内膜系统与蛋白质分选
细胞生物学 第七章_内膜系统与蛋白质的分选
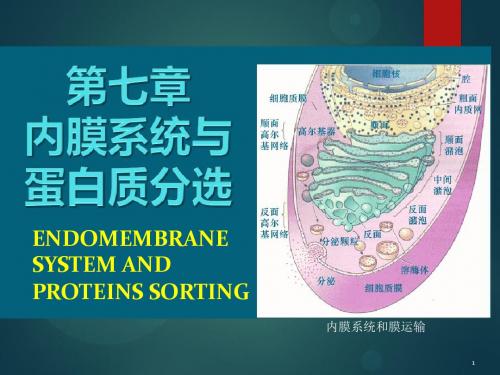
将细胞内部区分为不同的功能区域,保证各种生化反应所需的独
特的环境。
2
主要内容
内膜系统 内质网 高尔基体 溶酶体
蛋白质分选及其基本原理 膜泡运输
3
第一节 内膜系统 Endomembrane System
Endomembrane System : The structural and functional relationship organelles
混合功能氧化酶将底物羟基化 P354, Fig. 9-16
(1)被氧化的有毒底物(脂溶性)与细胞色素P-450结合; (2)细胞色素P-450中的铁原子被 NADPH还原; (3)氧与细胞色素P-450结合; (4)底物被一个氧原子氧化,另一个氧用于形成 水。被氧化的有毒底物带上羟基,由脂溶性转变为水溶性被排出体外。
sER合成磷脂酰胆碱 P352, Fig. 9-14
★ 内质网膜中脂肪酸与胞质溶胶中的磷酸甘油结合,然后脱磷生成甘油脂肪酸; ★ 在胆碱磷脂转移酶的作用下,将胞质中的CDP-胆碱与甘油脂肪酸结合形成磷脂酰胆碱; ★ 新合成的磷脂酰胆碱在内质网膜中磷脂转位酶的作用下翻转到内质网的腔面。
15
Detoxification of organiccompounds in liver cells. System of oxygenases ---cytochrome p450 family
O-linked: linked to the hydroxyl group serine or threonine via GalNac (in Golgi)
11
Quality control of newly synthesized proteins ---The role of N-linked glycosylationin ER protein folding
第九章 内膜系统与蛋白质分选和膜运输

第九章内膜系统与蛋白质分选和膜运输教学目的1、掌握信号肽假说和蛋白质转运的机制。
2、掌握高尔基体参与细胞分泌活动的作用。
3、掌握细胞内蛋白质的分选。
教学内容本章从以下6个方面讨论了细胞质质基质与内膜系统:1.细胞质膜系统及其研究方法2.内质网3.高尔基复合体4.溶酶体5.细胞的分泌与内吞作用6.小泡运输的分子机理计划学时及安排本章计划6学时。
教学重点和难点真核细胞在进化上一个显著特点就是形成了发达的细胞质膜系统,将细胞内环境分割成许多功能不同的区室。
内膜系统是指内质网、高尔基体、溶酶体和液泡(包括内体和分泌泡)等四类膜结合细胞器,因为它们的膜是相互流动的,处于动态平衡,在功能上也是相互协同的,其中包括膜运输系统。
本章是细胞生物学的重点章,包括六个方面的内容,其中内质网及信号肽假说、小泡运输的分子机理是本章的关键内容。
1.内质网是内膜系统中的重要膜结合细胞器,主要分清光面内质网和粗面内质网在功能上的差异。
对于粗面内质网,重点是信号肽假说和蛋白质转运的机制。
2.高尔基复合体是内膜系统中参与蛋白质加工与分选的细胞器,要求了解和掌握高尔基体参与细胞分泌活动的作用,即将内质网合成的多种蛋白质进行加工、分类与包装,然后分门别类地运送到细胞的特定部位或分泌到细胞外。
理解高尔基体在细胞内物质运输中所起的交通枢纽作用。
3.关于溶酶体,要求掌握溶酶体膜的稳定性、溶酶体的类型及特点、溶酶体的功能、溶酶体的生物发生。
4.细胞内蛋白质的分选是本章的核心内容之一,重点学习和掌握运输小泡的类型和分选信号、披网格蛋白小泡形成的机理、COP-被膜小泡形成的机理、小泡的定向运输、停靠和融合机理。
通过本章的学习要充分了解细胞内部结构的动态关系,蛋白质合成和分选的机制和“流水”作业的模式,从中获得启发。
教学方法讲授、讨论教学过程9.内膜系统与蛋白质分选和膜运输9.1 细胞质膜系统及其研究方法9.1.1 膜结合细胞器与内膜系统■ 膜结合细胞器的种类和功能● 膜结合细胞器种类与数量(表)● 膜结合细胞器的功能(表)● 膜结合细胞器在细胞内的分布(图)■ 内膜系统的动态性质内膜系统的最大特点是动态性质(图),这就使内膜系统的结构处于一个动态平衡。
