第十一章 核酸的物理化学性质
核酸的理化性质

线
尿嘧啶核苷酸
pK1 = 1.0 第一磷酸基
pK3 = 6.4 第二磷酸基
烯醇式羟基
21
pH
由于核苷酸含磷酸与碱基,为两性电解质,它们在不 同pH的溶液中解离程度不同,在一定条件下可形成兼性离 子。
在第一磷酸基和含N环解离曲线的交叉处,带负电荷的 磷酸基与带正电荷的含N环数目相等,此时pH即为核苷酸 的等电点:
完全水解 完全水解
嘧啶碱回收率高
6
二、碱水解
RNA的磷酸酯键对碱敏感、因此RNA易被碱水解, 产生核苷酸。 在室温,0.3~1mol/L KOH,24h,就可将RNA完全水 解,得到2′-或3′-核苷酸的混合物。
DNA无2′-OH,因此对碱有一定抗性:
生理意义: DNA更稳定 ,是遗传信息的载体。 RNA是DNA的信使,完成任务后迅速降解。
3.链球菌脱氧核糖核酸酶类
是一个内切酶,作用于DNA,产物为5’磷酸为末端的碎片,长度 不一,最适PH为7,需镁离子参与。
Dase只作用于DNA 12
4.限制性内切酶:
在细菌中发现有这类酶,主要降解外源DNA,第一个发现的限制 性内切酶是从大肠杆菌(E.coli.)中发现的(1968年)。
限制性内切酶的命名(以EcoR I 为例):
7
三、酶水解
(一)核酸酶的分类
①按底物专一性分类:
RNase(核糖核酸酶) DNase(脱氧核糖核酸酶)
核酸内切酶 ②按对底物作用方式分类: 核酸外切酶
小球菌核酸酶:内、外切均可 ③按磷酸二酯键断裂方式分类:
3′-OH与磷酸基之间断裂 如 蛇毒磷酸二酯酶
5′-OH与磷酸基之间断裂 如 牛脾磷酸二酯酶
磷酸基(含两个羟基)的解离
核酸的重要理化性质

RNA本身只有局部的双螺旋区,所以变性行 为所引起的性质变化没有DNA那样明显。 利用紫外吸收的变化,可以检测核酸变性的 情况。 因为天然状态的DNA在完全变性后,紫外吸 收(260 nm)值增加25-40%.而RNA变性后, 约增加1.1%。 增色效应:变性后DNA对260nm紫外光的吸 收率(A260)比变性前明显增加,这种现象 称为增色效应.
2.DNA变性后的表 现
A260值增加 粘度下降 浮力密度增大 分子量不变
3.DNA的热变性和解链温度(Fra bibliotekm)
用加热的方法使DNA变性叫做热变性 DNA的变性过程是突变性的,它在很窄的温度 区间内完成。因此,通常将DNA的变性达到 50%时,即增色效应达到一半时的温度称为 DNA的解链温度(melting temperature,Tm),Tm也称熔解温度或DNA 的熔点。
DNA复性
5、核酸的杂交(hybridization)
热变性的DNA单链,在复性时并不一定与同源 DNA互补链形成双螺旋结构,它也可以与在某 些区域有互补序列的异源DNA单链形成双螺旋 结构。 这样形成的新分子称为杂交DNA分子。DNA单 链与互补的RNA链之间也可以发生杂交。 核酸的杂交在分子生物学和遗传学的研究中具有 重要意义。
核 酸 的 杂 交
核酸杂交的应用
Southern blotting(Southern 印迹) Northern blotting(Northern 印迹) Western blotting(Western 印迹)
三、核酸的水解
1.核酸的酸解和碱解
