《现代分子生物学》第七章原核生物基因表达调控
分子生物学:基因表达调控

无论表达水平高低,管家基因较少受环境因素影响, 而是在个体各个生长阶段的大多数或几乎全部组织中 持续表达,或变化很小。区别于其他基因,这类基因 表 达 被 视 为 组 成 性 基 因 表 达 (constitutive gene expression)。
转录后调控,如RNA干扰。
特征:
(1)可遗传 (2)可逆性 (3)DNA不变
遗概 述 传 与 表 观 遗 传
(生殖系)
24
按对刺激的反应性,基因表达的方式分为: ➢ 基本(或组成性)表达 ➢ 诱导或阻遏表达
25
➢基因表达是受调控的。
在某一特定时期或生长阶段,基因组中只有一部分基 因处于表达。仅少数的基因处于高水平转录活性状态, 其余大多数基因处于静息状态或以极低的速率进行转录。 但是在一定的条件下可引发。
一、基本(或组成性)表达
基因表达调控
第一节 基因表达调控的概念及原理 第二节 基因表达的时间性及空间性 第三节 基因表达的方式 第四节 基因表达调控意义 第五节 表观遗传学——不涉及DNA序列改变的
基因表达
第一节 基因表达调控的概念及原理
一、基因表达调控是生命的必需 二、基因表达调控呈现多层次和复杂性 三、基因转录激活受到转录调节蛋白与启动子
可诱导基因在特定环境中表达增强的过程,称为诱导 (induction)。
如果基因对环境信号应答是被抑制,这种基因是可阻 遏基因(repressible gene)。可阻遏基因表达产物水 平降低的过程称为阻遏(repression)。
调控特点:可诱导或可阻遏基因 既受启动子或启动序列与RNA聚合 酶相互作用影响,又受其他机制 调节。因其调控机制中通常含有 特异刺激的反应元件。
分子生物学第七章原核生物基因表达调控

原核生物基因表达调控的特点
01
原核生物基因表达调控通常由特 定的转录因子、RNA聚合酶以及 其他调控蛋白介导,通过与DNA 的结合或解离来调节基因转录。
02
原核生物基因表达调控具有快速 响应环境变化的特点,能够在短 时间内调整基因表达模式,以适 应外界刺激和压力。
翻译后加工的调控
翻译后加工的调控
在翻译后加工阶段,新合成的蛋白质经过一系列修饰和加工,最终成为具有生物学活性的蛋白质。原 核生物通过控制翻译后加工酶的合成和活性来调控翻译后加工过程。此外,原核生物还可以通过控制 蛋白质的稳定性来影响其功能和表达水平。
总结
翻译后加工是基因表达调控的重要环节,原核生物通过控制翻译后加工酶的合成和活性,以及蛋白质 的稳定性来精细调控基因表达。
翻译延伸的调控
翻译延伸的调控
在翻译延伸阶段,核糖体沿着mRNA移动,将氨基酸组装成蛋白质。原核生物通过控制翻译延伸因子的合成和活 性,以及核糖体的合成和组装来调控翻译延伸。此外,原核生物还可以通过控制mRNA的结构和稳定性来影响翻 译延伸。
总结
翻译延伸是基因表达调控的重要环节,原核生物通过控制翻译延伸因子的合成和活性,以及核糖体的合成和组装, 以及mRNA的结构和稳定性来精细调控基因表达。
翻译起始的调控
原核生物通过控制翻译起始来调控基因表达。在翻译起始阶段, mRNA与核糖体结合,招募翻译所需的起始因子和其他成分。原 核生物通过控制起始因子的合成和活性,以及mRNA与核糖体的 结合来调控翻译起始。
总结
翻译起始是基因表达调控的重要环节,原核生物通过控制翻译起 始因子的合成和活性,以及mRNA与核糖体的结合来精细调控基 因表达。
分子生物学课程教学大纲

分子生物学课程教学大纲课程名称:分子生物学(Molecular Biology)课程编号:1313072215课程类别:专业课总学时数:68 课内实验时数:18学分:3.5开课单位:生命科学学院生物技术教研室适用专业:生物技术适用对象:本科(四年)一、课程的性质、类型、目的和任务分子生物学为高等学校生物技术专业学生必修的一门专业基础课,是从分子水平研究生命本质为目的的一门新兴边缘学科,主要研究核酸、蛋白质等生物大分子的功能、形态结构特征及其重要性和规律性的科学,是人类从分子水平上真正揭开生物世界的奥秘,由被动地适应自然界转向主动地改造和重组自然界的基础学科。
通过分子生物学的教学,应使学生了解分子生物学的发展历史以及最新研究成果;熟练掌握DNA的结构与功能、RNA在蛋白质合成中的功能、蛋白质的结构与功能、遗传密码及基因表达调控的本质;了解现代分子生物学基本研究方法,并能运用分子生物学的理论知识分析、研究和解决问题,为进一步学习有关专业课程及从事基因工程领域的研究工作奠定基础。
二、本课程与其它课程的联系与分工从学科角度来讲,分子生物学涵盖面非常广,与生物学、生物化学和细胞生物学、遗传学等生命科学课程有交叉,《生物化学》是先修课程。
三、教学内容及教学基本要求[1]表示“了解”;[2]表示“理解”或“熟悉”;[3]表示“掌握”;△表示自学内容;○表示略讲内容;第一章绪论第一节引言创世说与进化论[1];细胞学说[2];经典的生物化学和遗传学[3];DNA的发现[2]第二节分子生物学简史[1]第三节分子生物学研究的主要内容分子生物学的含义[3];DNA重组技术、基因工程技术概念[3];分子生物学研究的主要内容[3]第四节展望分子生物学的一些分支学科[1];分子生物学发展的趋势[1]重点:分子生物学的含义和研究内容难点:分子生物学的研究内容教学手段:多媒体教学教学方法:讲授法作业:1.简述阵德尔、摩尔根和沃森等人对分子生物学发展的主要贡献。
第七章原核生物的基因调控

