唐定中-植物抗病信号

植物抗病信号
唐定中
一、概况
1、研究植物抗病的原因:因为它既是重要的生物学问题也是重要的农业问题。每年稻
瘟病造成水稻损失50-60亿美元。所以作物病害对农作物生产造成的危害非常严重,直接影响粮食安全。另外,病原菌还具有变异迅速,扩散迅猛。例如Vg99引起的
小麦杆锈病,在世界范围内都有扩增。
2、研究较多的几种病原菌类型主要有:细菌(如:假单胞杆菌、黄单胞杆菌)、真菌
(如:白粉菌、稻瘟病)、卵菌(如:霜霉病菌、疫霉菌)、病毒(如:烟草花叶病
毒、黄瓜花叶病毒)。这些病菌可分为两类不同的植物病原菌,即a.死体营养型或
腐生型,b.活体营养型或寄生型。
3、病菌感染植物的一般过程或条件:a.锚定/进入宿主细胞,b.抑制宿主的抗性,c.改
变宿主的生理机能,如对营养和水的需求,d.在宿主细胞内扩增,e.最终导致疾病
的发生。
4、病菌感染植物后,植物会产生两种反应:即抗病(亦称非亲和性反应,此反应中病
菌是无毒的,植物产生抗性。)和感病(亦称亲和反应,病原菌有毒,植物是敏感
型的)
5、如何确定植物的病型?答:一是测定病原菌的生长情况。对于白粉菌来说要看其孢
子数,对于细菌来说,要测定其生长曲线。二是评价植物的损伤程度。
6、植物的抗病层次主要有:非寄主抗性、基础抗性(PTI)、R基因介导的抗性(ETI)。
二、非寄主性抗性
1、大部分植物都能对大多数病菌产生抗性。多数情况下从某一种植物中分离到的病
原菌不能再次感染或引起其它植物感染,也不能繁殖。非寄主性抗性就是植物能
够抵抗大部分潜在致病菌的原因,是最常见的植物抗病方式。
2、非寄主性抗性的机理:(1)消极防御也可以说是天然的防御系统,只要是植物表
面的物理障碍(如角质层、细胞壁等)以及一些代谢产物,大部分为抗菌素。(2)
诱导型的植物抗病机制。如植物抗毒素,它是植物在受到病原菌的侵害后应答从
头合成的一种抗菌物
3、非寄主性抗性所涉及的基因:多数情况下,植物针对真菌产生的非寄主性抗性是
在其侵入的过程中进行。拟南芥的PEN1, PEN2和PEN3基因可用于对大麦白粉病
产生抗性。(PEN1是一种突触融合蛋白,它在细胞膜的囊泡运输过程中具有重要
作用。PEN2是一种糖基水解酶。PEN3是ABC转运蛋白)
4、病原菌可通过多种方式破坏植物的非寄主性抗性系统。从病原菌进入植物的方式
看,可通过植物体表已有的缺口进入,如气孔、伤口;也可痛过分解酶分解角质;
或是通过纤维素酶、果胶酶、纤维素内切酶等破坏细胞壁。
三、基础抗性(PAMP triggered immunity, PTI)
PAMPs:病原相关的分子特征。MAMPs:细菌相关分子特征
PAMPs对于病原菌的生活方式来说是非常重要的,它在大部分微生物中都高度保守,但在宿主中却不存在。植物通过识别PAMPs来激活其基础抗性。
