园林植物同源四倍体的遗传特性(精)
四倍体刺槐的特征特性及育苗技术

四倍体刺槐的特征特性及育苗技术摘要阐述了四倍体刺槐的来源、生长特点、作用,介绍了砧木育苗和组培育苗等育苗技术,以供参考。
关键词四倍体刺槐;特征特性;育苗技术四倍体刺槐是韩国山林厅林木育种研究所经过人工诱变而来的刺槐新品种。
它具有速生、叶大且无刺、适应性强等优良特性。
四倍体刺槐的引进、试种和繁育技术被列为国家林业局“948”项目之一。
1 特点及应用1.1 特征特性四倍体刺槐在海拔2 000m以下、降水量不低于200mm、年平均气温不低于3℃、坡度在50°以下的地区均能生长。
饲料型四倍体刺槐具有速生、叶大、条粗、枝密、无刺等特点,叶厚为普通刺槐的2倍以上,叶肉肥厚,叶总干重及复叶干重均为普通刺槐的1.5倍以上,叶内粗蛋白、粗脂肪及灰分含量分别为优质匈牙利刺槐的118%、126%、180%,宜作动物饲料。
韩国的畜牧业主要以此为饲料。
用材型四倍体刺槐具有速生、抗旱、抗烟、耐盐碱、木材坚韧、纹理细致、有弹性、耐水湿、抗腐朽等特点,是保持水土、防风固沙、改良土壤的先锋树种与重要的用材树种。
尤其是矮化灌木类的四倍体刺槐,更为耐瘠薄,特别适合用作防沙治沙及滩涂开发。
1.2 生长特点经试验分析,四倍体刺槐与普通刺槐无性系相比,主要特点有:速生用材型与普通刺槐无性系相似,速生性较明显。
一年生嫁接苗高度可达 3.0m,地径 3.5cm;三年生树高5~7m,胸径6~9cm。
十八年生树高达19m,胸径24.5cm,单株材积0.283m3。
饲料型树枝无刺,呈灌木状,主干不明显,叶片是普通刺槐的2倍以上。
叶色浓绿,叶肉肥厚,复叶的干重和单株的总干重均为普通刺槐的1.5倍以上,叶富含多种维生素和矿物质。
此外,还含有钙、磷等牲畜所需要的微量元素,非常适宜作动物饲料。
1.3 应用价值四倍体刺槐的叶片和嫩枝是优良的禽畜饲料,广泛种植,可大力发展地方畜牧产业;生长快,干形通直,木材坚韧,纹理致密,抗腐朽,可广泛用于建筑业、矿产业,是营造速生丰产混交林和用材林的首选树种;根萌蘖性强,根系发达,具有根瘤菌,可起到改良土壤、增加土壤有机质含量和形成土壤团粒结构的作用;耐贫瘠,耐低温、烟碱,速生丰产,是优良的水土保持、沙漠化治理、防风固沙和“四旁”绿化的先锋树种;四倍体刺槐还是优质的蜜源植物,可用于发展地方养蜂业。
第七章 同源四倍体 ppt课件

2察,具40条染色体的4n玉米,产生的雄配 子中含有20条染色体的占42%,其余均为非整倍体。
Dawson(1962)研究,同源四倍体番茄的花粉 (n=24)只占75%,其它25%的花粉含有多于或少于
n=24的染色体数。
2020/10/28
14
○
2020/10/28
15
2、配子传递
产生的非整倍体配子,雌雄配子成活率相近,但 传递率不同,因为雄配子有竞争作用,所以没有雌配
子传递率高,也即整倍体配子通过花粉传递率高。
2020/10/28
16
同源四倍体后代当中,染色体数目存在差异:
例如,Randolph 1935 报告
4n玉米后Ⅰ20/20分离的占2/3
后代中 2n = 40
器官和组织变大。
2、代谢活性改变 四倍体细胞成份有所改变,如,水份、蛋白质、叶
绿素、纤维素、生长素等(有增有减)。
例如,生长素含量 4n<2n
∴一般4n生长缓慢,花期推迟,分蘖和分枝减少。大多数
水果、蔬菜,维生素C含量4n>2n;种子植物蛋白质含
量4n>2n;玉米,类胡萝卜素含量4n>2n,高43%
75-95%, 黑麦种在瑞典,结实率约为65%,在美国加
洲结实率达90%以上。
2020/10/28
18
U.Lsing 1967 研究四倍体大麦,F1优势不同结实率不同, F1优势越大,结实率越高,说明其非整倍体发育较好。
引起花粉和胚珠不育的原因不尽相同:
花粉育性:取决于小孢子发生过程
结实性:取决于(1)大孢子发生过程;
4n新种
(2)减数分裂异常 性母细胞(2n)→2n配子→有性结合→少数4n个体
不同物种或基因型产生2n配子频率不同:
园林植物遗传育种课件:园林植物性状遗传

花径、重瓣性与数量性状
花的直径表现为数量性状,重瓣性在大多数情下也 表现为数量性状,其遗传遵循数量性状的遗传规律。
如:牡丹、菊花等。
返回 目录
增加花径的途径
花径是数量性状,其遗传遵循数量性状的遗传机理, 利用这一属性,通常采用下列方法来增加花径。
❖ 改进栽培条件。充足的水肥,适当的管理及精心培育能使 花径变得更大。
C类突变体表型示意图
返回 目录
花型的发育 花型的发育受基因控制。
返回 目录
第二节 花色与彩斑的遗传
1.花色的形成 2.花色遗传 3.彩斑的遗传
屈金香
兰花
目录
花色的形成
花色的形成与花部所含色素种类和花被片海棉组织层的 物理特性有关,而主要受所含色素种类的影响。
花的色素有三大类群:类胡萝卜素、类黄酮、花青素。每 一类色素均包括很多种类。植物的不同花色是由不同的色素组 成的。
返回 目录
第四节 株型遗传
1.株型的多样性 2.植物激素与株型 3.株型的遗传
目录
株型的多样性
❖ 按植物的分枝特性,可分为乔木、灌木、藤本。 ❖ 按植株的高矮, 有乔化、矮化之分。 ❖ 按枝条的姿势可分为直枝、垂枝、曲枝。
如将上述因子组合则有十几种株型。
返回 目录
植物激素与株型
植物的株型,主要与其自身激素平衡有关。 1.分支特性 乔、灌、藤之分主要是顶端优势的作用,而顶端
园林植物遗传育种学
目录
园林植物性状遗传
第一节 花的发育 第二节 花色与彩斑的遗传 第三节 花径与重瓣性遗传 第四节 株型遗传 第五节 抗性遗传
首页 结束
第一节 花的发育
植物的发育从胚开始: 地下部分(根) ← 根尖 ← 胚 → 茎尖 → 地上部分(茎、叶、花)
草棉同源四倍体的诱导与鉴定

