第五章 噬菌体载体和柯斯载体
吴乃虎—基因工程原理

目录第一章基因与基因工程第一节基因研究的发展第二节基因的现代概念第三节基因工程的诞生及其主要的研究内容第二章基因操作的主要技术原理第一节核酸的凝胶电泳第二节核酸分子杂交第三节细菌转化1.肺炎球菌的转化2.大肠杆菌的转化3.细菌转化频率第四节DNA核苷酸序列分析第五节基因的化学合成第六节基因定点突变第七节基因扩增第八节研究DNA与蛋白质相互作用的方法第三章基因克隆的酶学基础第一节核酸内切限制酶与DNA分子的体外切割1.寄主控制的限制与修饰现象2.核酸内切限制酶的类型3.I 型和III型核酸内切限制酶的基本特性4.II型核酸内切限制酶的基本特性[1].基本特性[2].同裂酶[3].同尾酶[4].限制片段末端的连接作用5.核酸内切限制酶的命名法6.影响核酸内切限制酶活性的因素[1].DNA的纯度[2].DNA的甲基化程度[3].酶切消化反应的温度[4].DNA分子的结构[5].核酸内切限制酶的缓冲液7.核酸内切限制酶对DNA的消化作用[1].核酸内切限制酶与靶DNA识别序列的结合模式[2].核酸内切限制酶对DNA分子的局部消化作用[3].核酸内切限制酶对真核基因组DNA的消化作用第二节DNA连接酶与DNA分子的体外连接1.DNA连接酶2.粘性末端DNA片段的连接3.平末端DNA片段的连接[1].同聚物加尾法[2].衔接物连接法[3].DNA接头连接法4.热稳定的DNA连接酶[1].寡核苷酸连接测定法[2].连接酶链式反应(LCR)第三节DNA聚合酶1.DNA聚合酶I与核酸杂交探针的制备[1].DNA聚合酶I[2].DNA缺口转移[3].DNA杂交探针的制备2.大肠杆菌DNA聚合酶I 的Klenow片段与DNA末端标记3.T4 DNA聚合酶和取代合成法标记DNA片段4.依赖于RNA的DNA聚合酶与互补DNA的合成5.T7 DNA聚合酶6.修饰的T7 DNA聚合酶第四节DNA及RNA的修饰酶1.末端脱氧核苷酸转移酶与同聚物加尾2.T4多核苷酸激酶与DNA分子5’-末端的标记3.碱性磷酸酶与DNA脱磷酸作用第五节核酸外切酶1.核酸外切酶VII (exo VII)2.核酸外切酶III (exo III)3.λ核酸外切酶(λ exo)和T7基因6核酸外切酶第六节单链核酸内切酶1.S1核酸酶与RNA分子定位2.Bal1 核酸酶与限制位点的确定第四章基因克隆的质粒载体第一节质粒的一般生物学特性1.质粒DNA2.质粒DNA编码的表型3.质粒DNA的转移[1].质粒的类型[2].F质粒[3].质粒DNA的接合作用4.质粒DNA的迁移作用5.质粒DNA的复制类型6.质粒DNA的不亲和性[1].质粒的不亲和性现象[2].质粒不亲和性的分子基础7.第二节质粒DNA的复制与拷贝数的控制1.质粒DNA复制的多样性2.ColE 1质粒DNA复制的启动3.质粒DNA拷贝数的控制[1].天然质粒拷贝数的控制[2].杂种质粒拷贝数的控制4.质粒复制控制的分子模型[1].抑制蛋白质稀释模型[2].自体阻遏蛋白质模型5.第三节质粒DNA的分离与纯化1.氯化铯密度梯度离心2.碱变性法3.微量碱变性法4.影响质粒DNA产量的因素[1].寄主菌株的遗传背景[2].质粒的拷贝数与分子大小5.第四节质粒载体的构建与类型1.天然质粒用作克隆载体的局限性2.质粒载体必须具备的基本条件3.质粒载体的选择记号[1].高拷贝数的质粒载体[2].低拷贝数的质粒载体[3].失控的质粒载体[4].插入失活型的质粒载体[5].正选择的质粒载体[6].表达型的质粒载体4.第五节重要的大肠杆菌质粒载体1.pSC101 质粒载体[1].应用pSC101 质粒作基因克隆载体的实例一---葡萄球菌质粒基因在大肠杆菌中的表达[2].应用pSC101 质粒作基因克隆载体的实例二---在大肠杆菌中克隆非洲爪蟾2.Col 1质粒载体3.pBR322质粒载体[1].pBR322质粒载体的构建[2].pBR322质粒载体的优点[3].pBR322质粒载体的改良[4].应用pBR322质粒作为基因克隆载体的实例---水稻夜绿体光诱导基因psbA的结构分析4.pUC 质粒载体[1].pUC 质粒载体的结构[2].pUC 质粒载体的优点5.其他重要的质粒载体[1].丧失迁移功能的的质粒载体[2].能在体外转录克隆基因的质粒载体[3].穿梭质粒载体第六节质粒载体的稳定性问题1.质粒载体不稳定性的类型[1].结构的不稳定性[2].分离的不稳定性2.影响质粒载体稳定性的主要因素[1].新陈代谢负荷对质粒载体稳定性的效应[2].拷贝数差度对质粒载体稳定性的影响[3].寄主重组体系对质粒载体稳定性的效应3.随机分配的分子机理[1].通过精巧的控制环路使质粒拷贝数的差度限制在最低的水平[2].通过位点特异的重组作用消除天然质粒的寡聚体[3].通过调节细胞的分裂活动阻止无质粒细胞的产生[4].大肠杆菌素的合成增进了质粒的稳定性4.主动分配的分子机理[1].分配区的结构与功能[2].预配对模型[3].二聚体的解离有助于质粒的主动分配[4].寄主致死功能对质粒稳定性的效应5.第五章噬菌体载体和柯斯载体第一节噬菌体的一般生物学特性第二节λ噬菌体载体第三节柯斯质粒载体第四节单链DNA噬菌体载体第五节噬菌体载体第六章基因的分离与鉴定第一节DNA克隆片段的产生与分离1.基因组DNA克隆片段的产生与分离2.DNA片段的大小分部3.编码目的基因的克隆片段的富集第二节重组体DNA分子的构建及导入受体细胞1.外源DNA片段同载体分子的重组[1].外源DNA片段定向插入载体分子[2].