第七讲原核生物的基因调控科学家把这个从DNA到蛋白质的过程称为基因表达(gene expression),对这个过程的调节就称为基因表达调控(gene regulation或gene control)。
要了解动、植物发展发育的规律、形态布局特征和生物学功能,就必需弄清楚基因表达调控的时间和空间概念,掌握了基因表达调控的奥秘,我们手中就有了一把揭示生物学微妙的金钥匙。
基因表达调控主要暗示在以下几个方面:①转录程度上的调控(transcriptional regulation);②mRNA加工成熟程度上的调控(differential processing of RNAtranscript);③翻译程度上的调控(differential translation of mRNA).原核生物中,营养状况(nutritionalstatus)和环境因素(environmental factor)对基因表达起着举足轻重的影响。
在真核生物尤其是高等真核生物中,激素程度(hormone level)和发育阶段(developmental stage)是基因表达调控的最主要手段,营养和环境因素的影响力大为下降。
二、基因表达调控的底子道理〔一〕基因表达的多级调控基因的布局活化、转录起始、转录后加工及转运、mRNA降解、翻译及翻译后加工及蛋白质降解等均为基因表达调控的控制点。
可见,基因表达调控是在多级程度长进行的复杂事件。
此中转录起始是基因表达的底子控制点。
四个底子的调控点:〔1〕基因布局的活化。
DNA表露碱基后RNA聚合酶才能有效结合。
活化状态的基因暗示为:1.对核酸酶敏感;2.结合有非组蛋白及修饰的组蛋白;3.低甲基化。
〔2〕转录起始。
最有效的调节环节,通过DNA元件与调控蛋白彼此作用来调控基因表达。
〔3〕转录后加工及转运。
RNA编纂、剪接、转运。
〔4〕翻译及翻译后加工。
翻译程度可通过特异的蛋白因子阻断mRNA 翻译翻译后对蛋白的加工、修饰也是底子调控环节。
(2021年整理)现代分子生物学(第4版)_课后思考题答案

答:1,装上5′端帽子;2,装上3′端多聚A尾巴;3,剪接:将mRNA前体上的居间顺序切除,再将被隔开的蛋白质编码区连接起来。剪接过程是由细胞核小分子RNA参与完成的,被切除的居间顺序形成套索形;4,修饰:mRNA分子内的某些部位常存在N6—甲基腺苷,它是由甲基化酶催化产生的,也是在转录后加工时修饰的。
(2)复合型转座子:复合型转座子是一类带有某些抗药性基因(或其他宿主基因)的转座子,其两翼往往是两个相同或高度同源的IS序列,表明IS序列插入到某个功能基因两端时就可能产生复合转座子.一旦形成复合转座子,IS序列就不能再单独移动,因为它们的功能被修饰了,只能作为复合体移动.大部分情况下,这些转座子的转座能力是由IS序列决定和调节的。 除了末端带有IS序列的复合转座子外,还存在一些没有IS序列的,体积庞大的转座子(5000bp以上)-—TnA家族。
意义:该模型揭示了DNA作为遗传物质的稳定性特征,最有价值的是确认了碱基配对原则,这是DNA复制、转录和反转录的分子基础,亦是遗传信息传递和表达的分子基础。该模型的提出是20世纪生命科学的重大突破之一,它奠定了生物化学和分子生物学乃至整个生命科学飞速发展的基石。
7.DNA复制通常采取哪些方式?
①线性DNA双链的复制
(2) DNA复制的引发:RNA引物的合成 前导链:DNA双链解开为单链后,由引发酶(RNA聚合酶, Primase)在5’ →3’DNA模板上合成一段RNA引物,再由DNA 聚合酶从RNA引物3’端开始合成新的DNA链.然后以此为起点,进入DNA复制的延伸。后随链:后随链的引发过程由引发体(Primosome)来完成。引发体由6种蛋白组成的引发前体(Preprimosome)和引发酶(Primase)组成.引发体催化生成滞后链的RNA引物短链, 再由DNA聚合酶III 作用合成后续DNA,直至遇到下一个引物或冈崎片段为止。在滞后链上所合成的RNA引物非常短,一般只有3-5个核苷酸。而且,在同一种生物体细胞中这些引物都具有相似的序列。
原核、真核生物基因及表达调控

原核、真核生物基因及表达调控引言现代生物学中“基因”一词甚为流行,细胞学、遗传学、生物化学等,以及各种生物学课本中,都涉及到“基因”一词。
甚至象典型的宏观生物学科——生态学,也把一片森林称为一个“基因库”[1]。
现代生物学已经完全证明,DNA 分子是由称为核普酸的有机分子线性聚合而成。
基因就是核普酸按一定顺序排列而成的DNA分子片段,它携带着遗传信息。
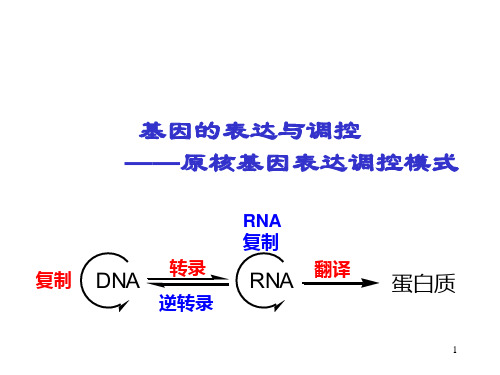
基因表达(gene expression)是指细胞在生命过程中,把储存在DNA顺序中遗传信息经过转录和翻译,转变成具有生物活性的蛋白质分子。
其实质就是遗传信息的转录和翻译。
在个体生长发育过程中,生物遗传信息的表达按一定的时序发生变化(时序调节),并随着内外环境的变化而不断加以修正(环境调控)[2]。
原核生物和真核生物的基因及表达过程有着差异。
随着世界分子生物学研究不断深入,基因表达技术有了很大的提高。
迄今为止,人们已经研究开发出多种原核和真核表达系统用以生产重组蛋白[3]。
一.原核、真核生物基因结构原核生物基因分为编码区与非编码区,所谓的编码区就是能转录为相应的信使RNA,进而指导蛋白质的合成,非编码区位于编码区的上游及下游。
[4]在调控遗传信息表达的核苷酸序列中最重要的是位于编码区上游的RNA聚合酶结合位点。
RNA聚合酶是催化DNA转录为RNA,能识别调控序列中的结合位点,并与其结合。
真核生物基因结构见图1:图1 真核生物基因结构二.原核、真核生物基因结构的区别最主要的在于真核基因是不连续的,而原核基因是连续的。
所谓真核基因的不连续,即一个基因的编码序列也叫外显子,被一个或多个非编码序列,又叫内含子所间隔。
[5]这些内含子和外显子同属一个转录单位,转录形成前体。
经过转录的加工,即切去内含子,重新连按外显子,从而得到成熟。
而绝大多数的原核基因是连续的,没有内含子的间隔,转录产生成熟。
不仅如此,而且凡在代谢途径上功能有关的多个基因可能紧密相联,与它们的调控基因一起组成一个操纵子,转录到一条链。
分子生物学 ch7原核生物基因表达调控

