(完整版)酵母细胞
(完整版)酵母细胞

酵母细胞表面展示技术是一种真核蛋白表达系统。
其基本原理是将外源靶蛋白基因与特定的载体基因序列融合后导入酵母细胞,融合蛋白诱导表达后,信号肽引导融合蛋白向细胞外分泌,外源蛋白的展示主要是借助于GPI锚的作用,GPI锚定蛋白的C端包含疏水多肽,当细胞蛋白被合成,前体蛋白通过羧基末端的疏水序列锚定在内质网膜上,其余蛋白则位于内质网腔中。
在极短的时间内,疏水序列在转酰氨基酶的作用下ω位点裂开同时被GPI锚置换,并被运输到高尔基体再通过分泌途径分泌至细胞膜外。
在蛋白水解酶作用下,分泌信号序列被切除,细胞蛋白被磷脂酰肌醇特异性磷脂酶C从细胞膜上释放并以GPI锚定形式与葡聚糖共价相连被转运至细胞壁外。
图1展示的是通过α-凝集素系统转运目的蛋白到细胞表面的分泌途径。
酵母表面展示技术具有表达偏差小,展示的融合蛋白相对分子质量范围大,分子数量多,筛选、纯化、活性测定方便等优点。
目前,最常见的酵母表面展示系统包括:凝集素展示表达和絮凝素展示表达[5]。
如图2所示,α-凝集素展示表达系统(图2A)是将外源基因与酵母编码α-凝集素C端编码序列连接后插入质粒载体的分泌信号肽(Secretion signal,SS)下游,融合蛋白诱导表达后,信号肽引导嵌合蛋白向细胞外分泌,由于融合蛋白C-末端存在含GPI锚定信号(GPIanchor attachment signal)的凝集素多肽序列,可将蛋白锚定在酵母细胞壁上,从而将蛋白分子展示表达在酵母细胞表面;a-凝集素展示系统(图2B)与α-凝集素展示系统不同的是其具有由二硫键连接的Aga1和Aga2两个亚基,目的蛋白通过与Aga2形成融合蛋白,再通过Aga1末端的GPI结构使目的蛋白表达于细胞表面;絮凝素展示表达系统利用絮凝素基因(flo1p)与外源基因融合,并将融合蛋白展示表达在细胞表面,此系统又包括两个子系统:GPI锚定系统(图2C)和絮凝结构域(Flo1p flocculation functional domain)锚定系统(图2D)。
(完整版)微生物学第三章真核微生物

第七授课单元一、教学目的通过该授课单元的学习,要求学生了解真核微生物和真菌所包括的主要类群,熟悉真核生物的细胞构造,掌握真核生物与原核生物的主要差别;熟悉单细胞真菌——酵母菌的细胞形态和大小、细胞结构、繁殖方式、生活史、菌落特征,并了解工业上常见的有益和有害的酵母菌。
二、教学内容第三章真核微生物的形态、构造和功能第一节真核微生物的概述第二节酵母菌一、分布及与人类关系二、细胞形态与构造三、酵母菌的繁殖方式和生活史四、酵母菌的菌落五、工业生产上常见的有益和有害的酵母菌三、教学重点、难点及其处理重点:1.真核生物的定义、细胞构造和生物学特性(1)定义:是一大类进化程度较高,细胞核具有核膜,能进行有丝分裂,细胞质中存在线粒体或同时存在叶绿体等细胞器的生物。
(2)真核生物的细胞结构:放在介绍酵母菌和霉菌相关章节中介绍。
(3)真核生物与原核生物的比较:真核生物在一系列生物学特性上均与原核生物有着明显的不同,主要差别如细胞核是真核或是原核;具膜细胞器的有无;细胞壁的成分是纤维素、几丁质还是肽聚糖、假肽聚糖;鞭毛的构造是复杂还是简单;以及细胞体积的大小等。
真核生物的细胞器都有完整的膜包围,如内质网、高尔基体、溶酶体、微体、线粒体和叶绿体等,而其细胞核更是发展到有膜包裹、具精巧染色体构造,其中的双链DNA长链还与组蛋白密切结合,成为结构十分精巧的遗传功能单位(通过列表的方式比较真核生物与原核生物的主要差别)。
2.真核微生物和真菌所包括的主要类群真核微生物主要包括属于植物界的微藻、属于动物界的原生动物和属于菌物界的真菌等微小生物。
真菌主要包括酵母菌(单细胞真菌)、霉菌(小型丝状真菌)和蕈菌(能形成大型肉质子实体)。
本章重点讨论酵母菌和霉菌。
“菌物界”的概念由我国学者裘维蕃1990年提出,指与动物界、植物界相并列的一大群无叶绿体、依靠细胞表面吸收有机养料、细胞壁一般含几丁质的真核微生物。
一般包括真菌、粘菌和假菌三类。
(完整版)海藻酸钠固定酵母细胞实验资料

海藻酸钠固定酵母细胞实验资料1实验原理海藻酸钠是应用最广泛的水溶性海藻酸盐。
海藻酸钠遇到钙离子可迅速发生离子交换,生成凝胶。
利用这种性质,将海藻酸钠溶液滴入含有钙离子的水溶液中可产生海藻酸钙胶球。
本实验便是将酵母细胞与海藻酸钠溶液混匀后,通过注射器或相似的滴注器将上述混合液滴入CaCl2溶液中,Ca2+从外部扩散进入海藻酸钠与细胞混合液珠内,使海藻酸钠转变为不溶于水的海藻酸钙凝胶,由此将酵母细胞包埋在其中。
2实验操作过程中的几个注意点2.1控制好配制海藻酸钠溶液的火候、浓度,实验成败的关键步骤配置海藻酸钠溶液。
加热使海藻酸钠溶化是操作中重要的一环,涉及到实验的成败。
教材提示是要用小火或者间断加热,反复几次,直到完全熔化。
在实验操作中,教师要提醒学生按照教材的提示进行,不然会发生焦糊现象。
笔者多次实际操作发现酒精灯上隔石棉网加热,匀速搅拌基本不会发生焦糊现象,而且需要时间略长,为节省时间可选用电炉加热,加快熔化,但一定要间断加热,可以用试管夹移动烧杯。
完全熔化的海藻酸钠成半透明状,最好要无颗粒无气泡,类似于平常家庭中冲调好食用的藕粉。
