第五章、噬菌体和粘粒载体
12第五章 噬菌体载体

234页
噬菌体:细菌病毒。 烈性噬菌体:感染寄主细胞后能进行复制、增殖,最后 引起细胞裂解的噬菌体。只有溶菌生长周期的噬菌体。 温和性噬菌体(溶原性噬菌体):感染细菌后能使细菌 产生溶原性的噬菌体。既能进入溶菌生命周期又能进入 溶原生命周期的噬菌体。 溶原性:温和噬菌体感染细胞形成原噬菌体的过程。 原噬菌体:在溶原性细菌内存在的整合的或非整合的噬 菌体DNA。 溶原性细菌:细胞中含有以原噬菌体状态存在着的温和 噬菌体的细菌。 噬菌斑:在含细菌的固体培养基上,噬菌体使细菌细胞 裂解而形成的空斑。
(243页)
4. 噬菌体的溶源生命周期
Ⅱ、溶源周期的主要特征(以λ噬菌体为原型): (1)噬菌体的DNA分子注入细菌细胞。 (2)经过短暂的转录。 (3)噬菌体的DNA分子插入到细菌染色体基因组 DNA上,变成原噬菌体。 (4)细菌继续生长、增殖,噬菌体的基因作为细 菌染色体的一部分进行复制。
(244页)
基因工程又称重组DNA技术 其实施至少要有四个必要的条件 1. 工具酶 2. 基因 3. 载体 4. 受体细胞
(《基因工程..》静国忠,85页)
(240页)
噬菌体定义: 是一类细菌病毒的总称。 噬菌体与质粒的区别 * 质粒:仅是一种含有复制起点的裸露的DNA分 子。 * 噬菌体:DNA分子除了复制起点之外,还有编 码外壳蛋白质的基因。
Ⅰ、若干基本概念
234页
噬菌体:细菌病毒。 烈性噬菌体:感染寄主细胞后能进行复制、增殖,最后 引起细胞裂解的噬菌体。只有溶菌生长周期的噬菌体。 温和性噬菌体(溶原性噬菌体):感染细菌后能使细菌 产生溶原性的噬菌体。既能进入溶菌生命周期又能进入 溶原生命周期的噬菌体。 溶原性:温和噬菌体感染细胞形成原噬菌体的过程。 原噬菌体:在溶原性细菌内存在的整合的或非整合的噬 菌体DNA。 溶原性细菌:细胞中含有以原噬菌体状态存在着的温和 噬菌体的细菌。 噬菌斑:在含细菌的固体培养基上,噬菌体使细菌细胞 裂解而形成的空斑。
第八讲单链噬菌体载体及噬菌粒载体

第八讲单链噬菌体载体及噬菌粒载体吴乃虎中国科学院遗传与发育生物学研究所第八讲单链噬菌体载体及噬菌粒载体一、单链噬菌体的一般生物学1.单链噬菌体的优越性2.M13噬菌体的生物学特性二、M13克隆体系1.M13克隆体系2.M13克隆体系-半乳糖苷酶的显色反应原理3.M13载体系列的发展4.M13载体系列的优点三、噬菌体展示载体1.噬菌体展示载体的构建原理2.噬菌体展示载体3.噬菌体表面展示文库4.应用噬菌体展示载体分离有关蛋白质的实例四、噬菌粒载体1.M13噬菌体载体克隆的若干难点2.噬菌粒3.若干常用的噬菌粒载体4.pBluescript噬菌粒载体5.pUC118和pUC119噬菌粒载体第八讲单链噬菌体载体一、单链噬菌体一般生物学大肠杆菌丝状单链DNA噬菌体有M13噬菌体、f1噬菌体及fd 噬菌体,它们均含有分子量约为6400个核苷酸的单链闭环DNA分子。
1.单链DNA phage的优越性A.具有双链的复制型DNA(RF DNA),可如质粒质粒一样进行遗传操作;RF DNA:Replication Form DNA。
B.RF DNA和ssDNA均可感染感受态的寄主细胞——形成phaque或colony。
C.不受包装的限制。
因为单链DNA phage的大小是受其DNA 多寡制约的。
D.可容易地测出外源DNA的插入取向。
E.可产生大量的含有外源DNA的单链DNA分子,这种单链DNA分子有如下用途(作为模板):*1用作双脱氧链终止法进行DNA测序*2制备单链的放射性标记的杂交用DNA探针*3利用寡核苷酸进行定点突变2.M13 phage的生物学特性A.M13 phage同f1 phage亲缘关系十分密切,例如:①基因组组织形式相同;②病毒颗粒大小、形状相近;③DNA同源性高达98%以上。
B.在M13 phage颗粒中只有(+)链DNA,感染具F性须的大肠杆菌菌株,因此M13噬菌体是雄性E.coli特有的;M13噬菌体的(+)链DNA,又称为感染性单链DNA。
第五章DNA克隆用载体

三、质粒的构建
(一) 质粒人工构建的方法 方法就是在天然质粒的基础上进行
DNA重组,拼拼接接,以改变其性 能。
(二) 质粒人工构建的目的
(1)加入合适的选择标记基因,如两个 以上,易于用作选择;
(2)增加或减少合适的酶切位点,便于 重组;
