垢剂TritonX-100洗涤去除膜碎片与膜蛋白。
同时,因为去垢剂得洗涤能力会随溶液离子强度升高而加强,所以在洗涤包涵体时可加入低浓度得尿素或高浓度得NaCl,使包涵体得纯度达到50%以上。
包涵体得溶解变性剂如尿素、盐酸胍,主要就是通过离子间得相互作用,打断包涵体蛋白分子间得各种化学键,使多肽延伸。
盐酸胍就是较尿素强得变性剂,它能溶解尿素不溶得包涵体。
SDS、正十六烷基三甲基铵氯化物、Sarkosyl等也可以破坏蛋白内得疏水键,溶解一些包涵体蛋白质。
另外,从某些含有半胱氨酸得蛋白质中分离出得包涵体,通常含有一些链间形成得二硫键与链内得非活性二硫键,这就还需加入还原剂,如巯基乙醇、二硫基苏糖醇(DTT)、二硫赤藓糖醇、半胱氨酸等。
这种还原性试剂能够同半胱氨酸形成各种二硫化物中间体,也会被复性用得二硫化物置换试剂所取代。
二硫键得形成与断裂就是可逆得,直到最有利得蛋白二硫键形成,这个平衡才会被破坏。
常用变性剂对比尿素(8-1较盐酸胍慢而弱,溶解度为7用尿素溶解具有不电离,呈中性,成0M)0-90%本低,蛋白质复性后除去不会造成大量蛋白质沉淀溶解得包涵体可选用多种色谱法纯化盐酸胍(6-8M)溶解能力达95%以上,且溶解作用快而不造成重组蛋白质得共价修饰成本高、在酸性条件下易产生沉淀、复性后除去可能造成大量蛋白质沉淀ﻫ对蛋白质离子交换色谱有干扰包涵体得复性复性就是指通过去除变性剂使目标蛋白从完全伸展得变性状态恢复到正常得折叠结构,同时去除还原剂确保二硫键正常形成、一般在尿素浓度4M左右时复性过程开始,到2M左右时结束;盐酸胍可以从4M开始,到1、5M时结束。
包涵体蛋白溶解和纯化复性

IFN-α重组蛋白包涵体溶解和蛋白纯化复性一、表达产物处理1、表达菌液8000rpm 4℃离心10min2、菌体沉淀按10:1(菌重5g,加50ml)裂解缓冲液,冰上超声至清亮(250W,超声5s,间歇5s,功率35%)3、取样取100ul 超声菌液并离心,标记超声上清,超声沉淀4、12000g 4℃离心15min,上清备用,标记超声上清5、超声沉淀用含2M 尿素的裂解缓冲液,以20:1 比例重悬,继续超声5min6、12000g 4℃离心20min7、超声沉淀用含1% Triton X-100 的裂解缓冲液重悬,4℃放置10min8、12000g 4℃离心15min,获得包涵体9、获得包涵体用Binding Bufer 重悬,4℃放置过夜二、包涵体的纯化1、放置过夜包涵体4℃高速离心,收集上清备用2、取样取离心上清,标记柱前3、使用Ni-NTA 基质进行纯化,用Binding Buffer 平衡Ni 柱,柱子平衡后低流速上样,整个上样过程使样品处于冰上,上样后用3~5 个柱体积的Wash Buffer 进行漂洗,最后用Elution Buffer 洗脱4、取样取柱后,标记柱后三、包涵体复性1、透析袋处理方法:把透析袋剪成适当长度(10~20cm)小段,在大体积的2% (W/V)NaHCO 3 和1mM EDTA (PH8.0)中将透析袋煮沸10min。
用蒸馏水彻底清洗透析袋。
放在1mM EDTA(PH8.0)中将之煮沸10min。
冷却后存放于4℃,必须确保透析袋始终浸没在溶液内。
从此时起取用透析袋时必须戴手套保持清洁,用前在透析袋内装满水然后排出,将之清洗干净2、复性采用梯度透析法,纯化后包涵体用含4M 尿素的透析缓冲液稀释蛋白浓度至约100ug/ml,从4M、2M、2M、0.5M 依次降低透析液中尿素浓度,每个浓度均4℃透析,4~6h 换新鲜透析液一次,高速离心去除沉淀。
最后用PBS(PH7.2) 4℃透析过夜。