调节蛋白
由调节基因lacI编码,单顺反子,有自身弱启 动子,能独立地组成型表达 阻遏蛋白一个结合位点是诱导物结合位点, 可被小分子诱导物结合,改变其构型,从而 影响与操纵基因结合的活性 阻遏蛋白一个结合位点是操纵基因结合位点, 分 调节蛋白以四聚体形式与操纵基因Olac结合, 子 阻遏结构基因的表达 生
物 学
CAP(降解物活化蛋白)或CRP(环腺苷酸受体 蛋白)是分子量为22.5kd的二聚体,CRP单体具有 DNA结合区和转录激活区,二聚体被单个cAMP活化, cAMP-CAP复合物与启动子结合,促进基因表达
葡萄糖分解代谢降低cAMP水平,使得其他分解代
谢受阻
CAP
RNA聚合酶结合
-35 cAMP
——阻遏蛋白(repressor)的结合操纵序列 当操纵序列结合有阻遏蛋白时,会阻碍
RNA聚合酶与启动序列的结合,或是RNA聚合酶
不能沿DNA向前移动 ,阻碍转录。
pol 启动序列 操纵序列 编码序列 阻遏蛋白
激活蛋白(activator)可结合启动序列邻近的
DNA序列,促进RNA聚合酶与启动序列的结合,增
无效应物(辅阻遏物)——基因表达
操纵子分类
四类: 可诱导的正调控型:(ara O): 可阻遏的正调控型 可诱导的负调控型(lac O)、 可阻遏的负调控型(trp O)
有 效 应 物 * 基 因 表 达 无 效 应 物 * 基 因 表 达
调节蛋白结合-阻遏基因表达 (阻遏蛋白)
负调控
调节蛋白结合-基因表达 (激活蛋白)
酶和转乙酰酶,结构基因由位于上游的一个lac启动子(lacP)起始
转录;lac操纵基因(lacO)位于lacP启和lacZ之间,并且和lacP有 部分重叠,其上可结合位于上游具有独立转录单位的lac调节基因
分子生物学复习总结题-第七章-基因表达调控

第七章基因表达调控一、选择单选:1. 关于“基因表达”的概念叙述错误的是A. 其过程总是经历基因转录及翻译的过程B. 某些基因表达产物是蛋白质分子C. 某些基因表达经历基因转录及翻译等过程D. 某些基因表达产物是RNA分子E. 某些基因表达产物不是蛋白质分子2. 关于管家基因叙述错误的是A. 在生物个体的几乎各生长阶段持续表达B. 在生物个体的几乎所有细胞中持续表达C. 在生物个体全生命过程的几乎所有细胞中表达D. 在生物个体的某一生长阶段持续表达E. 在一个物种的几乎所有个体中持续表达3. 目前认为基因表达调控的主要环节是A. 翻译后加工B. 转录起始C. 翻译起始D. 转录后加工E. 基因活化4. 顺式作用元件是指A. 基因的5’、3’侧翼序列B. 具有转录调节功能的特异DNA序列C. 基因的5’侧翼序列D. 基因5’、3’侧翼序列以外的序列E. 基因的3’侧翼序列5. 一个操纵子(元)通常含有A. 数个启动序列和一个编码基因B. 一个启动序列和数个编码基因C. 一个启动序列和一个编码基因D. 两个启动序列和数个编码基因E. 数个启动序列和数个编码基因6. 反式作用因子是指A. 对自身基因具有激活功能的调节蛋白B. 对另一基因具有激活功能的调节蛋白C. 具有激活功能的调节蛋白D. 具有抑制功能的调节蛋白E. 对另一基因具有功能的调节蛋白7. 乳糖操纵子(元)的直接诱导剂是A. 葡萄糖B. 乳糖酶C. β一半乳糖苷酶D. 透酶E. 别乳糖8. Lac阻遏蛋白结合乳糖操纵子(元)的A. CAP结合位点B. O序列C. P序列D. Z基因E. I某因9. cAMP与CAP结合、CAP介导正性调节发生在A. 葡萄糖及cAMP浓度极高时B. 没有葡萄糖及cAMP较低时C. 没有葡萄糖及cAMP较高时D. 有葡萄糖及cAMP较低时E. 有葡萄糖及CAMP较高时10. Lac阻遏蛋白由A. Z基因编码B. Y基因编码C. A基因编码D. I互基因编码E. 以上都不是11. 色氨酸操纵子(元)调节过程涉及A. 转录水平调节B. 转录延长调节C. 转录激活调节D. 翻译水平调节E. 转录/翻译调节12.基因表达的产物不包括A.蛋白质B. mRNAC. rRNAD. SnRNAE. tRNA13.真核基因调控中最重要的环节是A. 基因重排B. 基因转录C. DNA的甲基化与去甲基化D. mRNA的衰减E. 翻译速度14.RNA聚合酶结合于操纵子的A. 结构基因起始区B. 阻遏物基因C. 诱导物D. 阻遏物E. 启动子15. cAMP对转录的调控作用是通过A. cAMP转变为CAPB. CAP转变为CampC. 形成cAMP-CAP复合物D. 葡萄糖分解活跃,使cAMP增加,促进乳糖利用来扩充能源E. cAMP是激素作用的第二信使,与转录无关16. 原核生物与DNA结合并阻止转录进行的蛋白质称为A. 正调控蛋白B. 阻遏物C. 诱导物D. 反式作用因子E. 分解代谢基因激活蛋白17.增强子A. 是特异性高的转录调控因子B. 是真核生物细胞内的组蛋白C. 原核生物的启动子在真核生物中就称为增强子D. 是增强启动子转录活性的DNA序列E. 是在结构基因的5'-端的DNA序列18.关于色氨酸操纵子的错误叙述是:A.trpR参与阻抑调控B.色氨酸阻抑结构基因转录C.前导序列参与色氨酸操纵子的衰减调控D.色氨酰tRNA参与色氨酸操纵子的衰减调控E.前导序列的序列3和序列4形成衰减子结构多选:1、基因表达调控环节包括A.DNA复制B.转录起始C.转录后加工D. mRNA降解E.翻译2、关于原核生物基因表达A.每个原核细胞的一切代谢活动都是为了适应环境而更好地生存和繁殖B.操纵子是原核生物绝大多数基因的表达单位C.原核生物基因表达的特异性由 因子决定D.原核生物基因表达既存在正调控,又存在负调控E.转录起始是原核生物基因表达主要的调控环节3、原核生物基因的调控序列包括A.启动子B.终止子C.操纵基因D.增强子E.衰减子4、原核生物基因的调控蛋白包括A.特异因子B.起始因子C.延长因子D.激活蛋白E.阻抑蛋白5、乳糖操纵子包含以下哪些结构?cZB. lacAC. lacOD. lacPE. lacI6、关于乳糖操纵子的错误叙述是:A.乳糖操纵子编码催化乳糖代谢的3种酶cI促进乳糖操纵子转录C.别乳糖促进乳糖操纵子转录D.CAP促进乳糖操纵子转录E.cAMP抑制CAP的激活效应7、色氨酸操纵子的结构A.含trpYB.含trpAC.含trpOD.含trpPE.含前导序列8、与RNA聚合酶活性调控有关的成分有A.tRNAB.核糖体C.严谨因子D.鸟苷五磷酸E.鸟苷四磷酸9、以下关于cAMP对原核基因转录的调控作用的叙述,正确的A. 葡萄糖与乳糖并存时,细菌优先利用乳糖B. cAMP-CAP复合物结合于启动子上游C. 葡萄糖充足时,cAMP水平不高D. cAMP可与CAP结合成复合物E. 葡萄糖和乳糖并存时,细菌优先利用葡萄糖10、原核生物基因表达在翻译水平上的调控与那些因素有关?A.mRNA前体后加工B. mRNA稳定性C. SD序列D.翻译阻抑E.反义RNA11、以下哪些环节存在真核生物的基因表达调控A.DNA和染色质水平B.转录水平C. 转录后加工水平D. 翻译水平E. 翻译后加工水平12、与原核生物相比,真核生物的基因表达调控的特点是A.转录的激活与转录区染色质结构的变化有关B.转录和翻译分隔进行,具有时空差别C.转录后加工更复杂D.既有瞬时调控又有发育调控E.转录调控以正调控为主13、在真核生物基因表达调控过程中,DNA水平的调控包括哪些内容A.染色质结构改变B. DNA甲基化C. 基因重排D. 基因扩增E.染色质丢失14、关于真核生物基因表达转录水平的调控A.转录水平的调控实际上是对RNA聚合酶活性的调控B.RNA聚合酶Ⅱ是转录调控的核心C.转录水平的调控主要通过RNA聚合酶、调控序列和调控蛋白的相互作用来实现D.真核生物的调控序列又称顺式作用元件E.真核生物基因表达的调控蛋白即转录因子,又称为反式作用因子15、真核生物的调控序列有哪些?A.启动子B.终止子C.增强子D.沉默子E.衰减子16、哪些属于真核生物基因表达的调控蛋白A.转录因子B.反式作用因子C.通用转录因子D. 反式激活因子E.共激活因子17、哪些是真核生物调控蛋白所含的DNA结合域A.螺旋-转角-螺旋B.锌指C.富含脯氨酸域D.亮氨酸拉链E.螺旋-环-螺旋。
分子生物学复习7-9