chapter9内膜系统与蛋白质分选和膜运输

信号肽与起始跨膜序列
52
内部信号序列
53
内部信号序列
不位于N-末端,但具信号序列的作用 可作为蛋白质共翻译转移的信号被SRP识别,同时它也 是起始转移信号。 是不可切除的,又是疏水性, 是膜蛋白的一部分,如 果共翻译转运蛋白质中只有一个内含信号序列,那么合 成的蛋白就是单次跨膜蛋白
54
单次跨膜蛋白的定向I
在内质网合成的蛋白和 脂通过分泌活动进入分泌小 泡运送到工作部位(包括细胞 外);细胞通过内吞途径将细 胞外的物质送到溶酶体降解。
8
内膜系统的进化
◆起因∶
●遗传信息的扩大 ●体积的增大 ●表面积与体积比值失调 ●物质代谢速度受限
◆对策:细胞内部结构区域化 ◆途径:
●内共生途径 ●细胞质膜内陷途径。
11
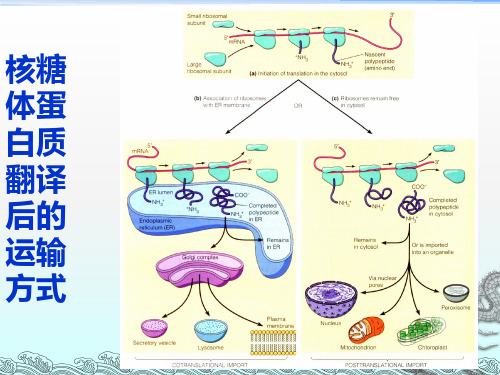
◆细胞中蛋白质的运输有两种方式: ●共翻译运输和翻译后运输
◆内膜系统参与共翻译运输,是分泌 蛋白质分选的主要系统。
12
细胞质中蛋白质合成和空间定位
细胞中各部位(包括细胞质)中 的蛋白质都是来自胞质溶胶,
内质网以上的细胞器,包括 细胞核、线粒体、过氧化物酶 体和质体所需蛋白是由胞质溶 胶直接运送的。
将内质网上合成的蛋白质或脂类转运到高尔基体。
●有两个面
内质网的外表面称为胞质溶胶面(cytosolic space), 内表面称为潴泡面(cisternal space)
26
内质网与核膜的关系
27
◆特殊类型的内质网
●微粒体(microsome)
在细胞匀浆和差速离心过程中获得的由破碎的内 质网自我融合形成的近似球形的膜囊泡状结构,
4
● 膜结合细胞器的功能
• 细胞核 基因组存在场所,DNA和RNA的合成地 • 内质网 大多数脂的合成场所,蛋白质合成和集散地 • 高尔基体 蛋白质和脂的修饰、分选和包装 • 溶酶体 细胞内的降解作用 • 内体 内吞物质的分选 • 线粒体 通过氧化磷酸化合成ATP • 叶绿体 进行光合作用 • 过氧化物酶体 毒性分子的氧化
chapter+9++内膜系统与蛋白质分选和膜运输

Chapter 9 内膜系统与蛋白质分选和膜运输9.1 内膜系统概述9.1.1 内膜系统与膜结合细胞器1. 内膜系统(endomembrane systems)内膜系统是指内质网、高尔基体、溶酶体和液泡(包括内体和分泌泡)、细胞核等五类膜结合细胞器, 因为它们的膜是相互流动的, 处于动态平衡, 在功能上也是相互协同的。
广义上的内膜系统概念也包括线粒体、叶绿体、过氧化物酶体等细胞内所有膜结合的细胞器。
2. 膜结合细胞器(membrane-bound organelles)或膜结合区室(membrane-bound compartments)指细胞质中所有具有膜结构的细胞器,包括细胞核、内质网、高尔基体、溶酶体、分泌泡、线粒体、叶绿体和过氧化物酶体等。
由于它们都是封闭的膜结构,内部都有一定的空间,所以又称为膜结合区室。
除核糖体和细胞骨架。
3. 膜结合细胞器的功能:①胞质溶胶代谢的主要场所;蛋白质合成部位②细胞核基因组存在场所,DNA和RNA的合成地③内质网大多数脂的合成场所,蛋白质合成和集散地④高尔基体蛋白质和脂的修饰、分选和包装⑤溶酶体细胞内的降解作用⑥内体内吞物质的分选⑦线粒体通过氧化磷酸化合成ATP⑧叶绿体进行光合作用⑨过氧化物酶体毒性分子的氧化4. 内膜结构特点与动态性质①独立性A 内膜封闭的区室B 执行独立的功能②动态性质A合成途径: 分泌性蛋白与脂的合成和加工内质网高尔基体B 分泌途径:内质网上合成的蛋白质和脂出芽小泡向外运输C内吞途径:细胞外的物质吞进内体和溶酶体内膜系统的动态特性:内膜系统将细胞中的生化合成、分泌和内吞作用连接成动态的、相互作用的网络。
在内质网合成的蛋白和脂通过分泌活动进入分泌小泡运送到工作部位(包括细胞外);细胞通过内吞途径将细胞外的物质送到溶酶体降解。
5. 内膜系统的进化①起因∶A 遗传信息的扩大B 体积的增大C 表面积与体积比值失调D 物质代谢速度受限②对策:细胞内部结构区域化③途径:A 内共生途径B 细胞质膜内陷途径。
9 内膜系统与蛋白质分选.