[原创]工程菌生产的蛋白包涵体的复性与纯化-秦
![[原创]工程菌生产的蛋白包涵体的复性与纯化-秦](https://img.taocdn.com/s3/m/b6184610ef06eff9aef8941ea76e58fafbb04554.png)
[原创]工程菌生产的蛋白包涵体的复性与纯化-秦第八章工程菌生产的蛋白包涵体的复性与纯化分子生物学的发展使蛋白质的表达和生产进入到一个新的时代。
大肠杆菌由于遗传背景清晰,容易培养,可以大规模发酵,以及大量可供选择利用的克隆和高效表达载体而成为目前人们克隆和表达外源基因的首选菌株。
但是,在实际操作中重组蛋白质在大肠杆菌中的高水平表达往往导致蛋白质聚集形成不溶性的包涵体。
包涵体的形成虽有不少优点,如可溶性形式存在的外源蛋白在细胞内容易受到蛋白酶的攻击,而包涵体形式存在的外源蛋白则不易被蛋白酶降解;包涵体的形成降低了细胞内外源蛋白的浓度,有利于表达量的提高;包涵体中杂蛋白含量较低,且只需要简单的低速离心就可以与可溶性蛋白分离,有利于分离纯化;包涵体对机械搅拌和超声破碎不敏感,易于破壁,并与细胞膜碎片分离等。
但是,无活性的包涵体蛋白质需经过复性使其折叠成为具有天然构象和生物活性的蛋白质,这通常是一件很困难的事情,而且不同的蛋白质其复性方法也各不相同,因此基因重组蛋白质的复性一直是生物工程下游纯化技术的研究热点之一。
现有的生物技术研究和产业化过程表明,重组蛋白包涵体的变性及复性是重组蛋白类药物生产中最为关键的技术之一,是基因工程产品商业化的瓶颈,也是一个世界性难题。
第一节包涵体形成的原因,(什么是包涵体所谓包涵体是指由于表达部位低电势及外源蛋白质分子的特殊结构(如Cys含量高),如低电荷,无糖基化等,使得重组蛋白质与核酸,周围杂蛋白质(如核糖体元件、RNA聚合酶、内毒素、外膜蛋白质ompC、ompF和ompA等),以及脂体、脂多糖等形成的聚合体(图8-1)。
包涵体一般大小为0.1~3.0 μm,具有很高的密度(约1.3 mg/ml),无定形,呈非水溶性,只溶于尿素、盐酸胍等变性剂。
包涵体中重组蛋白质一般占包涵体总成分的50%以上,它们没有生物学活性,因为蛋白质分子没有形成正确的构象。
有报道表明成熟的分子相比包涵体内蛋白质含有更多的β结构而较少的,结构,也有报道表明包涵体中的肽链已获得与天然蛋白质分子非常接近的空间构象。
包涵体制备,变复性,纯化

制备包涵体1、超声破碎细胞2、4 ℃,12000rpm,离心10 m in, 弃上清收集包涵体.3、所得的包涵体加入1\10 体积的体积分数为1% TritonX-100液洗涤2 次4、4 ℃、12 000 rpm、离心2 m in, 沉淀即为包涵体,包涵体蛋白的变性复性方法一1、在上述包涵体中加入19、7 mL 缓冲液A (50 mmol\L Tris-HCl, 0.5 mmol\LEDTA , 0.5mmol\L DTT , 50mmol\L NaCl, 5 % 甘油) 和0.3mL 20% N-十二烷基肌氨酸钠贮存液2、剧烈搅动重悬,25OC下磁力搅拌2-3h使沉淀缓慢溶解3、4 ℃,12 000 rpm、离心10 m in, 弃沉淀4、加入PEG-6000 (终体积分数为0、2% )、氧化型谷胱甘肽(终体积分数为1mmol\L )、还原型谷胱甘肽(终浓度为2 mmol\L ),静置2h5、以pH8、0 的PBS 缓冲液透析约20 h (期间换5~ 6 次透析液) , 然后用PEG-20000 浓缩到约1 mL.