PAMPs主要有:鞭毛蛋白、脂多糖、几丁质以及麦角脂醇等。
1、PAMP的识别及基础免疫
人们对植物对PAMPs的应答的认识大部分来自于鞭毛蛋白。鞭毛蛋白是细菌运动的
重要组成部分,其N端和C端都高度保守,这使其成为非常重要的PAMP,且其不
存在于植物中。
鞭毛蛋白的识别及下游调控:在拟南芥的研究中,鞭毛蛋白氨基末端高度保守的22个氨基小肽(flg22)足以用于与受体的结合。其下游调控途径为FLS2(RLK)(鞭毛蛋白的受体蛋白)—MAP激酶的级联反应—WRKY转录因子
植物的基础免疫应答:愈创葡萄糖的沉积作用,使细胞壁加厚;产生活性氧杀死病菌;激活抗性基因的转录;激活MAP激酶
那么病原菌又是如何进一步感染植物的呢?主要是有些病原菌具有抑制或逃避基
础性抗性的能力,如修饰PAMPs,根癌农杆菌能够修饰鞭毛蛋白,使其不能被FLS2受体识别,或者病原菌可将某些效应分子传递给植物细胞,抑制植物的基础免疫
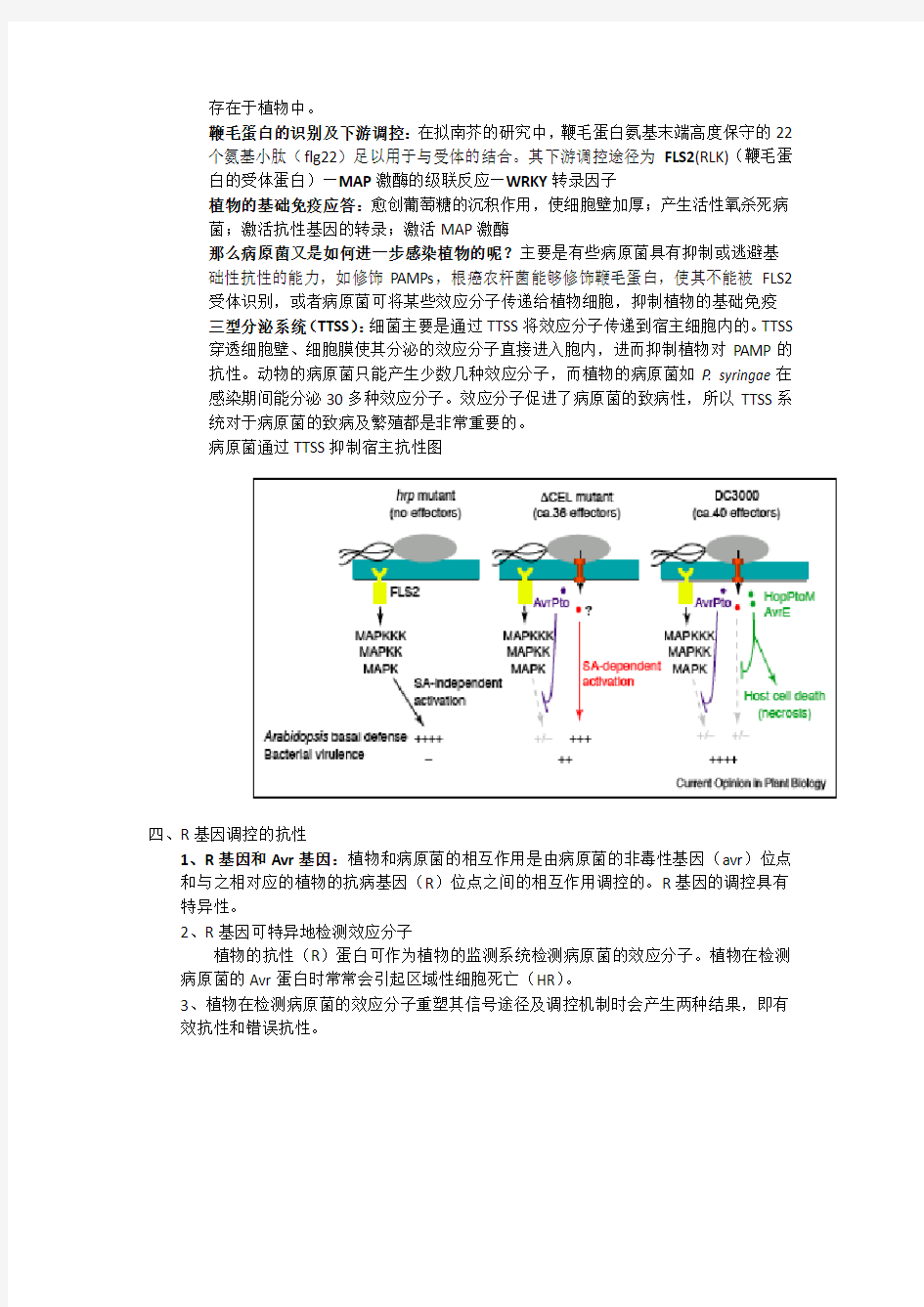
三型分泌系统(TTSS):细菌主要是通过TTSS将效应分子传递到宿主细胞内的。TTSS 穿透细胞壁、细胞膜使其分泌的效应分子直接进入胞内,进而抑制植物对PAMP的抗性。动物的病原菌只能产生少数几种效应分子,而植物的病原菌如P. syringae在感染期间能分泌30多种效应分子。效应分子促进了病原菌的致病性,所以TTSS系统对于病原菌的致病及繁殖都是非常重要的。
病原菌通过TTSS抑制宿主抗性图
四、R基因调控的抗性
1、R基因和Avr基因:植物和病原菌的相互作用是由病原菌的非毒性基因(avr)位点
和与之相对应的植物的抗病基因(R)位点之间的相互作用调控的。R基因的调控具有特异性。
2、R基因可特异地检测效应分子
植物的抗性(R)蛋白可作为植物的监测系统检测病原菌的效应分子。植物在检测病原菌的Avr蛋白时常常会引起区域性细胞死亡(HR)。
3、植物在检测病原菌的效应分子重塑其信号途径及调控机制时会产生两种结果,即有
效抗性和错误抗性。
4、R基因的分类
NBS-LRR(Nucleotide Binding Leucine Rich Repeats)是目前所克隆的抗性基因中最大的家族。其中LRR的长度为20-30个氨基酸。根据其N端的不同又可进一步分为(CC)-NBS-LRR和(TIR)-NBS-LRR
5、NBS-LRR蛋白的不同区域的作用:
其N端结构域用于与下游调控分子的结合,NBS结构域含有一些保守的区,如用于核苷酸结合的P-loop。LRR作为效应分子的结合区和调控区。
R基因识别效应分子激活抗性。
6、R蛋白通过两种不同的机制结合效应蛋白。直接结合,R蛋白作为受体直接与效应蛋白结合;间接结合(Guard假说)