非互补粘性末端DNA分子间的连接[3].最佳连接反应2.重组体分子导入受体细胞的途径[1].重组体DNA分子的转化或转染[2].体外包装的λ噬菌体的转导第三节基因克隆的实验方案1.互补作用基因克隆2.cDNA基因克隆[1].cDNA文库的建立[2].不同丰度mRNA的cDNA克隆[3].全长cDNA的合成[4].cDNA克隆的优越性3.基因组DNA克隆[1].应用 噬菌体载体构建基因组文库[2].应用柯斯质粒载体构建基因组文库4.基因定位克隆[1].拟南芥菜简介[2].RFLP分子标记[3].RFLP作图的原理与步骤[4].染色体步移[5].大尺度基因组物理图谱的构建第四节克隆基因的分离1.应用核酸探针分离克隆的目的基因[1].核酸探针的来源[2].寡核酸探针的的人工合成[3].假阳性克隆的克服2.应用差别杂交或扣除杂交法分离克隆的目的基因[1].差别杂交[2].差别杂交的局限性[3].扣除杂交3.应用mRNA差别显示技术分离克隆的目的基因[1].mRNA差别显示的原理[2].mRNA差别显示的基本过程[3].mRNA差别显示的局限性4.引用表达文库分离克隆的目的基因5.酵母双杂交体系[1].酵母双杂交体系的基本原理[2].酵母双杂交体系的寄主菌株[3].酵母双杂交体系的实验程序第五节重组体分子的选择与鉴定1.遗传检测法[1].根据载体表型特征选择重组体分子的直接选择法[2].根据插入序列的表型特征选择重组体分子的直接选择法2.物理检测法[1].凝胶电泳检测法[2].R-检测环法3.菌落或噬菌斑杂交筛选法4.免疫化学检测法[1].放射性抗体检测法[2].免疫沉淀检测法[3].表达载体产物之免疫化学检测法5.DNA蛋白筛选法6.转译筛选法[1].杂交抑制的转译[2].杂交选择的转译第七章基因的表达与调节第八章真核基因在大肠杆菌中的表达第一节真核基因的大肠杆菌表达体系第二节大肠杆菌的表达载体第三节克隆的真核基因在大肠杆菌中的表达第四节影响克隆基因在大肠杆菌中表达效率的因素第九章植物基因工程第十章哺乳动物基因工程第一节哺乳动物基因转移的遗传选择标记第二节外源DNA导入哺乳动物细胞的方法第三节SV 40病毒载体第四节反转录病毒载体第五节其他的病毒载体第十一章重组DNA与现代生物技术第十二章重组DNA与医学研究第一章基因与基因工程第一节基因研究的发展第二节基因的现代概念第三节基因工程的诞生及其主要的研究内容1.质的新组合,并使之参与到原先没有这类分子的寄主细胞内,而能够持续稳定的繁殖。
第五章DNA克隆用载体

三、质粒的构建
(一) 质粒人工构建的方法 方法就是在天然质粒的基础上进行
DNA重组,拼拼接接,以改变其性 能。
(二) 质粒人工构建的目的
(1)加入合适的选择标记基因,如两个 以上,易于用作选择;
(2)增加或减少合适的酶切位点,便于 重组;
(3)缩短长度,切去不必要的片段,提 高导入效率,增加装载量;
质粒pBR322 DNA
pBR322 质粒图谱
多克隆位点
氨苄青霉素 抗性基因
四环素抗 性基因
pUC系列的质粒载体
pUC系列的质粒载体集中了当时载体的诸多优点。 包括4个部分: ①来自pBR322的复制起点(ori); ②氨苄青霉素抗性基因(ampr); ③大肠杆菌乳糖操纵子的调节基因(laci)启动子 (Plac)和β 半乳糖苷酶的α -肽(lacZ); ④位于lacZ基因中靠近5′端的一段多接头,或称为 多克隆位点(MCS)。
因此通过蓝白斑筛选可 以筛选、鉴定重组体与非重 组体载体。
pGEX-6P-1 质粒物理图谱
SV40, 猴空泡病毒40
巨细胞病毒 (CMV)
(二)不相容性
利用同一复制系统的不同质粒(RNAI RNAII Rop因子)如果被导入同一细胞 中,它们在复制及随后分配到子细胞的过 程中,就会彼此竞争,它们在单细胞中的 拷贝数也会有差异,拷贝多的复制更快, 结果在细菌繁殖几代之后,细菌的子细胞 中绝大多数都含有占优势的质粒,因而这 两种质粒中只能有一种长期稳定地留在细 胞中,这就是所谓的质粒不相容性。
MAC (Mammalian Artificial Chromosome)
病毒载体
E.coli E.coli 酵母细胞 哺乳类细胞 动物细胞
环状 环状 线性染色体 线性染色体 环状
载体介绍

pBR322—→插入在Ampr 中,基因型为Tetr 、Amps 、—— →在含有氨卞青霉素培养基上不生长,在含有四环素培 养基可生长; 而在两种抗生素培养基上都生长的是非重组型。
这种在一个基因位点中插入外源DNA片段,从而使该 基因活性丧失的现象叫插入失活。
外源基因的插入:
PstI Amp r Tet r Amp s Tet r
酵母人工染色体载体(yeast artificial chromosome, YAC)
YAC的特点: 1.只能在酵母细胞中扩增 2.克隆容量大,一般为200kb-500kb 3.构建原理,按染色体结构构建
染色体复制和遗传的三个基本组件:
1.自主复制序列(ARS) (autonomous replicating sequence) DNA复制起始点(ori) 2.着丝粒(centromere,cen) 3.端粒(telomeres,tel)
P1 噬菌体载体和 P1 人工染色体载体
P1 噬菌体 :大肠杆菌的一种溶原性噬菌体。 P1 噬菌体载体 工作原理类似黏粒载体 外源片断70-100kb
P1 人工染色体载体 P1 噬菌体载体的改造,结合了P1 噬菌体载体和 BAC载体的优点。 克隆外源片断130-150kb
(详细介绍)质粒载体
什么是质粒?