海藻酸钠的浓度涉及到固定化细胞的质量。
如果海藻酸钠浓度过高,将很难形成凝胶珠;如果浓度过低,形成的凝胶珠所包埋的酵母细胞的数目少,影响后续实验效果。
教材提示是0.7 g海藻酸钠,加入10 mL蒸馏水,边加热边搅拌,直至完全熔化,用蒸馏水定容至10 mL。
实际操作中,很多学生忘记定容,也有教师认为不需要定容。
其实在加热搅拌的过程中,蒸馏水会蒸发,加热时间越长失去的蒸馏水越多,所以加热熔化结束后一定要定容,不然浓度肯定偏大,影响后续步骤中凝胶珠的形状。
2.2控制海藻酸钠溶液与酵母细胞混合时的温度,保证酵母细胞的活性海藻酸钠溶液与酵母细胞混合,一定要将溶化好的海藻酸钠溶液冷却至40℃以下,通常让学生自己用手感觉烧杯壁,感觉不到烫手就差不多。
再加入已活化的酵母细胞,进行充分搅拌,使其混合均匀。
(完整版)酵母菌实验
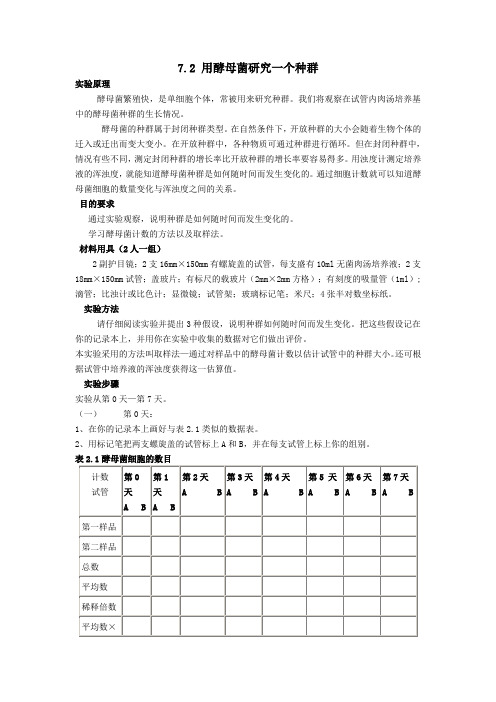
7.2 用酵母菌研究一个种群实验原理酵母菌繁殖快,是单细胞个体,常被用来研究种群。
我们将观察在试管内肉汤培养基中的酵母菌种群的生长情况。
酵母菌的种群属于封闭种群类型。
在自然条件下,开放种群的大小会随着生物个体的迁入或迁出而变大变小。
在开放种群中,各种物质可通过种群进行循环。
但在封闭种群中,情况有些不同,测定封闭种群的增长率比开放种群的增长率要容易得多。
用浊度计测定培养液的浑浊度,就能知道酵母菌种群是如何随时间而发生变化的。
通过细胞计数就可以知道酵母菌细胞的数量变化与浑浊度之间的关系。
目的要求通过实验观察,说明种群是如何随时间而发生变化的。
学习酵母菌计数的方法以及取样法。
材料用具(2人一组)2副护目镜;2支16mm×150mm有螺旋盖的试管,每支盛有10ml无菌肉汤培养液;2支18mm×150mm试管;盖玻片;有标尺的载玻片(2mm×2mm方格);有刻度的吸量管(1ml);滴管;比浊计或比色计;显微镜;试管架;玻璃标记笔;米尺;4张半对数坐标纸。
实验方法请仔细阅读实验并提出3种假设,说明种群如何随时间而发生变化。
把这些假设记在你的记录本上,并用你在实验中收集的数据对它们做出评价。
本实验采用的方法叫取样法—通过对样品中的酵母菌计数以估计试管中的种群大小。
还可根据试管中培养液的浑浊度获得这一估算值。
实验步骤实验从第0天—第7天。
(一)第0天:1、在你的记录本上画好与表2.1类似的数据表。
2、用标记笔把两支螺旋盖的试管标上A和B,并在每支试管上标上你的组别。
表2.1酵母菌细胞的数目3、教师将把0.1ml酵母菌贮存用培养物注入试管A中,轻轻倒转试管几次使酵母细胞分布均匀。
试管B不加任何东西。
将试管盖稍微拧松并将两支试管放在教师指定的地方。
设置试管B的目的是什么?4、试管A中为刚开始增长的酵母菌新种群。
就下周期间你认为可能会发生的变化评论你的假设。
5、为了确定酵母菌种群的增长速率,必须在实验过程中对酵母菌进行计数。
(完整版)酵母双杂交原理

酵母双杂交系统原理酵母双杂交系统(Yeast Two-hybrid System)由Fields和Song等首先在研究真核基因转录调控中建立。
典型的真核生长转录因子,如GAL4、GCN4、等都含有二个不同的结构域: DNA 结合结构域(DNA-binding domain)和转录激活结构域(transcription-activating domain)。
前者可识别DNA上的特异序列,并使转录激活结构域定位于所调节的基因的上游,转录激活结构域可同转录复合体的其他成分作用,启动它所调节的基因的转录。
二个结构域不但可在其连接区适当部位打开,仍具有各自的功能。
而且不同两结构域可重建发挥转录激活作用。
酵母双杂交系统利用杂交基因通过激活报道基因的表达探测蛋白-蛋白的相互作用。
主要有二类载体: a 含DNA -binding domain的载体; b 含DNA-activating domain的载体。
上述二类载体在构建融合基因时,测试蛋白基因与结构域基因必须在阅读框内融合。
融合基因在报告株中表达,其表达产物只有定位于核内才能驱动报告基因的转录。
例如GAL4-bd具有核定位序列(nuclear-localization sequence),而GAL4-ad没有。
因此,在GAL4-ad氨基端或羧基端应克隆来自SV40的T-抗原的一段序列作为核定位的序列。
双杂交系统的另一个重要的元件是报道株。
报道株指经改造的、含报道基因(reporter gene)的重组质粒的宿主细胞。
最常用的是酵母细胞,酵母细胞作为报道株的酵母双杂交系统具有许多优点: 〈1〉易于转化、便于回收扩增质粒。
〈2〉具有可直接进行选择的标记基因和特征性报道基因。
〈3〉酵母的内源性蛋白不易同来源于哺乳动物的蛋白结合。
一般编码一个蛋白的基因融合到明确的转录调控因子的DNA-结合结构域(如GAL4-bd,LexA-bd);另一个基因融合到转录激活结构域(如GAL4-ad,VP16)。
酵母菌的形态结构PPT课件

▪荚膜:有些菌能够形成荚膜,致使菌落粘稠。 如胶红酵 母。
2021
14
三、酵母菌的细胞结构
1、细胞壁