直接结合的证据:水稻的Pi-ta LRR与AVR-Pita的直接结合,拟南芥的RRS1直接与PopP2相互作用,亚麻的L抗性区直接与AvrL相互作用。
间接结合的证据:
(1)拟南芥的NBS-LRR蛋白RPM1既可以结合Pseudomonas syringae的AvrRpm1,又可
结合AvrB,这两个蛋白又都能与RIN4结合,且调控RIN4的功能。只要它们两个中
有一个存在就能调控RIN4的磷酸化。
(2)拟南芥的NBS-LRR蛋白PRS2可检测Pseudomonas syringae的AvrRpt2,但它们之间
不能直接结合,PRS2与RIN4结合,RIN4再与AvrRpt2结合。AvrRpt2是一个含半胱
氨酸的蛋白酶,可降解RIN4
(3)拟南芥的NBS-LRR蛋白PRS5可检测Pseudomonas syringae的AvrRpB。但它们之间
也不是直接作用的饿,PRS5结合PBS1,PBS1再结合AvrRpB。AvrRpB也是一个半胱
氨酸蛋白酶,在特定位点切割PBS1。所以,应该是PBS5通过调控PBS1的状态来结
合病菌的效应蛋白AvrRpB。
Guard假说:R蛋白用于调控(保卫)效应蛋白的靶标(被保护者),而效应蛋白对其靶标的修饰可激活R蛋白,使的R蛋白启动抗病机制。
7、植物和病原菌的进化过程:开始时病原菌无鞭毛,植物产生非宿主性抗性病原
菌就进化出鞭毛,使植物致病进而使植物进化出鞭毛蛋白的受体FLS2,产
生基础抗性病原菌再次进化出三型分泌系统,使植物致病植物再进化
出R基因,产生R基因介导的抗性病原菌再产生可抑制R基因的物质,再
次使植物感病植物则进化出R基因2,可间接抑制R基因的物质,同时作用
于下游调控,产生抗性。
五、R-Avr下游调控
1、R蛋白抗性调控所需基因:
从突变体中筛选对无毒病菌失去抗性的突变体,得到PBS1(RPS5特异性需要的蛋白激酶)、RIN4(R蛋白RPM1所需的一种蛋白)、RCR3(R蛋白Cf-2所需
蛋白)。
2、不同R基因调控的抗性基因进行筛选:
从对不同无毒菌株失去抗性的突变子中筛选:NDR1、RAR1、SGT1、EDS1和PAD4、NPR1
3、系统性获得抗性(SAR):受过感染的植物在受到同样的或相关的病原菌侵害时
会产生抗性。SAR的获得需要SA(水杨酸)。
六、抗性信号网
植物抵抗病原菌进行抗性调控的3个必需的信号分子:SA、JA、乙烯(Ethylene)
1、SA信号
SA是多数植物抗性应答的重要分子。
NahG:细菌水杨酸羟化酶基因SID2:异分支酸合成酶1基因
2、JA(茉莉酸)
JA常出现在土豆的诱导型损伤应答,JA和SA分别诱导不同系列的抗性基因
3、乙烯途径
SA途径主要用于针对活体营养型病菌产生抗性,JA/乙烯主要针对死体营养型病菌产生抗性
4、JA/乙烯途径与SA途径是拮抗的