核酸分子中的磷酸二酯键可在酸或碱性条件下水解切断 (降解)。 酸对核酸的作用因酸的浓度、温度和作用时间不同而不 同。嘌呤碱基比嘧啶碱基易被水解下来。 DNA和RNA对碱的耐受程度有很大差别。例如,在 0.3-1 mol/L NaOH溶液中,在室温至370C条件下 RNA几乎可以完全水解,生成2′-或3′-磷酸核苷; DNA在同样条件下则不受影响,若加温至1000C,4个 小时也可得到小分子的寡聚脱氧核苷酸。这种水解性能 上的差别,与RNA核糖基上2′-OH的羟基参与作用有 很大的关系。在RNA水解时,2′-OH首先进攻磷酸基, 在断开磷酯键的同时形成环状磷酸二酯,再在碱的作用
核酸的理化性质PPT

由于磷酸基团的酸性很强,所以pI较低 ,整个分子相当于多元酸。
利用核酸的两性解离可以通过调节核酸溶
液的pH来沉淀核酸,也可通过电泳分离纯化核酸
。
2020/4/10
5
三. 紫外吸收性质
嘌呤和嘧啶具有共轭双键,能强烈吸 收紫外光。在260nm处有最大吸收峰。对于纯 的DNA或RNA,可以通过测得A260来推测其核 酸含量。
核酸的理化性质
2020/4/10
1
一.一般的物理性质
1. 形 态
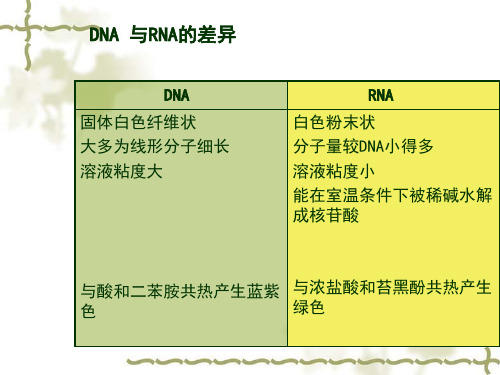
➢ DNA —— 白色纤维状固体 ➢ RNA —— 白色粉末状固体
2020/4/10
2
一.一般的物理性质
2. 溶 解性
➢ 微溶于水
➢ 不溶于乙醇、乙醚和氯仿等一般的有机溶剂
➢ RNA核蛋白体(RNP)易溶于0.14mol/L的NaCl溶液
这是由于变性的DNA双螺旋解体,藏于螺旋内 部的碱基暴露出来。
增色效应常可用来衡量 DNA变性的程度。
2020/4/10
9
4. 热变性曲线(熔解曲线)
(一) 变 性
在DNA发生热变性的过程中,A260随温度的变化曲线。
变性百分率 A260
不 同 DNA 的 熔 解
100
曲线不同,但很
类似。都是 —
A260/ A280值可以反映核酸的纯度。
纯的DNA:A260/ A280 =1.8 纯的RNA:A260/ A280 =2.0
2020/4/10
6
(一) 变 性
1. 变性的概念
四.变性与复性
核酸在某些物理或化学因素的作用下,其空 间结构发生改变,从而引起理化性质的改变及生 物活性的降低或丧失。
A260值升高 粘度下降
核酸的性质

核苷酸的紫外摩尔吸 收值最高
单链核酸紫外摩尔吸 收值其次
双螺旋核酸紫外摩尔 吸收值最低(双螺旋 结构使碱基对的p电子 云发生重叠而减少了 紫外光吸收)
核酸的变性、复性和杂交
DNA的热变性(熔解)过程是突变性的,它在很 窄的温度区间内完成。
将紫外吸收的增加量达最大量一半时的温度称熔解 温度,用Tm表示。 一般DNA的Tm值在70-85C之间。DNA的Tm值 与分子中的G和C的含量有关 G和C的含量高,Tm值高。测定Tm值,可反映 DNA分子中G, C含量,可通过经验公式计算: (G+C)%=(Tm-69.3)*2.44
核酸的物理化学性质
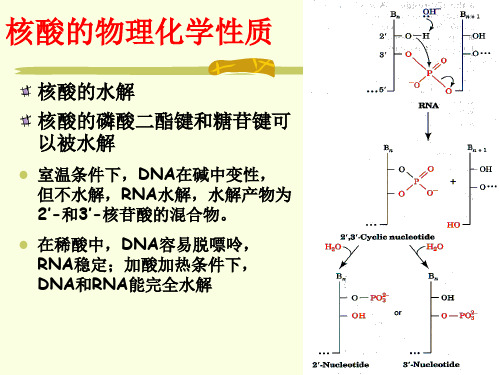
核酸的水解 核酸的磷酸二酯键和糖苷键可 以被水解
室温条件下,DNA在碱中变性, 但不水解,RNA水解,水解产物为 2’-和3’-核苷酸的混合物。
在稀酸中,DNA容易脱嘌呤, RNA稳定;加酸加热条件下, DNA和RNA能完全水解
核酸的酶水解
核酸水解酶的分类 根据底物:
核糖核酸酶(ribonuclease, RNase) 脱氧核糖核酸酶(deoxyribonuclease, DNase)
将热变性的DNA骤然冷却至低温时,DNA不 能复性(淬火)。变性的DNA缓慢冷却时可复 性,称为“退火” 影响复性的因素: DNA的浓度 ,DNA片段 的大小,温度,片段复杂性(重复序列数目)
复性的速度以Cot1/2来衡量,指复性完成一半 时的Cot值(Co是DNA的浓度,t是时间)
