神经元活动的一般规律和神经元的作用方式
动物生理学--神经系统
都有相应的受体
脑啡肽的镇痛功能
二、 中枢神经元的联系和活动
1 中枢神经元的联系方式
辐散式:兴奋或抑制的扩散 聚合式:总和或整合 连锁状:空间上加强了作用范围 环状:后放或及时终止
正、负反馈的基础 P265
2 中枢兴奋与中枢抑制 —中枢活动的两种基本过程
↓
肌紧张和肌运动↑
特 点 正常情况下活动较弱
正常情况下活动较强,
在肌紧张的平衡调节中占优势
4.3 小脑对躯体运动的调节
前庭小脑(原始小脑) 维持躯体姿势平衡 脊髓小脑(旧小脑) 调节肌紧张 皮层小脑(新小脑) 协调随意运动
4.4 大脑皮层对躯体运动的调节
❖ 发动和协调肌肉运动
起源 – 大脑皮层联络区;
肾上腺素能受体: 以兴奋为主: 1受体:1A/1B/1D 2受体:2A/2B/2C 以抑制为主: 1、2、3
3.2 中枢递质与受体
乙酰胆碱:感觉、运动、学习、记忆 单胺类:(去甲)肾上腺素、多巴胺、5-羟色胺。
情绪、觉醒、睡眠 氨基酸类:谷、天冬、甘、GABA 肽类:阿片样肽(脑啡肽,强啡肽 )与痛觉和镇痛有关;
b.去甲肾上腺素( NA or NE ) 肾上腺素能神经纤维
除支配汗腺和骨骼肌的舒血 管以外的交感神经节后纤维
c.嘌呤类或肽类 嘌呤能或肽能神经纤维
胃肠道壁内神经丛中的一些纤维释放ATP、血管活性肠肽、 促胃液素、生长抑素等。
受体
胆碱能受体: 以兴奋为主 毒蕈碱M型: M1/M2/M3/M4/M5 烟碱N型:N1/N2
突触前抑制 突触后抑制
传入侧支性抑制(交互抑制)
回返性抑制
图
突触前抑制:兴奋性递质释放的减少
大脑皮层的神经元活动规律及功能解析

大脑皮层的神经元活动规律及功能解析大脑是人类最神秘、最复杂的器官之一,拥有数十亿个神经元,这些神经元连接在一起,组成了大脑皮层,控制着人类的思考、记忆、行为和情绪等。
为了更好地了解大脑皮层的神经元活动规律以及功能解析,本文将对其进行探讨。
一、神经元的基本结构和功能神经元是构成神经系统的基本细胞,它具有三个主要部分:细胞体、轴突和树突。
其中细胞体是神经元的核心,包括细胞核和许多细胞器。
轴突则是神经元传递信息的主要通道,而树突则负责接收传入信息并将其发送给细胞体。
神经元的功能主要包括接收、处理和发射信息,实现神经系统的各种功能。
二、大脑皮层神经元的分类和分布大脑皮层是人类大脑外部的一层薄膜,包括六个层次,每个层次都有特定类型的神经元。
根据神经元的形态和功能,大脑皮层神经元可以分为三类:锥形细胞、星形细胞和小细胞。
锥形细胞是大脑皮层中最常见的神经元,主要位于第三、四和五层,其轴突可以连接到其他大脑皮层区域,也可以连接到深部结构。
星形细胞则位于第一层和第二层,主要承担支持和维护神经元生存的功能。
小细胞与锥形细胞类似,但其体积较小,主要分布在第六层。
大脑皮层神经元分布的不均衡性也是其研究的重点之一。
从神经元的数量上来看,前额叶和顶叶的神经元密度最高,而枕叶和颞叶的神经元密度相对较低。
从神经元的连接上来看,同一区域内的神经元之间会产生许多连接,而不同区域之间则会建立跨区域的连接,实现信息传递和整合。
三、神经元活动的规律与脑电图特征神经元的活动和信息传递通常通过细胞膜电位来进行,正常时神经元的细胞膜电位静息在低水平,当接收到外部刺激时,细胞膜电位会出现“快速上升-快速下降-慢速下降”的过程,称为动作电位。
动作电位的产生和传导需要大量的离子通道参与,各种离子的参与会表现出不同的电生理特征。
神经元的活动可以通过记录脑电图来进行研究。