JA/乙烯途径与SA途径是拮抗的,在假单胞菌感染植物的过程中,其产生的JA类似物冠菌素(COl1)可通过抑制SA调控的应答来致病。
5、MAPK激酶信号通路参与调控植物抗病反应
七、小结
抗性和感性、区域性细胞死亡(HR)、活体寄生型和死体寄生型、非宿主抗性、PAMPs、基础免疫(PTI)、TTSS及其效应分子、R基因和Avr基因、R基因调控的抗性及ETI、模式基因、Guard假说、植物与病原菌相互关系的进化、SAR、SA或JA以及乙烯途径。
八、唐定中课题组的研究
一些重要的未回答的问题:抗性应答的信号途径中仍有多种成分未知,植物应答时的细胞活动情况仍不了解。
研究方法:用遗传学方法鉴定抗性调控基因。分离能够增强抗性的突变体以鉴定抗性应答过程中的负调控因子;分离抑制抗性的突变子鉴定正调控因子。
利用拟南芥和白粉菌研究植物抗病的分子机制
edr1和edr2 突变体能够增强对白粉病的抗性。edr2对白粉病的调控依赖于SA。EDR2蛋白含有PH(Pleckstrin Homology普列克底物蛋白)区和START区(StAR (Steroidogenic Acute Regulatory protein)-related lipid transfer)。在体内实验中EDR2 PH 区结合到磷脂酰肌醇-4-磷酸上。对EDR2进行体内定位实验,发现其在内质网膜、细胞质膜、核内体上。
EDR2的作用模型:
Edts可抑制edr2,换句话说可促进植物的抗性应答。所以植物抗病反应中的重要基因主要有:EDTS1/BSK1(胞质型受体激酶)、EDTS2/SR1(转录因子)、EDTS3/RPN1a(蛋白降解途径)、EDTS5/ALD1(氨基转移酶)、EDTS6(未知功能的蛋白)
bsk1突变可抑制edr1, mlo2和对白粉菌的抗性。BSK1基因互补了bsk1-1突变体表型,BSK1同源基因的突变不抑制edr2对白粉菌的抗性
BSK1 是油菜素内脂(brassinosteroid,BR)受体BRI1的底物
bsk1 单突变体对白粉菌更感病
BSK1参与对细菌Pto DC3000和卵菌H. a. Noco2的抗性
bsk1积累较低的水杨酸(SA )
BSK1 定位在质膜上,膜定位对其功能很重要
BSK1 具有激酶活性,激酶活性对BSK1 的功能不可缺少
BSK1 与鞭毛蛋白的受体FLS2互作
bsk1突变不影响BSK1与FLS2的互作
bsk1突变表现flg22诱导的活性氧迸发的缺陷(ROS burst )
bsk1中flg22诱导的PR1表达低于野生型
bsk1 突变抑制edr2的白粉病抗性
BSK1 参与基础抗性和R基因介导的抗性
BSK1 是一个胞质型类受体激酶, 是BR 受体BRI1的底物
BSK1 与PAMP 受体FLS2互作.