第七章基因的表达与调控(上)——原核基因表达调控模式(一)基本概念1.基因表达:细胞在生命过程中,把蕴藏在DNA中的遗传信息经过转录和翻译,转变成为蛋白质或功能RNA分子的过程称为基因表达。
2.基因表达调控:围绕基因表达过程中发生的各种各样的调节方式都统称为基因表达调控。
rRNA或tRNA的基因经转录和转录后加工产生成熟的rRNA或tRNA,也是rRNA或tRNA 的基因表达,因为rRNA或tRNA就具有在蛋白质翻译方面的功能。
3.组成型表达:指不大受环境变动而变化的一类基因表达。
如DNA聚合酶,RNA聚合酶等代谢过程中十分必需的酶或蛋白质的表达。
管家基因:某些基因在一个个体的几乎所有细胞中持续表达,通常被称为管家基因。
管家基因无论表达水平高低,较少受到环境因素的影响。
在基因表达研究中,常作为对照基因适应型表达:指环境的变化容易使其表达水平变动的一类基因表达。
应环境条件变化基因表达水平增高或从无到有的现象称为诱导,这类基因被称为可诱导的基因;相反,随环境条件变化而基因表达水平降低或变为不表达的现象称为阻遏,相应的基因被称为可阻遏的基因。
4.结构基因:编码蛋白质或功能性RNA的任何基因。
所编码的蛋白质主要是组成细胞和组织基本成分的结构蛋白、具有催化活性的酶和调节蛋白等。
原核生物的结构基因一般成簇排列,真核生物独立存在。
结构基因簇由单一启动子共同调控。
调节基因:参与其他基因表达调控的RNA或蛋白质的编码基因。
①调节基因编码的调节物质通过与DNA上的特定位点结合控制转录是调控的关键。
②调节物与DNA特定位点的相互作用能以正调控的方式(启动或增强基因表达活性调节靶基因,也能以负调控的方式(关闭或降低基因表达活性)调节靶基因。
操纵子:由操纵基因以及相邻的若干结构基因所组成的功能单位,其中结构基因的转录受操纵基因的控制。
(二)原核基因调控的分类和主要特点一、原核生物的基因调控特点:(1)基因调控主要发生在转录水平上,形式主要是操纵子调控.(2)有时也从DNA水平对基因表达进行调控,实质是基因重排。
分子生物学(朱玉贤第四版)复习提纲思维导图 7.原核生物基因表达调控
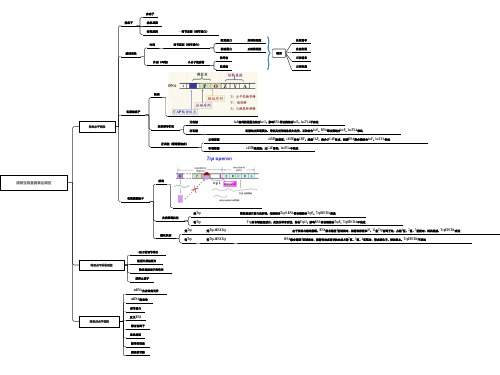
启动子 操纵子 操纵基因 结构基因 ←调节基因(调节蛋白) 阻遏蛋白 内因 基因表达 诱导物 外因(环境) 小分子效应物 阻遏物 正控阻遏 调节基因(调节蛋白) 激活蛋白 正转录调控 模型 正控诱导 负控阻遏 负转录调控 负控诱导
结构
乳糖操纵子
无乳糖 转录水平调控 负控诱导机制 有乳糖 无葡萄糖 正调控(葡萄糖效应) 有葡萄糖
2024版《现代分子生物学》朱玉贤第五版北大课件