核酸的杂交
指不同来源的单链核酸之间可通过碱基互补形 成双螺旋结构 利用核酸杂交可检测特定的核苷酸片段或研究 同源性等 Southern 杂交(Southern bolting): DNA-DNA Northern 杂交(Northern bolting): DNA-RNA
核酸的理化性质
限制性内切酶
原核生物(及病毒)中存在着一类能识别外源DNA双螺旋中 4-8个碱基对所组成的特异的具有二重旋转对称性的回文序列, 并在此序列的某位点水解DNA双螺旋链,产生粘性末端或平末 端,这类酶称为限制性内切酶(ristriction endonuclease)。 山东落花生花落东山 帘卷晚晴天,天晴晚卷帘 Was it a cat I saw?
二、核酸的酸碱性质
1、碱基的解离
2、核苷的解离
3、核苷酸的解离
结合及释放 质子的能力
第 1、碱基的解离 14 章 碱基:含氮碱基,杂环化合物(很多生物碱的结构) 核 1)、具有芳香环的结构特征,呈平面或近乎平面 酸 含有共轭双键体系,紫外区有吸收(260nm)。 的 物 2)、氮原子位于环上或取代氨基上 弱碱性来自于环上氮原子,pKa约9.5,倾向于接受H 理 取代氨基(或曰碱基环外的氨基)碱性很弱,生理 化 条件下不能被质子化。 学 性 质
核酸的物理化学性质
一、核酸的水解 二、核酸的酸碱性质 三、核酸的紫外吸收性质 四、核酸的变性、复性及杂交
一、核酸的水解
(一)酸水解 糖苷键比磷酸二酯键 更易被酸水解 嘌呤碱基的糖苷键比 嘧啶碱基的糖苷键对 酸更不稳定
NH 2
酯键
O
N N O
N 9 N
5' HO P O CH2 O
-
糖苷
1' H H 2' OH
H
H OH
腺苷酸
(二)碱水解 RNA的磷酸酯键易被碱 水解,产生核苷酸。 由于RNA的核糖上有2’OH基,在碱作用下形成 磷酸三酯。 磷酸三酯极不稳定,随 即水解产生,产生 2’,3’-环磷酸酯,再水 解成2’-核苷酸及3’-核 苷酸
核酸的性质
DNA的核苷酸序列测定
DNA的测序策略
DNA片断的序列测定
英国Sanger , 1975年加减法,1977年末端 终止法,目前广泛用于DNA的自动测序 美国Maxam和Gilbert , 1977年化学断裂法 基本原理:把DNA变成在不同碱基的核苷酸处 打断的四套末端标记的DNA片断,当相应于四 个不同碱基产生的四套DNA片断并排进行电泳 分离时,产生一个可以直接读出DNA顺序的区 带。
模板DNA的变性:双链DNA解离,使之成为单链,以便 它与引物结合 模板DNA与引物的退火(复性):模板DNA经加热变性 成单链后,温度降至55℃左右,引物与模板DNA单链的 互补序列配对结合(退火温度与Tm有关,温度太低非特 异扩增增加,温度过高引物和模板不易结合) 引物的延伸:DNA模板--引物结合物在Taq DNA聚合酶 的作用下,以dNTP为反应原料,靶序列为模板,按碱基 互补配对与半保留复制原理,合成一条新的与模板DNA 链互补的半保留复制链 重复循环变性--退火--延伸三过程就可获得更多的“半 保留复制链”,而且这种新链又可成为下次循环的模板
真核生物mRNA的分离:亲和层析
真核生物的mRNA有polyA的尾巴,能被oligo-dT柱 子吸附而和其他物质分离开来
真核生物mRNA的分离
核酸纯度鉴定与含量测定
根据A260/A280的比值判断核酸样品的纯度 (若样品中含有杂蛋白或苯酚,则 A260/A280比值明显降低)
纯DNA:A260/A280=1.8 纯RNA:A260/A280=2.0
核酸的物理化学性质