(3)缩短长度,切去不必要的片段,提 高导入效率,增加装载量;
质粒pBR322 DNA
pBR322 质粒图谱
多克隆位点
氨苄青霉素 抗性基因
四环素抗 性基因
pUC系列的质粒载体
pUC系列的质粒载体集中了当时载体的诸多优点。 包括4个部分: ①来自pBR322的复制起点(ori); ②氨苄青霉素抗性基因(ampr); ③大肠杆菌乳糖操纵子的调节基因(laci)启动子 (Plac)和β 半乳糖苷酶的α -肽(lacZ); ④位于lacZ基因中靠近5′端的一段多接头,或称为 多克隆位点(MCS)。
因此通过蓝白斑筛选可 以筛选、鉴定重组体与非重 组体载体。
pGEX-6P-1 质粒物理图谱
SV40, 猴空泡病毒40
巨细胞病毒 (CMV)
(二)不相容性
利用同一复制系统的不同质粒(RNAI RNAII Rop因子)如果被导入同一细胞 中,它们在复制及随后分配到子细胞的过 程中,就会彼此竞争,它们在单细胞中的 拷贝数也会有差异,拷贝多的复制更快, 结果在细菌繁殖几代之后,细菌的子细胞 中绝大多数都含有占优势的质粒,因而这 两种质粒中只能有一种长期稳定地留在细 胞中,这就是所谓的质粒不相容性。
MAC (Mammalian Artificial Chromosome)
病毒载体
E.coli E.coli 酵母细胞 哺乳类细胞 动物细胞
环状 环状 线性染色体 线性染色体 环状
基因工程生物化学

逆转录酶为依赖 RNA的 DNA聚合酶,具有 5’ 3’
2
聚合酶活性外,以 RNA为模板合成 DNA。
3
逆转录酶还具有核糖核酸酶H (RNase H) 活性,可
4
从 5’ 末端或 3’ 末端降解RNA:DNA杂合链中的
5
RNA。
6
依赖 DNA的 DNA聚合酶
7
逆转录酶无 3’ 5’ 外切酶活性。
8
禽类逆转录酶和鼠类逆转录酶的比较
以 mRNA为模板合成第一条 cDNA链,所用的引物包括 Oligo (dT)12-18 特定序列的寡核苷物 克隆特定的 cDNA:用特定序列的寡核苷酸引物 RT-PCR:用特定序列的寡核苷酸引物 制备 cDNA 探针:用特定序列的寡核苷酸引物或 随机序列的寡核苷酸引物
A
C
C
C
5'
DNA连接酶
基因工程 (Genetic engineering)
(Central dogma)
基因 (Gene) 能编码一条多肽链并具有一定长度的 DNA 分子。 基因组 (Genome) 一种生物全部染色体的遗传物质的总和,以全长 DNA 的碱基对 (bp) 数目表示。
5’ 凸端 (5’ overhang) 粘性末端
3’ 凸端 (3’ overhang) 粘性末端
CCCGGG GGGCCC
5’ - 3’ -
-3’ -5’
5’ - 3’ -
- OH 3’ - P 5’
CCC GGG
GGG CCC
5’ P - 3’ OH-
-3’ -5’
SmaI
-3’ -5’
细菌基因组 大肠杆菌: 4 x 106 bp
人类基因组:3 x 109 bp
入噬菌体及质粒载体

• ③质粒对宿主生存并不是必需的,某些质粒携带的基因
功能有利于宿主细胞的特定条件下生存。
质粒载体的种类
• 在基因工程中,常用人工构建的质粒作为 载体。人工构建的质粒可以集多种有用的 特征于一体,如含多种单一酶切位点、抗 生素耐药性等。常用的人工质粒运载体有 pBR322、pSC101。pBR322含有抗四环素 基因(Tcr)和抗氨苄青霉素基因(Apr),并含 有27种限制性内切酶的单一识别位点。
λ- 噬菌体及质粒载体
——郭苋 2014212346
噬菌体的一般生物学特性
• 依赖于寄主细胞存活。脱离了寄主细胞后既不能 生长也不能复制。
• 基本功能: • 保护自己的核酸分子(DNA或RNA)免遭环境化
学物质的破坏 • 将其核酸分子注入到被感染的细菌细胞 • 经被感染的细菌细胞转变成制造噬菌体的体系,
质粒载体的优缺点
• 质粒载体主要用于DNA分子的亚克隆,表达外源 蛋白,DNA序列的测定和基因的体外重新构建等。
• 1.与外源DNA重组操作简便; • 2.外源DNA在质粒载体中相对较稳定, • 3.质粒DNA的制备和纯化方便,快速简单,所以
得到广泛应用。
• 缺点: • 1.它容纳外源DNA片段有限,通常最多为15kb左
酶形成融合蛋白质的克隆载体。
质粒载体
质粒特点
• ①染色质外的双链共价闭合环形DNA(covalently closed