新生肽链经过加工修饰,如剪切、 折叠、修饰等,成为具有生物活性 的蛋白质。
20
蛋白质翻译后加工修饰类型举例
2024/1/28
N-端fMet或Met的切除
新生肽链N-端的甲硫氨酸或甲酰甲硫氨酸通常被切 除。
二硫键的形成
半胱氨酸残基之间可以形成二硫键,对蛋白质的稳 定性和活性有重要作用。
化学修饰
生物工程
表观遗传学机制可以影响细胞的分化和发育,因此通过表观遗传学手段来改造细胞或生物体可能成为一种新 的生物工程技术。例如,利用表观遗传学手段来实现细胞重编程和再生医学应用。
26
06
现代分子生物学技术应用与 发展趋势
2024/1/28
27
DNA测序技术原理及应用领域拓展
DNA测序技术原理
通过特定的生物化学方法,将 DNA片段化并逐一测定其碱基序 列,从而获得完整的基因序列信
组修复等。
DNA损伤修复对于维持细胞基 因组稳定性和防止突变具有重要
意义。
2024/1/28
11
基因突变与遗传多样性
基因突变是指DNA序列中碱基的替换、 插入或缺失。
基因突变是生物进化的原材料,对于 生物适应环境和进化具有重要意义。
2024/1/28
基因突变可以产生新的等位基因,增 加遗传多样性。
序列比对与注释
01
利用生物信息学方法对基因序列进行比对和注释,揭示基因功
能和进化关系。
基因表达谱分析
02
通过高通量测序技术,研究基因在不同条件下的表达谱变化,
解析基因调控网络。
蛋白质结构与功能预测
03
利用生物信息学方法预测蛋白质的三维结构和功能,为药物设
计和蛋白质工程提供理论支持。
《现代分子生物学》教学大纲

《现代分子生物学》教学大纲课程名称:现代分子生物学课程类别:专业必修课学时:48 学时学分:3学分考核方式:考试适用专业:生物技术开课学期:第5或6学期一、课程性质、目的任务分子生物学是从分子水平研究生物大分子的结构与功能从而阐明生命现象本质的科学。
自20世纪50年代以来,分子生物学一直是生物学的前沿与生长点,其主要研究领域包括蛋白质体系、蛋白质-核酸体系和蛋白质-脂质体系。
生物大分子,特别是蛋白质和核酸结构功能的研究,是分子生物学的基础。
现代化学和物理学理论、技术和方法的应用推动了生物大分子结构功能的研究,从而出现了分子生物学的蓬勃发展。
本课程是研究核酸等生物大分子的功能、形态结构特征及其重要性和规律性的学科,也是生物专业的主干课程,分子生物学已成为生物类各专业教学计划中重要的核心课程,因此它是十分重要的一门必修课程,也是培养造就生物技术和生命科学高层次专门人才所需基本素质的重要课程。
本门课程的主要内容包括:染色体与DNA、基因和基因组、现代分子生物学的研究方法与技术、转录、翻译、原核生物基因表达与调控、真核生物基因表达调控、发育与分子调控等,此外,还包括各种讲座。
总之,通过分子生物学知识的传授,培养学生从分子水平上去分析、理解生命现象与过程,提高学生思考与探索生命奥秘的能力,从而为生物技术的分子生物学实验提供详实的理论基础。
二、课程基本要求该课程要求学生掌握现代分子生物学基本理论和基本技术,为其它专业课的学习和今后的发展奠定基础。
在课程学习的同时,要求学生提高思想道德修养、自学能力、专业英语能力、应用知识能力、表达能力、创新能力和科研能力。
三、学时分配四、教学方法与考核(一) 教学方法1.以学科体系为主体,以应用为目的,教学过程加强针对性和实用性。
2.本课程以讲授为主、自学和讨论为辅的方式组织教学,并通过阅读主要参考书目、网上查询、资料整理和专题讨论,加深对细胞生物学了解,并掌握该学科的实验技能和操作。
现代分子生物学简答期末考试重点

1.核小体结构?2.核糖体活性位点?3.DNA二级结构?4.维持DNA双螺旋稳定性因素?5.原核生物中的DNA聚合酶(大肠杆菌)6.真核生物的RNA聚合酶Ⅱ的启动子结构特点?7.转座发生的机制、类型、遗传学效应8.证明遗传物质是核酸的实验依据是什么?9.设计实验证明DNA的半保留复制?10有何机制确保DNA复制的忠实性?11.原核生物(以大肠杆菌为例)DNA复制起始的步骤?12.简述原核生物转录起始的过程?13.大肠杆菌有两种类型的终止子(原核生物转录终止的两种机制)14.比较原核真核转录的差异?15.真核生物转录后加工?16.原核生物翻译起始过程?17.真核生物翻译后加工18.细菌与真核生物RNA翻译的机理的异同19.延伸因子的种类及作用机制?20.蛋白质合成的延伸步骤有21.蛋白质后加工22.简述真核生物核基因mRNA剪接的机制?23.真核生物基因表达调控的主要控制点有哪些?24.原核生物的基因表达调控分为几个层次25.真核生物基因表达调控的层次与方式26.真核生物和原核生物在基因表达调控上有以下几点不同27.什么是原核生物的正调控和负调控28.正调控和负调控的主要不同29.大肠杆菌链前导链和滞后链的协同合成30.启动子的作用是什么,原核生物启动子的结构特征31.TBP在三种真核RNA聚合酶的转录起始中的机制32.为什么说RNA编辑是中心法则的例外33.为什么说σ 因子的更替可对转录进行调控?34.Trapoxin是组蛋白去乙酰化酶的抑制剂。
你认为该抑制剂对转录的影响是什么,为什么?35.可变剪接调控机制?36.反式作用因子与顺式作用元件的相互作用存在于基因表达的各个水平上。
请分别举例说明:⑴DNA复制起始⑵转录起始和⑶翻译起始过程中二者的相互作用。
37.真核生物反式作用因子的功能域及其与DNA结合基序有哪些?38.真核生物的顺式作用因子和反式作用因子如何相互作用来调控基因的转录39.弱化子的作用机理?40.以色氨酸操纵子为例,论述原核生物基因表达的阻抑作用和弱化作用的机制(见上题)41.以大肠杆菌为例,说明色氨酸操纵子的特点和机制及乳糖操纵子的机制42.葡萄糖代谢是如何调控乳糖操纵子表达的?43.DNA的复制过程1.核小体的结构?①由核心颗粒和连接区构成;②核心颗粒包括由8个组蛋白分子(H2A,H2B,H3,H4各两个)构成的组蛋白核心和包绕在核心表面的DNA分子;③包绕在组蛋白核心表面的DNA长140bp,环绕1 ¾圈;④连接区由DNA分子和H1组蛋白分子构成,长度不定;2.核糖体活性位点?①mRNA结合位点:结合mRNA和IF因子②P位点:结合fMet-tRNA和肽基-tRNA③A位点:结合氨酰基-tRNA④E位点:结合脱酰tRNA⑤肽酰基转移酶:将肽链转移到氨基酰-tRNA上⑥EF-Tu结合位点:氨基酰-tRNA的进入⑦EF-G结合位点:移位3.DNA的二级结构?1953年,Watson和Crick提出DNA的反向平行右手双螺旋结构模型。
分子生物学课程教学大纲【范本模板】