脑电图是大脑皮层神经元活动的电信号表现形式,可以通过电极采集和放大,呈现出不同频率和振幅的波形。
神经调节

– 特征:突触前膜释放?递质 突触后膜? – ?离子内流 – ?突触后电位
• 抑制性突触传递过程
– 特征:突触前膜释放?递质 突触后膜? – ?离子内流 – ?突触后电位
(2)非定向突触传递
• 概念:不通过经典突触进行的化学传递 • 特点 – 曲张体与效应器不形成经典的突触联系
(3)紧张性作用在不同状态下不同
• 剧烈活动时:交感神经活动占优势
• 安静状态下:副交感神经活动就占优势
(4)整体生理功能调节不同
• 交感神经的作用是使人体迅速适应环境的急剧变化 :交感-肾上腺髓质系统 • 副交感神经的作用是促进消化吸收、积蓄能量及加 强排泄和生殖功能:迷走-胰岛素系统
(三)内脏活动的中枢调节
• 突触(synapse) 神经元与神经元之间相接 触并传递信息的特殊结构 • 结构:突触前膜
突触间隙
突触后膜
分类
• 按结构分
轴-树突触 轴-体突触 轴-轴突触 • 按功能分 兴奋性突触 抑制性突触
(2)突触传递过程:电——化学——电(动画)
动作电位——神经递质——突触后电位
突触传递过程小结
• 分布
– N1分布于自主神经节神经元的突触后膜
– N2分布于神经-肌接头的骨骼肌终板膜上 • N样作用:乙酰胆碱兴奋自主神经节后神经元引起 骨骼肌收缩
有机磷农药中毒
有机磷农药抑制胆碱酯酶活性
– 胆碱酯酶作用:降解乙酰胆碱
• 乙酰胆碱在胆碱能神经突触间隙蓄积 • 产生M样作用,N样作用以及中枢神经症状 • 可因呼吸中枢神经麻痹致死
突起
• 胞体:位于脑、脊髓、神经节中 是神经元代谢和营养的中心 2. 神经纤维:轴突外包髓鞘或神经膜
动物生理学第九章神经生理

神经冲动传导的原理 无髓纤维的传导 局部电流 有髓纤维的传导 郎飞氏结处形成局部电流
(二)神经纤维 的分类 (三)神经纤维 的轴浆运输
添加标题
突触
添加标题
轴-树突触
添加标题
突触的分类和基本 结构
添加标题
轴-胞突触
添加标题
突触的分类
添加标题
轴-轴突触
添加标题
根据突触的接触部 位不同
二.根据突触对下一个神经元的影响不同 一类是兴奋性突触,另一类为抑制性突触 二.根据突触工作的方式不同,分为电突触和化
具有精细定位
添加标题
04
支配不同部位有不同的 定位区
添加标题
椎体系统
05
添加标题
06
椎体外系统
添加标题
07
第五节 神经系统 对内脏活动的调节
添加标题
பைடு நூலகம்
08
交感神经和副交感 神经特征
二.交感神经和副交感神经的功能 三.植物性神经末梢的兴奋传递 四.神经递质 胆碱能纤维: 肾上腺素能纤维: 胆碱能纤维又分为烟碱型作用(N型),
添加标题
去大脑僵直
添加标题
基底神经节对躯体 运动的调节
添加标题
脑干对姿势反射的 调节
添加标题
小脑对躯体运动的 调节
添加标题
状态反射
添加标题
小脑的结构
添加标题
翻正反射
小脑的功能
原始小脑
旧小脑
新小脑
大脑皮质对躯 体运动的调节
添加标题
01
添加标题
02
添加标题
03
大脑皮层运动区
特点:1.两侧交叉
单击添加副标题
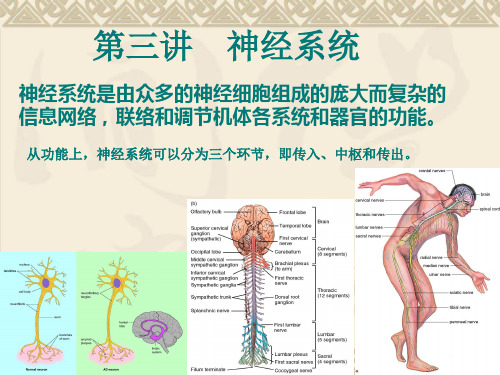
第 九章 神经生理
神经系统

时,大趾背屈,其它四肢展开如扇形。
(2) 节间反射(搔爬反射)
二、脑干对肌紧张和姿势的调节 脑干结构:腹面、背面
(一)脑干部位下行系统 1、下行易化系统:网状—脊髓通路(5);前庭—脊髓通路(6) 易化区: 分布于脑干中央区域,延髓网状结构的背外侧, 脑 桥的被盖,中脑的中央灰质和被盖,前庭,小脑前叶两侧。 2、下行抑制系统:皮质—延髓—网状通路(1);尾核—脊髓 通路(2);小脑—网状通路(3);网状—脊髓通路(4) 抑制区: 位于延髓网状结