草棉同源四倍体的诱导与鉴定李昱樱; 申状状; 荣二花; 郑赟; 吴玉香【期刊名称】《《山西农业大学学报(自然科学版)》》【年(卷),期】2019(039)006【总页数】5页(P40-44)【关键词】草棉; 多倍体诱导; 鉴定【作者】李昱樱; 申状状; 荣二花; 郑赟; 吴玉香【作者单位】山西农业大学农学院山西太谷 030801【正文语种】中文【中图分类】S334.3棉花属双子叶植物(Dicotyledons)、锦葵科(Malvaceae)、棉属(Gossypium)。
目前有51个种,其中46个为二倍体棉种(2n=2X=26),5个为异源四倍体棉种(2n=4X=52)[1]。
草棉(G. herbaceum L.)是二倍体,属于A染色体组栽培棉。
又名阿拉伯棉、小棉,也称非洲棉。
长期以来在棉属四倍体棉种起源的问题上,研究者普遍认为草棉和亚洲棉是四倍体棉种A基因组的祖先供体种[2]。
草棉虽然对现代纺织工业贡献不大,但与异源四倍体棉花A基因组密切相关,为异源四倍体棉花提供了纤维性状,草棉本身也具有其它棉花品种所没有的优良性状[3],如极早熟性、抗角斑病等,因而草棉在AD组棉花进化过程中起着十分重要的作用。
同时,A组亚洲棉和D组雷蒙德氏棉全基因组测序的完成[4,5],也为AD组棉种间种质互渗及异源四倍体进化的研究提供了更大的空间。
多倍化是物种形成的重要方式,蕴藏着丰富的遗传变异的潜力,多倍体生物不仅有很强的活力,还有很多性状都优于其二倍体亲本[6]。
多倍体研究对了解物种的起源和进化有十分重要意义,通常棉属被作为研究多倍体进化的模式植物,通过人工远缘杂交或加倍,可以为研究早期基因组互作和进化提供理论依据。
近年来通过陆地棉和斯特提棉的远缘杂交和染色体加倍,成功获得了染色体组完整的异源六倍体植株[7];以草棉做母本,雷蒙德氏棉做父本进行的远缘杂交也成功获得草雷杂种[8];也曾对亚洲棉[9]和草棉[10]进行过多倍体诱变。
四倍体泡桐冬季温室快速繁育及丰产栽培技术
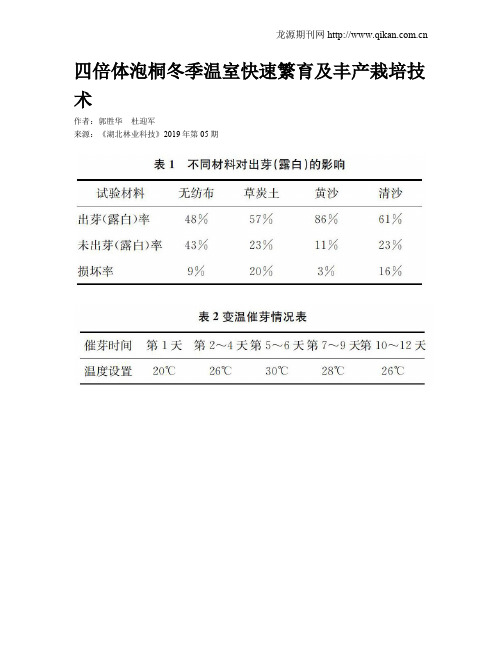
四倍体泡桐冬季温室快速繁育及丰产栽培技术作者:郭胜华杜迎军来源:《湖北林业科技》2019年第05期摘要:四倍体泡桐属玄参科泡桐属植物新品种,它是将二倍体泡桐染色体数目加倍培育的泡桐[1]。
泡桐为中国的特产树种,有悠久的栽培历史,是优良的速生用材树种,也是城市绿化园林景观树种,由于容易繁殖的特点,传统生产的无性繁殖造成泡桐性能退化,成材质量不高,病虫害严重等问题,影响了林农栽培的积极性。
四倍体泡桐新品种的培育,为泡桐产业的快速发展提供了新的材料支撑。
林木种苗标准化生产是林业产业发展的关键,造林栽培新技术的推广是林业产业创造效益的保证。
本文主要从四倍体泡桐采根催芽、容器基质配制,冬季温室容器育苗技术及丰产栽培管理等方面进行总结,为推广种植提供借鉴。
关键词:四倍体泡桐;快速繁育;容器育苗;栽培造林中图分类号:S753文献标识码:A文章编号:1004-3020(2019)05-0047-03传统的泡桐育苗,采取根插露地栽植培育。
因泡桐采取种根繁殖非常容易,而这种长期无性繁殖的培育方法造成了泡桐速生性能的退化,自然接杆率降低,丛枝病发病率增加。
河南农业大学泡桐研究所以毛泡桐P.tomentosa、兰考泡桐P.elongata、白花泡桐P.fortunei等无菌组培苗叶片为实验材料,利用秋水仙素处理,结合组织培养技术进行四倍体诱导,成功地培育了四倍体泡桐新品种[2]。
四倍体泡桐具有植物多倍体诸多优良特性,它具有材质优良、抗逆性强的优良特性,相比二倍体泡桐,它具有自然接杆率高,丛枝病显著降低等特性。
采用工厂化容器育苗有较高成活率,节省育苗周期,有利于机械化作业,促进苗木造林后快速生长等诸多优势。
冬季温室容器快速育苗,使泡桐造林周期提前了1年,使造林周期由一季拓展为三季,延长了造林周期[3]。
四倍体泡桐的工厂化育苗及标准化造林技术将助推泡桐产业快速发展。
1冬季温室快速育苗技术1.1采根及催芽1.1.1种源选择选用经河南省林木品种审定委员会审定的品种‘毛四’、‘兰四’。
蓖麻二倍体与同源四倍体光合特性和逆境生理指标的比较研究