9.内膜系统与膜运输真核细胞在进化上一个显著特点就是形成了发达的细胞质膜系统, 将细胞分成许多膜结合的区室,包括细胞核、内质网、高尔基体、溶酶体、内体和分泌泡等。
虽然这些区室具有各自独立的结构和功能,但它们又是紧密相关的,尤其是它们的膜结构是相互转换的,这种转换的机制则是通过蛋白质分选(protein sorting)和膜运输实现的(图9-1)。
图9-1 内膜系统和膜运输9.1 细胞质膜系统及其研究方法9.1.1膜结合细胞器与内膜系统关于真核细胞中具有膜结构的细胞器的总体描述通常有三个概念:膜结合细胞器(membrane-bound organelles)或膜结合区室(membrane-bound compartments)细胞质膜系统(cytoplasmic membrane system)内膜系统(endomembrane systems)虽然这三个概念都是指真核细胞中具有膜结构的细胞器,但是在含义上仍有一些差别。
膜结合细胞器的种类和功能膜结合细胞器种类与数量原核细胞内只有一个区室就是胞质溶胶(cytosol)。
真核细胞内有许多膜结合的区室,但与胞质溶胶相比, 所占比例都很小(表9-1)。
表9-1 肝细胞中主要膜结合细胞器的体积比细胞器每细胞所含数量细胞内的百分比胞质溶胶 1 54线粒体1700 22内质网 1 12细胞核 1 6高尔基体 1 3过氧化物酶体400 1溶酶体300 1内体200 1膜结合细胞器的功能膜结合细胞器在细胞的生命活动中具有重要作用(表9-2)。
表9-2 真核细胞膜结合区室的主要功能细胞器(区室) 主要功能胞质溶胶代谢的主要场所;蛋白质合成部位细胞核基因组存在的场所,DNA和RNA的合成地内质网大多数脂的合成场所,蛋白质合成和集散地高尔基体蛋白质和脂的修饰、分选和包装溶酶体细胞内的降解作用内体内吞物质的分选线粒体通过氧化磷酸化合成ATP叶绿体进行光合作用过氧化物酶体毒性分子的氧化在这些膜结合的细胞器中,线粒体、叶绿体、过氧化物酶体和细胞核等的独立性很强,并且有特别的功能;其他几种膜结合细胞器,如内质网、高尔基体、溶酶体和小泡,虽然有不同的结构和功能,但是它们都参与蛋白质的加工、分选和膜泡运输,形成了一个特别的细胞内系统。
05-5-内膜系统完整性与蛋白质分选

6.4 内膜系统完整性与蛋白质分选同学们好!前面我们分别阐述了内膜系统主要的细胞器是如何进行生物合成、加工、分选、包装、运输和消化分解等过程。
下面我们以蛋白质分选和交通运输为例,来谈谈细胞内膜系统结构和功能的整体性及蛋白质分选转运机制。
一、内膜系统整体性内膜系统的区域化、囊泡运输及膜流动等体现了内膜系统结构与功能的整体性。
1.区域化是内膜系统整体性的结构基础。
细胞内所有物质的合成与代谢活动都有各自的定位,细胞内膜系统的区域化为此构筑了结构基础。
真核细胞内部空间是由内膜分隔成无数相对独立的小房间,即细胞器,相应的蛋白质或酶便被局限在这些特定的细胞器中,起到对细胞的区域化作用。
这种区域化作用不仅增加了细胞内有限空间的表面积,同时使复杂的生理生化反应过程和特定的生物学功能分别在特定的空间、彼此独立地有序进行,减少相互干扰,提高了细胞的整体代谢水平、功能效率和可调控性。
这些区域化的结构在功能和发生上紧密联系,在细胞内物质代谢过程中分工协作,为细胞整体生命功能提供结构保障。
这就像一栋建筑,只有合理配置每个房间的效用,才能凸显其整体性。
2.囊泡运输是内膜系统功能整体性的运行方式。
细胞生命活动依赖于胞内运输系统,细胞内的定向物流是由囊泡运输方式来实现的,囊泡运输是生物物质在细胞膜性结构之间按时空进行分拣、装卸、运送、加工与拆分等事件的运行方式。
囊泡是真核细胞中十分常见的膜泡结构,是细胞中专门负责蛋白质运输的各种膜性小泡的总称。
囊泡运输是一个受精密调控而高度有序的物质转运过程,是真核细胞特有的一套完整的物质转动形式。
归纳起来,囊泡运输有如下功能特点:1)它是生物物质在细胞质中定向运输的基本途径;2)它是一个高度有序并受到严格选择和控制的物质转运过程;3)它是实现细胞膜及内膜系统功能结构转换和代谢更新的桥梁;4)它是生物物质特异性卸载和回收再利用的基本保证。
如此,在细胞整个生命活动中,囊泡往返穿梭于各细胞器之间,实现生物物质的定时、定点、定量配置。
内膜系统与蛋白质分选-细胞生物学

内膜系统与蛋⽩质分选(protein sorting) 蛋⽩质是由核糖体合成的,合成之后必须准确⽆误地运送到细胞的各个部位。
在进化过程中每种蛋⽩形成了⼀个明确的地址签(address target),细胞通过对蛋⽩质地址签的识别进⾏运送,这就是蛋⽩质的分选(protein sorting)。
细胞中蛋⽩质的运输有两种⽅式:共翻译运输和翻译后运输,内膜系统参与共翻译运输,是分泌蛋⽩质分选的主要系统。
■蛋⽩质分选定位的时空概念 所谓蛋⽩质分选定位的时空概念包括两种含义:①合成的蛋⽩质何时转运?②合成蛋⽩质在细胞中定位空间及转运中所要逾越的空间障碍是什么? ●从时间上考虑,蛋⽩质的合成分选有两种情况:先合成,再分选和⼀边合成⼀边分选。