BSK1 参与FLS2介导的免疫反应。
BSK1作用的分子机理
edts2抑制edr2调控的白粉病抗性,它编码SR1(一种结合转录因子的钙调蛋白)SR1 结合到NDR1和EIN3启动子上。
edts3抑制edr2调控的白粉病抗性
RPN1是26S蛋白酶体的一个亚基
EDOS1/HPR1 is required in edr1-mediated resistance
HPR1 is involved in mRNA transport
Screen additional edr1 and edr2 like mutants
EDR4未知功能,与类受体激酶互作
EDR5与植保素合成有关
EDR6/ATG2与细胞自噬有关
EDR7与囊泡运输有关
edr4 displays enhanced disease resistance to G. cichoracearum。EDR4 is novel protein, and it interacts with two receptor like kinases, may function in vesicle
edr6 is resistant to powdery mildew
EDR6 encodes ATG2, an autophagy related protein
atg2 shows defects in autophagosome accumulation
总之,多种细胞过程参与植物抗病:囊泡运输、自噬作用、蛋白降解、蛋白磷酸化
植物抗病基因在进化中形成了几种共有的进化形式
植物抗病基因在进化中形成了几种共有的进化形式。植物祖先抗病基因的复制创造了新基因座。基因间和基因内重组导致了变异,也导致了新特异性抗病基因的产生。另外,与特异性识别相关的富含亮氨酸重复区顺应于适应性选择。同样,类转座元件在抗病基因座中的插入加速了抗病基因的进化。随着抗病基因的进化,抗病反应也呈现出多样化,代表着植物与病原物动态进化的不同阶段。 几种抗病基因进化模式得到提出。重复拷贝对创造新的抗病基因起着重要的作用。抗病基因的复制与随后序列的差异性能创造或扩大基因家族中另一基因簇。不对等重组与基因转化(基因内)创造了基因数量上的多样性。基因外重组与基因转化能创造新的特异性抗病基因。而有的这些重组事件发生在高保守区域上。LRR区域的多态性为识别、配位及防卫大量病原物提供了进化优势。转座元件插入到某些抗病基因座中造成基因断裂或染色体重排,加速了抗病基因的进化。基因座内的过多重组将导致抗病基因特异性丧失,而寄主植物与病原物不断相互作用——双方相互施加压力并不断适应与反适应于选择压力,进行着协同进化,那么抗病基因就必须维持着序列的特异性。实际上,抗病基因的进化是基因变异与基因序列保守性之间的平衡。在抗病基因不断进化的推动下,抗病基因控制下的抗病反应表现出多样化(如过敏性反应、非过敏性反应、系统过敏性反应以及极度抗病等),不同类型的抗病反应代表着植物与病原物动态进化的不同阶段。有关与抗病基因的进化研究还存在一定困难,涂礼莉等人借助其他物种已获得的信息,利用生物信息学的方法来研究海岛棉抗病基因的抗病机制及抗病基因进化。这种研究方法可能也适用于其他农作物,可以说对抗病机制的研究、抗病基因的转育及抗病基因进化的研究具有重要的意义。 近年来抗性基因研究的突破性进展、抗性基因的克隆和序列分析所揭示的其编码蛋白的组 成、拓扑学和亚细胞定位等特征,为揭开抗性基因的作用特点提供了线索。一般来讲,基因克 隆的策略可分为两种:正向遗传学途径和反向遗传学途径。前者以欲克隆的基因所表现的功 能为基础,通过鉴定其产物或某种表型的突变进行,如功能克隆( Functional Cloning) 和表 型克隆( Phenotype Cloning) ;后者则着眼于基因本身特定序列或者在基因组中的特定位 置进行,如定位克隆( Positional Cloning) 和序列克隆( Sequence Cloning) 。