核酸的水解 核酸的磷酸二酯键和糖苷键可 以被水解
室温条件下,DNA在碱中变性, 但不水解,RNA水解,水解产物为 2’-和3’-核苷酸的混合物。
核酸的物理化学性质和常用的研究方法

25
七、聚合酶链反应(PCR)
2、优化反应条件 包括模板、引物、 dNTP、DNA聚 合酶和Mg2+,PCR中常用的聚合酶是 Taq DNA聚合酶 3、 选择热循环温度 PCR 过程的温度控制十分关键。 热循环温度:变性温度,退火温度(低 于引物Tm值的2-3℃),延伸温度
A:光吸收值 C:磷的摩尔浓度 L:比色杯内径 W:每升溶液中磷的重量(g)
4
一、核酸的紫外吸收
DNA的ε(P):6600 RNA的ε(P):7700-7800
核酸的ε(P)比核苷酸单体低 单链多核苷酸的ε(P)比双螺 旋多核苷酸的ε(P)要高 增色效应:核酸变性时, ε(P)增加 减色效应:当核酸复性时, ε(P)降低
第十九章
核酸的物理化学性质 和常用的研究方法
1
核酸的物理化学性质
天然DNA分子细丝状的双螺旋结构 赋予DNA一系列显著的物化特性: 极大的粘度; 易于断裂; 易形成纤维状物质; 在稀盐溶液中热变性; 熔点高 RNA分子没有如此明显的物化特性
2
一、核酸的紫外吸收
1、核酸具有紫外吸收 吸收范围:240-290 nm λmax = 260nm (定量测定) 2、根据紫外吸收判断样品纯度 不纯的样品
29
九、DNA的限制酶图谱
细菌内有二种不同功能的酶(Arber等) : 限制性内切酶:识别并水解DNA的某特 定碱基序列 修饰酶(甲基化酶):识别限制酶识别的碱 基顺序并将其甲基化 被甲基化了的DNA不会被限制酶降解, 所以细菌自身的DNA不会被自身酶降解。 而当异源DNA侵入细胞时,就会被限制酶 降解
核酸检测物理知识点总结

核酸检测物理知识点总结一、核酸的结构与性质1.1 核酸的化学结构核酸是一种由核苷酸经过磷酸二脂酸酯键连接形成的生物大分子,包括DNA和RNA两种类型。
DNA由脱氧核糖核苷酸组成,RNA由核糖核苷酸组成。
核苷酸由核苷和磷酸二脂酸组成,核苷包括一个含氮碱基和一个糖分子,磷酸二脂酸作为链的连接部分。
1.2 核酸的物理性质核酸具有许多特殊的物理性质,如双螺旋结构、碱基配对、DNA超螺旋等。
其中双螺旋结构是DNA的典型结构,由两条螺旋形成,而碱基配对是通过氢键将两条链连接在一起,碱基的配对规律是腺嘌呤(A)与胸腺嘧啶(T)之间形成两个氢键,鸟嘌呤(G)与胞嘧啶(C)之间形成三个氢键。
此外,DNA还具有超螺旋结构,这种结构形式使得DNA在细胞分裂时更容易分离。
1.3 核酸的光学性质核酸具有一定的光学性质,如吸收光谱、荧光光谱等。
DNA和RNA在紫外光下有显著的吸收,其中DNA在260nm处有最大吸收峰,而RNA在260nm处有一个稍微红移的吸收峰。
此外,核酸还具有荧光发射的性质,一些荧光染料可以与核酸结合产生荧光信号,用于核酸的检测和定量分析。
二、核酸检测的原理与技术2.1 核酸检测的原理核酸检测的原理是通过特定的技术手段来识别和检测样品中的核酸序列,常用的技术包括PCR(聚合酶链式反应)、分子杂交、核酸电泳、原位杂交等。
PCR是最常用的核酸扩增技术,通过模拟细胞内DNA复制的过程来扩增目标DNA序列,从而实现对目标基因的检测和分析。
2.2 核酸检测的技术手段核酸检测的技术手段包括一系列的实验方法和设备,如核酸提取、PCR扩增、凝胶电泳、原位杂交、微阵列技术等。
其中核酸提取是核酸检测的首要环节,其目的是从样品中提取出目标DNA或RNA序列,为后续的PCR扩增和检测做准备;PCR扩增是一种快速、高效、特异性强的核酸扩增技术,可将目标核酸的复制数量扩大上百万倍,从而实现对微量核酸的检测和分析。