为了适于蛋⽩质分选的时间上的需要,核糖体在合成蛋⽩质时就有两种存在状态:游离的或与内质结合的。
●从蛋⽩质定位的空间看,包括了细胞内各个部分,即使是具有蛋⽩质合成机器的线粒体和叶绿体也需要从细胞质中获取所需蛋⽩质。
细胞中各部位(包括细胞质)中的蛋⽩质都是来⾃胞质溶胶,不过内质以上的细胞器,包括细胞核、线粒体、过氧化物酶体和质体所需蛋⽩是由胞质溶胶直接运送的。
⽽从内质以下的各种细胞器,包括内质、⾼尔基体、溶酶体、内体、分泌泡、细胞质膜以及细胞外基质等所需的蛋⽩质虽然起始于胞质溶胶,但要经过内质和⾼尔基体的中转。
■蛋⽩质分选定位的空间障碍及运输⽅式 从蛋⽩质定位的细胞内空间部位结构来看,可分为三种类型:①没有膜障碍的,如胞质溶胶,包括胞质溶胶中的细胞⾻架蛋⽩和各种酶及蛋⽩分⼦;②有完全封闭的膜障碍,如线粒体、叶绿体、内质、⾼尔基体等;③有膜障碍,但是膜上有孔,如细胞核。
根据三种不同的空间障碍,合成的蛋⽩质通过三种不同⽅式进⾏运输定位(图9-6)。
●核孔运输(transport through nuclear pore)胞质溶胶中合成的蛋⽩质穿过细胞核内外膜形成的核孔进⼊细胞核,被运输的蛋⽩需要有核定位信号。
内膜系统与蛋白质的分选

蛋白质的分选
• 蛋白质分选的信号假说
新合成的蛋白质分子内包含有某种分拣 信号,该分拣信号具有决定蛋白质在细 胞内的去向或定位作用。 信号假说具有普遍性。
• 蛋白质的分拣信号与运输途径
蛋白质的分拣信号是一段特殊的氨基 酸序列
游离核糖体合成蛋白质
号 号氨 (核 ) (基 核定 端 输位 端有 入信 的 信号 螺螺 号 旋旋 ) 信信 细胞核 N
内膜系统与细胞内蛋白质的分 选
内膜系统
内膜系统(endomembrane systems)是指内质网、高尔基体、溶酶体和液泡 (包括内体和分泌泡)等四类膜结合细胞器, 因为它们的膜是相互流动的, 处于 动态平衡, 在功能上也是相互协同的。 (广义上的内膜系统概念也包括线粒体、叶绿体、过氧化物酶体、细胞核 等细胞内所有膜结合的细胞器。)
• 使蛋白质分子能准确定位到功能部位, 使其能准确行使其生物学功能; • 分选过程中伴随着对蛋白质分子的加 工和修饰,使真核细胞蛋白质分子的 结构和功能更加多样化。
谢谢观赏
第8小组
调节性分泌小泡 溶酶体
蛋 白 质 的 运 输 方 式
主要有三种:
门孔运输 跨膜运输 囊泡运输
门孔运输 定 位
位 置
跨膜运输
线粒体,过氧化 物酶体,微体, 内质网内的
囊泡运输
细胞核内
高尔基复合 体,溶酶体, 内吞体
核膜上的核孔 复合体
CELL器膜上的蛋 白质传导通道
囊泡
蛋白质分选的生物学意 义
• 提高细胞对蛋白质的合成和利用效率;
内膜系统区域化(房室化):内膜系统的各细胞器在细胞内形成了一些相互 分隔膜性区室。
细胞核 胞质溶胶 胞质溶胶 线粒体 内质网
内膜系统、蛋白质分选和膜泡运输

击 此
七 白 处
添
质 章 加
正 文
, 文
分 字 真 是
您
选 思 核 想 与 的
提
细 炼 膜 ,
请
胞 言 泡 简 内 意
赅
运 的 膜 阐 输 述 系 您
的
观 点
统
。
、
目录
壹
细细 胞胞 内质 膜基 系质 统的 及涵 其义 功与 能功
能
貳
内 质 网
叁
高 尔 基 体
肆
溶 细酶 胞体 内与 蛋过 白氧 质化 的物 分酶 选体 与 膜 泡 运 输
具有异质性,由单层膜围绕而成。
过氧化物酶体中常含有两种酶:
○ 依赖于黄素(FAD)的氧化酶,其作用是将底物氧化形成 H2O2;
○ 过氧化氢酶,作用是将H2O2分解,形成水和氧气。
Peroxisome of hepatocyte
特征 形态大小
酶种类 pH 值 是否需O2 功能 发生
识别的标志酶
异质性(heterogenous)的细胞器 。
初级溶酶体(primary lysosome)
由高尔基体分泌形成, 含多种酸性水解酶。
ቤተ መጻሕፍቲ ባይዱ
次级溶酶体(secondary lysosome)
是正在进行或完成消化作 用的溶酶体,分为自噬溶 酶体和异噬溶酶体。由初 级溶酶体与胞吞泡或自噬 泡或异噬泡融合形成。
溶酶体 多呈球形,直径0.2~0.5μm, 无酶晶体 酸性水解酶 5 左右 不需要 细胞内的消化作用 酶在粗面内质网合成经高尔基体 出芽形成 酸性水解酶等
微体 球形,哺乳动物细胞中直径多在 0.15~ 0.25μm,内常有酶的晶体 含有氧化酶类 7 左右 需要 多种功能 酶在细胞质基质中合成,经分裂 与组装形 成 过氧化氢酶
第六章细胞内膜系统与蛋白质分选

第六章细胞内膜系统与蛋白质分选与原核细胞物不同的是真核细胞具有复杂的由内膜构成的功能区隔。
细胞内膜系统指在结构,功能或发生上相关的细胞内膜形成的细胞结构,包括核被膜、内质网、高尔基体及其形成的溶酶体和分泌泡等,以及其它细胞器如线粒体,质体和过氧化物酶体等膜包围的细胞器(膜性细胞器)。
内膜系统形成了一种胞内网络结构,其功能主要在于两个方面:其一是扩大膜的总面积,为酶提供附着的支架,如脂肪代谢、氧化磷酸化相关的酶都结合在细胞膜上。