1.“严紧型”复制控制的质粒(stringent plamid):拷 贝数少(1-5个); 2.“松弛型”复制控制的质粒(relaxed plasmid):拷 贝数多(10-200个)。
质粒DNA的转移
1. 质粒的类型 据能否自我转移可分为:
接合型(conjugative plasmid)
非接合型(non-conjugative plasmid) 2. F质粒(fertility factor)
第七讲 噬菌体载体与柯斯载体

第七讲噬菌体载体与柯斯载体吴乃虎中国科学院遗传与发育生物学研究所2005年8月目录一、噬菌体的一般问题1.何谓噬菌体2.噬菌体效价测定与双层平板法3.噬菌体的生命周期4.噬菌体的溶源生命周期5.超感染免疫6.溶源性的诱发7.Campbell模型二、λ噬菌体载体1.λ噬菌体的生物学概述2.λ噬菌体载体的构建(1)λ噬菌体载体构建的基本原理(2)插入型载体(3)替换型载体3.λ噬菌体载体的改良(1)Spi-正选择的λ噬菌体载体(2)具有内删除特性的λ噬菌体载体4.λ重组体DNA分子的体外包装(1)λ重组体DNA的转染作用(2)λ噬菌体的体外包装5.λ噬菌体DNA的包装限制问题(1)包装限制的概念(2)包装限制与λ噬菌体的克隆能力(3)包装限制的生物学意义6.λ重组体分子的选择方法(1)cI基因功能选择法(2)lacZ基因功能选择法(3)Spi-选择法三、柯斯质粒载体1.柯斯质粒的定义及其构建(1) 噬菌体的克隆能力(2)柯斯质粒载体定义(3)柯斯质粒pHC79的构建(4)柯斯质粒克隆能力及其组成2.柯斯质粒载体的特点3.柯斯质粒载体的改良4.柯斯克隆(1)定义(2)理论依据(3)柯斯质粒包装条件(4)柯斯克隆程序(5)柯斯克隆的优点5.柯斯克隆的改良(1)柯斯克隆的局限性及其克服办法(2)Ish-Horowicz-Burke克隆方案(3)Bates-Swift克隆方案噬菌体载体与柯斯质粒载体一、噬菌体的一般问题1.何谓噬菌体?(1)定义噬菌体英文名为Bacteriophage,简称phage,来源于希腊文“phages”,系指吞食之意,乃是一类细菌病毒的总称需要指出的是,“phage”这个英文单词既是单数又是复数形式。
作单数使用时是指一个噬菌体,如试管中含1个T4 phage;而用作复数使用时,则是指同一种噬菌体的众多颗粒,如试管中含有上百个的T4 phage。
“phages”是噬菌体的复数词,是指多种不同的噬菌体。
大学《基因工程学》教学大纲
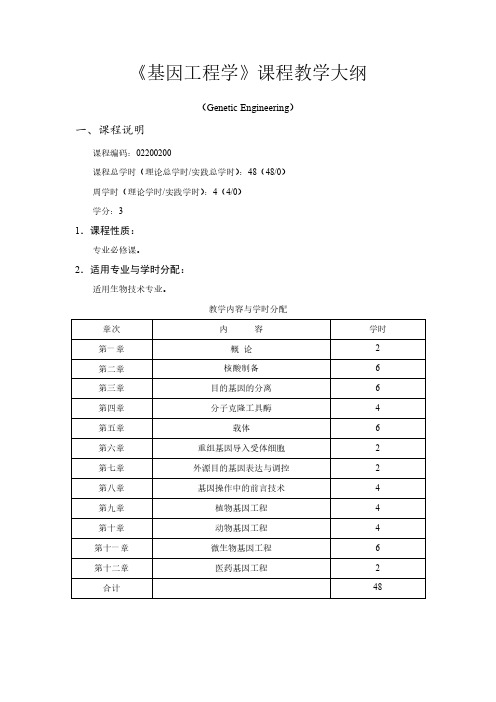
《基因工程学》课程教学大纲(Genetic Engineering)一、课程说明课程编码:02200200课程总学时(理论总学时/实践总学时):48(48/0)周学时(理论学时/实践学时):4(4/0)学分:31.课程性质:专业必修课。
2.适用专业与学时分配:适用生物技术专业。
教学内容与学时分配3.课程教学目的与要求:本课程的授课对象是生物技术专业的本科生。
课程简介:《基因工程》是生物技术专业的专业必修课程。
其以分子遗传学理论为基础,以分子生物学和微生物学的现代方法为手段而建立起来的一门技术学科。
基因工程兴起于20世纪70年代初,它的问世带动了生物技术的兴起和发展,是现代生物技术的核心内容。
基因工程课程的主要内容包括基因的分离、基因的克隆、基因的表达、植物基因工程、动物基因工程、药物基因工程和基因治疗等。
它是生命科学学院生物技术专业本科生的主干专业课程之一,它是生物工程(包括基因工程、细胞工程、酶工程、发酵工程)中最重要的课程,其它三大工程是建立在基因工程基础之上的,同时也为生物技术制药等后继学科奠定了重要的理论基础。
课程目标:设置本课程是为了让生物技术专业的学生理解和掌握基因工程的技术原理,通过本课程学习,掌握基因操作的工具酶,基因克隆常用载体,目的基因的分离与合成,重组体的构建,重组体向宿主细胞的导入,重组体克隆的筛选与鉴定以及克隆基因的表达,同时了解基因工程在生物学领域中的应用与发展前景。
对学生达到毕业要求贡献如下:1)了解基因工程学的历史、发展和前沿知识。
2)掌握基因工程学的基础理论、基本知识和基本技能;教学要求:学完基因工程学后,学生将具备以下能力:1)具有良好的自学能力;2)综合运用所掌握的基因工程学理论知识和技能、从事生物科学及其相关领域科学研究的能力。
4.本门课程与其它课程关系:先修课程为生物化学、微生物学、分子生物学、细胞学等,具备基础理论知识及实验能力是基因工程学课程的基础。
第四、五章 噬菌体载体及粘粒及目的基因的准备、重组及重组子的鉴定

•3、核酸探针(screening a library)
• • • • • • • •
4、免疫化学检测 *标记抗体探针 *免疫沉淀 5、转译筛选法 *杂交抑制的转译-丰富mRNA *杂交选择的转译-低丰度mRNA-insulin 6、正负筛选法 组织特异性、发育阶段特异性、特殊处理后 基因表达差异