2.3 核酸检测的应用核酸检测技术在临床医学、疾病预防和控制、食品安全监测等领域有着广泛的应用,如临床诊断中的传染病检测、肿瘤基因检测、遗传病筛查等;疾病预防和控制中的病毒核酸监测、病原微生物检测、环境污染监测等;食品安全监测中的食源性疾病的检测、转基因食品的检测等。
核酸性质

核酸的凝胶电泳
核酸的序列测定 DNA聚合酶链反应 DNA的化学合成
第十五章 核酸的研究方法
一 核酸的分离、提纯和定量测定
核酸制备
总的要求:防止核酸的降解和变性,要尽量保
持其在生物体内天然状态
注意的事项:条件温和,防止过酸、过碱、避
免剧烈搅拌,防止核酸酶作用
P513
第十五章 核酸的研究方法
(一)DNA的分离
Chapter14
核酸的物理化学性质
四
核酸的变性、复性及杂交
(三)、核酸的杂交
2. 常见杂交的类型
(1)Southern blotting (2)Northern blotting (3)Western blotting
(1). Southern印迹杂交
将在电泳凝胶中分离的DNA片段转移并结合在适当的 滤膜上,然后通过与标记的单链DNA或RNA探针的杂 交作用检测这些被转移的DNA片段
经验式: (G-C)%=(Tm-69.3)×2.44
(3)介质中的离子强度 一般在较高的离子强 度时,DNA的Tm值较高,而且熔解过程发生在 一个较小的温度范围之内。
P509
Chapter14
核酸的物理化学性质
四
核酸的变性、复性及杂交
(二)、复性 (renaturation)
变性DNA在适当条件下,又可使两条彼此分开的链重新缔合成为双螺 旋结构,这过程称复性 1. DNA复性的特点:
一、 核酸的水解
二、核酸的酸碱性质
三、核酸的紫外吸收
四、核酸的变性、复性及分子杂交 五、核酸的沉降特性
502
Chapter14
核酸的物理化学性质
生物化学-生化知识点_核酸的物理化学性质

7-3 核酸的物理化学性质上册P502一一一核酸的水解:所有糖苷键和磷酸酯键都能被水解。
一1一酸水解:糖苷键比磷酸二酯键易被水解,嘌呤碱糖苷键比嘧啶碱更易水解。
一2一碱水解:磷酸酯键易水解,RNA比DNA易水解,因为RNA核糖上有2‘-OH,水解过程见P502。
一3一酶水解:为水解磷酸二酯键的酶,专一水解核酸的为核酸酶。
1.核酸酶的分类:按底物专一性分为RNase(核糖核酸酶)和DNase(脱氧核糖核酸酶)。
按对底物作用方式分为内切酶(作用点在核糖核酸酶内部)和外切酶(作用点在末端)。
2.RNase:如牛胰核糖核酸酶(EC 2.7.7.16),内切酶,作用位点为嘧啶核苷(Py)-3‘-磷酸与其他核苷酸之间的连键。
3.限制性内切酶:为DNase。
剪裁DNA的工具,可用于核酸测序和基因工程。
在细菌中发现,目前已找到限制性内切酶数千种。
限制性内切酶往往与甲基化酶成对存在,自身酶作用位点的碱基被甲基化,内切酶不再降解,因而可识别和降解外源DNA。
断裂位点处常有二重旋转(轴)对称性(回文结构,正读反读相同),在特定位点两条链切断后形成粘末端或平末端。
限制性内切酶命名:如E. coRⅠ,第1个字母E(大写),为大肠杆菌(E.coli)属名的第一个字母,第2、3两个字母co(小写)为种名头两个字母,第4个字母R,表示菌株,最后一个罗马字为该细菌中已分离这一类酶的编号。
一一一核酸的酸碱性质:核苷和核苷酸都是兼性离子,碱基和磷酸基均能解离,见P505,具有酸碱性。
由于DNA酸碱变性,使酸碱滴定曲线不可逆。