circuar DNA,cccDNA)可自然形成超螺旋结构,不同质 粒大小在2-300kb之间,<15kb的小质粒比较容易分离纯
化,>15kb的大质粒则不易提取。
• ②能自主复制,是能独立复制的复制子(autonomous
基因工程第五章-载体

• 最后:4℃ 12000rpm离心10分钟,将上清 转移到另一离心管中,上清中是质粒DNA 。 加等体积氯仿,振荡混匀,
4℃12000rpm离心10分种,将上清转移到 另一离心管中。用2倍体积的无水乙醇沉淀 双链DNA。温和振荡混匀,于-18℃放置20 分钟。用微量离心机于4℃以12000rpm离 心10分钟。弃上清,加入70%冷乙醇洗沉 202淀0/3/310 -2次真空干燥DNA沉淀后,将其溶于TE
• 共价闭合超螺旋形式:(covalently closed circular,简写 cccDNA)
• 开放环结构:一条保持着完整的环结构,另一条有一个或数个 切刻。
• 线性结构:质粒在限制性酶作用下,双链断裂,形成线性,又 叫L型。 克隆只有超螺旋可用。
2020/3/30
2. cccDNA分子分离特性:
2020/3/30
2、pUC质粒载体:
有一个或多个利于检测的遗传表型,易于识别和筛选。如抗 药性、显色表型、营养缺陷型、荧光表型等. 有一个或几个限制性酶切酶的单一识别位点,便于外源基因 的插入。
适当的拷贝数。高拷贝数不仅利于载体的制备,也有利于克 隆基因的剂量的增加。
载体的分类:质粒载体――可容纳15KB的外源片断 噬菌体载体――25-45KB 人工染色体--45KB以上
• 由于cccDNA分子团压成一个紧密 的结构,当我们用EBr、丫啶橙染 料分子作用后,线性分子染料插入 的分子个数多,cccDNA插入的分 子个数少,密度大,所以沉降系数 大,进行密度梯度离心时可将其分 开
• cccDNA在较高高的温度和PH值条件下才会变性,(较线 性和开放环结Байду номын сангаас的DNA),如果变性,由于团在一起,条 件恢复后,会很快复性,形成cccDNA,线性和环状的变 性后,复性会形成杂乱的大分子团状结构,从而从体系中 沉淀下来。
大学《基因工程学》教学大纲
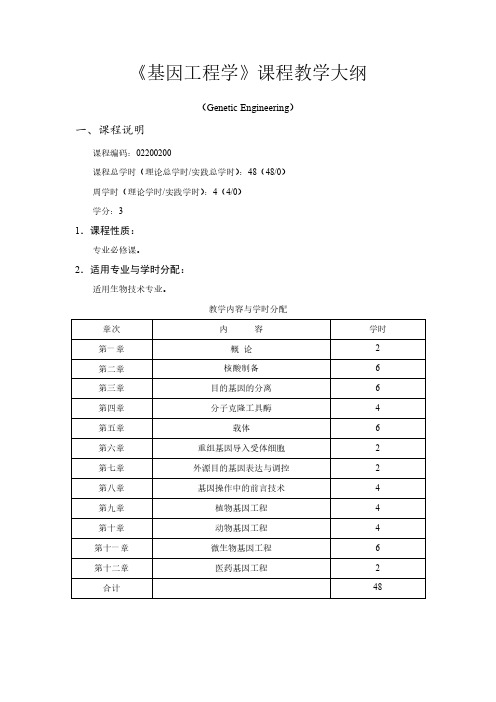
《基因工程学》课程教学大纲(Genetic Engineering)一、课程说明课程编码:02200200课程总学时(理论总学时/实践总学时):48(48/0)周学时(理论学时/实践学时):4(4/0)学分:31.课程性质:专业必修课。
2.适用专业与学时分配:适用生物技术专业。
教学内容与学时分配3.课程教学目的与要求:本课程的授课对象是生物技术专业的本科生。
课程简介:《基因工程》是生物技术专业的专业必修课程。
其以分子遗传学理论为基础,以分子生物学和微生物学的现代方法为手段而建立起来的一门技术学科。
基因工程兴起于20世纪70年代初,它的问世带动了生物技术的兴起和发展,是现代生物技术的核心内容。
基因工程课程的主要内容包括基因的分离、基因的克隆、基因的表达、植物基因工程、动物基因工程、药物基因工程和基因治疗等。
它是生命科学学院生物技术专业本科生的主干专业课程之一,它是生物工程(包括基因工程、细胞工程、酶工程、发酵工程)中最重要的课程,其它三大工程是建立在基因工程基础之上的,同时也为生物技术制药等后继学科奠定了重要的理论基础。
课程目标:设置本课程是为了让生物技术专业的学生理解和掌握基因工程的技术原理,通过本课程学习,掌握基因操作的工具酶,基因克隆常用载体,目的基因的分离与合成,重组体的构建,重组体向宿主细胞的导入,重组体克隆的筛选与鉴定以及克隆基因的表达,同时了解基因工程在生物学领域中的应用与发展前景。
对学生达到毕业要求贡献如下:1)了解基因工程学的历史、发展和前沿知识。
2)掌握基因工程学的基础理论、基本知识和基本技能;教学要求:学完基因工程学后,学生将具备以下能力:1)具有良好的自学能力;2)综合运用所掌握的基因工程学理论知识和技能、从事生物科学及其相关领域科学研究的能力。
4.本门课程与其它课程关系:先修课程为生物化学、微生物学、分子生物学、细胞学等,具备基础理论知识及实验能力是基因工程学课程的基础。
第五章 微生物基因工程育种

1960年,F.Jacob和J.Monmd提出了操纵元 (操纵子)的概念,揭示了原核生物基因表达 调控的重要规律。