• • • • •
4、重组DNA分子的构建 噬菌体载体构建 柯斯质粒载体构建 5、重组体的筛选 四、PCR方法与化学合成
生产 DNA 片段
限制性内切酶处理 机械剪切 从 mRNA 合成 DNA 基因化学合成 PCR 法
DNA 片段与载 体 DNA 分子连接
同聚物加尾
粘性末端连接
平端连接
连接子连接
四、凯伦噬菌体载体(charon)
• 两类 • 特点:酶切位点多、含LacZ、容量大 • 克隆程序:分离线性噬菌体DNA-酶切分 离左右臂+中央区段-与外源DNA混合-包 装 • 几kb~23kb • 噬菌体载体及派生载体的优点:感染效 率高
五、柯斯质粒载体
• • • • • • 1、柯斯质粒载体的构建 定义:(cos site-carrying plasmid) eg.pHC79(+pBR322) cos+与噬菌体包装有关的序列 ori+Ampr+Tetr 容量:31~45kb
第五章 目的基因的准备、重组及• 表达 • 测定基因结构(内含子 外显子)、时间调节基因 表达特征分析 • 2、缺点: • 无调节序列 • 低丰度mRNA克隆难
• • • • • •
4、cDNA两链的合成 5、双链cDNA分子与载体的连接 *同聚物加尾 *linker-连接子 6、重组体导入宿主 转化、转染、电脉冲
基因工程载体噬菌体载体

基因转殖
噬菌体载体可以在转基因植 物和动物中嵌入外源基因, 实现基因转殖和遗传改良。
洗牌策略
利用噬菌体载体的洗牌策略, 可以分离和筛选特定的DNA 或蛋白质序列,用于疾病诊 断和药物研发。
噬菌体载体的优势与限制
1 优势
高效的DNA复制和表达能力、容易获取和构建、适用于广泛的宿主细胞、可高表达目标 基因。
农业改良
噬菌体载体在基因工程研究中仍 有很大潜力,可以用于新型基因 编辑技术和药物运荷系统的开发。
噬菌体载体可以应用于药物运输、 基因疗法和肿瘤免疫治疗等领域 的创新药物开发。
噬菌体载体可用于改良农作物、 提高农作物的抗害能力和产量。
2 限制
容量有限、宿主细胞影响表达水平、病毒复制可能导致细胞毒性和免疫反应。
噬菌体载体构建与筛选方法
1
构建方法
通过DNA重组技术将目标DNA序列插入到噬菌体载体的适当位置,并使用DNA连接酶连接两 者。
2
筛选方法
利用选择标记和可视化标记(如荧光蛋白)来鉴定带有目标DNA序列的噬菌体载体。
噬菌体载体在基因表达与蛋白质生产中的 应用
基因工程载体噬菌体载体
了解噬菌体载体的定义、结构与类型以及在基因工程中的应用。探讨其优势 与限制,构建与筛选方法,以及在基因表达与蛋白质生产中的应用。最后, 展望噬菌体载体的未来发展趋势。
噬菌体载体的定义
噬菌体载体是指噬菌体基因组中携带外源DNA序列的DNA分子。它可以被用 来传递、复制和表达外源基因,对研究和应用领域具有重要意义。
Байду номын сангаас
基因表达
噬菌体载体可用于高效表达外 源基因,生产重组蛋白质,如 药物、酶和抗原。
蛋白质纯化
基因工程的载体种类

基因⼯程的载体种类基因⼯程的载体对于外源基因的复制、扩增、传代乃⾄表达⾄关重要,其必需具备以下条件:①具有有效运载能⼒,能够进⼊宿主细胞;②对多种限制酶有单⼀或较少的切点,最好是单⼀切点,即本⾝是⼀个复制⼦,携带外源基因前后均能在宿主细胞内⾃主复制,或者能够整合到宿主细胞中;③在宿主中能控制外源基因的表达活动;④要有筛选标记,鉴定⽅便,装卸⼿续简单;⑤容易控制,安全可靠。
在基因⼯程(DNA重组)中,使⽤的载体有:①克隆载体(clone vector),即以繁殖DNA分⼦为⽬的的载体;②穿梭载体(shuttle vecto),⽤于真核⽣物DNA⽚段在原核⽣物中增殖,然后在转⼊真核⽣物细胞宿主表达;③表达载体(express vector),⽤于⽬的基因的表达。
现在对载体提出了更⾼的要求,如:⾼拷贝数、具有强启动⼦和稳定的mRNA、具有⾼的分离稳定性和结构稳定性、转化频率⾼、宿主范围⼴、插⼊外源基因容量⼤且可以重新完整地复制与转录、和宿主细胞匹配等。
此外,载体在宿主不⽣长或低⽣长速率时仍能⾼⽔平地表达⽬的基因。
但达到上述要求的载体很少,尤其是当动物细胞作为宿主细胞时,⽬前能⽤的主要时病毒,进⼊宿主的⽬的基因⼀般只能是⼀个基因,⽽以基因组或多个基因同时进⾏重组还有⼀定困难。
⼀、质粒克隆载体除酵母杀伤质粒(killer plasmid)为RNA外,其他质粒多位环状DNA分⼦,每个质粒都有⼀段DNA复制起始点的序列,帮助实现质粒的复制。
质粒⼀般决定抗⽣素的抗性、产⽣抗⽣素酶系、糖酵解酶系、降解芳⾹族化合物酶系、肠毒素及限制-修饰酶系等。
其中严紧型复制控制质粒的复制与宿主染⾊体同步,并与宿主蛋⽩质合成有关,与DNA聚合酶I活性⽆关,蛋⽩质合成停⽌,质粒与宿主染⾊体复制亦停⽌,故只有1个或少数⼏个拷贝;⽽松弛型复制控制质粒的复制与宿主染⾊体复制不同步,与蛋⽩质合成⽆关,与DNA聚合酶I活性有关,蛋⽩质合成停⽌,质粒仍可复制,故可以在宿主有10—206个拷贝。