第二章 中枢神经系统的功能
本章概述:
一、神经系统如何接受感觉信息(感觉机能); 二、神经系统如何支配躯体骨骼肌的运动及内 脏肌肉的运动(运动机能); 三、神经系统有哪些高级机能及其特征(高级 机能)。
第一节 神经元活动的一般规律
一、 神经元和神经纤维
NC是神经系统中最基本的 结构和功能单位,故称为神经 元(Neuron)。 按照生理机能,一般可将 神经元分为三类 (1)感觉(传入神经元) (2)运动(传出神经元) (3)中间(联合神经元)
三、 反射的分类
按反射形成的特点 将所有的反射区分为非条件反射
和条件反射两大类。
非条件反射是动物生来就有的。 条件反射不是先天就具有的,是动物个体后天通过学 习和训练所获得的。
四、 中枢神经元的联系方式
1、辐散 2、聚合
3、链锁状与环状联系
五、 反射弧中枢部分兴奋传布的特征 1、单向传布 在中枢内兴奋传布只能由传入向传出 的方向进行,而不能逆向传布。 2、中枢延搁 从刺激感受器起至效应器开始出现反 射活动为止,所需的全部时间称为反射时。兴奋通过 中枢部分较慢。 3、总和 如果由同一传入纤维先后连续传入多个冲 动,或许多条传入纤维同时传入冲动至同一神经中枢 ,则阈下兴奋可以总和。当达到一定水平就能发放冲 动,引起反射活动。
306西医综合考研真题笔记生理复习神经系统一

306西医综合考研真题笔记生理复习神经系统(一)考纲要求1.神经元活动的一般规律:神经纤维传导的特征,速度,神经纤维的分类以及神经的营养性作用,神经胶质细胞的功能。
2.突触与突触传递:兴奋性突触与抑制性突触传递的过程和原理,突触前抑制。
神经递质。
突触传递的特点。
3.反射中枢的概念,中枢兴奋和抑制的过程。
4.神经系统的感觉机能:感觉的特异与非特异投射系统及其在感觉形成中的作用。
痛觉。
5.神经系统对躯体运动的调节:骨骼肌的运动单位,牵张反射,肌紧张及其调节。
锥体系统及锥体外系统在运动调节中的作用,中枢神经调节系统其他部位对运动的调节作用。
6.神经系统对内脏机能的调节:植物性神经系统及其化学传递,低位脑干对内脏机能的调节,下丘脑对内脏活动的调节。
7.脑的高级机能:条件反射的形成和生物学意义,人类条件反射的特征。
大脑皮层的语言中枢及两侧大脑半球的职能分工。
8.两种睡眠状态及其特点。
考纲精要一、神经元和神经纤维1.神经元即神经细胞,是神经系统的基本结构和功能单位。
神经元由胞体和突起两部分组成,胞体是神经元代谢和营养的中心,能进行蛋白质的合成;突起分为树突和轴突,树突较短,一个神经元常有多个树突,轴突较长,一个神经元只有一条。
胞体和突起主要有接受刺激和传递信息的作用。
2.神经纤维即神经元的轴突,主要生理功能是传导兴奋。
神经元传导的兴奋又称神经冲动,是神经纤维上传导的动作电位。
神经元轴突始段的兴奋性较高,往往是形成动作电位的部位。
3.神经胶质:主要由胸质细胞构成,在神经组织中起支持、保护和营养作用。
二、神经冲动在神经纤维上传导的特征1.生理完整性:包括结构和功能的完整,如果神经纤维被切断或被麻醉药作用,则神经冲动不能传导。
2.绝缘性:一条神经干内有许多神经纤维,每条神经纤维上传导的神经冲动互不干扰,表现为传导的绝缘性。
3.双向传导:神经纤维上任何一点产生的动作电位可同时向两端传导,表现为传导的双向性,但在整体情况下是单向传导的。
解剖生理学第九章神经系统

(2)与躯体运动有关:
其对骨骼肌的调节作用,主要是 其下行纤维(网状脊髓束),终
于脊髓前角运动细胞(、r细 胞)。
(3)参与调节内脏活动:
脑干网状结构中有呼吸中枢、血 管运动中枢、血压调节中枢和呕 吐中枢等(生命中枢)。
髓节段。
脊髓节段与椎骨的对应关系
脊髓节段 第1—第4颈节 第5颈节—第4胸节 第5—第8胸节 第9—第12胸节 第1—第5腰节 全部骶节和尾节
椎骨的椎体 第1—第4颈椎 (一对一) 第4颈椎—第3胸椎(高一) 第3—第6胸椎 (高二) 第6—第9胸椎 (高三) 第10—第12胸椎 第12胸椎和第1腰椎
神经核—功能相同的神经元胞体集中形
成的灰质团块
白质—神经纤维集中处色泽白亮
纤维束—起止和功能基本相同的神经纤
维集合成束
神经系统
网状结构——灰质、白质混合形成
的结构
周围神经系统:
神经节—神经元胞体集中处形成的
结节状结构
神经—神经纤维聚集成束,并被结
缔组织包裹形成圆索状的
结构
第一节 神经元活动的一般规律
第四脑室向上经中脑水管通第三脑室,向下通脊髓中央管,并借 正中孔和外侧孔与蛛网膜下腔相通。
脑干内构特点
1.由灰质、白质和网状结构构成。 2.中央管开放形成第四脑室底(菱形窝), 使灰质核团由腹背方向排列变成内外方向排 列。感觉柱位于界沟的外侧;运动柱位于界 沟的内侧;与内脏相关的靠近界沟;与躯体 相关的则远离界沟。 3.神经纤维左右交叉(锥体交叉、内侧丘系 交叉、三叉丘系交叉、斜方体、小脑上脚交 叉)使灰质柱断裂成细胞团块。即包括脑神 经核、非脑神经核、网状核、中缝核。
神经元活动的一般规律和神经元的作用方式

神经元活动的一般规律:神经系统神经元,神经纤维突触神经递质.受体学说.神经营养性作用神经元是神经系统的结构与功能单位。
结构上大致都可分成细胞体和突起两部分,突起又分树突和轴突两种。
轴突往往很长,由细胞的轴丘分出,其直径均匀,开始一段称为始段,离开细胞体若干距离后始获得髓鞘,成为神经纤维。
习惯上把神经纤维分为有髓纤维与无髓纤维两种,实际上所谓无髓纤维也有一薄层髓鞘,并非完全无髓鞘。
(一)神经纤维传导的特征神经传导是依靠局部电流来完成的。
因此它要求神经纤维在结构和功能上都是完整的;如果神经纤维被切断或局部受麻醉药作用而丧失了完整性,则因局部电流不能很好通过断口或麻醉区而发生传导阻滞。
一条神经干中包含着许多条神经纤维,但由于局部电流主要在一条纤维上构成回路,加上各纤维之间存在结缔组织,因此每条纤维传导冲动时基本上互不干扰,表现为传导的绝缘性。
人工刺激神经纤维的任何一点引发冲动时,由于局部电流可在刺激点的两端发生,因此冲动可向两端传导,表现为传导的双向性。
由于冲动传导耗能极少,比突触传递的耗以小得多,因此神经传导具有相对不疲劳性。
(二)神经纤维传导的速度一般地说,神经纤维的直径越大,其传导速度也越大;有髓纤维的传导速度与直径成正比,其大致关系为:传导速度(m/s)=6×直径(μm)。
一般据说有髓纤维的直径是指包括轴索与髓鞘在一起的总直径,而轴索直径与总直径的比例与传导速度又有密切关系,最适宜的比例为0.6左右。
神经纤维的传导速度与温度有关,温度降低则传导速度减慢。
经测定,人的上肢正中神经的运动神经纤维和感觉神经纤维的传导速度分别为58m/s和65m/s。
当周围神经发生病变时传导速度减慢。
因此测定传导速度有助于诊断神经纤维的疾患和估计神经损伤的预后。
表10-1 神经纤维的分类(一)表10-2 神经纤维的分类(二)(三)神经纤维的分类1.根据电生理学的特性分类主要是根据传导速度(复合动作电位内各波峰出现的时间)和后电位的差异,将哺乳类动物的周围神经的纤维分为A、B、C 三类(表10-1)。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