方法二1、在上述包涵体中加入5mL 浓度为8mol\L 的尿素剧烈搅动, 25 ℃下用磁力搅拌器不断搅拌2~ 3 h 使沉淀缓慢溶解.2、4 ℃,12 000 rpm、离心10m in, 弃沉淀3、所得上清加入15mL pH8、0 的PBS 溶液(使尿素的浓度降低以利于复性) 以pH8、0 的PBS 缓冲液透析约20 h (期间换5~ 6 次透析液) , 然后用PEG-20000 浓缩到约1 mL.检测1、取出30 uL 备用2、向30 uL样品中加入7、5 uL 的5 倍LSB上样缓冲液, 100℃沸水处理5~10 min, 离心后取5 uL 上清上样, SDS-PAGE 检测相关试剂Triton X-100是一种比较温和的去垢剂(表面活性剂或称界面活性剂),常作为添加剂使蛋白保持稳定,尤其是膜蛋白。
包涵体蛋白体外复性的研究进展

17卷6期2001年11月生 物 工 程 学 报Chinese Journal o f Biotechnology V ol.17 N o.6N ovember 2001收稿日期:2001204213,修回日期:2001207220。
基金项目:国家863项目(8632102209204201)资助。
3通讯作者。
T el :86210264889350;Fax :86210264857285;E 2mail :hlhuang @ 包涵体蛋白体外复性的研究进展方 敏 黄华 3(中国科学院遗传研究所,北京100101)摘 要 外源基因在大肠杆菌中高水平表达时,通常会形成无活性的蛋白聚集体即包涵体。
包涵体富含表达的重组蛋白,经分离、变性溶解后须再经过一个合适的复性过程实现变性蛋白的重折叠,才能够得到生物活性蛋白。
近年来,发展了许多特异的策略和方法来从包涵体中复性重组蛋白。
最近的进展包括固定化复性以及用一些低分子量的添加剂等来减少复性过程中蛋白质的聚集,提高活性蛋白的产率。
关键词 重组蛋白,包涵体,重折叠,复性,二硫键形成中图分类号 Q51 文献标识码 C 文章编号100023061(2001)0620608205 DNA 重组技术的发展使得蛋白质的生产进入了一个新时代。
大肠杆菌表达体系因其具有低廉性、高效性和稳定性等优点在科研生产中被广泛应用。
然而,重组蛋白在大肠杆菌中的高水平表达经常导致蛋白聚集而形成不溶的、无活性的包涵体。
因此,自从应用大肠杆菌体系表达基因工程产品以来,人们就一直通过以下两个方向的研究以期望得到高活性、高产量的重组蛋白:(1)促进重组蛋白在大肠杆菌中的可溶表达:改变大肠杆菌的生长条件,或使重组蛋白与其它蛋白(如分子伴侣、折叠酶等)融合表达或共表达,或使重组蛋白分泌表达至细菌周质腔中等策略使重组蛋白在大肠杆菌中表达成可溶的生物活性形式。
(2)优化复性过程,将包涵体蛋白在体外复性得到生物活性蛋白。
包涵体的纯化和复性总结全

包涵体的纯化和复性总结二、包涵体的洗涤1、包涵体的洗涤问题通常的洗涤方法一般是洗不干净的,我以前是这么做的,先把包涵体用6M盐酸胍溶解充分,过滤除去未溶解的物质,注意留样跑电泳,然后用水稀释到4M,离心把沉淀和上清分别跑电泳,如此类推可以一直稀释到合适的浓度,你可以找到一个合适去除杂质的办法,其实这就是梯度沉淀的方法,我觉得比通常的直接洗脱效果好。
包涵体一般难溶解,所以你要注意未溶解的部分,你可以跑电泳对比,因为有时候难溶解的就是你的目标蛋白,所以每次处理都要把上清和沉淀跑电泳对比,免得把目标蛋白弄丢了。
此外刚处理完的包涵体好溶解。
冷冻后难溶解,溶解也需要长点时间,也需要大量的溶剂。
如果说是不少不溶解的不是你要的,那就不用管了。
2、如何得到比较纯的包涵体对于包涵体的纯化,包涵体的前处理是很重要的。