▪壁外组成: •出芽痕和诞生痕: 酵母出芽繁殖时,子细胞与 母细胞分离,在子、母细胞 壁上都会留下痕迹。在母细 胞的细胞壁上点称出芽痕,子细胞细胞壁上的位点称诞 生痕。由于多重出芽,致使酵母细胞表面有多个小突起。 根据酵母细胞表面留下芽痕的数目,就可确定某细胞产 生过的芽体数,因而可估计该细胞的菌龄。
宽度:2.5—10μm 长度:4.5—21μm 酵母的大小、形态与菌龄、环境有关。一般成熟的 细胞大于幼龄的细胞,液体培养的细胞大于固体培养的 细胞。有些种的细胞大小、形态极不均匀,而有些种的 酵母则较为均匀。
2021
7
二、酵母菌的形态
(二)、酵母菌的菌落特征 1、固体培养菌落特征: • 菌落大而厚,圆形,光滑湿
➢能发酵糖类物质进行产能;
➢细胞壁常含有甘露聚糖;
➢适合在含糖量较高、偏酸性ຫໍສະໝຸດ 环境中生长,因而有人 称其为糖菌。
2021
2
一、酵母菌概述
2、酵母菌的应用
酵母菌种类较多,目前已知有56属500多种,同时, 酵母菌分布广,在果蔬表面、土壤、牛奶、动物的排泄 物以及空气中均有存在,与人类关系密切。 ➢在食品方面——酿酒、制作面包、生产调味品等。 ➢在医药方面——生产酵母片、核糖核酸、核黄素、氨 基酸等。 ➢在化工方面——使石油脱腊、以石油为原料生产柠檬 酸等。 ➢在农业方面——生产饲料(例如SCP)。 ➢在生物工程方面——作为基因工程的受体菌。 ➢在环保方面——废水处理。
2021
3
一、酵母菌概述
3、酵母菌的危害: ➢腐生性酵母菌能使食物、纺织品和其他原料腐败变质; ➢少数耐高渗的酵母菌和鲁氏酵母、蜂蜜酵母可使蜂蜜和果酱 等败坏; ➢有的酵母菌是发酵工业的污染菌,影响发酵的产量和质量; ➢某些酵母菌会引起人和植物的病害,例如白假丝酵母可引起 皮肤、粘膜、呼吸道、消化道等多种疾病.
(完整版)微生物学第三章酵母菌

假菌丝:藕节状的细胞串 真菌丝:竹节状的细胞串
任何细胞上的蒂痕仅一个, 而芽痕有一至数十个, 根据它的多少还可测定 该细胞年龄。
(single-cell protein,SCP)。 ➢在医药方面-生产酵母片、核黄素、乳糖酶、脂肪酶、
氨基酸、细胞色素c、B族维生素等。
维
假 丝
生 素
酵 母
B2 产 生
菌
3.重要的科研模式微生物
酿酒酵母(Saccharomyces cerevisiae),1996年,第一个 完成全基因组序列测定的真 核生物。
液泡:是单层膜包围的一种细胞器 线粒体:是两层单位膜包围的一种细胞器 微体:由单层膜包裹的小球形细胞器
线粒体电镜图
微 体 电 镜 图
三、酵母菌的繁殖方式和生活史
无性繁殖
酵母菌繁殖方式
有性繁殖
芽殖:各属酵母菌都存在
裂殖:Schizosaccharomyces(裂殖酵母菌)
酵 无性 母
节孢子:Geotricum(地霉属)等
等疾病。
二、酵母菌细胞的形态和构造
酵母菌细胞直径一般约是细菌 的10倍,是典型的真核微生物。
细胞大小:(2.5~10) m×(4.5~21) m
酵母菌细胞形态
圆形、椭圆形 球形、卵形、梨形 圆柱形、腊肠形
酵母菌的细胞形态
酵 母 菌 的 细 胞 形 态
酵母菌细胞构造的模式图
(一)细胞壁
假酵母(拟酵母):只有无性繁殖过程。 真酵母:无性繁殖+有性繁殖。
(一) 无性繁殖
1. 芽殖(budding)
最常见的一种繁殖方式,成熟 细胞长出一个小芽,母细胞的细胞 核分裂成两个子核,一个随母细胞 的细胞质进入芽体内,当芽体接近 母细胞大小时,自母细胞脱落成为 新个体,如此继续出芽。
微生物(酵母菌)

四、酵母菌的代表属
1、酵母菌属
例:啤酒酵母和葡萄汁酵母
2、裂殖酵母属
3、假丝酵母属
例:热带假丝酵母、解脂假丝酵母和产朊假 丝酵母 4、球拟酵母属 5、红酵母属
6、掷孢酵母属:属于担子菌亚门,
冬孢菌纲,黑粉菌目,掷孢酵母科。 孢子是由卵圆形的营养细胞生出的 小突起形成的,然后由一种机制有 力的射出,形成红至鲑肉粉红色的
宽度:2.5—10μm 长度:4.5—21μm 酵母的大小、形态与菌龄、环境有关。一般成 熟的细胞大于幼龄的细胞,液体培养的细胞大 于固体培养的细胞。有些种的细胞大小、形态 极不均匀,而有些种的酵母则较为均匀。
二、酵母菌的细胞结构
一般具有: 细胞壁、细胞膜、细胞核、液泡、线粒 体、内质网、微体、微丝、及内含物等, 有的菌体还有出芽痕、诞生痕。
三、繁殖方式
(一)繁殖方式: 分无性繁殖和有性繁殖两大类,主要是 无性繁殖。 无性繁殖:包括芽殖 、裂殖和产生无性 孢子 有性繁殖:主要是产生子囊孢子。 假酵母:只有无性繁殖过程。 真 酵 母 : 既 有 无 性 繁 殖 , 又 有 有 性繁 殖 过程。
无性繁殖: 1、芽殖:是酵母菌无性繁殖的主要 方式。 一个酵母能形成的芽数是有限的。(平 均24个) 出芽方式:多边出芽、两端出 芽、三边出芽、单边出芽。 环境适宜时,可出现假菌丝。
在农业方面——生产饲料(例如SCP)。
在生物工程方面——作为基因工程的受体菌。
酵母菌
酵母菌的危害: 1、腐生性酵母菌能使食物、纺织品和其他原 料腐败变质; 2、少数耐高渗的酵母菌如鲁氏酵母、蜂蜜酵 母可使蜂蜜和果酱等败坏;
3、有的酵母菌是发酵工业的污染菌,影响发 酵的产量和质量;
4、某些酵母菌会引起人和植物的病害,例如 白假丝酵母可引起皮肤、粘膜、呼吸道、消化道 等多种疾病.