基因工程原理讲义:噬菌体载体与柯斯载体

第七讲噬菌体载体与柯斯载体中国科学院遗传与发育生物学研究所2017年8月目录一、噬菌体的一般问题1.何谓噬菌体2.噬菌体效价测定与双层平板法3.噬菌体的生命周期4.噬菌体的溶源生命周期5.超感染免疫6.溶源性的诱发7.Campbell模型二、λ噬菌体载体1.λ噬菌体的生物学概述2.λ噬菌体载体的构建(1)λ噬菌体载体构建的基本原理(2)插入型载体(3)替换型载体3.λ噬菌体载体的改良(1)Spi-正选择的λ噬菌体载体(2)具有内删除特性的λ噬菌体载体4.λ重组体DNA分子的体外包装(1)λ重组体DNA的转染作用(2)λ噬菌体的体外包装5.λ噬菌体DNA的包装限制问题(1)包装限制的概念(2)包装限制与λ噬菌体的克隆能力(3)包装限制的生物学意义6.λ重组体分子的选择方法(1)cI基因功能选择法(2)lacZ基因功能选择法(3)Spi-选择法三、柯斯质粒载体1.柯斯质粒的定义及其构建(1) 噬菌体的克隆能力(2)柯斯质粒载体定义(3)柯斯质粒pHC79的构建(4)柯斯质粒克隆能力及其组成2.柯斯质粒载体的特点3.柯斯质粒载体的改良4.柯斯克隆(1)定义(2)理论依据(3)柯斯质粒包装条件(4)柯斯克隆程序(5)柯斯克隆的优点5.柯斯克隆的改良(1)柯斯克隆的局限性及其克服办法(2)Ish-Horowicz-Burke克隆方案(3)Bates-Swift克隆方案噬菌体载体与柯斯质粒载体一、噬菌体的一般问题1.何谓噬菌体?(1)定义噬菌体英文名为Bacteriophage,简称phage,来源于希腊文“phages”,系指吞食之意,乃是一类细菌病毒的总称需要指出的是,“phage”这个英文单词既是单数又是复数形式。
作单数使用时是指一个噬菌体,如试管中含1个T4 phage;而用作复数使用时,则是指同一种噬菌体的众多颗粒,如试管中含有上百个的T4 phage。
“phages”是噬菌体的复数词,是指多种不同的噬菌体。
植物基因工程中的λ噬菌体载体

•右臂:长约10kb,控 制溶菌和溶原生长最重 要的调控基因和序列、 以及λDNA复制起始均 在这区域内。
➢ 基因组长约50kb ,至少包括61个基因,除少数例外,大多数编码基因均是按功能的相似 性成簇排列。左右臂包含λDNA复制、噬菌体结构蛋白合成、组装成熟噬菌体、溶菌生 长所需全部序列;对溶菌生长来说,中段是非必需的。 植物基因工程中的λ噬菌体载体
植物基因工程中的λ噬菌体载体
λ早使用的载体 系统,其主要优点是插入片段的装载容量大,适合于 全长的eDNA克隆,不仅质量高、代表性好,而且重组 噬菌体颗粒的感染活性在替换或插入某些标志基因,如上述的可 供蓝白筛选lacZ’序列和多克隆位点等。
• 建立重组λDNA分子的体外包装系统。
• 与一般的质粒载体不同, 噬菌体转染前需要利用噬菌体外 壳蛋白和噬菌体DNA加工酶组成的混合物(包装抽提物
• )在体外将连接好的DNA线性分子包装成噬菌体颗粒
植物基因工程中的λ噬菌体载体
植物基因工程中的λ噬菌 体载体
2020/11/20
植物基因工程中的λ噬菌体载体
载体(vector)是把一个有用的目的DNA片段通过重 组DNA技术,送进受体细胞中去进行繁殖和表达的工 具。 植物基因工程技术中尤为重要的是载体,不同目的 基因需要采用不同的载体。 组成植物基因工程载体系统常见的几种载体有:
右臂,填充片断的存在会影响连接效率, 必须纯化左臂和右臂。
植物基因工程中的λ噬菌体载体
(3)外源片断与载体的连接 • 通过载体的粘性末端,将载体连接成多联
体,以利于将两个cos位点之间的片断装入 噬菌体颗粒
植物基因工程中的λ噬菌体载体
(4)重组噬菌体的体外包装,形成有感染力 的噬菌体颗粒
噬菌体载体和柯斯载体

•λ 噬 菌 体 分 子 量 31×106dal,中等大 小的温和噬菌体,目 前已经定位61个基因, 其中一半参与了生命 周期,为必要基因; 余下的为非必要基因。
溶菌阶段(复制和释放)来自溶源阶段(整合寄主染色体)
一、λ噬菌体的分子生物学概述 λ phage
1、基因组结构 (1)cos位点 • 线性双链DNA分子两端各有一条12nt组成的 彼此完全互补的5’突出单链 • 注入宿主后,粘性末端互补形成双链区,成 为cos位点。
四、噬菌体的溶源生命周期
溶源生长周期:是指在感染过程中没有产 生出子代噬菌体颗粒,噬菌体的DNA是整 合到寄主细胞染色体DNA上,成为它的一 个组成部分。
现知道只有双链DNA的噬菌体才具有溶源 周期
1、概念 温和噬菌体:既能进入溶菌生命周期又能进
入溶源生命周期。 溶源性细菌:具有一种完整的噬菌体基因组
有 些 噬 菌 体 的 DNA 碱 基 不 是 由 标 准 的 A/T/C/G组成的。 Example:
T4噬菌体DNA没有C碱基,取代的是5-羟甲基胞
嘧啶(HMC);SP01噬菌体DNA中没有T碱基, 取代的是5-羟甲基尿嘧啶(HMU)。
二、噬菌体的感染性 噬菌斑形成
噬菌体
感染高浓度细菌
在未发生第一次裂解 前涂布平板
一、 噬菌体的结构和其核酸类型 结构:无尾部结构的二十面体型、
具尾部结构的二十面体型、 线状体型 核酸类型:最常见的是双链线性DNA、
双链环形DNA、 单链环形DNA、 单链线形DNA、 单链RNA。
不同的噬菌体之间的核酸分子量的差异也是 比较大的。大分子量的噬菌体生命周期比较 复杂;对寄主的依赖性较低
五、重组噬菌体的分离
第五章、噬菌体和粘粒载体

第一节 噬菌体的一般生物学特性 第二节 λ噬菌体载体 第三节 柯斯质粒载体 第四节 单链噬菌体载体 第五节 噬菌粒载体
第一节 噬菌体的一般生物学特性
5.1.1.1 结构及核酸
噬菌体颗粒基本类型:无尾二十面体型、 具尾二十面体型和线状体型。
噬菌体的核酸,最常见的是双链线性 DNA。除此之外,还发现有双链环形 DNA、 单 链 环 形 DNA、 单 链 线 性 DNA以及单链RNA等多种形式。
这类载体有插入型的,如凯伦2;也有替换型的,如凯伦30。
许多Charon噬菌体载体,都带有编码着从半乳糖苦酶基因以及它的操纵 基因和启动子的laeZ取代区段。
用替换型载体克隆外源DNA包括三个步骤:
第一,核酸内切酶消化载体, 除去可取代的DNA区段。
第二,λ DNA臂同外源 DNA 片段连接。
第三,重组体的 DNA分子进 行包装和增殖,得到有感染性 的λ重组噬菌体。
返回第五章
5.1.2.1 溶源周期的主要特征
返回第五章
已知存在两种不同类型的溶源周期,其中最普遍的一种是以λ噬菌体 为原型,具有如下特性:
①噬菌体的DNA分子注入细菌细胞。
②短暂的转录期之后,合成一种整合酶,之后转录活性便被一种阻 遏物所关闭。
③噬菌体DNA分子插入到细菌染色体基因组DNA上,变成原噬菌体。
能否发展出一种可将插入的DNA片段直接从λ载体转移到质粒载体的快 速简便的体内系统呢?
λZAP本质上是插入型载体。基因组中含有 一个可以在体内发生删除作用的pBluescript 噬菌粒(见本章第五节)的DNA片段,两 侧是由两个单链DNA噬菌体fi的复制信号。
pBluescript SK (+/-)
噬菌粒载体

二.pUCll8和 pUCll9噬菌粒载体
Puc118和PUCll9是一对分别由pUC18和pUC19质粒与野生型M13噬菌 体的基因间隔区(IG)重组而成的噬菌粒载体。除了多克隆位点区的 序列取向彼此相反而外,两者的分子结构完全一样(图5-15)。 如果有某一种特定的外源基因被插入在这一对噬菌体载体多克隆位点 区的同一种限制位点上,那么所形成的重组载体中的一个将转录克隆 基因的正链DNA,而另一重组载体则转录克隆基因的负链DNA。所以, 应用Puc118和PUCll9噬菌粒作载体,克隆基因的两条链l9噬菌粒载体的复制模式 具有两种不同的复制模型(图5-16)。 1.在寄主细胞没有感染上辅助噬菌体M13的情况下,pUCll8和pUCll9噬 菌粒载体的复制就如同双链质粒载体DNA一样,是受起源于ColE1质粒的LK复 制起点控制的。按这种复制模式产生的双链载体DNA分子仍然保留在寄主细 胞内[图5-16(a)]。 2.当寄主细胞被辅助噬菌体M13感染之后,pUCll8和pUCll9载体便转向 按照M13噬菌体的滚环模型进行复制。它是受存在于这两个PUC载体上的M13 噬菌体复制起点控制的。所产生的单链载体DNA分子可以被包装进由辅助噬 菌体提供的外壳蛋白质中,形成的噬菌体颗粒随后被挤压出寄主细胞,分布 在周围的培养基中〔图5-16(b)〕。
(2)pUCll8和pUCll9噬菌粒载体的优点 主要的有如下几个方面: (i)具有小分子量的共价、闭合、环形的基因组DNA,可克隆高达10kb的外源DNA片段,并易于进行体外分离与 操作; (ii)编码有一个ampr基因作为选择记号,因此只有携带着pUCll8或pUCll9噬菌粒载体的大肠杆菌转化子细胞, 才能够在含有氨千青霉素的培养基中生长,便于转化子的选择; (iii)拷贝数含量高,每个寄主细胞可高达500个,所以只要用少量的大肠杆菌细胞培养物,便可制备出大量的 载体DNA; (iv)存在着一个多克隆位点区,因此许多种不同类型的外源DNA限制片段,不经修饰便可直接插入到载体分子 上; (v)由于多克隆位点区阻断了大肠杆菌lacZ基因的5′-端编码区,故可按照Xgal-IPTG组织化学显色反应试验, 筛选重组体分子; (vi)lacZ基因是置于lac启动子的控制之下,这样插入的外源基因(当其读码结构没有发生改变的情况下)便 会以融合蛋白质形式表达,即产生出β 半乳糖管酶与外源蛋白质的融合产物; (vii)含有一个质粒的复制起点,因此在没有辅助噬菌体的情况下,克隆的外源基因可以像质粒一样按常规方 法,复制形成大量的双链DNA分子; (viii)带有一个M13噬菌体的复制起点,所以在有辅助噬菌体感染的寄主细胞中,可以合成出单链DNA拷贝,并 包装成噬菌体颗粒分泌到培养基中; (ix)在Puc118和pUCll9这两个载体中,多克隆位点区的核着酸序列取向是彼此相反的,于是它们当中的一个可 转录克隆基因的正链DNA,另一个则可转录负链DNA。 (x)可以直接对克隆的DNA片段进行核着酸序列测定,免去了从质粒载体到噬菌体的这一烦琐的亚克隆步骤。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2、替换型载体(取代型载体)