基因的现代概念
移动基因(movable gene) 断裂基因(split gene) 假基因(pseudogene) 重复基因(repeated genes) 重叠基因(overlapping genes) 或嵌套基因(nested genes)
基 因 工 程 流 程 示 意 图
基因工程的应用
基因工程技术已经在医学、工业、 农业等各个领域得到了广泛的应用。
在医学上的应用
基因工程被用于大量生产过
去难以得到或几乎不可能得到的
蛋白质-肽类药物。
转基因动物和植物
转基因动物首先在小鼠获得成功。现在
转基因动物技术已用于牛、羊,使得从 牛/
第五章 工业微生物基因育种
王陶 2012年2月
目的与要求
了解和掌握基因工程育种的原理与方法; 了解基因工程的主要载体与基因定位诱变的 原理与方法。
教学重点:质粒的特性与基因工程操作原理 教学难点:基因定位诱变的原理
教学内容
1、概述 基因工程在微生物育种中的地位和作用,原理 2、基因工程载体 几种常见的载体 3、基因工程所用的酶 限制性内切酶,核酸酶,连接酶,聚合酶等 4、基因工程的主要步骤 5、基因定位诱变
孟 德 尔 研 究 的 七 对 性 状
豌豆杂交操作
孟 德 尔 分 离 律
孟 德 尔 自 由 组 合 律
黄圆 绿圆 黄皱 绿皱
1909年,丹麦的遗传学家W.
Johanssen 根据希腊语“给予生命”之义,创造了 “gene‖一词。但它只是一个抽象的单 位,并不代表物质实体。
噬菌体载体

第三章噬菌体载体一、填空题1.噬菌体之所以被选为基因工程载体,主要有两方面的原因:一是—-----------—;二是----------——。
2.第一个报道的全测序的单链DNA噬菌体是ФX174,DNA长5386个碱基对,共一个基因,为一环状DNA分子,基因组的最大特点是—----------—。
3.λ噬菌体的基因组DNA为———————kb,有——多个基因。
在体内,它有两种复制方式,扩增时(早期复制)按—-----—复制,成熟包装(晚期复制)则是按—--------—复制。
它有一个复制起点,进行—-------—向复制。
λ噬菌体的DNA既可以以线性存在又可以环状形式存在,并且能够自然成环。
其原因主要是在λ噬菌体线性DNA分子的两端各有一个——个碱基组成的天然黏性末端。
这种黏性末端可以自然成环。
成环后的黏性末端部位就叫做——————位点。
4.根据噬菌体的包装能力,将野生型λ噬菌体的基因组DNA改造成插入型载体,该载体的最小分子大小约为————kb,插入的外源片段最大不超过——————kb。
5.野生型的M13不适合用作基因工程载体,主要原因是————和--------------—。
6.黏粒(cosmid)是质粒—噬菌体杂合载体,它的复制子来自——、COS位点序列来自—--------—,最大的克隆片段达到—----------—kb。
7.有两类改造型的λ噬菌体载体,即插入型和取代型。
从酶切点看,插入型为——个,取代型为——个。
8.野生型的λ噬菌体DNA不宜作为基因工程载体,原因是:(1)---------------——(2)——————————(3)—---------------------—。
9. M13单链噬菌体的复制分为三个阶段:(1)————————(2)—-------------—,(3)———————————。
10.噬菌粒是由质粒和噬菌体DNA共同构成的,其中来自质粒的主要结构是—-----—,而来自噬菌体的主要结构是—-------------------—。
基因工程载体噬菌体载体课件

•24
(左右臂连接)
(三段自身连接) (插入片段)
•基因工程载体噬菌体载体
•25
(三)体外包装
λ噬菌体DNA体外重组后,一般必须经 过体外包装,然后以噬菌体感染的方式将
重组DNA导入E.coli细胞内。这是因为以感
染方式导入细胞的频率可达 106~108/μgDNA,而以转染(translation) 的方式导入的频率仅为103~105/ μgDNA。
特点是功能相近的基因在基 因组中聚集在一起。
•基因工程载体噬菌体载体
•12
λ噬菌体
组成特点:
双链线状DNA分子,
全长50kb,
含65个基因,
在其分子两端各含有12
个碱基的互补单链,是天
然的粘性末端,被称为
COS位点。
•基因工程载体噬菌体载体
•13