(完整版)细菌、古菌和真菌(主霉菌、酵母菌)的比较

细菌、古菌和真菌(主霉菌、酵母菌)的比较2013生技基一班刘雨桐1.细胞构造比较细菌:包括细胞壁、细胞膜、细胞质和核区,特殊环境下形成的特殊构造有鞭毛、菌毛、性菌毛、糖被(包括荚膜和黏液层)和芽孢。
古菌:细胞壁,细胞膜,细胞质和核区。
真菌:细胞壁,细胞核,细胞质和细胞器(细胞基质和细胞骨架,内质网与核糖体,高尔基体,溶酶体,微体,线粒体,叶绿体,液泡,膜边体,几丁质酶体,氢化酶体)细菌古菌真菌细胞壁有有有细胞膜有有有细胞质和细胞器有细胞质无细胞器有细胞质无细胞器有细胞质和众多细胞器2. 细胞膜结构与组分的比较细菌:内外两暗色层间夹着一浅色中间层的一种双层膜结构,20%~30%磷脂、50%~70%蛋白质。
其中每个磷脂分子由一个带正电荷且能溶于水的极性头和一个不带电荷不溶于水的非极性端所构成,所有细菌细胞膜上都含磷脂酰甘油。
G-细菌中还富含磷脂酰乙醇胺。
磷脂双分子中嵌埋着整合蛋白或膜内在蛋白。
细胞膜是合成细胞壁和糖被有关成分的重要场所;膜上含有与氧化磷酸化或光合磷酸化等能量代谢有关的酶系,是细胞产能基地;是鞭毛基体着生部位。
原核细胞膜上一般不含胆固醇等甾醇。
古菌:细胞膜具有某些独特性和多样性。
①其磷脂的亲水头仍由甘油组成,但疏水尾却由长链烃组成,一般为异戊二烯重复单位。
②亲水头和疏水尾间由特殊的醚键连接,而其他原核或真核生物是酯键③古菌的细胞膜中存在独特的单分子层或单双分子层混合膜。
④在甘油分子C3位上,可连接多种与细菌和真核生物细胞膜上不同的基团。
⑤细胞膜上含有多种独特脂质。
真菌:与细菌的细胞膜构造和功能十分相似。
但其膜上有甾醇,磷脂种类为磷脂酰胆碱和磷脂酰乙醇胺。
在高等真菌中含有的是偶数碳原子的饱和或不饱和脂肪酸,在低等真菌中含有奇数碳原子的不饱和脂肪酸。
真菌细胞膜上有糖脂,具有细胞间识别受体的功能。
真菌上可进行胞吞作用。
甾磷脂种类脂肪酸种类糖电子基因转胞吞细胞核 无核膜包被完整细胞核 无核膜包被完整细胞核 有核膜包被完整细胞核醇脂传递链移运输作用细菌无磷脂酰甘油和磷脂酰乙醇胺等直链或分支,饱和或不含饱和脂肪酸;每一磷脂分子常含饱和与不和饱和酸各一无有有无真菌有磷脂酰胆碱和磷脂酰乙醇胺等高等真菌:含偶数原子的饱和或不饱和脂肪酸低等真菌:含奇数碳原子的不饱和脂肪酸有无无有古菌①其磷脂的亲水头仍由甘油组成,但疏水尾却由长链烃组成,一般为异戊二烯重复单位。
2019-2020学年高二生物人教版选修一课下能力提升:(十一) 酵母细胞的固定化 Word版含解析

课下能力提升(十一)酵母细胞的固定化【基础题组】1.下列属于固定化酶应用特点的是( )①可以被反复利用 ②有利于酶与产物分离 ③能自由出入载体 ④一种固定化酶只催化一种酶促反应 ⑤酶多用包埋法固定化A.①②③ B.③⑤C.①②④D.①②⑤解析:选C 酶被固定化后可与产品分离,故可反复使用,但不能自由出入依附的载体。
通常情况下固定化酶种类单一,所以只催化一种酶促反应。
2.下列关于固定化酶和固定化细胞的叙述,错误的是( )A.从操作角度来考虑,固定化细胞比固定化酶更容易B.固定化细胞比固定化酶对酶活性的影响更小C.固定化细胞固定的是一种酶D.将微生物的发酵过程变成连续的酶促反应应选择固定化细胞技术解析:选C 细胞内有许多种酶,因此固定化细胞固定的是一系列酶。
3.下列关于使用固定化酶生产高果糖浆的说法,正确的是( )①生产高果糖浆需要使用果糖异构酶 ②在反应柱的顶端装上分布着许多小孔的筛板,防止异物的进入 ③将葡萄糖溶液从反应柱的上端注入,果糖溶液从反应柱的下端流出 ④反应柱可以降低生产成本,提高果糖的产量和质量A.①② B.②③C.③④D.①④解析:选C 高果糖浆的生产需要使用葡萄糖异构酶,①错误;在反应柱内的底端装上分布着许多小孔的筛板,防止固定化酶漏出,②错误;将葡萄糖溶液从反应柱的上端注入,果糖溶液从反应柱的下端流出,③正确;反应柱能连续使用多次,降低了生产成本,提高了果糖的产量和质量,④正确。
4.相对固定化酶而言,固定化细胞对酶的活性影响较小,根本原因是( )A.避免了细胞破碎、酶的提取纯化过程B.固定化细胞在多种酶促反应中连续发挥作用C.催化反应结束后,能被重复利用D.细胞结构保证了各种酶在细胞内的化学反应中有效地发挥作用解析:选D 固定化细胞维持了细胞结构的稳定性,为酶充分发挥作用提供了稳定的环境。
5.有关制备固定化酵母细胞的叙述,正确的是( )A.首先用蒸馏水把干酵母、CaCl2、海藻酸钠配制成溶液,但配制CaCl2溶液需要加热B.配制海藻酸钠溶液时不需要加热,因为海藻酸钠在水中溶解速度很快C.配制酵母细胞海藻酸钠溶液,要用玻璃棒搅拌,使酵母细胞与海藻酸钠混合均匀D.酵母细胞溶液首先要和CaCl2溶液混合解析:选C 配制CaCl2溶液不需加热;配制海藻酸钠溶液需要加热,直至将海藻酸钠完全溶化;溶化好的海藻酸钠冷却至室温后加入活化的酵母细胞,进行充分搅拌,使其混合均匀。
高中生物选修一知识点(完整版)

【果酒和果醋的制作】一、果酒制作原理1.原理:微生物:___________,主要繁殖方式:_________代谢类型___________。
在有氧条件下,可以进行 呼吸,大量_______,反应式为:_____________________________________;在无氧条件下,进行 (或无氧呼吸),反应式为:________________________________________。