蓖麻二倍体与同源四倍体光合特性和逆境生理指标的比较研究陆建农;施玉珍;李卫锦;刘柳彬;关雪燕;曾洁;陈森;汪亚菲;殷学贵【摘要】[目的]评价同源四倍体蓖麻的育种意义.[方法]以二倍体和同源四倍体蓖麻为材料,测定光合速率,过氧化物酶(POD)、超氧化物歧化酶(SOD)活性,及叶绿素、可溶性糖、可溶性蛋白、游离脯氨酸(Pro)、丙二醛(MDA)等含量,并比较上述指标的种间差异.[结果]同源四倍体蓖麻的叶绿素含量、净光合速率、叶片可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量、过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性均显著地高于二倍体,分别提高30.00%、53.47%、19.23%、25.22%、11.72%、262.07%和24.20%,丙二醛(MDA)的积累明显低于二倍体,降低30.57%.[结论]四倍体蓖麻可为蓖麻种质资源提供较宝贵新种质.%[Objectives] In order to evaluate the breeding significance of autotetraploidcastor.[Method] comparative studies were carried out on photosynthetic characteristics and stress physiological indexes between diploid and autotetraploid in castor in this paper.[Result] The chlorophyll content,the net photosynthetic rate,the soluble sugar content,the soluble protein content,the free proline content,the peroxidase (POD) and superoxide dismutase (SOD) activity in leaves of the autotetraploid were significantly higher than those in diploid,increased by30%,53.47%,19.23%,25.22%,11.72%,262.07% and 24.20%respectively.Meanwhile the malondiadehyde (MDA)accumulation in autotetraploid was significantly reduced by 30.57%.[Conclusion] Tetraploid castor provides valuable new germplasm for germplasm resources of castor.【期刊名称】《广东海洋大学学报》【年(卷),期】2018(038)001【总页数】4页(P87-90)【关键词】蓖麻;同源四倍体;光合特性;逆境生理指标【作者】陆建农;施玉珍;李卫锦;刘柳彬;关雪燕;曾洁;陈森;汪亚菲;殷学贵【作者单位】广东海洋大学农学院,广东湛江 524088;广东海洋大学化学与环境学院,广东湛江 524088;岭南师范学院生命科学与技术学院,广东湛江 524048;广东海洋大学农学院,广东湛江 524088;广东海洋大学农学院,广东湛江 524088;广东海洋大学农学院,广东湛江 524088;广东海洋大学农学院,广东湛江 524088;广东海洋大学农学院,广东湛江 524088;广东海洋大学农学院,广东湛江 524088【正文语种】中文【中图分类】S565.6蓖麻(Ricinus communis L.),大戟科(Euphorbiaceae)蓖麻属作物,是世界十大油料作物之一。
同源四倍体