λ噬菌体感染细菌后的溶菌性生长和溶源性生长
溶菌性生长(lytic pathway) 噬菌体感染细菌后,借助其2个COS位点互补成环,在宿
主菌体内连续复制,合成大量基因产物,进而装配成噬菌体 颗粒,裂解宿主菌,释放出来的噬菌体又可感染其他细菌。 溶源性生长(lysogenic pathway)
噬菌体感染细菌后,可将自身DNA整合到细菌的染色体中 去,与之一起复制,并遗传给子代细胞,宿主细胞不被裂解 。
•基因工程载体噬菌体载体
•14
噬菌体
•41
•基因工程载体噬菌体载体
•42
Phage M13 replication in the host cell: Nicked by gene 2 protein
RNA pol +
RF DNA polIII
±
+
第3.4.5章载体

(2)插入失活 插入失活(insertional inactivation) 插入失活
在质粒pBR322的抗四环素基因上插入一 的抗四环素基因上插入一 在质粒 个外源基因后,导致抗四环素基因失活,变成 个外源基因后,导致抗四环素基因失活, 只对氨苄青霉素有抗性, 只对氨苄青霉素有抗性,这样就可通过对抗生 素是 双抗 还是 单抗 来筛选是否有外源基因片 段插入到载体中,这种筛选方法称为插入失活。 段插入到载体中, 这种筛选方法称为插入失活。
按工作方式
噬菌体载体、 噬菌体载体、 粘粒载体、 粘粒载体、 人工染色体载体等
克隆载体 按功能: 按功能 表达载体
分泌表达载体 细胞内表达载体
原核细胞载体 按受体细胞: 按受体细胞 真核细胞载体
载体的功能
运送外源基因高效转入受体细胞。 运送外源基因高效转入受体细胞。 为外源基因提供复制能力或整合能力。 为外源基因提供复制能力或整合能力。 为外源基因的扩增或表达提供必要的条件。 为外源基因的扩增或表达提供必要的条件。
有过百个限制性内切酶切点, 有过百个限制性内切酶切点,一种限制性内切酶只 有单一切口的位点也多达数十个, 有单一切口的位点也多达数十个,几乎具备了所有常 用限制酶都能切开并插入目的基因的优越条件。此外, 用限制酶都能切开并插入目的基因的优越条件。此外, pBR322DNA, pBR322DNA,被限制性内切酶消化后产生的片段大小均 已知道,可以作为核酸电泳的分子质量标准。 已知道,可以作为核酸电泳的分子质量标准。
iii)卡那霉素(kanamycin,Kan) )卡那霉素( , ) a)杀菌原理 ) 通过与70S核糖体结合,导致mRNA发 核糖体结合,导致 通过与 核糖体结合 发 生错读。杀死细菌。 生错读。杀死细菌。 b)细菌抗性原理 ) Kanr 编码的氨基糖苷磷酸转移酶,对 编码的氨基糖苷磷酸转移酶, 卡那霉素进行修饰, 卡那霉素进行修饰,阻断其与核糖体结 合作用。 合作用。
基因克隆的载体噬菌粒载体

基因克隆的载体噬菌粒载体
第3页
4. 存在着一个多克隆位点区,所以许各种不一样类型外 源DNA片段,不经修饰便可直接插入到载体分子上;
5. 因为多克隆位点区阻断了大肠杆菌lacZ基因5’端编码区,可按照IPTG组织化学显色反应试验, 筛选重组子;
基因克隆的载体噬菌粒载体
第2页
常见噬菌粒载体pUC118和pUC119
是一对分别由pUC18和pUC19质粒与野生型M13噬菌体基 因间隔区(IG)重组而成噬菌粒载体。
1. 含有较小分子量共价、闭合、环形基因组DNA,可克 隆10kb外源DNA片段,并易于进行分离与操作;
2. 编码一个ampr基因作为选择记号,所以只有携带 pUC118或pUC119噬菌粒载体大肠杆菌转化子细胞,才 能够在含有氨苄青霉素培养基中生长,便于转化子选择;
6. lacZ基因是置于lac开启子控制之下,这么插入 外源基因便会以融合蛋白质形式表示;
7. 含有质粒复制起点,在没有辅助噬菌体情况下, 克隆外源基因能够像质粒一样按常规方式,复 制形成大量双链DNA分子
基因克隆的载体噬菌粒载体
第4页
8. 带有一个M13噬菌体复制起点,在有辅 助噬菌体感染寄主细胞中,能够合成出 单链DNA拷贝,并包装成噬菌体颗分泌 到培养基中;
第6页
pBluescript噬菌粒载体
基T3和T7噬菌体开 启子,能够定向指导外源基因转录活动;
2. 同时含有一个单链噬菌体M13或f1复制起点 和一个来自ColE1质粒复制起点,在不一样情 况下,能够采取不一样复制形式,分别合成单 链或双链DNA;