一一一核酸的紫外吸收:嘌呤环与嘧啶环具有共轭双键,核苷和核酸的吸收波段在240~290nm,最大吸收值在260nm附近(蛋白质最大吸收值280nm)。
一1一可用于测样品纯度(测吸光度A):A260/A280比值,纯DNA应大于1.8,纯RNA应达到2.0,若样品混有杂蛋白,比值明显降低。
对于纯样品,从260nm的A值即可算出含量。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
盐浓度对DNA变性的影响
RNA的变性:RNA也具有螺旋 线团 之间的转变。但这种较变不如DNA明显。 变性曲线不陡, Tm值较低。tRNA具有较 多的双螺旋区,所以具有较高的Tm值, 变性曲线也较陡。
(二)复性 变性核酸的互补链在适当的条件下,重新缔合成为双 螺旋结构的过程称为复性。
核酸分子杂交(DNA/DNA or DNA/RNA)
技术
核酸分子杂交技术,是在1968年由华盛顿 卡内基学院(Cavnegie Institute of Washington) 的Roy Britten及其同事发明的。所依据的原理 是,带有互补的特定核苷酸序列的单链DNA或 RNA,当它们混合在一起时,其相应的同源区 段将会退火形成双链的结构。
<2>核酸分子杂交的技术 Southern Blotting(南印迹):用于钓基因,即用已知 的DNA单链或RNA,钓取未知DNA分子中的基因,方 法如下: 未知的DNA --DNA限制性内切酶 DNA片段 琼脂糖电泳分离 碱液变性 影印在硝酸纤维薄 膜上 与放射性标记的已知DNA单链或RNA杂交 放射自显影 Northern Blotting(北印迹):用已知的DNA钓 mRNA,方法如下: 众多未知的RNA 电泳分离 变性 影印 用标记的已知DNA单链杂交 放射自显影 Western Blotting(西印迹):蛋白质与抗体的杂交, 跟核酸无关。
四.核酸的变性,复性及杂交 (一) 变性
稳定核酸双螺旋的次级键断裂,空间结构破坏,变 成单链结构的过程。核酸的一级结构(核苷酸顺序)保 持不变。 变性表征 生物活性部分丧失、粘度下降、浮力密度升高、紫 外吸收增加(增色效应) 变性因素 pH(>11.3或<5.0) 变性剂(脲、甲酰胺、甲醛) 低离子强度 加热
(三)核酸的杂交 DNA单链与在某些区域有互补序列的异源 DNA单链或RNA链形成双螺旋结构的过程。 这样形成的新分子称为杂交DNA分子。 核酸的杂交在分子生物学和遗传学的研究中具 有重要意义。 Southern 杂交(Southern bolting) Northern 杂交(Northern bolting) Western 杂交 (Western bolting)
(a)
基因组DNA
Southern
Hale Waihona Puke DNA限制片段凝胶转移
杂交技术
(b)
( c)
(d)硝酸纤维素滤膜
同探针同源杂交 的基因DNA片段 X光底片
( e)
Southern Blotting
DNA
DNA Marker
migration
Paper towels
Gel Salt
solution
Northern RNA印迹技术(Northern blotting)
DNA复性后,一系列性质将得到恢复,但是生物活性一 般只能得到部分的恢复,具有减色效应。
将热变性的DNA骤然冷却至低温时,DNA不可能复性。 变性的DNA缓慢冷却时可复性,因此又称为“退火”。 退火温度=Tm-25℃ 复性影响因素 片段浓度/片段大小/片段复杂性(重复序列数目)/ 溶液离子强度