Ⅱ式联会交换的结果,每条染色体上基因成员分别为:
① A1a3/ ②a3A1/③a2a4/④a4a3(即为① Aa/ ③Aa/②aa/ ④aa) 分离结果同Ⅰ式 Ⅲ式联会交换的结果,每条染色体上基因成员分别为: ① A1a4/ ②a4A1/③a2a3/④a3a2(即为① Aa/ ④Aa/②aa/ ③aa) 分离结果同Ⅰ式 所以,Aaaa总的完全均衡分离结果为:1AA:10Aa:13aa
§7-2 同源四倍体的细胞学行为及可育性 一、同源四倍体的细胞学行为 二、同源四倍体可育性
一、同源四倍体的细胞学行为
1、联合与分离
同源四倍体的同源染色体之间存在相同的亲合力,但 由于同源区段只能两两相互配对,所以,存在如下联合 形式:
Ⅳ
分离:2/2 3/1
Ⅱ+Ⅱ
2/2
Ⅲ+Ⅰ
2/2 3/1 2/1
归纳:
• AAAa完全均衡分离结果 • 同理推出AAaa结果 • Aaaa结果
AA:Aa:aa = 13:10:1 AA:Aa:aa = 2:5:2 AA:Aa:aa = 1:10:13
2) AAaa完全均衡分离:可改写为A1A2a3a4
如果都发生交换的话: Ⅰ式联会交换的结果,每条染色体上基因成员分别为: ① A1A2/ ②A2A1/③ a3a4/ ④a4a3 (即为① AA/ ②AA/③ aa/ ④aa)
(2)秋水仙加倍处理分生组织
(3)组培 + 秋水仙处理 可提高再生株中4n的频率
(4)体细胞杂交 两种体细胞原生质体融合→诱发再生植株→异源四倍体。 烟草、茄科和十字花科植物已获成功。
二、表型效应
1、巨大型: 同源四倍体,细胞体积变大,分生组织更明显,使
器官和组织变大。 2、代谢活性改变
小果型西瓜同源四倍体的诱导及其特性研究
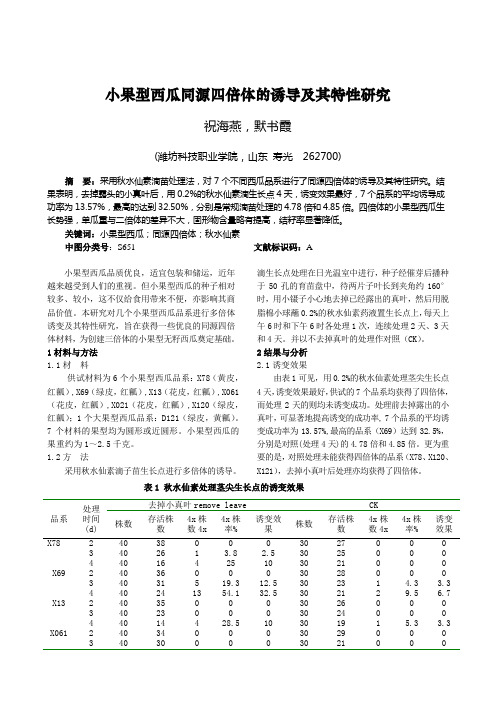
小果型西瓜同源四倍体的诱导及其特性研究祝海燕,默书霞(潍坊科技职业学院,山东寿光 262700)摘要:采用秋水仙素滴苗处理法,对7个不同西瓜品系进行了同源四倍体的诱导及其特性研究。
结果表明,去掉露头的小真叶后,用0.2%的秋水仙素滴生长点4天,诱变效果最好,7个品系的平均诱导成功率为13.57%,最高的达到32.50%,分别是常规滴苗处理的4.78倍和4.85倍。
四倍体的小果型西瓜生长势强,单瓜重与二倍体的差异不大,固形物含量略有提高,结籽率显著降低。
关键词:小果型西瓜;同源四倍体;秋水仙素中图分类号:S651 文献标识码:A小果型西瓜品质优良,适宜包装和储运,近年越来越受到人们的重视。
但小果型西瓜的种子相对较多、较小,这不仅给食用带来不便,亦影响其商品价值。
本研究对几个小果型西瓜品系进行多倍体诱变及其特性研究,旨在获得一些优良的同源四倍体材料,为创建三倍体的小果型无籽西瓜奠定基础。
1材料与方法1.1材料供试材料为6个小果型西瓜品系:X78(黄皮,红瓤),X69(绿皮,红瓤),X13(花皮,红瓤),X061(花皮,红瓤),X021(花皮,红瓤),X120(绿皮,红瓤);1个大果型西瓜品系:D121(绿皮,黄瓤)。
7个材料的果型均为圆形或近圆形。
小果型西瓜的果重约为1~2.5千克。
1.2方法采用秋水仙素滴子苗生长点进行多倍体的诱导。
滴生长点处理在日光温室中进行,种子经催芽后播种于50孔的育苗盘中,待两片子叶长到夹角约160°时,用小镊子小心地去掉已经露出的真叶,然后用脱脂棉小球蘸0.2%的秋水仙素药液置生长点上,每天上午6时和下午6时各处理1次,连续处理2天、3天和4天。
并以不去掉真叶的处理作对照(CK)。
2结果与分析2.1诱变效果由表1可见,用0.2%的秋水仙素处理茎尖生长点4天,诱变效果最好,供试的7个品系均获得了四倍体,而处理2天的则均未诱变成功。
处理前去掉露出的小真叶,可显著地提高诱变的成功率, 7个品系的平均诱变成功率为13.57%,最高的品系(X69)达到32.5%,分别是对照(处理4天)的4.78倍和4.85倍。
第七章 同源四倍体

3、对环境条件要求改变
4、改变自交不亲和性 二倍体自交不亲和系(2n)加倍→4n→自交不亲和性
变弱或消灭,在配子体不亲和性中表现更甚。
§7-2 同源四倍体的细胞学行为及可育性 一、同源四倍体的细胞学行为 二、同源四倍体可育性
子传递率高,也即整倍体配子通过花粉传递率高。
同源四倍体后代当中,染色体数目存在差异:
例如,Randolph 1935 报告
4n玉米后Ⅰ20/20分离的占2/3
后代中 2n = 40
占50%
=37-39
18.5
=41-42
31.5
苏联4n黑麦 2n = 28
89.5%
2n≠28
10.5%
二、同源四倍体可育性
第七章 同源四倍体
§7-1 同源四倍体的产生途径及表型效应
一、产生途径 二、表型效应
一、产生途径
1、自然产生
(1)2n体细胞自然加倍→4n细胞或局部组织→无性繁殖(扦插或嫁接)
4n新种
(2)减数分裂异常 性母细胞(2n)→2n配子→有性结合→少数4n个体
不同物种或基因型产生2n配子频率不同:
例如,芸苔属
2、AAaa
3、Aaaa
表7-4 同源四倍体以染色体和染色单体分离后代预期结果
上述分离结果只是理论上的,而实际分离 结果往往更接近后者。因为基因与着丝点之 间总有一定的距离,要发生一定的交换,而 交换也并非所有的性母细胞中都发生交换, 所以也不可能都按染色单体进行随机分离, 因而出现下面第三种分离情况。
后Ⅱ,着丝点一裂为二,复制的两成员(染色单体)彼 此分向两极,这叫均衡分离。减数分离的前后两次细胞分裂, 分别称为前减数和后均衡。
同源四倍体玉米的细胞遗传学研究