3. 编码有一个胺苄青霉素抗性基因,供作转化 子记号;
第五章、噬菌体和粘粒载体

第一节 噬菌体的一般生物学特性 第二节 λ噬菌体载体 第三节 柯斯质粒载体 第四节 单链噬菌体载体 第五节 噬菌粒载体
第一节 噬菌体的一般生物学特性
5.1.1.1 结构及核酸
噬菌体颗粒基本类型:无尾二十面体型、 具尾二十面体型和线状体型。
噬菌体的核酸,最常见的是双链线性 DNA。除此之外,还发现有双链环形 DNA、 单 链 环 形 DNA、 单 链 线 性 DNA以及单链RNA等多种形式。
这类载体有插入型的,如凯伦2;也有替换型的,如凯伦30。
许多Charon噬菌体载体,都带有编码着从半乳糖苦酶基因以及它的操纵 基因和启动子的laeZ取代区段。
用替换型载体克隆外源DNA包括三个步骤:
第一,核酸内切酶消化载体, 除去可取代的DNA区段。
第二,λ DNA臂同外源 DNA 片段连接。
第三,重组体的 DNA分子进 行包装和增殖,得到有感染性 的λ重组噬菌体。
返回第五章
5.1.2.1 溶源周期的主要特征
返回第五章
已知存在两种不同类型的溶源周期,其中最普遍的一种是以λ噬菌体 为原型,具有如下特性:
①噬菌体的DNA分子注入细菌细胞。
②短暂的转录期之后,合成一种整合酶,之后转录活性便被一种阻 遏物所关闭。
③噬菌体DNA分子插入到细菌染色体基因组DNA上,变成原噬菌体。
能否发展出一种可将插入的DNA片段直接从λ载体转移到质粒载体的快 速简便的体内系统呢?
λZAP本质上是插入型载体。基因组中含有 一个可以在体内发生删除作用的pBluescript 噬菌粒(见本章第五节)的DNA片段,两 侧是由两个单链DNA噬菌体fi的复制信号。
pBluescript SK (+/-)
分子生物学载体课件

• 穿梭质粒 装有针对两种不同受体的复制 子,便于基因能在两种不同的受体细胞中 复制。
• 探 针 质 粒 筛选克隆或寻找基因元件。
23
如何阅读质粒图谱
• 1.首先看Ori的位置,了解质粒的类型(原核/真 核/穿梭质粒)
• 2.再看筛选标记,如抗性,决定使用什么筛选标记。 1 Ampr 水解β -内酰胺环,解除氨苄的毒性。 2 Tetr 可以阻止四环素进入细胞。 3 Camr 生成氯霉素羟乙酰基衍生物,使之失 去毒性。 4 neor(Kanr)氨基糖苷磷酸转移酶使G418 (卡那霉素衍生物)失活 5 Hygr 使潮霉素β 失活。
40
T4 Bacteriophage
41
42
2.2 噬菌体的生活周期(有两种)
1. 溶菌周期 —— 烈性噬菌体(virulent phage) 感染细菌后立即在菌体内复制和合成蛋白质外壳,重
新组装成噬菌体颗粒,并导致细菌细胞解体,释放出大量 子代噬菌体。 2. 溶原周期 —— 温和噬菌体(temperate phage)
• 2.8kb
• 元件来源 ① 复制起点:pBR322的 ori ② Ampr 基因:pBR322的Ampr基因,但其上失去了克 隆位点。
③ lacZ’基因:大肠杆菌lacZ的α-肽链序列, 是LacZ 的氨基端片断。在lacZ 基因中有一段人工合成的含多 种内切酶的单一切点,在此位点上插入外源DNA都能 灭活下游lacZ基因。
3 温控的质粒载体(runaway plasmid vectors) 这是一类温度敏感型复制控制质粒。
温度低(低于37 ℃),拷贝数很少; 温度增加(>40 ℃)时,拷贝数会很快增加到 1000个以上。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
阻遏物能够持续地合成,产量就其操纵基因而言是略为超量的。 由于阻遏物同OL和OR结合,PL和PR这两个早期启动子开始的转录 被抑制的。使原噬菌体保持“关闭”的形式,这就是溶源性细菌能够不 停地生长而不发生溶菌裂解的原因。 同时,后进入的同类噬菌体由于PL和PR被抑制,无法溶原化。称这 种抑制作用同源免疫超感染抗性。
DNA、 单 链 环 形 DNA、 单 链 线 性
DNA以及单链RNA等多种形式。
西北师范大学 精品课程 武国凡
基因工程 5.1.1.2 噬菌体的感染性
第五章、噬菌体和粘粒载体 返回第五章
少量噬菌体感染高浓度寄主细胞后,涂板,结果在平板上, 以最初被感染的细胞为中心,形成大量的噬菌斑。
西北师范大学 精品课程 武国凡
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