2.条件:酵母菌繁殖最适温度为 ,酒精发酵时一般控制在 ℃。
3.菌种来源: 二、果醋制作原理1.原理:微生物:___________,繁殖方式:_________代谢类型___________。
对氧气含量特别敏感。
中断通入氧气,会引起______________;当氧气、糖源充足时,醋酸菌将葡萄汁中的 分解成 ,其反应式为 ;当缺少糖源时,醋酸菌将 变为乙醛,再将乙醛变为 ,其反应简式为:________________________________________________。
2.条件:醋酸菌的最适生长温度为 ℃,此外还需要充足的 。
3.菌种来源:可从食醋中分离;到生产食醋的工厂或菌种保藏中心购买。
三、实验设计流程图四、关键操作步骤1.材料的选择与处理:选择新鲜的葡萄,榨汁前先将葡萄进行冲洗,但不能2.防止发酵液被污染:榨汁机要清洗干净,并晾干;发酵装置要清洗干净,并用70%的酒精 ;装入榨好的葡萄汁后, 。
3.控制好发酵条件:将葡萄汁装入发酵瓶,留大约 的空间。
制葡萄酒的过程中,将温度严格控制在 ℃,时间控制在 天左右;制葡萄醋的过程中,将温度严格控制在 ℃,时间控制在 天左右,并注意适时通过充气口 。
五、结果分析与评价可通过嗅闻、观察和品尝初步鉴定。
(可能的结果如下)酒精发酵 醋酸发酵 气味和味道酒味 酸味 气泡和泡沫有气泡和泡沫 无气泡和泡沫 发酵液状态 浑浊 浑浊,液面形成白色菌膜六、课题延伸:发酵后是否有酒精产生,可用 来检验。
(完整版)高中生物选修一试题(含答案)

人教版高中生物选修一练习题(含答案)第I卷(选择题)1.在制果酒、果醋、泡菜、腐乳时,所用微生物的代谢类型分别是A. 兼性厌氧型、需氧型、厌氧型、需氧型B. 厌氧型、厌氧型、厌氧型、需氧型C。
兼性厌氧型、需氧型、需氧型、厌氧型D。
厌氧型、兼性厌氧型、需氧型、厌氧型2.下列关于“DNA粗提取与鉴定"的实验,叙述正确的是A。
酵母、菜花和猪的红细胞均可作为提取DNA的材料B。
DNA易溶于氣化钠溶液,不溶于酒精溶液C。
向切碎的植物组织中加入适量蒸馏水,可获得含大量DNA的溶液D。
将提取的丝状物溶解后,加入二苯胺试剂后水浴呈蓝色3.下列有关生物技术实践应用的叙述,正确的是A. 固定化酶技术在多步连续催化反应方面优势明显B. 牛肉膏蛋白胨培养基是常用的细菌基础培养基,可用高压蒸汽灭菌法灭菌C. 相同pH时,加酶洗衣粉洗涤效果好于普通洗衣粉D。
向鸡血细胞中加入冷却的95%的酒精,可以获得无杂质的DNA4.下列关于果酒、果醋和腐乳制作的叙述中,正确的是A. 一般含糖量较高的水果可用来制作果酒B. 果醋发酵包括有氧发酵和无氧发酵C. 制作腐乳时,应防止毛霉以外的其它微生物生长D。
都是利用微生物胞内酶催化获得最终产品5.果胶酶能使果胶水解,使榨取果汁变得更容易,提高水果的出汁率。
某同学为探究温度对果胶酶活性的影响,在不同温度下,将等量的果胶酶加入到等量的苹果泥中,在反应同样时间后,再将反应液过滤同样时间,用量筒测出滤出苹果汁的体积.下列能正确反映实验结果的曲线是6.相对分子质量不同的两种蛋白质在凝胶色谱柱中的行进过程,可表示为图中哪一个()【注:图中大圆点代表相对分子质量大的蛋白质】7.下列有关酵母细胞固定化实验操作目的及结果分析,错误的是()A.干酵母细胞处于休眠状态,需置于蒸馏水中活化B.配制海藻酸钠溶液时需要小火或间断加热,以防海藻酸钠焦糊C.若海藻酸钠的浓度偏低时凝胶珠呈白色,固定的细胞数目过少D.凝胶珠在CaCl2溶液中浸泡时间的长短,不影响发酵效果8.如图所示的固定化方式中,适宜固定化酶的方法是 ( )A.①②③ B.①② C.②③ D.①③9.果酒和果醋制作过程中,发酵条件的控制至关重要,相关措施正确的是 ( )A.葡萄汁要装满发酵瓶,造成无氧环境,有利于发酵B.在葡萄酒发酵过程中,每隔12 h左右打开瓶盖一次,放出CO2C.果酒发酵过程温度控制在30℃,果醋发酵过程温度控制在20℃D.在果醋发酵过程中,要适时通过充气口充气,有利于醋酸菌的代谢10.下列不属于纤维素酶组分的是A.C1酶 B.Cx酶C.葡萄糖苷酶 D.半乳糖苷酶11.能合成脲酶的微生物所需要的碳源和氮源分别是A.C02和N2 B.葡萄糖和NH3C.C02和尿素 D.葡萄糖和尿素12.下列不属于菌落特征的是A.菌落的形状 B.有无荚膜C.菌落的大小 D.隆起程度13.变酸的酒表面有一层膜、泡菜坛表面有一层膜、腐乳外面有一层致密的皮,它们分别是A.醋酸菌、乳酸菌、毛霉菌丝B.醋酸菌、毛霉菌丝、毛霉菌丝C.醋酸菌、酵母菌、毛霉菌丝D.酵母菌、醋酸菌、乳酸菌14.从土壤中筛选纤维素分解菌的实验设计正确的是A.土壤取样→梯度稀释→稀释涂布平板→发酵培养B.土壤取样→选择培养→稀释涂布平板→挑选菌落C.土壤取样→梯度稀释→选择培养→挑选菌落D.土壤取样→梯度稀释→稀释涂布平板→选择培养→挑选菌落15.下列有关分解纤维素微生物的分离的叙述,错误的是A.纤维素酶的发酵方法有液体发酵和固体发酵B.对分解纤维素的微生物进行了初步筛选后,无需再进行其他实验C.纤维素酶的测定方法,一般是对所产生的葡萄糖进行定量测定D.纤维素酶是一种复合酶16.含氨苄青霉素的LB培养基在基因工程中有很重要的应用。