植物与病原菌互作和抗病性的分子机制
中国农业科学 1999,32(增刊):94~102 Scientia A gricultrua Sinica 植物与病原菌互作和抗病性的分子机制3 刘胜毅1 许泽永1 何礼远2 (1中国农业科学院油料作物研究所,武汉 430062;2中国农业科学院植物保护研究所) 提要 概述了近几年在寄主植物抗病基因与防卫反应基因、病原菌毒性基因、寄主抗病性机制和抗病基因工程策略等方面取得的主要进展,重点分析了抗病反应的一般过程、毒性基因 产物胞外水解酶和毒素的作用与关系、作物抗毒素基因工程策略。 关键词 植物;抗病基因;防卫基因;毒性基因;基因工程策略 早在40年代末50年代初,F lo r(1947;1955)在对亚麻和亚麻锈菌互作的遗传规律研究中,提出了基因对基因假说(gene2fo r2gene hypo thesis)〔4,5〕,这标志着对植物与病原菌互作的认识深入到了基因水平,从而为应用分子生物学手段研究植物抗病性奠定了基础。本文概要地综述近几年在寄主植物抗病基因、病原菌致病基因、寄主抗病机制等方面取得的主要进展,并试图侧重分析概括抗病反应的一般过程及毒素的作用与基因工程策略。 1 抗病相关基因 根据基因的作用性质,可把抗病反应过程中起作用的基因分为两类:抗病基因和防卫反应基因。抗病基因是决定寄主植物对病原菌的专化性识别,并激发抗病反应的基因。即按F lo r的基因对基因理论,它与病原菌的无毒基因互补;按Keen(1990)提出的用来解释基因对基因理论分子机制的配体2受体模型〔6〕,它的产物是抗病反应信号传导链的起始组分,即信息链的前端,当它与病原菌的无毒基因直接或间接编码产物互补结合后,启动信号传导激发植物的抗病反应。防卫反应基因是一类在抗病机制中最终起作用的基因,它们的编码产物直接或间接地作用于病原。除此之外,抗病基因和防卫反应基因的区别还有:(1)抗病基因编码产物具有特异性,而防卫反应基因编码产物具有普遍性,即不同的寄主植物中有一套类似的防卫反应基因,如植保素合成链中的酶基因、病程相关(PR)蛋白基因、植物细胞壁成分合成酶基因等。(2)抗病基因产物是植物防卫反应基因表达的直接或间接调节因子。防卫反应基因一般是受病原菌诱导表达的,编码产物比较容易分离的一类基因,而抗病基因是组成型表达的,编码产物不容易分离的一类基因。因此在基因克隆、基因编码产物的结构和功能分析等方面的研究工作中,防卫反应基因均早于抗病基因。所以植物防卫基因既有普遍性,又有特殊性。除有一部分是相似的外,还有一部分是不同的,如对真菌、细菌毒素的解毒基因,因毒素不同而不同。而人工赋予植物的解毒基因则可能更加不同,有动物源的,也有微生物源的。 1.1 抗病基因 接收病原菌信号,启动植物抗病反应信号转导的是植物抗病基因的编码产物,这是分子植物病理学研究寄主植物的重点和难点。自1992年应用转座子标签法分离出第一个抗病基 收稿日期 1999207215
植物抗病基因研究进展