超感染的λ噬菌体DNA将会发生什么样的变化呢? 这种DNA可形成超盘旋的结构,由于处在超盘旋结构状
态的λDNA复制基因很难进行复制,而细菌细胞却没有受
到影响,仍能正常地生长和分裂,结果经过若干世代之 后,在细菌细胞群体中,感染的DNA分子便逐渐地被稀
西北师范大学 精品课程 武国凡
பைடு நூலகம்因工程
第五章、噬菌体和粘粒载体 返回第五章
西北师范大学 精品课程 武国凡
基因工程 5.1.2.2 超感染免疫性
第五章、噬菌体和粘粒载体 返回第五章
溶源性细菌有两个重要特点: 第一,溶源性细菌不能够被头一次感染并使之溶源化的同种噬菌体
感染。溶源性细菌所具有的这种抗御同种噬菌体感染的性质,叫超
许多Charon噬菌体载体,都带有编码着从半乳糖苦酶基因以及它的操纵
基因和启动子的laeZ取代区段。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
用替换型载体克隆外源DNA包括三个步骤:
第一,核酸内切酶消化载体,
除去可取代的DNA区段。
第二,λ DNA臂同外源 DNA 片段连接。 第三,重组体的 DNA 分子进 行包装和增殖,得到有感染性 的λ重组噬菌体。
西北师范大学 精品课程 武国凡
基因工程 5.1.2.4 溶源噬菌体的诱发
第五章、噬菌体和粘粒载体 返回第五章
溶源性细菌中的阻遏基因处于失活状态时,操纵基因就将
是自由的,转录就会正常启动,诱发噬菌体进入裂解途径。 噬菌体的诱发还需要合成一种叫做删除酶的噬菌体产物。 删除酶的功能是将原噬菌体 DNA 从细菌染色体 DNA 上删 除下来,而后环化成环形的DNA分子。。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
5.2.3 λ噬茵体载体的构建及其主要类型 (1)构建λ噬菌体载体的基本原理 消去多余的限制位点和非必要的区段:
野生型的 λ,有过多的限制位点,例如有 5 个 EcoRI 的限制
位点和7个Hindlll的限制位点。
所以说,构建λ载体的基本原理是多余限制位点的删除。
基因工程 第五章、噬菌体和粘粒载体 第一节 噬菌体的一般生物学特性 第二节 λ噬菌体载体 第三节 柯斯质粒载体 第四节 单链噬菌体载体 第五节 噬菌粒载体
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
第一节 噬菌体的一般生物学特性 5.1.1.1 结构及核酸
噬菌体颗粒基本类型:无尾二十面体型、 具尾二十面体型和线状体型。 噬菌体的核酸,最常见的是双链线性 DNA。除此之外,还发现有双链环形
基因至少有 61 个,其中有一半参与生命周期的活动,称为必须基 因;另一部分基因为非必须基因。 溶源周期中整合在染色体DNA上(λ作为克隆载体的重要特性)。 其它优点: 研究历史悠久; 寄主范围较质粒狭窄,更加安全; λ噬菌体DNA两端具有由 12个核着酸组成的粘性末端,可用来 构建质粒类载体。
基因工程
第五章、噬菌体和粘粒载体 返回第五章
②开半乳糖苷酶失活的插入型载体 基因组中含有一个大肠杆菌的lacZ 区段
lacZ 指示菌:lac一 培养基:IPTG和Xgal
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
替换型载体
又叫做取代型载体:提高克隆外源DNA片段的能力。
M.Thomas等人(1974):替换型载体 AgtAc,可取代片段 含有att、int及xis三个基因。 野生型载体可以处于稳定的溶源状态,并能产生出混浊型的 噬菌斑。 重组体载体失去了整合作用的功能,不能进入溶源状态,产 生出清晰型的噬菌斑。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
西北师范大学 精品课程 武国凡
基因工程 5.2.2 λ噬菌体DNA的复制
早期,按式复制
晚期,滚环复制。 受 λgam 基因产物控制, 抑 制 RecBC 酶 ( 内 、 外 切 酶)
第五章、噬菌体和粘粒载体 返回第五章
gam+ 的噬菌体只能产生
出多连体的DNA分子, gam - 噬 菌 体 只 能 产 生 出 单体分子。
感染免疫性。 第二,经过许多世代之后,溶源性细菌便能够开始进入溶菌周期,
这一过程叫溶源性细菌的诱发。在诱发过程中,噬菌体基因组以单