关 词三 旦 里 竖 苎生 减 分 育 移 键 塑擅 塑 敷 裂 性 胞 任 : 荽
细胞遗 传和 育种 学者认 为利用 多倍体 ,在改进 某些 作物产 量上是重 要 的手段 之一 .它是 为 了增加 细 胞 桉构 造 的成 分 —— 染 色体 的数量 ( 而拟定 的 )作用于 细胞分 裂 的方 法 .这 是现 代实 验遗 传学和 细胞 学 的一个 重要发 现 . n 经查 阅有关 的 资料 ,多倍体 育种 已在三个 不 同的方 面成 功地 应 用于 植 物育种 中 .首
置载玻 片 上 .挤压 .滴 加 K -I 液染 色 1 I 溶 O分钟 ,显徽 镜下观 察 .有 活性 的 花粉粒 被染成 蓝色 ,无 活性
的花 粉粒不 被染色 .分 别计数 并 随机测 量 l ~2 0 O十花 粉直径 的数值 ,求其 平均 值 。 1 2 3 根尖 细胞 染 色体 的观 察 .按李 懋学染 色体 研究技 术 。 .. 】 3 同源 四倍 体玉 米结实率 的测 定 . 从 收 获的 同潭 四倍体 玉米 穗 中随机抽 取 l 十 穗子进行 统计 完全成 熟的粒 数 .求其结 实率 百分 数 。 0
注 ・上 表 中 的 数 字 为 1 0十 细 胞 的 平 均 数 0
2 { 同源四倍体 玉米 结 实率 的 测定 . 以收 获的 同源 四倍体 玉米 穗中随 机抽取 1 个 ,通过 统计 结实率 为 8 2 ( 0 .4 详见 表 3 ,结果说 明诱 ) 变后 的 同源 四倍 体 玉米结 实率 相当低 。 关于 多倍体 结实率 降 低 ,目前主 要有两 方面 的解释 . 一方 面是观 察到 减数 分裂过 程 中染 色体行 为异 常 t 以就跟 着 出现配子 的败育 现象 ,另一方 面的解释是 认 为诱变 多倍体 能育性 降低是 由于 它 的遗 传性 所
同源四倍体荞麦的细胞遗传学研究

( 一)从许多文献报道看, 不同作物的同源
四倍体的结实率是不相同的,即使同一作物的
质[ ad p ] 4 。R no h石研尤中发现, l 四倍儿中的不 育性,不可能归于与多价体有关的减数分砚的 不规则性, 高度可育和高度不育的玉米, 减数分 裂的染色体行为以及其配子体的染色体数最关
大。如榆林县 18 9 7年荞麦盛花期时遇到严 重
干旱,又伴有 4 天 3-3℃ 的高温,结果 -5 0 5 二倍体荞麦和四倍体荞麦的结实率都明显低于 18 96年 ( 3。同一品种开花多时结实率低, 表 ) 开花少时结实率高。四倍体荞麦与二倍体的结 实率比较无明显差异, 有时还有提高 ( 30 表 )
45 8 - 61 6 .3 .7 7 7 7. 0 5 7 7. 5
38。1 9 8. 77 72.1 . 0
大麦
莫斯 科 1 1 2
和稻 4 2 2 水稻
粉育性结果表明, 四倍体荞麦花粉育性为8 %, 5 二倍体荞麦的花粉育性: 9 q( I, X 3 表 ) _ '
表 1 四倍体 荞麦和二 倍体荞麦花 粉母 细胞减数 分裂时 染色体行为及花 粉育性比较
不同品种的同原四倍体的结实率也有差异 ( 表 系基本上是相 同的〔 。减数分裂中都是二价休 ) 3 1 2。从表2中可以看出, ) 高粱、 大麦、水稻同源 的同源四倍体一希却是高度不育的「 首 切。文献报 四倍沐的结买率都高于二倍体荞麦的结实 率, 道,影响水稻同沁四倍体结实率的主要因索是 可见养麦本身均结实率极低。 遗传因素,在理论上肯定了选择的效果f 事 , , 。 ( 二)荞麦结实率极 低与减数分裂 不 规 则 实上对于同源四倍体荞麦逐代进行选择也是有 性没有规律性的联系。观察四倍体荞麦的花粉 效果的f。但是影响荞炭结实率的主要因素不 ‘ 。 〕 母细胞减数分裂过程中具落后染色体的细胞数 是配子肩一 性。 及不育花粉率均高于二倍休荞麦( 1,这是 表 ) ( 三)荞麦 ( 包括同源四倍体荞麦) 结实率 极低的主要原因是由荞麦自身的花器结怡,外 母细胞只占4 %, . 2 0 不育花粉率也只占1%, 5 如 哭气候等外界决定的。国内也有学者认为荞麦 果不育胚囊也占同样的百分数,则在理论上由 产量不高的主要原因,除了亲本的遗传基础和 此造成的不实率应为 4 一巧%之间, . 2 0 但事实 外界自然条件之外,与 花器的形态结构和开花 上, 无论是四倍体荞麦还是二倍体荞麦, 不实率 生物学 特性 关系也很 大〔。 习 均在 9呢 以上,远远超过不育胚囊的百分数。 0 ( 四)四倍体荞麦比二倍体荞麦渭 产的 主
园林植物同源四倍体的遗传特性

园林植物同源四倍体的遗传特性摘要多倍体在园林植物中广泛存在,由于多倍体具有一些独特的特征,已逐渐成为广大遗传育种学家研究的热点。
就同源四倍体与相应二倍体核DNA、雌雄配子育性、受精过程的差异;同源四倍体结实率(或育性)降低的原因及其随世代推移的变化;叶绿体、线粒体与核基因的关系等方面进行了总结和探讨。
关键词园林植物;同源四倍体;多倍化;遗传特性一个物种生物体内细胞染色体组数达到3组或3组以上时,称为多倍体。
多倍体作为高等植物中存在的一种普遍现象,在园林植物中广泛存在,如景天科、蔷薇科、锦葵科、禾本科、鸢尾科内多倍体种特别多。
细胞核内染色体组加倍以后,常带来一些形态和生理上的变化,这些改变都与基因剂量有关。
一般表现在叶大、茎粗、花大、色浓;育性低;抗逆性强;碳水化合物、蛋白质、维生素、植物碱等表现偏高。
如四倍体紫罗兰、桂竹香芳香性强、蜜腺多。
自然界的多倍体的产生多出现在分布区的一些边缘地带,多在气候条件恶劣的地区,这些地区多倍体的出现常伴随着抗逆性的相对提高,如报春花原产温带,我国云南很多,原始种为二倍体,而新生的异源四倍体分布在二倍体区域内的高山上,三倍体和八倍体分布在更北或更南的高山上,而十四倍体生长在极地。
20世纪30年代,自人们发现用秋水仙素诱导多倍体的方法以来,育种家们在植物倍性育种方面作出了较多的探索,形成了一些人工多倍体的商业品种。
在花卉方面,矮牵牛、金鱼草、鸡冠花等多倍体植物多表现为叶片肥厚、花色艳丽、花期长、花瓣多等特点,观赏价值得到了提高;林木方面,四倍体桑树及刺槐在生长量及抗逆性方面都较之二倍体对照有了较大提高。
同源四倍体育性的降低是由多方面原因引起的,既有同源四倍体核内染色体在减数分裂时,染色体间配对不正常致使育性降低,也有其他原因。
植物的育性主要受核内染色体减数分裂及雌雄配子的育性、授粉受精过程等方面的影响。
由于染色体倍性的变化,植物同源四倍体在前述几个方面较其起源二倍体均有所差异。
同源四倍体水稻糯性基因的遗传分析1)