《分子生物学》课程教学大纲课程中文名称:分子生物学课程英文名称:Molecular Biology课程类别:必修课课程编号: 0901100003课程归属单位:农学院修定时间:2007年7月一、课程的性质、任务1. 课程的基本类型、基本属性分子生物学是目前自然科学中进展最迅速,最具有活力和生气的领域,也将是21世纪的带头学科。
它是研究核酸等生物大分子的功能、形态结构特征及其重要性和规律性的科学,分子生物学已成为农学类有关专业教学计划中重要的核心课程,因此它是十分重要的一门必修基础课程,是培养造就农学类高层次专门人才所需基本素质的重要课程。
2。
教学的基本要求通过对本课程的学习,使学生从分子水平上认识、理解生命现象及其过程,培养学生思考与探索生命奥秘的能力.为学生的分子生物学实验提供强实的理论基础.本课程要求学生掌握现代分子生物学的基本理论和基本技术,为其它专业课的学习和今后的发展奠定基础。
3. 适用专业与学时数本教学大纲适用于农学、植保、园艺、农产品质量与安全、中草药栽培与鉴定等专业大学本科生。
总学时数为54学时,其中理论部分36个学时,实验部分18个学时。
4. 本门课程与其他课程关系本门课程与其他课程关系较为密切,以微生物学、生物化学、遗传学等基础课为理论支撑,故学生在学习本课程之前应具备以上课程的基础知识。
5。
推荐教材及参考书推荐教材:卢向阳主编,《分子生物学》,中国农业出版社,2004 。
参考书:(1)、阎隆飞、张玉麟主编,《分子生物学》,中国农业大学出版社,1997。
(2)、朱玉贤等编著,《现代分子生物学》,高等教育出版社,1995(3)、张玉静主编,《分子遗传学》,科学出版社,20006.主要教学方法与媒体要求:本课程教学所采取的主要教学方法有课堂讲授、室内实验.课堂讲授主要采用板书教学结合多媒体教学手段进行。
二、各章教学内容和要求绪论(2学时)基本要求:使学生了解分子生物学的含义,研究内容、范畴,以及分子生物学的发展、现状和展望.基本知识点:分子生物学的含义、研究内容,分子生物学与农业科学的关系。
分子生物学原核生物基因表达调控ppt课件
一、原核基因表达调控环节
1、转录水平上的调控
(transcriptional regulation)
2、转录后水平上的调控
(post-transcriptional regulation)
① mRNA加工成熟水平上的调控 ② 翻译水平上的调控
15
二、操纵子学说
1、操纵子模型的提出 1961年,Monod和Jacob提出 获1965年诺贝尔生理学和医学奖
54
55
③ 操纵基因是DNA上的一小段序列(仅为26bp), 是阻遏物的结合位点。
56
RNA聚合酶结合部位
阻遏物结合部位
57
操纵位点的回文序列
58
④当阻遏物与操纵基因结合时,lac mRNA的转 录起始受到抑制。
59
未诱导:结构基因被阻遏
阻遏物 四聚体
LacI P O
lacZ
lacY
lacA
32
酶合成的诱导操纵子模型
调节基因
操纵基因
结构基因
阻遏蛋白
调节基因
操纵基因
结构基因
诱导物
如果某种物质能够促使
阻遏蛋白
mRNA
细菌产生酶来分解它,
这种物质就是诱导物。
诱导物
酶蛋白
33
• 可阻遏调节:基因平时是开启的,处在产生蛋白质 或酶的工作过程中,由于一些特殊代谢物或化合物 的积累而将其关闭,阻遏了基因的表达。 例:色氨酸操纵子 合成代谢蛋白的基因
1、根据操纵子对调节蛋白(阻遏蛋白或激活蛋白) 的应答,可分为: 正转录调控 负转录调控
29
调节基因
操纵基因
结构基因
激活蛋白 阻遏蛋白
正转录调控 负转录调控
原核生物基因表达调控的方式

原核生物基因表达调控的方式
1.DNA、染色体水平调控:基因丢失、基因修饰、基因重排、基因扩增、染色体结构变化。
2.转录水平调控(主要调控方式):转录起始、延伸、终止均有影响。
原核生物借助于操纵子,真核生物通过顺式作用元件和反式作用因子相互作用进行调控。
3.转录后水平调控:主要指真核生物原初转录产物经过加工成为成熟的mRNA,包括加帽、加尾、甲基化修饰等。
4.翻译水平调控:对mRNA稳定性的调控、反义RNA对翻译水平的调控等。
5.翻译后水平调控:蛋白质的剪切、化学修饰(磷酸化、乙酰化、糖基化等)、转运等。
6.mRNA降解的调控。
分子生物学第七章原核生物基因表达调控