其二是将细胞内部区分为不同的功能区域,保证各种生化反应所需的独特的环境。
本章主要介绍内质网、高尔基体、溶酶体、过氧化物酶体的功能和蛋白质分选,关于线粒体、叶绿体和细胞核的功能与蛋白质分选将分别在第七章(线粒体与叶绿体)和第十二章(细胞核与染色体)中讲解。
第一节内质网由KR. Porter、A. Claude 和EF. Fullam等人于1945年发现,他们在观察培养的小鼠成纤维细胞时,发现细胞质内部具有网状结构,建议叫做内质网endoplasmic reticulum,ER,后来发现内质网不仅仅存在于细胞的“内质”部,通常还有质膜和核膜相连,并且与高尔基体关系密切,并且常伴有许多线粒体。
一、形态与组成内质网膜约占细胞总膜面积的一半,是真核细胞中最多的膜。
内质网是内膜构成的封闭的网状管道系统。
具有高度的多型性。
可分为粗面型内质网(rough endoplasmic reticulum,RER,图6-20)和光面型内质网(smooth endoplasmic reticulum,SER,图6-21)两类。
RER呈扁平囊状,排列整齐,膜围成的空间称为ER腔(lumen),膜外有核糖体附着。
SER呈分支管状或小泡状,无核糖体附着。
肌肉细胞中的肌质网是一种特化的SER,称为肌质网,可贮存Ca2+,引起肌肉收缩。
细胞不含纯粹的RER或SER,它们分别是ER连续结构的一部分。
ER主要功能是合成蛋白质和脂类,分泌性蛋白和跨膜蛋白都是在ER中合成的。
【细胞生物学】内膜系统与蛋白质分选和膜运输2
●狭义:高尔基体外侧网络的分选作用
●分选作用主要 是由信号序列和 受体之间的相互 作用决定的
9.4 溶酶体(lysosome)
◆动物细胞内的一种细胞器
◆小球状,直径一般0.25~0.8μm, 由一层单位膜包被
◆溶酶体含有50多种水解酶类,
在细胞内起消化和保护作用
◆当细胞被损伤时,溶酶体可
释放出水解酶类,使细胞自溶
生产加工流水线
酶 顺面 脂肪酰基转移酶 √ 甘露糖苷酶 Ⅰ √ 乙酰葡萄糖胺转移酶 Ⅰ 甘露糖苷酶 Ⅱ NADPase 磷酸酶 腺苷酸环化酶 √ 5'核苷酶 √ 酸性磷酸酶 核苷二磷酸酶 唾液酸转移酶 硫胺素焦磷酸酶 半乳糖基转移酶
中间膜囊 反面
√ √ √ √ √ √
√ √ √ √ √ √ √
高尔基体的功能
●N-乙酰葡萄糖苷酶(内质网) ●N-乙酰葡萄糖胺磷酸转移酶(TGN)
信号斑
◆M6P受体蛋白(M6P receptor protein)
●M6P 受体蛋白在 pH 为 6.5 ~ 7 的条件下与 M6P 结合 ●在酸性条件下(pH=6)脱落 ●M6P受体蛋白主要存在于高尔基体外侧网络 ●在某些细胞中 M6P 受体蛋白也存在于细胞质 膜上。?
甘油三酯和磷脂 解毒作用 P450的作用 Ca2+离子浓度的调节作用 Ca2+-ATP酶
糖原分解与游离葡萄糖释放
葡萄糖磷酸化酶 葡萄糖磷酸易位酶 葡萄糖-6-磷酸酶
乙酰辅酶A
磷脂的合成
脂酰辅酶A 磷酸甘油 二酰甘油
翻转酶
光面内质网的解毒作用
在混合氧化酶、NADPH、O2共同作用下
脂溶性有毒物
从CGN向TGN的运输
高尔基潴泡的融合和
真核细胞内膜系统蛋白质分选和

真核细胞内膜系统蛋白质分选和
真核细胞内膜系统蛋白质分选和
分泌性蛋白的合成与转运
➢游离核糖体上信号肽的合成
➢SRP识别信号肽,信号识别颗粒(SRP)-核糖体复合体形成,翻译暂停
程主 要 过
➢核糖体与内质网膜结合,形成SRP-SRP受体-核糖体复合体 ➢SRP脱离并参加再循环,核糖体翻译继续进行
➢信号肽被切除,成熟的蛋白质落入内质网腔
动物和真菌细胞内。
真核细胞内膜系统蛋白质分选和
第三节 高尔基复合体 Golgi complex
一、高尔基复合体的形态结构 二、高尔基复合体的化学组成 三、高尔基复合体的功能 四、高尔基复合体的来源
真核细胞内膜系统蛋白质分选和
一、高尔基复合体的形态结构
真核细胞内膜系统蛋白质分选和
一、高尔基复合体的形态结构
主要特点: 细胞质基质是一个高度有序的体系;
通过弱键而相互作用处于动态平衡的结构体系。
真核细胞内膜系统蛋白质分选和
细胞质基质的功能
➢ 完成各种中间代谢过程 ➢ 蛋白质的分选与运输 ➢ 与细胞质骨架相关的功能 ➢ 蛋白质的修饰
真核细胞内膜系统蛋白质分选和
细胞质基质的功能
蛋白质的修饰
(1)辅基或辅基与酶的共价结合 (2)磷酸化与去磷酸化,调节很多蛋白质的生物活性 (3)糖基化。糖基化主要发生在内质网和高尔基体中 (4)对某些蛋白质的N端进行甲基化修饰 (5)酰基化
二、高尔基复合体的化学组成
蛋白质 60% ;脂类 40%
集中在高尔基复合体中的酶,有硫氨素焦磷酸酶和若 干糖基转移酶,也含有酸性磷酸酶和其它溶酶体的酶。 其中糖基转移酶是高尔基复合体最具特征的酶,他能把 低聚糖转移到蛋白质上形成糖蛋白。
细胞生物学总结(复习重点)——7.内膜系统、蛋白质分选、膜泡运输

1、细胞质基质:真核细胞的细胞质中除去细胞器和内含物以外的、较为均质半透明的液态胶状物称为细胞质基质或胞质溶胶。