H R D S ei ) 0 ) - 1 18 E E fA ( i g 1 ( :9 1 8 Bj n 1 - 9
同 倍体水稻糯性基因的遗传分析” 源四
宋 文昌 陈 勇 张 华 吴 瑜 志 玉 德
〔 中国农业科 学院作物所 . 北京)
系 H 8, 4 H 9 杂交, : 86 H 9 和 76 8 F代种子为非 传。他们指出糯性胚乳是单基因隐性 遗 传的。 糯, 用碘液染呈蓝黑色 。播种后从F 植株上得 : ,代种子, 以株为单位收获脱粒, 用小型鲁 二倍体杂种 W 。 的自交后代有 3 x 种基因型, 到 F 显隐性比例为31 : 。同源四倍体糯稻在 。 二位 谷机碾出糙米,先从外观上将糯米与非糯米区 点上有4 个等位基因,它与非糯稻的杂种后代 分开,而将难以区别的米粒顶端切去一些,用 的基因型种类和表现型比例要比二倍体复杂得 I I溶液鉴别。分别统计每株的糯、 - K 非糯的位
1 。如果是同一大样本分布中左右两极端部分
的偏差,那么这两个偏差的非糯之和与糯米之 和的比例理论上仍应为 3 :。计算结果, 1 51 这 7 株共有非糯米 59 粒、糯米 18 90 6 粒,比 例为
- 30
一14 -28
16 2
192 5 65 330 63 427 651 232 237 245 255 29 7 69 4 544 1 0 00
一7
50 江米 XH76 0拉 9 50 0 粒江 米 X 76 H 9 50 江米 XH76 0粒 9 50 江米 XH76 0粒 9
35 6 1909 971
345 1858 941 14 46
11 51 30 34
024 .41 103 .92 019 .50 000 .03
同源四倍体刺槐花序变异表型观察与分析

曾经受 到过 限 制 。这 一 局 面 在 Ba ele等 ( 9 7 lks e 13 ) 成 功利 用秋 水仙 素诱 导体 细 胞染 色体 加倍 后得 到 了
收 8— 5 2 1 —0 9 21 0 2 。
基 金 项 目 :国 家 自然 科 学 基 金 项 目( 17 6 9 3 60 3 ; 业 公 益 行 业 专 项 ( 0 14 1 ) 3 10 2 ; 0 40 6) 林 2 10 0 3 ;北 京市 农 委 项 目(0 8 14 北 京 林 业大 学 2 0 0 0 );
Ph n t pi b e v to n e o y c O s r a i n a d Ana y i fI fo e c n e l ss o n l r s e c
V a i to f Aut t t a 0 d Ro i i e d a a i ra i n O o e r pl i b n a ps u o c ca
d fe e c s i h e g h o f r s e c e we n a t t t p o d a d d p o d R.p e d a a i i r n e n t e l n t fi l e c n e b t e u o e r l i n i l i n o a s u o c ca,b tt e n mb r o o e s wa u h u e ff r t s l s g ii a t r n a t t ta l i h n t e d p o d i n f n l mo e i u o e r p o d t a h i l i .Th o e s o h u o e r p o d R.p e d a a i n d fe e tp s t n c y e f r t ft e a t t ta l i l s u o c ca i i r n o i o s i
南方泡桐同源四倍体的诱导及其体外植株再生

南方泡桐同源四倍体的诱导及其体外植株再生范国强;魏真真;杨志清【摘要】[目的] 用秋水仙素诱导出南方泡桐同源四倍体,建立其体外植株高效再生系统,为泡桐新品种的培育奠定基础.[方法] 以二倍体南方泡桐幼苗叶片为外植体,预培养泡桐不同时间(0,8,16 d)后,置于含不同质量浓度(5,10,20 mg/L)秋水仙素的固液双层培养基上处理泡桐不同时间(24,48,72 h),以诱导其同源四倍体植株,并通过根尖细胞染色体计数和叶片单细胞DNA相对含量的测定,进行变异植株的倍性分析;同时,以其四倍体叶片、茎段和叶柄为外植体,利用不同植物激素不同质量浓度的组合培养基,筛选建立体外植株再生系统的最适培养基.[结果] 秋水仙素质量浓度和处理时间对南方泡桐同源四倍体诱导率影响显著,外植体预培养时间影响不显著;在含10 mg/L秋水仙素的MS培养基上处理72 h,未经预培养叶片的四倍体诱导率高达18.8%;同源四倍体幼苗叶片较二倍体大而且厚,叶片单气孔器变大,孔密度变小,叶绿素含量和SOD活性升高,MDA含量和POD活性降低.MS+0.1 mg/L NAA+8 mg/L BA、MS+0.3 mg/L NAA+12 mg/L BA和1/2 MS分别是其愈伤组织诱导、芽诱导和根诱导的适宜培养基.[结论] 诱导获得了南方泡桐同源四倍体,建立了同源四倍体南方泡桐的体外植株再生系统;叶片是同源四倍体南方泡桐体外植株再生的最佳外植体.【期刊名称】《西北农林科技大学学报(自然科学版)》【年(卷),期】2009(037)010【总页数】8页(P83-90)【关键词】南方泡桐;秋水仙素;同源四倍体;体外植株再生【作者】范国强;魏真真;杨志清【作者单位】河南农业大学,泡桐研究所,河南,郑州,450002;河南农业大学,泡桐研究所,河南,郑州,450002;河南农业大学,泡桐研究所,河南,郑州,450002【正文语种】中文【中图分类】S792.43;S722.3+5多倍体植物具有生物量大和抗逆性强等特点,自20世纪30年代德国首次育成了四倍体黑麦以来,国内外科技工作者对其进行了大量研究,并培育出了多种多倍体植物[1-16],有的多倍体植物已在农林业生产上产生了显著的经济和社会效益[17-18]。
园艺植物育种学》复习资料--整理版-改动