一DNA片段的形式从寄主染色体DNA上删除下来。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
λ噬菌体的超感染免疫性现象--阻遏一操纵体系
阻遏基因 cI 编码的阻遏蛋白,与两 个分别同启动基因PL、PR相邻的操 纵基因OL和OR结合。
④原噬菌体:在溶源性细菌内存在的整合的或非 整合的噬菌体DNA。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
5.1.2.1 溶源周期的主要特征
已知存在两种不同类型的溶源周期,其中最普遍的一种是以λ噬菌体 为原型,具有如下特性: ①噬菌体的DNA分子注入细菌细胞。 ②短暂的转录期之后,合成一种整合酶,之后转录活性便被一种阻 遏物所关闭。
凯伦噬菌体载体
的一种特殊的噬菌体载体。
凯伦噬菌体是 F.R.Blattner等人于 1977年在λ噬菌体基础上发展出来
凯伦( Charon)是古希腊神话中的一位老摆渡工,专司在斯蒂克斯河 (River Styx)上运送亡灵到阴间。所以 Blattner形象地称自己所构建的 噬菌体为凯伦载体。
这类载体有插入型的,如凯伦2;也有替换型的,如凯伦30。
西北师范大学 精品课程 武国凡
基因工程 5.2.4 λ噬茵体载体的改良
基因工程
第五章、噬菌体和粘粒载体 返回第五章
(1)red和gam控制DNA的重组作用,在感
染的早期使寄主 RecBC 蛋白质失去功能
( RecBC 蛋白质是一种多功能的核酸酶, 它具有核酸内切酶和核酸外切酶活性)。
(2)xis和int基因 (3)N和Q编码抗终止因子,分别控制早期功能和晚期功能的调节; (4)cI基因编码的蛋白质是一种阻遏物;cro蛋白也是一种阻遏物,能同 操纵基因OL和OR结合而抑制转录
λDNA首先环化,然后通过在两个 别 λDNA 和细菌 DNA 的附着位点, att物理性的破裂,并进行准确的噬 菌体DNA和寄主DNA的再接合,从 并能催化这两种 DNA之间发生链 而发生整合作用 的交换。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
疑问--插入之后为何不马上删除呢?
基因工程
第五章、噬菌体和粘粒载体 返回第五章
5.1.2.2 噬菌体的溶源生命周期
感染过程中没有产生出子代噬菌体颗粒,噬 菌体DNA是整合到寄主细胞染色体DNA上, 成为它的一个组成部分。
只有双链DNA的噬菌体才具有溶源周期。
①温和噬菌体: ②溶源性细菌:整合有一套完整的噬菌体基因组 的细菌。
③溶源化:用温和噬菌体感染细菌使之形成溶源 性细菌的过程。
基因工程 具体过程:
(1)尾部末端吸附;
第五章、噬菌体和粘粒载体 返回第五章
(2)通过尾轴把DNA全部注入到细 胞,蛋白质外壳则留在胞外;
(3)噬菌体的DNA在细菌体内合成自
身DNA和蛋白质; (4)新合成DNA与蛋白质组装子代噬 菌体; (5)细菌的解体,子代噬菌体释放。
西北师范大学 精品课程 武国凡
删除应该比插入更容易发生,因为此 时两个附着位点都是存在于同一 DNA分子上。 实际上:整合与删除不是同一过程。
其原因是整合后的位点是杂合的。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
第二节 λ噬菌体载体
研究得最为详尽,双链DNA,分子量31×106,48502bp;中等大小, 温和噬菌体。
释掉了。
西北师范大学 精品课程 武国凡
基因工程 5.1.2.3 异源免疫性
第五章、噬菌体和粘粒载体 返回第五章
有些温和噬菌体,如噬菌体 21 和 434 ,各自具有自身的 免疫体系,具有自身的阻遏基因和操纵基因。因此,其阻 遏物不能同λ的操纵基因结合;反过来,λ的阻遏物,也不 能够同21和434 噬菌体的操纵基因结合。这样的一对噬菌 体,互称为异源免疫性。 温和噬菌体可以在异源免疫性的溶源性细菌的菌苔上形 成噬菌斑。 噬菌体DNA的免疫性区段(包括cI基因、操纵基因和启 动基因),以imm表示,如immλ、imm21和imm434等。
西北师范大学 精品课程 武国凡
基因工程
第五章、噬菌体和粘粒载体 返回第五章
①免疫功能失活的插入型载体:
基因组中有一段免疫区 imm
当外源DNA插入时,合成活性阻遏物的功能遭受破坏,而不能进入溶源
周期。