4、内膜系统:细胞内在结构、功能乃至发生上相关的、由膜围绕的细胞器或细胞结构的统称,主要包括内质网、高尔基体、溶酶体、胞内体、分泌泡等。
2、微粒体:为了研究ER的功能,常需要分离ER膜,用离心分离的方法将组织或细胞匀浆,经低速离心去除核及线粒体后,再经超速离心,破碎ER的片段又封合为许多小囊泡(直径约为100nm),这就是微粒体。
3、糙面内质网:细胞质内有一些形状大小略不相同的小管、小囊连接成网状,集中在胞质中,故称为内质网。
内质网膜的外表面附有核糖体颗粒,则为糙面内质网,为蛋白质合成的部位。
核糖体附着的膜系多为扁囊单位成分,普遍存在于分泌蛋白质的细胞中,其数量随细胞而异,越是分泌旺盛的细胞中越多。
5、分子伴侣:细胞中,这类蛋白能识别正在合成的多肽或部分折叠的多肽,并与多肽的一定部位相结合,帮助这些多肽的转移、折叠或组装,但其本身并不参与最终产物的形成。
6、溶酶体:溶酶体几乎存在于所有的动物细胞中,是由单层膜围绕、内含多种酸性水解酶类、形态不一、执行不同生理功能的囊泡状细胞器,主要功能是进行细胞内的消化作用,在维持细胞正常代谢活动及防御方面起重要作用。
7、残余小体:在正常情况下,被吞噬的物质在次级溶酶体内进行消化作用,消化完成,形成的小分子物质可通过膜上的载体蛋白转运至细胞质中,供细胞代谢用,不能消化的残渣仍留在溶酶体内,此时的溶酶体称为残余小体或三级溶酶体或后溶酶体。
残余小体有些可通过外排作用排出细胞,有些则积累在细胞内不被排出,如表皮细胞的老年斑、肝细胞的脂褐质。
8、蛋白质分选:细胞中绝大多数蛋白质均在细胞质基质中的核糖体上开始合成,随后或在细胞质基质中或转至糙面内质网上继续合成,然后,通过不同途径转运到细胞的特定部位并装配成结构与功能的复合体,参与细胞的生命活动的过程。
又称定向转运。
细胞内膜系统与蛋白质分选

返回内质网 -Lys-Asp-Glu-Leu-COO-
蛋白质分选运输机制
门控运输(gated transport) 跨膜运输(transmembrane transport) 膜泡运输(vesicular transport)
蛋白质分选运输机制
蛋白质分选运输机制——膜泡运输——信号肽假说
G. Blobel 等 1 9 7 5 年 提 出 了 信 号 假 说 ( Signal hypothesis),认为蛋白质N端的信号肽,指导蛋白质转 至内质网上合成,因此获1999年诺贝尔生理医学奖。
输入过氧化 物酶体
-Ser-Lys-Leu-COO-
输入内质网 +H3N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-AlaThr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-Val-Phe-Gln-
Prelysozyme
Amino Acid Met-Lys-Trp-Val-Thr-Phe-Leu-Leu-Leu-Leu-
Phe-Ile-Ser-Gly-Ser-Ala-Phe-Ser↓Arg...
Met-Asp-Met-Arg-Ala-Pro-Ala-Gln-Ile-PheGly-Phe-Leu-Leu-Leu-Leu-Phe-Pro-Gly-
蛋白质合成的区域化——膜泡运输——信号肽假说
SRP受体(SPR receptor):内质网膜的整合蛋白,异二聚体, 可与SRP特异结合。
转位因子(translocator,translocon),由3-4个Sec 61蛋白 构成的通道,每个Sec61由3条肽链组成。
蛋白质合成的区域化——膜泡运输——信号肽假说
第五章细胞内膜系统与蛋白质分选

蛋白质都是在核糖体上合成的,但是有些蛋白质在合成开始不久后 便转在内质网上合成。包括:①向细胞外分泌的蛋白 ②跨膜蛋白 ③构成细胞器中的可溶性驻留蛋白
(二)蛋白质的修饰与加工
修饰包括糖基化、羟基化、酰基化、二硫键形成等 糖基化类型: N-连接的糖基化 :起始于内质网,完成于高尔基体 O-连接的糖基化:主要发生在高尔基体上
2020/9/26
2020/9/26
图4-16 两类分选信号
二、蛋白质分选运输的途径与类型
1.蛋白质分选的两条基本途径 (1)翻译后转移:在细胞质基质中完成多肽链的
合成,再转运至细胞器(如线粒体、叶绿体、 微体、细胞核等)或细胞质基质的特定部位。 (2) 翻译共转移:在细胞质基质中多肽链的合成 起始后,边合成边转入内质网腔中,随后经高 尔基体运至溶酶体、质膜、分泌到细胞外。
• 动力素是一种GTP酶,它将小泡柄部的膜尽可能地 拉近,调节小泡以出芽形式脱离膜。
2020/9/26
2020/9/26
图4-18 笼形衣被小泡的组成
网格蛋白分子由3个重链和3个轻链组成,形成一个具有3个曲臂的 形状。许多网格蛋白的曲臂部分交织在一起,形成一个具有5边形 网孔的笼子。
202图0/94/2-619 网格蛋白的结构,A电镜照片,B分子模型 ,C衣被模型
2020/9/26
图4-8 胰岛素分子的加工成熟