神经元活动的一般规律:神经系统神经元,神经纤维突触神经递质.受体学说.神经营养性作用神经元是神经系统的结构与功能单位。
结构上大致都可分成细胞体和突起两部分,突起又分树突和轴突两种。
轴突往往很长,由细胞的轴丘分出,其直径均匀,开始一段称为始段,离开细胞体若干距离后始获得髓鞘,成为神经纤维。
习惯上把神经纤维分为有髓纤维与无髓纤维两种,实际上所谓无髓纤维也有一薄层髓鞘,并非完全无髓鞘。
(一)神经纤维传导的特征神经传导是依靠局部电流来完成的。
因此它要求神经纤维在结构和功能上都是完整的;如果神经纤维被切断或局部受麻醉药作用而丧失了完整性,则因局部电流不能很好通过断口或麻醉区而发生传导阻滞。
一条神经干中包含着许多条神经纤维,但由于局部电流主要在一条纤维上构成回路,加上各纤维之间存在结缔组织,因此每条纤维传导冲动时基本上互不干扰,表现为传导的绝缘性。
人工刺激神经纤维的任何一点引发冲动时,由于局部电流可在刺激点的两端发生,因此冲动可向两端传导,表现为传导的双向性。
由于冲动传导耗能极少,比突触传递的耗以小得多,因此神经传导具有相对不疲劳性。
(二)神经纤维传导的速度一般地说,神经纤维的直径越大,其传导速度也越大;有髓纤维的传导速度与直径成正比,其大致关系为:传导速度(m/s)=6×直径(μm)。
一般据说有髓纤维的直径是指包括轴索与髓鞘在一起的总直径,而轴索直径与总直径的比例与传导速度又有密切关系,最适宜的比例为0.6左右。
神经纤维的传导速度与温度有关,温度降低则传导速度减慢。
经测定,人的上肢正中神经的运动神经纤维和感觉神经纤维的传导速度分别为58m/s和65m/s。
当周围神经发生病变时传导速度减慢。
因此测定传导速度有助于诊断神经纤维的疾患和估计神经损伤的预后。
表10-1 神经纤维的分类(一)表10-2 神经纤维的分类(二)(三)神经纤维的分类1.根据电生理学的特性分类主要是根据传导速度(复合动作电位内各波峰出现的时间)和后电位的差异,将哺乳类动物的周围神经的纤维分为A、B、C 三类(表10-1)。
A类:包括有髓鞘的躯体的传入和传出纤维,根据其平均传导速度又进一步分为α、β、γ、δ四类。
B类:有髓鞘的自主神经的节前纤维。
C类:包括无髓鞘的躯体传入纤维(drC)及自主神经节后纤维(sC)。
2.根据纤维的直径的大小及来源分类将传入纤维分为I、Ⅱ、Ⅲ、N四类(表10-2),I类纤维中包括I a和I b两类。
(四)神经纤维的轴浆运输神经元的细胞体与轴突是一个整体,胞体和轴突之间必须经常进行物质运输和交换。
实验证明,轴突内的轴浆是经常在流动的。
轴浆流动是双向的,一方面部分轴浆由胞体流向轴突末梢,另一方面部分轴浆由轴突末梢反向流向胞体。
胞体内具有高速度合成蛋白质的结构,其合成的物质借轴浆流动向轴突末梢运输;而反向的轴浆流动可能起着反馈控制胞体合成蛋白质的作用目前知道,自胞体向轴突末梢的轴浆运输分两类。
一类是快速轴浆运输,指的是具有膜的细胞器(线粒体、递质囊泡、分泌颗粒等)的运输,另一类是慢速轴浆运输,指的是由胞体合成的蛋白质所构成的微管和微丝等结构不断向前延伸.在缺氧、氰化物毒化等情况下,神经纤维的有氧代谢扰乱使ATP减少到50%以下时,快速轴浆运输流动即停止,说明它是一种耗能过程。
逆向流动的速度约为快速顺向运输速度的一半左右。
有人认为,破伤风毒素、狂犬病病毒由外周向中枢神经系统转运的机制,可能就是逆向轴浆流动。