(三)、阻遏物 lac I 基因产物及功能
Lac 操纵子阻遏物 mRNA 是由弱启动子控制下组 成型合成的,该阻遏蛋白具有4个相同的亚基,每个亚 基均含347个氨基酸残基。
lacI 基因为组成型,通过启动子的上升突变体可获 得较多的阻遏蛋白;
阻遏物 2022/10/18
β-半乳糖苷酶 透过酶 转乙酰3酶2
2022/10/18
16
调节机理:
细胞中某一氨基酸或嘧啶的浓度发生改变
氨酰 – tRNA的浓度变化
核糖体在转录产物RNA上的结合位置不 同,使得RNA形成特定的二级结构 由RNA的二级结构判断基因能否继续转录
2022/10/18
17
3、降解物对基因活性的调节P252
葡萄糖效应或降解物抑制作用:细菌培养基中在 葡萄糖存在的情况下,即使加入乳糖、半乳糖等 诱导物,与其对应的操纵子也不会启动,这种现 象称为葡萄糖效应或降解物抑制作用。
这是通过阻止乳糖操纵子表达来完成的,这种 效应称为降解物抑制(catabolite repression)。
2022/10/18
35
(五)、cAMP与代谢物激活蛋白
葡萄糖
葡萄糖-6-磷酸
甘油 某些代谢产物抑制活性
腺苷酸环化酶
ATP
cAMP
编码
cAMP-CAP
Crp基因
代谢物激活蛋白 CAP
葡萄糖对其它糖的代谢抑制,是通过对 cAMP的抑制完成的。
2022/10/18
22
一、酶的诱导 ——
lac 体系受调控的证据
两种含硫的乳糖类似物:
异丙基巯基半乳糖苷
(IPTG)
巯甲基半乳糖苷(TMG)
E. coli 在不含乳糖的培养基生 长时,β-半乳糖苷酶含量极低;
分子生物学基础第七章真核基因表达的调控第三节真核基因表达转录水平的调控