包涵体中主要含有重组蛋白,但也含有一些细菌成分,如一些外膜蛋白、质粒DNA和其它杂质。
洗涤常用1%以下的中性去垢剂,如Tween、Triton、Lubel和NP40等加EDTA 和还原剂2-巯基苏糖醇(DTT)、β-巯基乙醇等反复多次进行,因去垢剂洗涤能力随溶液离子强度升高而加强,在洗涤包涵体时可加50mM NaCL。
这样提取的包涵体纯度至少可达50%以上,而且可保持元结构。
也可用低浓度的盐酸胍或尿素/中性去垢剂/EDTA/还原剂等洗去包涵体表面吸附的大部分不溶性杂蛋白。
洗涤液pH以与工程菌生理条件相近为宜,使用的还原剂为0.1-5mM。
EDTA为0.1-0.3mM。
去垢剂如Triton X-100、脱氧胆酸盐和低浓度的变性剂如尿素充分洗涤去除杂质,这一步很重要,因为大肠杆菌外膜蛋白Omp T(37KDa)在4-8mol/L尿素中具有蛋白水解酶活性,在包涵体的溶解和复性过程中可导致重组蛋白质的降解。
对于尿素和盐酸胍的选择:尿素和盐酸胍属中强度变性剂,易经透析和超滤除去。
它们对包涵体氢键有较强的可逆性变性作用,所需浓度尿素8-10M,盐酸胍6-8M。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
包涵体蛋白的纯化和复性1.菌体的收集和破碎用50 ml离心管分别收集诱导表达后的培养物,8000 rpm 4℃离心10 min。
沉淀用适量的超声破菌缓冲液重悬。
【备注】超声破菌缓冲液(20 mmol/L Tris-HCl pH8.0、0.5 mol/L NaCl、1 mmol/L EDTA、1 mg/mL溶菌酶)重悬后的菌液于冰浴中进行超声波破碎,其条件为:功率为300W,占空比50%,每个循环30 s,总时间20 min。
破碎后的匀浆,4℃、12000 rpm离心15 min。
分别取上清液和沉淀进行12% SDS-PAGE分析,确定目的蛋白是否以包涵体的形式表达。
2.包涵体的处理洗涤:沉淀用适量的包涵体洗涤缓冲液重悬后,搅拌洗涤20~30 min,于4℃,12000 rpm离心15 min,弃去上清液。
重复一次。
再用适量的50 mmol/L Tris pH8.0溶液洗涤一遍(以去除残留的EDTA),于4℃12000 rpm离心15 min,弃去上清液。
【备注】包涵体洗涤缓冲液(20 mmol/L Tris-HCl pH 8.0,0.5 mol/L NaCl,2 mol/L尿素,2%Triton)2%Triton X-100溶液:量取2mlTriton X-100(聚乙二醇辛基苯基醚)液,加M缓冲液98ml即可。
溶解:沉淀用适量的包涵体溶解缓冲液重悬,室温搅拌5-6小时或过夜。
12000 rpm 4℃离心15 min,收集上清液。
【备注】包涵体溶解缓冲液(20 mmol/L Tris-HCl pH 8.0,0.5 mol/L NaCl,8 mol/L尿素,0.2 mmol/L DTT或100 mmol/L β-巯基乙醇,2%Triton)3.目的蛋白的纯化亲和层析基于目的蛋白与固相化的配基特异结合而滞留,其他杂蛋白会流过柱子。
谷胱甘肽S-转移酶(GST)是最常用的亲和层析纯化标签之一,带有此标签的重组蛋白可用交联谷胱甘肽的层析介质纯化,但本方法有以下缺点:首先,蛋白上的GST必须能合适的折叠,形成与谷胱甘肽结合的空间结构才能用此方法纯化;其次,GST标签多达220个氨基酸,如此大的标签可能会影响表达蛋白的可溶性,使形成包涵体,这会破坏蛋白的天然结构,难于进行结构分析,有时即便纯化后再酶切去除GST标签也不一定能解决问题。