园艺植物育种学一.名词解释 1. 品种:具有在特定条件下表现为不妨碍利用的优良、适应、整齐、稳定和特异性的家养动植物群体。
2. 良种:常说的良种有两层含义:一是优良品种;二是优良种子,即优良品种的优良种子。
即优良品种的优良种子。
3. 种质资源:即携带种质的材料或载体的总称。
即携带种质的材料或载体的总称。
4. 引种:人类为了某种需要把植物从其原分布区移种到新的地区。
人类为了某种需要把植物从其原分布区移种到新的地区。
5. 无性系: 指具有相同遗传组成的植物利用无性繁殖方式所得来的群体。
指具有相同遗传组成的植物利用无性繁殖方式所得来的群体。
6. 无性系品种:采用无性繁殖方法育成的遗传组成同质,个体杂合的一类品种。
采用无性繁殖方法育成的遗传组成同质,个体杂合的一类品种。
7. 自交系品种:群体遗传组成基本同质,个体基本纯合的可用于生产的群体群体遗传组成基本同质,个体基本纯合的可用于生产的群体8. 杂交种品种:指用遗传上纯合的亲本在控制授粉条件下生产特定组合的一代杂种群体指用遗传上纯合的亲本在控制授粉条件下生产特定组合的一代杂种群体9. 遗传力:广义—基因型方差占表现型方差的百分率;狭义—基因型加性方差占表现型方差的百分率。
基因型加性方差占表现型方差的百分率。
10. 芽变:发生在芽内分生组织细胞中的突变,属于体细胞突变的一种。
发生在芽内分生组织细胞中的突变,属于体细胞突变的一种。
11. 异花授粉植物:在人工选择的情况下构成的一个遗传基础比较复杂又在主要经济性状相对一致而保持遗传平衡的异质群体。
传平衡的异质群体。
12. 有性杂交育种:经过有性杂交途径获得新品种的过程经过有性杂交途径获得新品种的过程13. 组合育种: 又叫常规杂交育种,是通过人工杂交,把分散于不同亲本上的优良性状组合在杂种中,对其后代进行多代培育选择,获得基因型纯合或接近纯合的新品种的育种途径。
后代进行多代培育选择,获得基因型纯合或接近纯合的新品种的育种途径。
青花菜同源四倍体与二倍体农艺性状及营养品质比较

青花菜同源四倍体与二倍体农艺性状及营养品质比较李俊星;张蜀宁;刘惠吉;侯喜林【摘要】以同源四倍体及二倍体青花菜为材料,比较研究农艺性状及营养品质.结果表明:四倍体青花菜维生素C含量极显著高于二倍体,增加25.6%,可溶性蛋白质、可溶性糖含量显著高于二倍体,干物质积累量、叶绿素a+b含量与二倍体无显著差异;四倍体植株表现出巨大型,花球单球重比二倍体增加15.7%;最大叶片重比二倍体增加达70.5%,株高、茎粗、开展度显著高于二倍体,分别增加6.1%、16.5%、12.6%;四倍体熟期比二倍体提早了5 d.【期刊名称】《江苏农业科学》【年(卷),期】2010(000)003【总页数】2页(P175-176)【关键词】青花菜;同源四倍体;农艺性状;营养品质【作者】李俊星;张蜀宁;刘惠吉;侯喜林【作者单位】南京农业大学/农业部南方蔬菜遗传改良重点开放实验室,江苏南京,210095;南京农业大学/农业部南方蔬菜遗传改良重点开放实验室,江苏南京,210095;南京农业大学/农业部南方蔬菜遗传改良重点开放实验室,江苏南京,210095;南京农业大学/农业部南方蔬菜遗传改良重点开放实验室,江苏南京,210095【正文语种】中文【中图分类】S635.301青花菜(Brassica oleracea var. italica Plenck)是以绿色或紫色花球供食用的蔬菜,为十字花科芸薹属甘蓝种的一个变种。
20世纪90年代末青花菜在我国已成为一种新兴的优质蔬菜。
营养学分析表明,青花菜的营养价值远远高于一般蔬菜。
一般认为同源多倍体其抗性、品质、产量都优于二倍体,上述研究在小白菜[1]、黄瓜[2]等蔬菜有报道。
本实验室在获得同源四倍体青花菜以来,已对其光合特性[3]、花粉母细胞减数分裂及雄配子体形成[4]、核型分析[5]进行了深入研究。
对四倍体青花菜的农艺性状及营养品质研究还未见报道。
本研究旨在为优质四倍体青花菜品种选育及应用奠定理论基础。
008-园林植物育种学-倍性育种