第三节 溶酶体与过氧化物酶体
一、溶酶体的结构 二、溶酶体的功能 三、溶酶体的发生 四、溶酶体与疾病 五、过氧化物酶体
2020/9/26
一、溶酶体的结构类型
溶酶体是单层膜围绕、内含多种酸性水解酶类的囊泡状细 胞器。其主要功能是进行细胞内的消化作用。 • 溶酶体膜的特征:
(细胞生物学(王金发版)章节总结)第九章 内膜系统与蛋白质分选和膜运输

第九章内膜系统与蛋白质分选和膜运输9.1内膜系统9.1.1内膜系统概述9.1.2内质网9.1.3高尔基体9.1.4溶酶体9.2细胞的分泌和内吞作用9.2.1细胞分泌过程9.2.2细胞内吞过程9.2.3膜泡运输机制9.3膜的生物发生9.3.1膜脂9.3.2膜整合蛋白和外周蛋白9.3.3脂锚定蛋白形成9.1内膜系统9.1.1内膜系统概述(1)内膜系统的组成内质网、高尔基体、溶酶体、细胞核、液泡。
功能上紧密联系,形成协调统一的整体。
(2)内膜系统的结构特点既相互独立,又相互间处于动态平衡状态。
通过三种途径:生化合成途径;分泌途径;内吞途径。
(3)内膜系统的重要功能—蛋白质分选蛋白质分选三种途径:核孔运输;跨膜运输;膜泡运输。
信号肽指导内膜系统蛋白质的运输,其对蛋白质没有特异性,对不同的膜结合细胞器具有特异性。
(4)内膜系统的进化与生物学意义原因:遗传信息量增大;细胞体积增大;表面积与体积比下降;物质代谢速度受限。
途径:内共生途径;质膜内陷。
生物学意义:形成了特定的功能区域与微环境,有不同的物质浓度及代谢系统,合理利用了资源,工作效率上升;通过各种活动,形成统一整体。
(5)内膜系统的研究方法放射性自显影;差速离心分离与功能分析;突变技术;绿色荧光蛋白定位法。
9.1.2内质网(1)结构与组成平行扁平囊泡(粗面内质网)或管状囊泡(光面内质网)组成。
粗面内质网可与核膜、质膜结构连续,外表面称为胞质溶胶面,内表面为潴泡面。
标志酶为葡萄糖-6-磷酸酶。
(2)功能①光面内质网糖原分解释放葡萄糖;类固醇激素的合成;脂的合成与转运;解毒作用(P450);钙库。
②粗面内质网膜结合核糖体的蛋白质运输:信号假说。
信号序列,SRP识别信号肽、停止翻译、识别停靠蛋白,停靠蛋白,蛋白质运输通道,袢环状过膜。
起始转移信号(信号序列及内部信号序列)与蛋白质运输通道受体位点结合,停止转运信号和内部信号序列决定穿膜次数。
信号序列被信号肽酶切除,内部信号序列保留。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
G-1-P
Glycogen granules in a hepatocyte.
G-1-P
葡 萄 糖 磷 酸 变 位 酶
G-6-P
G-6-P
葡萄糖-6-磷酸酶
葡萄糖 + Pi
◇类固醇激素的合成 光面内质网能够合成 胆固醇,然后将胆固 醇氧化、还原、水解 进一步转变成各种类 固醇激素。
some organs (adrenal gland and gonads, for example) use cholesterol as a precursor for steroid hormone production 甲羟戊酸
活化的异戊二烯
三十碳六烯
孕烯醇酮 黄体酮 肾上腺酮 睾丸激素
皮质醇
◇信号假说的补充 • Signal recognition particle, SRP
• 停靠蛋白(docking protein, DP) a. 是SRP在内质网膜上的受体蛋白; b. 两个亚基:亲水的α亚基, 疏水的β亚基. •易位子( translocon) a. Multiple copies of the Sec61 complex assemble to form a cylinder translocon channel b. 5~6 nm high and 8.5 nm in diameter, with a central pore, roughly 2 nm in diameter c. 哺乳动物细胞中有三种类型的Sec61, 即α、β 和γ d. Sec61α有10个跨膜α螺旋
膜结合细胞器的种类和功能
肝细胞中主要膜结合细胞器的体积比 细胞器 胞质溶胶 线粒体 内质网 细胞核 高尔基体 过氧化物酶体 溶酶体 每细胞所含数量/个 1 1700 1 1 1 400 300 体积比/% 54 22 12 6 3 1 1
(胞)内体
200
1
真核细胞膜结合区室的主要功能
细胞器(区室) 胞质溶胶 细胞核 内质网 高尔基体 溶酶体 (胞)内体 线粒体 叶绿体 过氧化物酶体
The formation of coated vesicles in a cell-free system
◇Study of genetic mutants
9.1.3 内膜系统与蛋白质分选 ◇蛋白质分选定位的时空概念 •合成的蛋白何时转运? •蛋白质在细胞中的定位
◇蛋白质分选定位的三种机制 •Transport through nuclear pore
• 信号肽(signal peptide):指导内膜系统的蛋白质 运输.