将热变性的DNA骤然冷却时,DNA不可能复 性 DNA的片段越大,复性越慢 DNA的浓度越大,复性越快 在一定条件下,复性反应的速度用Cot 1/2来衡 量,Co为变性DNA复性时的初始浓度,t为时 间, Cot ½表示复性一半时的DNA浓度。 两种浓度相同但来源不同的DNA,复性时间的 长短与基因组的大小有关 有很多重复序列的DNA复性也快
热变性与Tm
DNA的变性过程是突变性的,它在很窄的温度 区间内完成。因此,通常将紫外吸收的增加量 达最大量一半时的温度称熔解温度,用Tm表示。 (或者把加热变性使DNA的双螺旋结构失去一 半时的温度称为该 DNA的熔点或熔解温度)。 一般DNA的Tm值在82-95C之间。
DNA的Tm值大小与下列因素有关: (1)DNA的均一性。均质DNA,如一些病毒的 DNA,人工合成的poly d(A-T),poly d(G-C)熔 解过程发生在一个较小的范围内。异质DNA熔 解的过程发生在一个较宽的范围内。所以Tm 值可作为衡量DNA样品均一性的标准。 (2)分子中G和C的含量。 G和C的含量高,Tm值高。因而测定Tm值,可 反映DNA分子中G, C含量,可通过经验公式计 算:(G+C)%=(Tm -69.3) Χ 2.44 (3)介质的离子强度。一般离子强度较低的介 质中,DNA的熔解温度较低,而且溶解温度的 范围较宽。
核酸的杂交
<1>核酸分子杂交的意义: 发现原核生物的基因是连续基因,而真核生物 的基因是断裂基因。 连续基因:基因中的bp序列能够连续的在成熟 的蛋白质中找到其相应的AA,电镜显示这种 基因能够和它的成熟mRNA形成平滑的杂交分 子。 断裂基因:基因中的bp序列能够断续的在成熟 的蛋白质中找到其相应的AA,电镜显示这种 基因和它的成熟mRNA只能形成带泡的杂交分 子。 发现癌基因的普遍性:肿瘤病毒的RNA能够与 人类正常的DNA分子形成带泡的杂交分子。
核糖核酸酶举例 牛胰核糖核酸酶( RNase I),内切酶,水解嘧 啶3'-磷酸和其他核苷酸5 ' -OH形成的酯键,产 生3 ' -磷酸核苷。 核糖核酸酶T1,内切酶,发现于米曲霉,水解 3 ‘ -鸟苷酸与其他核酸5 ’ -OH形成的酯键, 产物为3 ‘ -鸟苷酸为末端的寡核苷酸。
脱氧核糖核酸酶举例
1.牛胰脱氧核糖核酸酶(DNase I),内切酶,水解DNA 3 ‘ -磷酸酯键,产生5 ’ -磷酸的寡核苷酸,平均长 度为4个核苷酸。 2.牛脾脱氧核糖核酸酶(DNase II),内切酶,水解DNA 5 ‘ -磷酸酯键,产生3 ’-磷酸的寡核苷酸,平均长 度为6个。 3.限制性内切酶,可识别特异的碱基序列并水解断开,产 物带5 ‘ -磷酸基,已发现数千种,是基因工程的重要 工具酶。 N-糖苷酶 水解糖苷键,产物为含氮碱基和无碱基末湍的寡核苷酸
1979年,J.C.Alwine等人发展而来,是将RNA分 子从电泳凝胶转移到硝酸纤维素滤膜或其他化学修饰 的活性滤纸上,进行核酸杂交的一种实验方法。由于 这种方法与Southern DNA印迹杂交技术十分类似,所 以叫做Northern RNA印迹技术(Northern blotting)。 将蛋白质从电泳凝胶中转移到硝酸纤维素滤膜上, 然后同放射性同位素125I标记的特定蛋白质之抗体进行 反应,这种技术叫做Western蛋白质杂交技术 (Western blotting)。
二.核酸的酸碱性质
两性解离:DNA无,只有酸解,碱基被屏蔽 (在分子内部形成了氢键)。RNA有两性解离, 有pI。核酸的碱基、核苷、核苷酸均能发生解 离,因此核酸是两性电解质 碱基的解离:嘧啶和嘌呤环上的 N及各取代基 具有结合和释放质子的能力。 核苷的解离:碱基和核糖具有结合和释放质子 的能力 核酸的解离:磷酸基具有较强的酸性,核酸的 两性解离主要决定于磷酸基和含氮碱基。