酵母细胞的固定化(公开课)

酵母细胞的固定化(公开课)酵母细胞的固定化(公开课)介绍酵母是一种常见的微生物,被广泛应用于食品工业、酿酒工业和生物技术等领域。
酵母细胞的固定化是指将酵母细胞固定在一定的载体上,以便于对其进行分离和重复利用。
固定化技术可以有效提高酵母细胞的稳定性和生产效率,具有重要的实际应用价值。
1. 酵母细胞固定化的原理酵母细胞固定化的原理是利用一定的载体材料将细胞固定在固定床内,形成“细胞载体复合物”。
载体材料通常选择具有良好的稳定性和生物相容性的材料,如多孔陶瓷、海藻酸钙和高分子材料等。
载体表面的孔隙结构提供了良好的生长环境和微观结构,有利于细胞的附着和生长。
2. 酵母细胞固定化的优势相比于游离状态下的酵母细胞,固定化酵母细胞具有以下优势:- 稳定性提高:固定化酵母细胞不易受到环境条件的影响,能够在不同的温度、pH值和抑制物浓度下保持较高的活性。
- 操作简便:固定化酵母细胞可以灵活地进行操作和控制,方便分离和回收,减少了生产过程中的工艺复杂性。
- 循环利用:固定化酵母细胞能够重复利用,提高了酵母细胞的利用率和产能,减少了资源的浪费。
3. 酵母细胞固定化的应用酵母细胞固定化技术在食品工业、酿酒工业和生物技术等领域有着广泛的应用。
3.1 食品工业酵母细胞固定化在食品工业中常被用于发酵产品的生产,如面包、酸奶和啤酒等。
固定化酵母细胞能够提高发酵效率和产品品质,同时还能够减少生产过程中的能耗和废水排放。
3.2 酿酒工业在酿酒工业中,固定化酵母细胞能够提高酿酒过程中的稳定性和酒品质量。
固定化酵母细胞还可以在高温和高浓度的条件下继续进行发酵,从而减少了生产时间和成本。
3.3 生物技术在生物技术领域,固定化酵母细胞常被用于生产生物活性物质,如药物、酶和蛋白质等。
固定化酵母细胞能够提高生产效率和纯度,并且易于回收和再利用,有利于大规模生产。
4. 酵母细胞固定化的方法酵母细胞固定化的方法多种多样,常见的方法包括胶包固定化、吸附固定化、凝胶固定化和共价固定化等。
(完整版)实验:探究酵母菌细胞呼吸的方式
选一选
适宜的温度条件下,在下列装置中都加入活酵母,其
中适于产生酒精的装置是( A)
A 加入葡萄糖和水
B 加入葡萄糖
C 加入水
D
加入葡萄糖和水 并不断搅拌
+ A装置的酵母菌滤液
不变灰绿色
+ B装置的酵母菌滤液
溶有重铬酸 钾的浓硫酸
灰绿色
五、观测、记录结果
ห้องสมุดไป่ตู้
条件 澄清石灰水/出现时间 重铬酸钾-浓硫酸溶液
有氧 无氧
变混浊/快 变混浊/慢
无变化 出现灰绿色
六、得出结论
1、酵母菌在有氧和无氧情况下均产生了CO2, 能使澄清石灰水变浑浊。 2、酵母菌的有氧呼吸比无氧呼吸释放的CO2要 多。 3、酵母菌在有氧情况下,没有酒精生成,不能 使重铬酸钾溶液发生显色反应;在无氧情况下, 生成了酒精,使重铬酸钾溶液发生灰绿色显色 反应。
有氧呼吸装置
无氧呼吸装置
四、实验步骤
1、配制酵母菌培养液(等量原则)置于A、B锥形瓶 2、组装有氧呼吸和无氧呼吸装置图,放置在25-35℃、 环境下培养8-10小时。 3、检测CO2的产生 4、检测酒精的产生 (1)取2支试管编号; (2)各取A、B锥形瓶酵母菌培养液的滤液2毫升注入 试管; (3)分别滴加0.5ml重酪酸钾-浓硫酸溶液,轻轻震荡、 混匀。
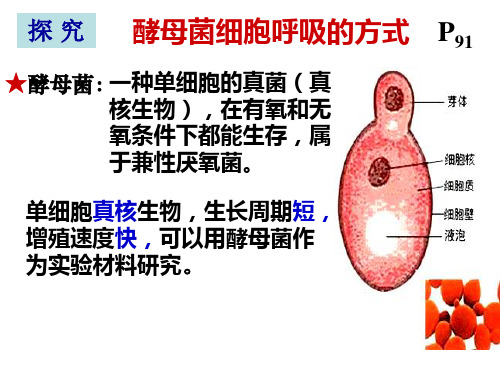
探 究 酵母菌细胞呼吸的方式 P91
★酵母菌:一种单细胞的真菌(真 核生物),在有氧和无 氧条件下都能生存,属 于兼性厌氧菌。
单细胞真核生物,生长周期短, 增殖速度快,可以用酵母菌作 为实验材料研究。
一、实验原理 1、酵母菌是单细胞真菌属于兼性厌氧菌。 进行有氧呼吸产生水和CO2 ,无氧呼吸产 生C2H5OH和CO2。 2、 CO2的检测方法 (1)CO2使澄清石灰水变浑浊 ★(2)CO2使溴麝香草酚蓝水溶液由蓝变 绿再变黄 3、酒精的检测 ★橙色的重酪酸钾溶液在酸性下与酒精发生 反应,变成灰绿色。
酵母演示文稿1

(二)有性生殖 酵母菌进行的有性繁殖主要是产生子囊和子囊孢子。 其基本过程包括:质配、核配和减数分裂三个基本过 质配: 当一些酵母菌发育至一定阶段或在特定的培养条件 两个性别不同的细胞各伸出小突起相互接触, 形成一个通道;两细胞质融合,称为质配。 核配: 两单倍体的核移到融合管中融合成二倍体的核。此 倍体细胞称为合子。 减数分裂: 在合适条件下,二倍体细胞的核进行一次减数分裂 1~2次有丝分裂,形成4~8个有性的子囊孢子 成子囊孢子的细胞,称为子囊。
与人类关系密切
酵母菌概述 酵母的应用
酵母菌是人类应用比较早的微生物。
在食品方面——酿酒、制作面包、生产调味品等。
在医药方面——生产酵母片、核糖核酸、核黄素、 细胞色素C、B族维生素、乳糖酶、脂肪酶、氨基酸 等。 在化工方面——使石油脱腊、以石油为原料生产 柠檬酸等。
在农业方面——生产饲料(例如SCP)。 在生物工程方面——作为基因工程的受体菌。
酵母菌的危害:
腐生性酵母菌能使食物、纺织品和其他原料 腐败变质;
少数耐高渗的酵母菌和鲁氏酵母、蜂蜜酵母 可使蜂蜜和果酱等败坏;
有的酵母菌是发酵工业的污染菌,影响发酵 的产量和质量;
某些酵母菌会引起人和植物的病害,例如白 假丝酵母可引起皮肤、粘膜、呼吸道、消化道 等多种疾病.