另一种可应用的亲和纯化标签是6组氨酸标签,组氨酸的咪唑侧链可亲和结合镍、锌和钴等金属离子,在中性或弱碱性条件下带组氨酸标签的目的蛋白与镍柱结合,在低pH下用咪唑竞争洗脱。
组氨酸标签与GST相比有许多优点:首先,由于只有6个组氨酸,分子量很小,一般不需要用酶切去除;其次,可以在变性条件下纯化蛋白,在高浓度的尿素和胍中仍能够保持结合力;另外6组氨酸标签无免疫原性,重组蛋白可直接用来注射动物,也不影响免疫学分析。
虽然有这么多的优点,但此标签仍有不足,如目的蛋白易形成包涵体、难以溶解、稳定性差及错误折叠等。
镍柱纯化时金属镍离子容易脱落漏出混入蛋白溶液,不但会通过氧化破坏目的蛋白的氨基酸侧链,而且柱子也会非特异吸附蛋白质,影响纯化效果。
若目的蛋白可与某种碳水化合物特异结合,或者需要某种特殊的辅因子,可将该碳水化合物或辅因子固相化制成亲和柱,结合后目的蛋白可用高浓度的碳水化合物或辅因子洗脱。
融合蛋白的表达和纯化过程:1.挑单菌落于3-5 ml 2YT或LB 培养中,37℃过夜培养。
2.按1:100的比例取过夜培养液转接。
37℃扩大培养至OD 0.5左右。
3.加入IPTG终浓度0.2~0.4 mM/L。
37℃摇床培养4~6h。
4.12000g离心15min,去上清。
按40ml/100ml菌液加入1×Binding Buffer {破碎缓冲液组成:50mM Tris pH7.0~8.5左右,1mM EDTA,溶菌酶(10 礸/g cell),;或20 mmol/l Tris-HCl,pH 7.5,1mmol/lEDTA, 0.1mM PMSF, DNAse (10 礸/g cell) }细胞破碎缓冲液中不含脲等变性剂,故不溶解包涵体。
或4×PBS重悬细胞。
5.超声波破碎:占空比50%,超声15s,停15s,10~20min。
破碎后的匀浆在4℃,12,000 rpm 下离心30 min,弃去上清液。
6.洗涤包涵体:2M尿素在50mM Tris pH7.0~8.5左右,1mM EDTA ,10% Triton X-100中洗涤。
洗涤2~4h 去除膜碎片和膜蛋白。
8M尿素在50mM Tris pH7.0-8.5左右,1mM EDTA ,10% TritonX-100,二硫键还原剂中洗涤。
洗涤4~8h,使包涵体变性可溶。
7. 过Ni柱:Ni2+亲和层析柱纯化包涵体溶解后的上清液,用镍亲和层析柱纯化。
操作过程参照Amersham Pharmacia Biotech公司提供的操作指南进行。
具体过程如下:转移树脂到柱子,待完全沉淀后,用三蒸水洗涤镍亲和层析柱,以除尽基质中的乙醇和空气,并防止下一步Ni2+沉淀;5×柱体积的Charge Buffer充电;20×柱体积的三蒸水洗涤,去尽游离的Ni2+;10×柱体积的Binding Buffer平衡柱子;手工上样,根据蛋白的浓度来确定上样体积;20×柱体积的Binding Buffer 洗涤,至检出无蛋白,并收集流出液,用于12%SDS-PAGE电泳检测;6×柱体积的Wash Buffer 洗涤,至检出无蛋白;Elution Buffer洗脱,每次1 mL,收集洗脱流出液,洗脱至检出无蛋白;10×柱体积的Binding Buffer平衡。
此时,即可用于纯化同种蛋白,不需重新充电。
纯化操作完成后,加5×柱体积的Strip Buffer洗涤,去尽Ni2+;10×柱体积的三蒸水洗涤去尽EDTA;加5×柱体积的0.5 mol/L NaOH,静置0.5-2 h,去沉淀的蛋白质;三蒸水洗涤,至流出液的pH值为7.0;20%乙醇洗涤,再加适量20%乙醇,于4℃保存。