信号序列对蛋白质没有特异性
9.2 内质网(endoplasmic reticulum) 9.2.1 形态结构和化学组成 ◇类型
1.Rough endoplasmic reticulum (RER) 2.Smooth endoplasmic reticulum (SER) 3.特殊类型的内质网 微粒体(microsome)
易位子
◇蛋白质共翻译转运的机制—信号假说
◇膜蛋白的共翻译 转运机理
•Start-transfer signal
在跨膜时保持正电荷 一端朝向胞质溶胶面
•内部信号序列与单次跨膜蛋白
•终止转移肽与单次跨膜蛋白
•二次跨膜蛋白与多次跨膜蛋白
◇Bip蛋白在ER蛋白质的转运和装配中的作用
◇蛋白质在内质网中的修饰 •N-糖基化(protein glycosylation) 指在rER上合成的蛋白与寡糖 基共价连接
•羟基化 除了N-糖基化以外,新生肽的脯 氨酸和赖氨酸生在少数蛋白质身上.
Protein glycosylation in the rough ER
•形成脂锚定蛋白
GPI:糖基磷脂酰肌醇
•双硫键的形成
PDI可以切断二硫键形成自由能最低的构象,以帮助新合成的 蛋白质重新形成二硫键并处于正确折叠的状态。
9.2.3 糙面内质网的功能—蛋 白质合成与转运
◇信号序列的发现和证实
•Redman和Sabatini用分离 的RER小泡研究膜结合核 糖体合成的蛋白质去向. •1971年美国洛克菲勒大 学的Blobel等提出了信号 序列(signal sequence) 的概念。
•信号序列的间接证据 1972年, Milstein 和他的同事发现: a.从细胞质分离的IgG轻链:多20个氨基酸残基 b.释放到细胞外的IgG轻链:少20个氨基酸残基
主要功能 代谢的主要场所,蛋白质合成部位 基因组存在场所,DNA和RNA的合成地 脂的合成场所,蛋白质合成和集散地 蛋白质和脂的修饰,分选和包装 细胞内降解作用 内吞物的分选 氧化磷酸化位点 光合作用 毒性分子的氧化
膜结合细胞器在细胞内的分布是有层次的.
内膜系统的特点与动态性质
◇特点 •内膜封闭的区室 •执行独立的功能 ◇动态性质 •Biosynthetic pathway •Secretory pathway •Endocytosis pathway
Silver grains Rough ER
Secretory granule
◇Green Fluorescent Protein (GFP) GFP is a small protein which was encoded by a gene of jellyfish. GFP
The eleven βstrands form the staves of a barrel.
•Across membrane transport
•Vesicle translocation
◇信号序列指导蛋白质的正确运输 信号序列的类型 • 入核信号(nuclear localization signal):指导核蛋 白的运输;
• 引导肽(leading peptide):指导线粒体,叶绿体,和 过氧化物酶体蛋白的运输;
雌二醇
醛甾酮
◇脂的合成与转运 •合成甘油三脂(triacylglycerol)
•磷脂合成 在光面内质网上合成的磷脂先作为内质网膜的构 成成分,然后再转运到其他的膜上.
•磷脂转运
◇解毒作用(detoxification) 光面内质网含有丰富的氧化酶系统(如细胞色素 P450、NADH细胞色素C还原酶等)能使许多有害 物质由脂溶性转变成水溶性而被排出体外.此反应 主要在肝细胞的sER中进行. ◇Ca2+的调节作用 肌质网是细胞内特化的光面内质网,有高浓度的 钙结合蛋白,每个钙结合蛋白可以结合30个左右 的Ca2+.
To use this technique, DNA encoding GFP is fused to DNA encoding the protein to be studied and the resulting chimeric DNA is introduced into cells that can be observed under the microscope.
•信号肽的证实
a. 加与不加rER小泡,产物不同 b. 蛋白水解酶水解实验 c. 多聚核糖体的离体翻译
蛋 白 水 解 酶 水 解 实 验
多聚核糖体实验
◇信号序列的一般特征及早期信号假说
• 一般特征
长度: 15~35 氨基酸残基;
结构: N端含1或多个带正电荷氨基酸,其后是 6~12个连续的疏水残基; 功能: 在蛋白质合成中将核糖体引导到内质 网,进入内质网后通常被切除. •1975年,Blobel等正式提出了信号假说
膜结合细胞器的进化
◇起因∶ •遗传信息的扩大 •体积的增大 •表面积与体积比值失调 •物质代谢速度受限 ◇对策:细胞内部结构区域化 ◇途径: •内共生途径 •细胞质膜内陷途径。
9.1.2 A few approaches to the study of cytomembranes
◇Autoradiography
VSV: vesicular stomatitis virus
疱疹病毒
◇Biochemical Analysis of Subcellular Fractions
Vesicles are isolated from endomembranes by cell fractionation and then their composition and functional properties can be determined by biochemical analysis and immunoelectromicroscopy. For example, enzymatic activities and the capacities of transporting newly synthesized proteins of the microsomes and the role of Golgi complex in the stepwise assembly of complex carbohydrates.
1. 合成始于游离核糖体; 2. N 端信号序列靠自由碰撞与内质网膜接触 , 然后 插入内质网的膜; 3. 蛋白质以袢环形式穿过内质网的膜; 4. 蛋白的存在方式: a.若信号序列被信号肽酶切除:完全进入腔 b.若是停止转移信号: 膜蛋白
Blobel received the Nobel Prize for medicine in 1999
9.3 高尔基体
Golgi body, Golgi apparatus,Golgi complex)
9.3.1 高尔基体的功能
9.3.1.1 蛋白质和脂的运输 • ER与高尔基体内侧面间的运输
•蛋白质从高尔基体内侧网络向外侧网络运输
9.3.1.2 蛋白质的糖基化 • 高尔基体对蛋白质的糖基化修饰作用是在高尔基 体的各个特异的功能区室中进行.
第九章 内膜系统与蛋白质分选和膜运输