二、酵母菌的形态特征与大小
(二)酵母菌的生活史类型
1. 单倍体型
2. 双倍体型
3. 单双倍体型
1. 单倍体型
以八孢裂殖酵母为代表 特点:营养细胞是单倍体;无 性繁殖以裂殖方式进行; 双倍体细胞不能独立生活, 故双倍体阶段短,一经生 成立即减数分裂。
2、双倍体型
以路德类酵母为代表 特点:营养体为双倍体, 不断进行芽殖,双倍体 营养阶段长,单倍体的 子囊孢子在子囊内发生 接合。单倍体阶段仅以 子囊孢子形式存在,故 不能独立生活。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
酵母细胞表面展示技术是一种真核蛋白表达系统。
其基本原理是将外源靶蛋白基因与特定的载体基因序列融合后导入酵母细胞,融合蛋白诱导表达后,信号肽引导融合蛋白向细胞外分泌,外源蛋白的展示主要是借助于GPI锚的作用,GPI锚定蛋白的C端包含疏水多肽,当细胞蛋白被合成,前体蛋白通过羧基末端的疏水序列锚定在内质网膜上,其余蛋白则位于内质网腔中。
在极短的时间内,疏水序列在转酰氨基酶的作用下ω位点裂开同时被GPI锚置换,并被运输到高尔基体再通过分泌途径分泌至细胞膜外。
在蛋白水解酶作用下,分泌信号序列被切除,细胞蛋白被磷脂酰肌醇特异性磷脂酶C从细胞膜上释放并以GPI锚定形式与葡聚糖共价相连被转运至细胞壁外。
图1展示的是通过α-凝集素系统转运目的蛋白到细胞表面的分泌途径。
酵母表面展示技术具有表达偏差小,展示的融合蛋白相对分子质量范围大,分子数量多,筛选、纯化、活性测定方便等优点。
目前,最常见的酵母表面展示系统包括:凝集素展示表达和絮凝素展示表达[5]。
如图2所示,α-凝集素展示表达系统(图2A)是将外源基因与酵母编码α-凝集素C端编码序列连接后插入质粒载体的分泌信号肽(Secretion signal,SS)下游,融合蛋白诱导表达后,信号肽引导嵌合蛋白向细胞外分泌,由于融合蛋白C-末端存在含GPI锚定信号(GPIanchor attachment signal)的凝集素多肽序列,可将蛋白锚定在酵母细胞壁上,从而将蛋白分子展示表达在酵母细胞表面;a-凝集素展示系统(图2B)与α-凝集素展示系统不同的是其具有由二硫键连接的Aga1和Aga2两个亚基,目的蛋白通过与Aga2形成融合蛋白,再通过Aga1末端的GPI结构使目的蛋白表达于细胞表面;絮凝素展示表达系统利用絮凝素基因(flo1p)与外源基因融合,并将融合蛋白展示表达在细胞表面,此系统又包括两个子系统:GPI锚定系统(图2C)和絮凝结构域(Flo1p flocculation functional domain)锚定系统(图2D)。
GPI锚定系统是利用絮凝素Flo1p的C末端含有的GPI结构锚定外源蛋白,此系统与α-凝集素展示相似;絮凝结构域锚定系统是利用Flo1p的中间絮凝功能结构域与外源蛋白融合,通过絮凝功能结构域识别酵母细胞壁中的甘露聚糖链并通过非共价作用诱导细胞粘附、聚集成可逆性絮状物。
进而使外源蛋白表达于酵母细胞表面。
近年来,S.cerevisiae细胞表面展示技术迅速发展并在多个领域得到应用,展现出广阔的发展前景。
可通过使外源蛋白固定化于S.cerevisiae细胞表面,从而生产微生物细胞表面蛋白,包括淀粉酶、纤维素酶等在生物乙醇生产中起关键作用的酶蛋白,从而使酵母表面展示技术在生物乙醇生产应用中不断发展与完善,本文着重介绍了近年来酵母细胞表面工程在生物乙醇菌种开发中的应用与研究进展。
1酵母细胞表面技术展示淀粉分解酶以淀粉类生物质为主要成分的蔗糖和糖蜜可以实现大规模的乙醇生产,然而,以淀粉为原料生产生物乙醇要经历多步反应而且生产成本偏高。
导致成本偏高的原因主要有两个:一是由于S.cerevisiae不能直接利用淀粉类物质,因此在发酵过程中需要额外添加大量的葡糖淀粉酶(Glucoamylase)和α-淀粉酶(α-Amylase);二是为获得较高的乙醇产量,首先要将淀粉在140℃~180℃的高温下进行糊化,而这也就增加了能耗。
为此,大量的研究集中尝试在S.cerevisiae细胞表面展示淀粉分解酶,从而实现以淀粉类物质为底物的大规模乙醇生产,并取得了一定的成果。
利用细胞表面展示技术构建能利用淀粉的全细胞催化剂,可将编码淀粉酶的外源基因导入酵母细胞内使其能够分泌淀粉酶[7]。
已经构建好的质粒pGA11带有编码米根霉Rhizopus oryzae的葡糖淀粉酶基因,转化子细胞能够在以1%可溶性淀粉为唯一碳源的培养基中迅速生长并产生乙醇,葡糖淀粉酶活力稳定至少100h,而且所表达的酶,其酶学性质与天然酶相当[8]。
Murai等[9]是研究细胞表面表达淀粉酶工作的先驱,他们创造性地通过在S.cerevisiae 细胞表面展示淀粉酶来开发重组酿酒酵母的研究策略,构建了带有S.cerevisiae MT8-1的GAPDH启动子及含有信号肽和glucoamylase/α-凝集素融合基因的载体,glucoamylase来自于R.oryzae,能有效切割淀粉α-1,4和α-1,6糖苷键。
随后,Kondo[10]等在具絮凝功能特性的酵母细胞表面展示了葡糖淀粉酶,他们的工作表明,展示在细胞表面的葡糖淀粉酶对酵母的正常生长及乙醇产生无任何影响;厌氧发酵150h后乙醇浓度达到20~30g/L。
但Kondo 等构建的展示葡糖淀粉酶的重组絮凝酵母在发酵300h的情况下还能保持较高的乙醇产率(0.6~0.7g/(L·h))[10],同时,发酵培养基中低浓度的葡萄糖对减少污染也极为有利。
展现了在S.cerevisiae细胞表面展示淀粉酶的优越性。