(一)插入式载体
• λ噬菌体载体相对于质粒载体来插入失活
•如λgt10、λNM1149等载体,在cI基因上有EcoR I及
Hind III的酶切位点。外源基因插入后将导致cI基因的
失活。cI基因失活后将导致噬菌体不能溶源化,产生 清晰的噬菌斑。相反,产生混浊的噬菌斑。
基 因 red 和 gam 位 于 非 必 需 区 域 内 , 外 源 DNA片段能置换基因red和gam形成重组体, 其重组体不能在recA-的P2溶源菌中繁殖
可以在recA+ 的P2溶源菌中繁殖并且显示出 Spi-的表型。因此只要选用recA+的P2溶源菌 作为宿主菌,就可以对重组体进行筛选。
阻遏物被一种蛋白酶切割,失活,使噬菌
体转录。
需要删除酶将噬菌体的DNA从大肠杆菌染
色体上删除下来,使之形成环形的DNA分
子,恢复溶源周期开始时的状态。
Hif
整合酶必须首先与宿主编码的宿主整合因子(host integration factor,Hif)结合形 成复合体,才能催化 attP和 attB在O区进行重组而形成原噬菌体。
cI
LA 32.7
λgt 10
EcoR I
RA10.6
λgt 10
•
Lac Z基因插入失活:如charon16A,λgt 11载 体,在非必需区段引入lac Z基因,在lac Z基 因上有EcoRⅠ位点,插入失活后利用X-gal法 筛选(兰白筛选)。
lac Z
LA 19.9
EcoR I
RA21.9
如 果 这 个 具 有 supE 基 因 的 EcoRⅠ 片 段 被 外 源
DNA取代了,那么所形成的重组体噬菌体,在
上述指示培养基上只能产生出无色的噬菌斑。
• 替换型载体克隆外源DNA的步骤
1、应用适当的核酸内切酶消化λ载体,除去可 取代的DNA片段; 2、将上述所得的 λDNA载体臂同外源DNA片 段的连接; 3、对重组体的λDNA分子进行包装和增殖, 以得到有感染性的λ重组噬菌体
第五章 噬菌体载体和 柯斯载体
本章主要内容
噬菌体一般特性 λ噬菌体 柯斯质粒 M13噬菌体 1、M13噬菌体特性及基因组结构 2、M13噬菌体载体类型及特点 噬菌粒
比较
第一节 噬菌体的一般生物学特性
(Bacteriophage,phage)
• 噬菌体可以在脱离寄主 细胞的状态下保持生命, 但一旦脱离了寄主细胞, 就不能生长和复制 • 利用寄主的核糖体、蛋 白质合成因子、氨基酸 和产能体系,进行生长 和繁殖
λ 取代载体
2、组装 1. 连接
3、侵染
(三)凯伦噬菌体载体
• 在λ噬菌体基础上发展起来的 • 承受外源DNA的能力:几个kb到23kb
(左右臂连接)
(三段自身连接)
(插入片段)
三、 λ噬菌体载体的改良
改良的目标:
扩大克隆容量 设计可对重组体分子作正选择的克隆载体 构建可以方便的通过转录作用制备外源 DNA插入序列之RNA探针的克隆载体 发展可以使插入的真核cDNA与β-半乳糖 苷酶形成融合蛋白的克隆载体
•λ 噬 菌 体 分 子 量 31×106dal,中等大 小的温和噬菌体,目 前已经定位61个基因, 其中一半参与了生命 周期,为必要基因; 余下的为非必要基因。
溶菌阶段
(复制和释放)
溶源阶段
(整合寄主染色体)
一、λ噬菌体的分子生物学概述
1、基因组结构
λ phage
(1)cos位点
• 线性双链DNA分子两端各有一条12nt组成的
菌周期,称为溶源性诱发。
① 超敏感免疫性
阻遏-操纵体系:cⅠ基因编码阻遏蛋白,启 动子PL和PR,操纵基因OL和OR
cⅠ PL OL
OR
PR
• 阻遏蛋白同操纵基因结合使RNA聚合酶不能起 始转录,这种状态有充分的能力使原噬菌体保 持“关闭”形式,溶源细菌能够正常生长。
② 溶源噬菌体的诱发
依赖于阻遏基因与操纵基因的相互作用。
recA+的P2溶源菌
red gam 导入
red gam P2 λ +
λ
无噬菌斑
外源DNA 取代 导入 λ
-
P2
滚环复制
λDNA 的 复 制 是 由 θ 型 向 滚 环 型 的 转 变 , 受 到 gam基因控制 基因gam功能:蛋白产物和寄主细胞的外切核酸 酶Ⅴ(宿主recB和recC基因产物)结合成复合物, 失去对λDNA的作用活性,因而λ噬菌体能够合 成成熟的多联体DNA,供作包装底物