A、过柱前的注意事项:a、样品必须澄清、无颗粒物,否则会堵塞柱子、缩短其使用寿命;b、包含体洗涤的最后一步及溶解时所用的Buffer中不能含EDTA、DTT、2ME;c、确保样品及Binding Buffer中有适量的NaCl,以防止杂离子与Ni离子竞争;B、过Ni柱的操作步骤:①用DDW洗涤,洗去20%的乙醇、除尽基质中的空气,并能防止下一步中Ni离子沉淀;②⑩5×柱体积的Charge Buffer(50mM NiSO4)充电;③5×柱体积的DDW洗涤,去游离的Ni离子;④10×柱体积的Binding Buffer(20mM Tris-HCl,0.5M NaCl,5mM咪唑,pH8.0)平衡;⑤上样(手工上样,样品体积应根据目的蛋白的浓度来确定);⑥10×柱体积的Binding Buffer洗涤,至无蛋白检出,用三氯醋酸检测,收集流出液;⑦Elution Buffer(20mM Tris-HCl,0.5M NaCl,500-1000mM咪唑,pH8.0)洗脱,每次1ml,应呈现正态分布(两边稀,中间浓);⑧10×柱体积的Binding Buffer洗涤。
此时,即可用于纯化同种蛋白,很少需要重新充电。
⑨用20%的乙醇充满柱子,保存在4℃。
注:同种蛋白纯化5~10次后,应进行strip和re-charge。
C、柱的清洗与保存(最好每天清洗一次):(1)去Ni离子:①5×柱体积的Strip Buffer(20mMTris-HCl,0.5M NaCl,50mM EDTA,pH7.4)洗涤;②10×柱体积的DDW洗涤。
(2)去沉淀的蛋白质:①5×柱体积的0.5M NaOH装满柱子,静置0.5~2hr;②DDW洗涤,至流出液的pH值为7。
(3)保存:20%乙醇洗涤,再加20%乙醇保存于4℃。
D、柱的再生:当流速很慢、充电后基质不变成蓝绿色时,应进行柱的再生。
① 2×柱体积的6M盐酸胍洗涤,然后3×柱体积的DDW洗涤;② 1×柱体积的2%SDS洗涤;③ 5×柱体积的0.5M NaOH装满柱子,静置0.5~2hr;④ 5×柱体积的DDW洗涤,然后5×柱体积的100mM EDTA,pH8.0洗涤;⑤ 3×柱体积的DDW洗涤,然后3×柱体积的20%乙醇洗涤;⑥ 20%乙醇保存于4℃。
【备注】Ni2+亲和层析柱纯化缓冲液(1×Buffer):?/P>4.目的蛋白的SDS-PAGE分别取加有IPTG的空载体pET22(b+)表达的上清液、未加IPTG和加IPTG诱导的重组子全提取物、超声波破碎后的上清液和沉淀(即包涵体)、纯化后的蛋白溶液,进行12%SDS-PAGE分析。
通过蛋白质分子量标准,判断目的蛋白的相对分子质量,并对纯化后的蛋白进行纯度分析。
5、蛋白质的复性目的蛋白在大肠杆菌中以包含体形式存在,要获得有活性的产物,必须对纯化的目的蛋白进行复性。
将收集的洗脱流出液装入透析袋,对含适量甘油的0.01 mol/L PBS pH8.0透析48 h,每4~8 h更换一次PBS溶液。
然后,将上述透析袋包埋在聚乙二醇粉末中,对蛋白溶液进行浓缩。
【备注】PBS配方:配制0.01 M PH 7.4 PBS 2L:0.2 mol/L Na2HPO4(mL) 81.00.2 mol/L NaH2PO4(mL) 19.0NaCL 17 g用水稀释到2 L即可。
另外,可加入下列重折叠(复性)介质:①氧化还原系统:还原/氧化型谷胱甘肽(10:1~3:1)②L-Arg或Gly(0.5mol/L)③甘油:10%④聚乙二醇(浓缩)⑤高摩尔浓度的Tris(0.4~1mol/L)。