二、神经元间相互作用的方式(一)经典的突触概念神经元之间在结构上并没有原生质相连,每一神经元的轴突末梢仅与其他神经元的胞体或突起相接触,引相接触的部位称为突触。
主要的突触组成可分为三类:①轴突与细胞体相接触;②轴突与树突相接触;③轴突与轴突相接触(图1 0-1)。
在电子显微镜下观察到,突触的接触处有两层膜,轴突末梢的轴突膜称为突触前膜,与突触前膜相对的胞体膜或树突膜则称为突触后膜,两膜之间为突触间隙。
突触前膜和后膜较一般的神经元膜稍增厚,约7.5nm左右。
突触间隙约20nm左右,其间有粘多糖和糖蛋白(图10-2)。
在突触小体的轴浆内,含有较多的线粒体和大量聚集的囊泡(突触小泡)。
它们含有高浓度的递质(图10-3)。
不同突触内含的泡大小和形状不完全相同,突触小泡在轴浆中分布不均匀,常聚集在致密突起处。
由于突触传递功能有兴奋性和抑制性两种,因此有人认为,突触在形态上也可能存在两种类型。
兴奋性突触的前膜释放兴奋性递质,它对突触后膜的作用是产生兴奋性突触后电位;抑制性突触的前膜释放抑制性递质,它对突触后膜的作用是产生抑制性突触后电位。
一个神经元的轴突末梢一般都分支形成许多突触小体,与其后的神经元构成突触,所以一个神经元能通过突触传递作用于许多其他神经元。
另一方面,一个神经元的树突或胞体可以接受许多神经元的突触小体构成突触,因此一个神经元又可接受许多不同神经元的作用。
据估算,一个脊椎前角的运动神经元的胞体和树突上可有2000个左右突触,而一个大脑皮层锥体细胞则约有30000个突触。
(二)电突触神经元之间除了上述的经典突触联系外,还存在电突触。
电突触的结构基础是缝隙连接,是两个神经元膜紧密接触的部位。
两层膜间的间隔只有2-3nm,连接部位的神经元膜没有增厚,其旁轴浆内无突触小泡存在(图10-4)。
连接部位存在在沟通两细胞胞浆的通道,带电离子可通过这些通道而传递电信号,这种电信号传递一般是双向的。
因此,这种连接部位的信息传递是一种电传递,与经典突触的化学递质传递完全不同。
电突触的功能可能是促进不同神经元产生同步性放电。
电传递的速度快,几乎不存在潜伏期。
电突触可存在于树突与树突、胞体与胞体、轴突与胞体、轴突与树突之间。
图10-4 几种不同形式突触的模式图箭头示传递方向(三)非突触性化学传递由于荧光组织化学等新方法的应用,目前已明确除了经典的突触能进行化学传递外,还存在非突触性化学传递(non-synaptic chemical transmission )。
关于这方面的研究,首先在交感神经肾上腺素能神经元上进行。
实验观察到,肾上腺素能神经元的轴突末梢有许多分支,在分支上有大量的念珠状曲张体(varico sity)。
曲线体内含有大量的小泡(图10-5),是递质释放和部位。
一个神经元的轴突末梢可以具有30000个曲线体,因此一个神经元具有大量的递质释放部位。
但是,曲张体并不与效应细胞形成经典的突触联系,而是处在效应细胞附近。
当神经冲动抵达曲张体时,递质从曲张体释放出来,通过弥散作用到效应细胞的受体,使效应细胞发生反应。
由于这种化学传递不是通过经典的突触进行的,因此称为非突触性化学传递。
在中枢神经系统内,也有这种传递方式存在。
例如,在大脑皮层内具有直径很细的无纤维,这种纤维是去甲肾上腺素能性的,纤维分支上具有许多曲张体,能释放去甲肾上腺素递质;这种曲张体绝大部分不与支配的神经元形成经典的突触,所以进行的是非突触性化学传递。
又如在黑质中,多巴胺能纤维也有许多曲张体,且绝大多数也进行非突触性化学传递。
此外,中枢内5-羟色胺能纤维也能进行非突触性化学传递。