在批式补料发酵过程中,由于缺少溶解酶——α-淀粉酶,单一展示葡糖淀粉酶的酵母菌株在发酵过程中会有不溶性淀粉的积累。
为克服这一问题,Shigechi等[11-14]构建了共展示来自R.oryzae的glucoamylase和来自嗜热脂肪芽胞杆菌Bacillus stearothermophilus的α-amylase基因,并以可溶性淀粉为底物,进行补料批式发酵。
结果表明,将淀粉颗粒处理到与酵母细胞大小相近时可极大程度地提高乙醇产率,发酵100h后乙醇浓度达到60g/L,而且共展示葡糖淀粉酶和α-淀粉酶菌株的絮凝能力与只展示葡糖淀粉酶的菌株基本相同,发酵过程中酵母絮凝能力并不发生改变。
在后续的研究中,Shigechi等[14]构建了含有葡糖淀粉酶结合结构域(SBD)突变子的酵母展示株系,结果发现,带有突变子SBD葡萄淀粉酶的菌株较野生型菌株的淀粉降解能力提高1.4倍。
Shigechi等[13]又利用低温糊化的淀粉(80)℃取代可溶性淀粉作为唯一碳源进行乙醇发酵,结果表明:共展示葡糖淀粉酶和α-淀粉酶的菌株与单展示葡糖淀粉酶的菌株相比,乙醇产率明显增加,尤其是共展示菌能更快速地产生乙醇且无延滞期,说明随机水解淀粉α-1,4糖苷键的α-淀粉酶在淀粉水解过程中起关键作用。
比较共展示两种淀粉酶的菌株在低温和高温糊化情况下的发酵效率,无论是最大乙醇浓度、乙醇产率还是底物消耗率,两种发酵系统都基本相同,乙醇产量为0.5g/g,相当于理论值的97.2%。
更好地说明了共展示两种淀粉酶菌株的优势。
从牛链球菌Streptococcus bovis中分离的淀粉酶为生料淀粉的高效水解提供了新的研究思路[15]。
Shigechi等[12]分别以α-凝集素和絮凝素为基础,共展示来自R.oryzae的葡糖淀粉酶和来自S.bovis148的α-淀粉酶,并以生料淀粉为底物进行发酵试验,结果表明α-淀粉酶活性依赖于锚定蛋白,而葡糖淀粉酶活性则独立于锚定蛋白;对于α-淀粉酶活性,以絮凝素系统为基础构建的菌株要比以凝集素为基础构建的菌株酶活性高40倍。
目前已经报道了一些α-淀粉酶具有结合生料淀粉的能力,且降解淀粉的区域位于C-末端[16]。
以α-凝集素为基础的共展示菌能以生料淀粉为原料生产乙醇[17-19]。
另一方面,基于α-凝集素和絮凝素Flo1p共展示的葡糖淀粉酶和α-淀粉酶在生料发酵过程中不需要额外添加淀粉酶,在发酵过程中,淀粉浓度迅速下降,72h乙醇浓度达到61.8g/L,乙醇产量为0.44g/L,相当于理论值的86.5%。
利用酵母细胞表面展示技术从最初的单展示葡糖淀粉酶到共展示葡糖淀粉酶和α-淀粉酶,均取得了较好的成果,为利用淀粉类物质实现大规模乙醇生产提供了新的思路。
不过随着生物乙醇生产技术的不断发展,人们开始着眼于利用木质纤维素生产燃料乙醇,从而避免了第一代生物乙醇存在的与人争粮的缺陷。
2酵母细胞表面技术与第二代生物乙醇由于第一代生物乙醇的生产存在着与人争粮、与粮争地的缺陷,发展第二代生物燃料是人们面对能源、环境与粮食问题所采取的积极措施。
利用丰富廉价的可再生木质纤维素生产乙醇,是规模化发展车用燃料乙醇的基础。
木质纤维素是地球上最丰富的可再生资源,占地球总生物量的40%左右,高效、廉价地利用木质纤维素转化为如燃料乙醇等化工产品,是由石油基向生物基经济社会过渡的基础。
为此大量的研究开始转向展示纤维素分解酶、木糖代谢酶等,从而实现新的突破与尝试,为实现第二代生物燃料的开发与利用提供了一定的理论依据。
2.1展示纤维素分解酶在纤维素乙醇生产经历的糖化步骤形成葡萄糖的过程中,S.cerevisiae不能直接利用纤维素类物质,因此在S.cerevisiae中稳定表达纤维素酶是降低生产成本的有效方法。
基于α-凝集素C端构建的pCMC11含有来自棘孢曲霉Aspergillusaculeatus的内切羧甲基纤维素酶CMCase)编码基因,Murai等[20]通过一系列方法证明了cmcase基因成功地锚定在细胞表面,内切羧甲基纤维素酶活性稳定至少120h。
当pCMC11和含有β-bgl1的载体(pBG211)共表达时,可以快速地将纤维低聚糖转化为葡萄糖,从而显著地提高了利用纤维素的效率。
目前已报道了很多在S.cerevisiae中表达多种纤维素酶基因和能够糖化可溶性纤维寡糖的重组酿酒酵母[21-22]。
Fujita等同样基于α-凝集素系统在S.cerevisiae细胞表面展示来自丝状真菌里氏木霉Trichoderma reesei的内切葡聚糖酶Ⅱ的融合蛋白,酵母菌株能明显提高对大麦β-葡聚糖的水解能力;随后,构建了共表达内切葡聚糖酶Ⅱ和来自A.aculeatus的β-葡糖苷酶Ⅰ的酵母株系,这种酵母能在以大麦β-葡聚糖为唯一碳源的培养基上生长,在50h内将β-葡聚糖发酵生成乙醇,乙醇产量为0.48g/g,相当于理论值的93.3%,且发酵培养基中的β-葡聚糖能在酵母细胞表面连续水解为葡萄糖,在胞内酶的作用下,立即被利用,转变为乙醇。
同时,酵母细胞的酶活测定结果和流式细胞计数分析结果表明:共展示菌中内切葡聚糖酶Ⅱ和β-葡糖苷酶Ⅰ分子的数量大于单展示菌中内切葡聚糖酶Ⅱ或β-葡糖苷酶Ⅰ分子数的总和[23]。
其后,Fujita研究小组[24]将来自T.reesei的纤维二糖水解酶Ⅱ也增加到展示系统中,从而使酵母细胞真正具有水解无定形纤维素能力,同步糖化发酵生成乙醇,结果显示共展示内切葡聚糖酶Ⅱ和纤维二糖水解酶Ⅱ的菌株比只展示内切葡聚糖酶Ⅱ的菌株对无定形纤维素的水解能力高,且主要产物是纤维二糖,单展示纤维二糖水解酶Ⅱ的菌株主要产物是少量的纤维二糖,单展示内切葡聚糖酶Ⅱ的菌株主要产物是纤维二糖、纤维三糖,而共展示β-葡糖苷酶Ⅰ和内切葡聚糖酶Ⅱ的菌株无乙醇产生。