样品中如含有杂蛋白及苯酚,比值明显 降低。 对于纯的样品,只要读出260nm的A值即 可算出含量。通常以A值为1相当于 50µ g/ml 双螺旋DNA,或40 µ g/ml单链 DNA(或RNA),或20 µ g/ml寡核苷酸 计算。
有时核酸的紫外吸收以摩尔磷的吸光度来表示,摩尔 磷即相当于摩尔核苷酸。 先测定核酸溶液中的磷含量及紫外吸收值,然后求出 摩尔磷吸光系数ε(P) 。 ε(P)=A/cL A为吸收值,c为每升溶液中磷的摩尔数,L为比色杯内径。 c=每升中磷的重量(克)W/磷的原子量(30.98) ε(P)=30.98 A/WL 一般天然DNA的ε(P)为6 600,RNA为7 700-7 800。核 酸的ε(P)值较所含核苷酸单体的ε(P)要低40%-45%。 单链多核苷酸的ε(P)值比双螺旋结构多核苷酸的ε(P)要 高,所以核酸发生变性时, ε(P)升高约25%,此现象 称为增色效应(hyperchromic effect)。复性后ε(P)又 降低,这现象称减色效应(hypochromic effect)。
第十一章 核酸的物理化学性质
一.核酸的水解 核酸分子中的磷酸二酯键可在酸或碱性条件下 水解切断。 DNA和RNA对酸或碱的耐受程度有很大差别。 例如,在0.1 mol/L NaOH溶液中,RNA几乎可 以完全水解,生成2′-或3′-磷酸核苷;DNA在 同样条件下则不受影响。这种水解性能上的差 别,与RNA核糖基上2′-OH参与作用有很大的 关系。在RNA水解时,2′-OH首先进攻磷酸基, 在断开磷酯键的同时形成环状磷酸二酯,再在 碱的作用下形成水解产物。
三.核酸的紫外吸收 在核酸分子中,由于嘌呤碱 和嘧啶碱具有共轭双键体系, 因而具有独特的紫外线吸收 光谱,一般在260nm左右有 最大吸收峰,可以作为核酸 及其组分定性和定量测定的 依据。 以A260/A280进行定性、定量 DNA和RNA溶液中加入溴化 乙锭(EB),在紫外线下发 出荧光。 用A260/A280还可用来表示 核酸的纯度: 大于1.8,DNA很纯; 大于2, RNA很纯。
Southern DNA印迹杂交 根据毛细管作用的原理,使在电泳凝胶中 分离的DNA片段转移并结合在适当的滤膜上, 然后通过同位素标记的单链DNA或 RNA探针 的杂交作用检测这些被转移的DNA片段,这种 实验方法叫做DNA印迹杂交技术。由于它是由 E.Southern于1975年首先设计出来的,故又叫 Southern DNA 印迹转移技术。
酸易水解N-糖苷键,嘌呤碱基比嘧啶碱 基易被酸水解,脱氧核糖比核糖的糖苷 键易水解,DNA的嘌呤碱在pH2以下即 脱落而成嘌呤酸。 碱易水解磷酸酯键,RNA在0.3N碱中即 被水解,DNA对碱相对稳定。
酶水解
生物体内存在多种核酸水解酶。这些酶可以催化水解 多聚核苷酸链中的磷酸二酯键。 核酸酶的分类 按 底 物 专 一 性 分 : 以 DNA 为 底 物 的 DNA 水 解 酶 (DNases)和以RNA为底物的RNA水解酶(RNases)。 按作用方式分:核酸外切酶和核酸内切酶。 核酸外切酶的作用方式是从多聚核苷酸链的一端( 3′端或5′-端)开始,逐个水解切除核苷酸;核酸内切酶 的作用方式刚好和外切酶相反,它从多聚核苷酸链中 间开始,在某个位点切断磷酸二酯键。 按作用键分:水解3-磷酸酯键和5-磷酸酯键的核酸酶, 产物分别是5-磷酸核苷和3-磷酸核苷