第七章 真核基因表达的调控
第三节 真核基因表达转录水平的调控
一、真核基因转录与染色质结构变化的关系 DNA绝大部分都在细胞核内与组蛋白等结合成染色质, 染色质的结构影响转录,至少有以下现象: 1.染色质结构影响基因转录 在真核细胞中以核小体为基本单位的染色质是真核基 因组DNA的主要存在方式。DNA盘绕组蛋白核心形成核小体, 妨碍了与转录因子及RNA聚合酶的靠近和结合,使基因的 活性受到抑制。 2.组蛋白的作用 组蛋白H1及核心组蛋白共同参与核小体的组装与凝聚。 在特殊氨基酸残基上的乙酰化、甲基化或磷酸化等修饰, 可改变蛋白质分子表面的电荷,影响核小体的结构,从而 调节基因的活性。
第三节 真核基因表达转录水平的调控
图7-6 碱性螺旋-环-螺旋结构图
第三节 真核基因表达转录水平的调控
螺旋-转角-螺旋结构域是最早发现于原核生物中的一个关键因子, 该结构域长约20个aa,主要是两个α-螺旋区和将其隔开的β转角。 其中的一个被称为识别螺旋区,因为它常常带有数个直接与DNA序列 相识别的氨基酸。其结构如图7-3所示。
图7-3 螺旋-转角-螺旋结构及其与 DNA的结合
第三节 真核基因表达转录水平的调控
2.增强子 增强子是指能使基因转录频率明显增加的DNA序列。增强子的作 用有以下特点。 ①增强效应十分明显。一般能使基因转录频率增加10~200倍,有 的可以增加上千倍, ②增强效应与其位置和取向无关。 ③大多为重复序列。 ④增强效应有严密的组织和细胞特异性。说明只有特定的蛋白质 (转录因子)参与才能发挥其功能。 ⑤没有基因专一性,可以在不同的基因组合上表现增强效应。 ⑥许多增强子还受外部信号的调控,如金属硫蛋白的基因启动区 上游所带的增强子,就可以对环境中的锌、镉浓度做出反应。 ⑦增强子要有启动子才能控
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
诱导物 和阻遏
蛋白结 合的模
型
维持阻遏
阻遏物结合在 操纵基因上 aaaa aaa a a 诱导物 a
过量的阻遏物可结
合在 DNA 的其它位点
诱导
所有的阻遏物都结合
aa
在 DNA 的随机位点上
a
a
a
aa 诱导物解离
a 建立阻遏
阻遏物恢复
活性状态
a
a
a
阻遏蛋白通过直接取代
a
从随机位点移向
a
操纵基因
a
a
在细菌中,cAMP与CAP(catabolite activator protein,CAP)二聚体结合形成二 元复合物共同发挥作用。只有cAMP存在时, CAP才有活性。
CAP是一个正调控因子,在依赖CAP的启动 子上起始转录必须有CAP参与。cAMP下降, CAP就不能与控制区结合,RNA聚合酶就 不能启动转录。
乳糖操纵子的阻抑 蛋白为两个二聚体 结合在一起形成的 四聚体结构。四个 亚基的螺旋连在一 起,维持四聚体结 构。
每个二聚体都 有两个头段, 两个头段在一 起结合一个操 纵基因序列。 这使整个阻抑 蛋白能够同时 结合操纵基因 的两个位点。
完整的四聚 体阻抑蛋白 能够同时与 操纵基因结 合,亲和力 较不完整的 阻抑蛋白高 很多。
诱导作用和阻抑作用都能对细菌的代谢能力 进行调控。前者调节细菌使用某种物质的能 力,后者调节合成特定代谢中间产物的能力。 每种调节的诱发剂都是小分子物质,或为酶 的底物,或为酶所催化合成的产物。
诱导物或辅阻抑物的作用是高度特异的,仅 有特定的或密切相关的分子能发挥作用。但 这些小分子并不直接与其调控的目标酶分子 相互作用而实现其调控功能。
导 阻遏物 阻遏
阻
诱导物 诱导
失活的阻遏物 Trp O
失活的活性蛋白 阻遏
活化的激活蛋白 诱导物
诱导
未发现
遏 失活的活性蛋白
诱导
辅-阻遏物 阻遏
活化的激活蛋白 诱导
辅-阻遏物 失活的活性蛋白 阻遏
图 16-1 原核生物结构基因的 4 种表达调控类型 (仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.21)
4.操纵基因和阻抑蛋白
操纵基因(operator)是与启动子相邻的顺式 作用位点,是阻抑蛋白的靶点。阻抑蛋白 (repressor)是调节基因的产物,与操纵基 因的结合可以阻止受调节基因的表达。
当阻抑蛋白与操纵基因结合时,就会阻止 RNA聚合酶启动转录,基因的表达就被关闭。 在无阻抑蛋白时,RNA聚合酶可以识别受调 节基因的启动子,使这种基因得到表达。
二、乳糖操纵子
1 乳糖操纵子与负控诱导系统
乳糖操纵子(lactose operon,lac )的三个结 构基因成簇排列,编码参与β-半乳糖苷(如 乳糖)分解代谢所需的三种蛋白质:lacZ编 码β-半乳糖苷酶,lacY编码β-半乳糖苷透性酶, lacA编码β-半乳糖苷转乙酰基酶。
lacI基因(调节基因)正好与结构基因相邻, 但它不与结构基因属于同一转录单位,它 有自己独立的转录单位,含有自己的启动 子和终止子。
C端为两组亮氨酸拉链,形成四聚体。
Helix-turn-helix Core domain1 Core domain2
1
51
80
360
DNA binding Hinge
Inducer binding Oligomerization
图 16- 阻遏蛋白单体的结构和功能
2. 阻抑蛋白结构与 功能的关系
控制位点(controlling site) 诱导(induction)
第二节 转录水平的调控
原核生物的基因调控可以发生在转录和翻 译等不同阶段,但也是以转录水平为主。
原核生物许多功能相关的结构基因,特别 是编码同一代谢途径的酶的基因,一般成 簇排列,能受单一启动子的共同控制,结 果是整簇基因或者都表达或者都不表达。
genes.
Mutations that inactivate the lacI gene operon to be constitutively expressed,because the mutant repressor protein cannot bind to the operator.
反式作用因 子通常为蛋 白质或RNA, 其特征为可 以从合成地 扩散到目标 场所发挥作 用。
顺式作用元件(cis-acting factor)是指对基因表 达有调节活性的DNA序列,其活性只影响与其自 身同处在一个DNA分子上的基因。顺式作用元件 通常不编码蛋白质,多位于基因旁侧或内含子中。
乳糖操纵子同样存在正调控。
在没有诱导物存在的情况下,存在lac操纵 子的本底水平的表达。
2 cAMP对乳糖操纵子转录的激活作用
细菌在富含葡萄糖的培养基上生长的时候, 葡萄糖可以抑制β-半乳糖苷酶表达的一个很 重要的因素是葡萄糖降低了细菌体内cAMP 的水平。
生化和遗传学实验证明,cAMP可以结合到 启动子的某个部位而激活操纵子的转录。
加入诱导物以后,释放出阻抑蛋白,被关闭的 复合物转变为开放的复合物,起始转录。
(a)
(b)
诱导 物的
结合
解离 模型
图 16-10 诱导物作用模型。(a)平衡模型,诱导物与游离的阻遏物结合,打破了其游离与 结合之间的平衡;(b)解离模型,诱导物直接与结合在操纵位点上的阻遏物结合,使其释放 (仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.13)
a
a
a
a 图 16-14 在细胞中所有的阻遏蛋白都结合在 DNA 上 (仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.18)
gal aroH trp trpR lac
Startpoint
Promoter Operator location
图 16-13 操纵基因可位于启动子不同的相对位点
第七章 原核生物基因表达调控
机体可以在基因 表达过程(DNA 到蛋白质)的任 何阶段进行调控, 一般以转录水平 上的调控为主。
第一节 基因表达调控的基本概念
1. 反式作用因子与顺式作用元件
基因表达的产物(蛋白质或RNA)从合成的场 所扩散到目标场所而发挥作用的过程称为 反式作用(trans-acting),此基因表达产物 被称为反式作用因子(trans-acting factor) 。
lacI编码可扩散的产物,理论上它不必位于 结构基因附近。将lacI基因转移到其他任何 地方都能很好地发挥作用,因此lacI的表达 产物属于反式作用因子。
lacI基因的表达产物称为乳糖操纵子阻抑蛋 白(lac repressor)。它的功能是阻止结构 基因的表达。因此,乳糖操纵子的调控属 于负调控。
顺式作用(cis-acting)的概念用于任一不转变为 任何其他形式的DNA序列,它只在原位发挥 DNA序列的作用,仅影响与其物理上相连的 DNA序列的活性。
基因活性的调控主要通过反式作用因子(通常是 蛋白质)和顺式作用元件(通常在DNA上)相互 作用而实现。
2. 结构基因和调节基因
结构基因(structural gene)是编码蛋白质或 RNA的任何基因。结构基因编码大量的功 能各异的蛋白质,如组成细胞和组织的结 构蛋白和酶等。
正调控和负调控
正调控 负调控 诱导(induction) 阻遏(repression) 诱导物(inductor) 阻遏物(repressor) 辅阻遏物(corepressor) 使阻遏蛋白具有活性
或使活性蛋白失去活性的物质称~。
负调控
Lac O
诱
正调控 Ara O
与诱导物结合可直 接引起阻抑蛋白构 象发生改变,使两 个二聚体的头段不 再同时与DNA结合, 使多亚基阻抑蛋白 失去优势,降低了 与操纵基因结合的 亲和力。
3. 阻抑蛋白对RNA聚合酶功能的影响
阻抑蛋白和RNA聚合酶可同时与DNA结合, 而且阻抑蛋白与DNA的结合能够增强RNA聚 合酶结合DNA的能力,但是结合着的RNA聚 合酶不能起始转录。
原核生物操纵子中的全部结构基因从同一个ቤተ መጻሕፍቲ ባይዱ启动子开始转录成单个mRNA分子。
一、诱导作用和阻抑作用
细菌应答某种特定物质出现而合成特定酶 的过程,称为诱导作用(induction)。大肠 杆菌乳糖操纵子是这种机制最好的范例。
如果某种小分子物质能够促使细菌产生酶 将其自身分解,这种小分子物质就叫做诱 导物(inducer)。
CAP激活转录有两种方式:
阻抑蛋白的作用机制
1. 阻抑蛋白及其靶DNA序列的对称性 阻抑蛋白识别位点的共有特点就是具有对
称性。靠近对称轴的一对反向重复序列在 阻抑蛋白结合过程中起主要作用。阻抑蛋 白的结构也具有对称性。
阻遏蛋白的结构:
N端1~59aa-头部片段:为HTH,与操纵 基因DNA的大沟结合;
核心区:有6个折叠,诱导物就结合在两 个核心区之间的裂缝中;
调节基因(regulator gene)仅指参与其他基 因表达调控的RNA和蛋白质的编码基因。 调节基因编码的调节物通过与DNA上的特 定位点结合而控制受调节基因的转录是基 因表达调控的关键。
3. 启动子和终止子
位于转录单位开始和结束位置上的DNA序 列,称为启动子(promoter)和终止子 (terminator)。两者都是典型的顺式作用 元件,能受同一类反式作用因子RNA聚合 酶的识别。
异丙基硫代-β-D-糖苷(IPTG)虽然不能被 β-半乳糖苷酶分解,但它是乳糖操纵子非常 有效的诱导物。
这种能诱导酶合成,但不能被酶分解的分 子称为义务诱导物(gratutious inducer)。 义务诱导物由于能在细胞内保持原型不变, 因此很有用处,而真正的诱导物会被代谢 分解掉。