四、噬菌体的溶源生命周期
溶源生长周期:是指在感染过程中没有产 生出子代噬菌体颗粒,噬菌体的DNA是整 合到寄主细胞染色体DNA上,成为它的一 个组成部分。 现知道只有双链DNA的噬菌体才具有溶源 周期
1、概念
温和噬菌体:既能进入溶菌生命周期又能进
入溶源生命周期。
溶源性细菌:具有一种完整的噬菌体基因组 的细菌 溶源化:用温和噬菌体感染细菌培养物使之 形成溶源性细菌的过程
• Charon 4A载体是用Lac 5(乳糖操纵子的大部分系 列,包括完整的Lac Z)替换入噬菌体的中间区段,同 时将Lac 5作为选择标记 •使用时用EcoRI水解,去掉中间的片段,再与欲克隆 片段在体外进行重组、包装。而后,感染E.coli使之在 E.coli内繁殖,并裂解E.coli,形成空斑 Charon 4A Lac 5
att:提供整合酶的识别位点
③ 控制基因 • cIII、N、cI、cro、cII ④ 溶菌基因——S、R
Protein coat
DNA
λ phage
Long (left) arm short (right) arm
cos
12bp
Nonessential region
cos
Exogenous DNA (~20-23 kb)
整合:噬菌体的DNA是被包容在寄主细胞 染色体DNA中 原噬菌体:在溶源细菌内存在的整合的或 非整合的噬菌体 超感染免疫性:溶源性细菌不能够被头一 次感染并使之溶源化的同种噬菌体再感染。
2、溶源周期的主要特征
λ噬菌体和P1噬菌体
λ噬菌体的特征:
(1) 噬菌体的DNA分子注入细菌细胞
(2) 经过短暂的转录之后,需要合成一种整合酶,于 是转录活性便被一种阻遏物所关闭 (3) 噬菌体的DNA分子插入到细菌染色体基因组DNA 上,变成原噬菌体
charon16A
(二)λ 替换型载体(取代型载体)
eg: EMBL3, DASH
• 外源DNA取代噬菌体染色体中对于噬菌体的 感染和复制非必要的片段 (~ 20 kb) • 高感染效率 (109 转化株/ug 载体 DNA,比质 粒高100-倍) • 替换型噬菌体λ是使用最广泛的载体。
• Charon 4A载体
短片段 重复替换区
LA 19.2
MCS
MCS
RA 9.6
•λNM781
在λNM781替换型载体中,可取代的EcoRⅠ片段, 编码有一个supE基因(大肠杆菌突变体tRNA基
因),这种λNM781噬菌体感染细胞后,寄主细
胞lacZ基因的琥珀突变被抑制,因此能在X-gal琼
脂培养基上产生出蓝色的噬菌斑。
(4) 细菌继续生长、增值,噬菌体的基因作为细菌染 色体的一部分进行复制。
噬菌体P1基本特征:
不存在噬菌体DNA分子的整合作用体系,而
是变成了一种进行独立复制的环形的质粒
DNA分子。
3、超感染免疫性
(1)溶源细菌的两个重要特点:
不能被头一次感染的噬菌体再次感染,称为
超感染免疫性;
经过许多世代以后,溶源性细菌可以进入溶
EcoR I
• λEMBL 3/ 4、Charon40
• Charon 40的中间可取代区段是一种DNA短片段 多次重复而成,称为多节段区,两个短片段之间可 被NaeⅠ识别,因此用它做载体可以有效的将其中 的多节段区清除,从而得到的多是重组的噬菌体。 克隆能力达到9~22kb Charon 40
彼此完全互补的5’突出单链
• 注入宿主后,粘性末端互补形成双链区,成 为cos位点。
cos位点的两个作用:
使进入细菌的DNA 分子成为环状 ,这 是插入细菌基因组 的先决条件。
作为识别位点,由 内切酶将滚环复制 的多联体DNA切开
The phage cos ends
5‘-CGGGGCGGCGACCTCG-3’ 3’-GCCCCGCCGCTGGAGC-5’ Circular form
一、 噬菌体的结构和其核酸类型 结构:无尾部结构的二十面体型、
具尾部结构的二十面体型、
线状体型 核酸类型:最常见的是双链线性DNA、 双链环形DNA、 单链环形DNA、 单链线形DNA、 单链RNA。
不同的噬菌体之间的核酸分子量的差异也是
比较大的。大分子量的噬菌体生命周期比较
复杂;对寄主的依赖性较低
Cleavage Ligation (during packaging)
(after infection)
GGGCGGGCGACCTCG-3’ 5’-CG + GC-5’ 3’-GCCCCGCCGCTGGA Linear form
(2)基因组
右侧区 N右侧
左侧区 A~J
中间区 J~N
λ噬菌体的基因组结构
二、λ噬菌体载体的主要类型
1、插入式载体
一种只具有一个可供外源DNA插入的克隆位点的 派生载体。只能插入较小分子量的外源(< 10kb),广泛用于cDNA及小片段DNA的克隆 具有成对的克隆位点,在这两个位点之间的 λDNA区段可以被外源插入的DNA片段所取代。 可以克隆较大分子量的外源,克隆高等真核生物 DNA