由此看来,单胺类神经纤维都能进行非突触性化学传递。
已知,非突触性化学传递也能在轴突末梢以外的部位进行,轴突膜也能释放化学递质(如释放胞浆中的乙酰胆碱),树突也能释放化学递质(如黑质中、树突可释放多巴胺)。
图10-5 交感神经肾上腺能神经示意图非突触性化学传递与突触性化学传递相比,有下列几下特点:①不存在突触前膜与后膜的特化结构;②不存在一对一的支配关系,一个曲张体能支配较多的效应细胞;③曲张体与效应细胞间的距离至少在20nm以上,距离大的可达几十微米;④递质弥散的距离大,因此传递花费的时间可大于1s;⑤递质弥散到效应细胞时,能否发生传递效应取决于效应细胞上有无相应的受体。
(四)局部回路神经元和局部神经元回路中枢神经系统中存在长轴突的神经元,也有大量短轴突和无轴突的神经元。
长轴突的神经元是投射性神经元,它们投射到远隔部位,起到联系各中枢部位功能的作用;其轴突末梢通过经典的突触联系和非突触性化学传递的方式,完成神经元间的相互作用。
短轴突和无轴突神经元不投射到远隔部位,它们的轴突和树突仅在某一中枢部位内部起联系作用;这些神经元称为局部回路神经元(local circuit neuron)从进化来看,动物越高等,局部回路神经元数量越多,它们的突起越发达。
局部回路神经元的活动可能与高级神经功能有密切的关系,例如学习、记忆等。
由局部回路神经元及其突起构成的神经元间相互作用的联系通路,称为局部神经元回路(local nuronal circuit)。
三、神经递质前文已述及突触传递是通过突触前膜释放化学递质来完成的(非突触性化学传递的情况也是如此)。
一个化学物质被确认为神经递质,应符合以下条件:①在突触前神经元内具有全盛递质的前体物质和合成酶系,能够合成这一递质;②递质贮存于突触小泡以防止被胞浆内其它酶系所破坏,当兴奋冲动抵达神经末梢时,小泡内递质能释放入突触间隙;③递质通过突触间隙作用于突触后膜的特殊受体,发挥其生理作用,用电生理微电泳方法将递质离子施加到神经元或效应细胞旁,以模拟递质释放过程能引致相同的生理效应;④存在使这一递质失活的酶或其他环节(摄取回收);⑤用递质拟似剂或受体阻断剂能加强或阻断这一递质的突触传递作用。
(一)外周神经递质1.乙酰胆碱:释放乙酰胆碱作为递质的神经纤维,称为胆碱能纤维节前纤维的递质也是乙酰胆碱。
节前纤维和运动神经纤维所释放的乙酰胆碱的作用,与菸碱样作用(N样作用);而副交感神经节后纤维所释放的乙酰胆碱的作用,也毒蕈碱的药理作用相同,称为毒蕈碱样作用(M样作用)。
2.去甲肾上腺素为交感神经节后纤维的递质。
在高等动物中由交感神经节后纤维释放的递质仅是去甲肾腺上素,而不含肾上腺素;因为在周围神经末梢中不含合成肾上腺素所必需的苯乙醇胺氮位甲基移位酶。
释放去甲肾上腺素作为递质的神经纤维,称为肾上腺素能纤维。
但是,不是所有的交感神经节后纤维都是肾上腺素能纤维,像支配汗腺的交感神经和骨骼肌的交感舒血管纤维却是胆碱能纤维。
3.嘌呤类和肽类递质自主神经的节后纤维除胆三能和肾上腺素能纤维外,还有第三类纤维。
第三类纤维末梢释放的递质是嘌呤类和肽类化学物质。
第三类纤维是非胆碱能和非肾上腺素能纤维,主要存在于胃肠(二)中枢神经递质1.乙酰胆碱前角运动神经元支配骨骼肌的接头处递质为乙酰胆碱乙酰胆碱肯定是中枢的递质,而且分布比较广泛。
2.单胺类递质是指多巴胺、去甲肾上腺,色胺多巴胺递质系统主要包括三部位:黑质-纹状体部分、中脑边缘系统部分和结节、漏斗部分。
去甲肾上腺素系统比较集中,极大多数的去甲肾上腺素能神经元位于低位脑干5-羟色胺递质系统也比较集中,其神经元主要位于低位脑干近中线区的中缝核内。