分类
依据染色体组的来源不同分类 同源多倍体 异源多倍体
根据染色体数整数倍还是非整数倍性
整数多倍体 非整数多倍体(异数多倍体)
(1)同源多倍体 形成多倍体的染色体组来源于同一个物种。
以A代表一个染色体组 AA为同源二倍体 AAA为同源三倍体 AAAA为同源四倍体
如美国已育成同源四倍体的金鱼草、麝香百合等。
(3)异数多倍体(非整数多倍体) 指细胞中染色体数目有零头的多倍体。
例如栽培菊花大多为六倍体(2n=6x=54) 有的为47条染色体(即5x+2) 有的为71条染色体(即8x-1)
二、多倍体的特点
1、巨大性 在体形和细胞上都表现出明显的巨大性:叶片 变宽增厚、茎粗壮;花、果实、种子增大等。 多倍体形态上的巨大性还表现在气孔与花粉的
注射法
采用微量注射器将秋水仙素溶液注入植株顶芽或侧 芽中。
复合处理法
秋水仙素与辐射复合处理
如对好望角苣苔属植物的处理 先用秋水仙素处理,再用X射线照射,染色体加倍率达到 60% 单独用秋水仙素处理时为30%。
人工诱导多倍体主要采用化学法
秋水仙素诱导多倍体的方法
(1)诱导多倍体材料的选择 Nhomakorabea 染色体倍数较低的植物; 染色体数目极少的植物; 异花授粉植物; 能利用根、茎、叶等进行无性繁殖的植物; 杂种后代(包括远缘杂种和常规杂种)。
(2)秋水仙素的理化性质、配制与贮藏
来源:从百合科植物秋水仙 (Colchicum autumnale)的根、茎、 种子等器官中提炼出来的一种植物碱. 分子式:C22H25O6NO6· 1.5H2O。 物理性质:淡黄色粉末,针状无色结晶 (纯品),性极毒,融点为155℃,易 溶于水、酒精、氯仿和甲醛中,不易溶 解于乙醚、苯。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
园林植物同源四倍体的遗传特性
[ 10-12-14 10:34:00 ] 作者:尚瑛男杨鹏
鸣编辑:studa20
摘要多倍体在园林植物中广泛存在,由于多倍体具有一些独特的特征,已逐渐成为广大遗传育种学家研究的热点。
就同源四倍体与相应二倍体核DNA、雌雄配子育性、受精过程的差异;同源四倍体结实率(或育性)降低的原因及其随世代推移的变化;叶绿体、线粒体与核基因的关系等方面进行了总结和探讨。
关键词园林植物;同源四倍体;多倍化;遗传特性
一个物种生物体内细胞染色体组数达到3组或3组以上时,称为多倍体。
多倍体作为高等植物中存在的一种普遍现象,在园林植物中广泛存在,如景天科、蔷薇科、锦葵科、禾本科、鸢尾科内多倍体种特别多。
细胞核内染色体组加倍以后,常带来一些形态和生理上的变化,这些改变都与基因剂量有关。
一般表现在叶大、茎粗、花大、色浓;育性低;抗逆性强;碳水化合物、蛋白质、维生素、植物碱等表现偏高。
如四倍体紫罗兰、桂竹香芳香性强、蜜腺多。
自然界的多倍体的产生多出现在分布区的一些边缘地带,多在气候条件恶劣的地区,这些地区多倍体的出现常伴随着抗逆性的相对提高,如报春花原产温带,我国云南很多,原始种为二倍体,而新生的异源四倍体分布在二倍体区域内的高山上,三倍体和八倍体分布在更北或更南的高山上,而十四倍体生长在极地。
20世纪30年代,自人们发现用秋水仙素诱导多倍体的方法以来,育种家们在植物倍性育种方面作出了较多的探索,形成了一些人工多倍体的商业品种。
在花卉方面,矮牵牛、金鱼草、鸡冠花等多倍体植物多表现为叶片肥厚、花色艳丽、花期长、花瓣多等特点,观赏价值得到了提高;林木方面,四倍体桑树及刺槐在生长量及抗逆性方面都较之二倍体对照有了较大提高。
同源四倍体育性的降低是由多方面原因引起的,既有同源四倍体核内染色体在减数分裂时,染色体间配对不正常致使育性降低,也有其他原因。
植物的育性主要受核内染色体减数分裂及雌雄配子的育性、授粉受精过程等方面的影响。
由于染色体倍性的变化,植物同源四倍体在前述几个方面较其起源二倍体均有所差异。
植物同源四倍体的染色体倍性的变化或其变化过程而造成同源四倍体遗传物质或遗传物质之间的互作效应在DNA水平上与其起源二倍体的差异,研究这些变异有益于从分子水平上揭示同源四倍体表现型变异的原因。
1同源四倍体与相应二倍体核DNA、雌雄配子育性、受精过程的差异
1.1同源四倍体与其起源二倍体DNA遗传结构的差异
随着细胞生物学和分子生物学的不断发展,人们把细胞生物学和分子生物学分析技术引入到同源四倍体DNA遗传结构变异分析工作之中。
从报道的若干植物种类分析结果可以看出,在基因和同工酶水平上,同源四倍体的DNA遗传结构与其起源二倍体均有不同程度的差异,表现为DNA分子标记或同工酶的电泳图谱与起源二倍体相比有特异性谱带的增减。
综合产生上述差异的原因可归纳为以下2点:一是非加倍因素。
即伴随着染色体加倍而产生的染色体结构变异、基因突变及重组、核-质之间及合子胚与胚乳之间协调关系的变化等[1]。
二是加倍
因素,即纯粹的染色体数目增加。
桑树二倍体(2n=2x=28)与其同源四倍体
(2n=4x=56)在遗传结构上的AFLP分子标记结果表明,经秋水仙素诱变得到的同源4x与2x在DNA分子遗传结构上产生了一定程度的变异,但2x与同源4x间的遗传差异小于种间差异[2]。
1.2雌雄配子的育性及授粉受精过程的变异
由于减数分裂的不正常,同源四倍体的雌雄配子的育性均低于其起源二倍体,不同植物种类和同种植物不同类型或品种之间的配子败育率和败育方式不同。
同源四倍体的一部分花粉是双单倍体,因而其体积多大于起源二倍体植株的花粉(单倍体)。
同源四倍体的胚囊结构也因倍性的改变而发生了变异,表现为配囊内的卵细胞数量增加、助细胞及反足细胞的有无及形态的不相同等。
柑桔同源四倍体花粉育性和花粉粒大小的测定结果显示,其花粉育性平均为36%,约为起源
二倍体(62%)的60%,花粉粒体积平均为起源二倍体的2倍或双亲之和[3],南荻
同源四倍体花粉直径比起源二倍体花粉大20.65%[4]。