植物抗病基因研究进展 摘要:植物抗病基因的研究是目前植物病理学科的热点及难点之一。近年来,通过基因工程技术培育抗病毒植物已经成为抵抗植物病毒的有效手段。本文简要讨论了近年来植物抗病毒基因工程的方法策略, 并对植物抗病基因工程的研究取得的成绩、存在的问题及展望进行了简介。 关键词植物病毒、抗病基因、基因工程、前景 一、植物抗病基因工程原理 植物抗病基因工程指的是用基因工程(遗传转化)的手段提高植物的抗病能力,以此获得转基因植物的方法。植物抗病基因工程主要包括:抗病及其他相关基因的分离和克隆、与合适的载体及标记基因构成适于转化的重组质粒、用不同的转化方法向受体植物导入重组质粒、筛选转化因子并鉴定转基因植株。此外,还有一种可以获得抗病转基因植物的方法即把具有抗病能力的植物或微生物的DNA 直接导入受体植物,从后代中筛选具有抗病能力的个体,经过稳定转化得到转基因抗病植株。 植物病毒每年给世界各地的农作物生产造成严重损失,每年全世界的农作物因病毒侵害的损失数百亿美元,传统的防治方法已远远无法满足现代农业的生产要求。病毒侵染之所以复杂,在于一方面病毒的高突变率所致的植物抗病品种抗性丧失速度远高于常规植物抗病育种速度;另一方面病毒在隐症野生植物中的储存;第三,无亲缘关系的病毒复合侵染以及病毒侵染的持久性,特别是以线虫和真菌传播的植物病毒能在土壤中存活许多年。因此,在适宜病毒介体生长的温度条件下,大面积连作缺乏抗病基因的植物,造成的经济损失会更高。Hamilton[1]于 20 世纪 80 年代初首先提出了基因工程保护的设想,在转基因植物中表达病毒基因组序列可能是防御病毒侵染的途径之一。近 20 多年来,基因工程的发展,为防治病毒病开辟了新途径。 二、利用非病毒来源的基因策略 1.植物自身基因介导的病毒抗性 一些植物在病毒侵染的时会启动主动防御机制,最普遍最常见的主动防御机制就是通常所说的过敏反应,也就是那些最初被病原侵染点周围的细胞发生程序性死亡最终在病原最初侵染点周围形成坏死斑。如番茄中的 Tm-1 或 Tm-2 和Tm-22基因,马铃薯的 Rx,Ry,烟草中的 N 基因等等[2]。这类基因通常称为 R 基因。根据其抗性水平的不同还分为:真实免疫指病毒复制完全不能发生、阈下侵染指病毒的复制仅局限于受侵染的细胞。不管 R 基因是在模式植物还是在
植物抗病、抗虫及抗除草剂基因与基因工程
植物抗病、抗虫及抗除草剂基因与基因工程 张永强 (西南大学植物保护学院, 重庆 400716) 摘 要:病虫草害历来是植物保护工作的重中之重,农药为病虫草害防治立下了汗马功劳。近来由于大量使用、滥用农药给环境带来了巨大的负面影响。20世纪70年代兴起的基因工程为这一问题的解决带来了新的途径。本文就植物抗病基因分类、最新报道的相关基因;抗虫基因的来源、最新报道的抗虫基因及试验结果;抗除草剂基因以及基因工程技术在现代农业中的应用予以综述。 关键词:植物抗病;植物抗虫;抗除草剂;基因工程 农药伴随人类改造自然,征服自然已经有100多年的历史,在促进农业发展和对人类发展做出卓越贡献的同时,也不可避免的带来许多负面影响,如:对非靶标生物的毒害、对环境的污染、对生态系统的破坏以及病虫草抗药性的产生等。特别是化学农药对动物和人类健康的影响,已经成为全人类普遍关心和急需解决的全球性问题。诞生在20世纪70年代的基因工程技术为这些问题的解决提供了一条新的途径。进入20世纪90年代具有实用价值的转基因生物品种因其诸多的优势,逐渐被人们所接受,而迅速走向商品化和产业化。 1 植物抗病基因与基因工程 植物受病原菌侵染时,会诱导相关的基因产生一系列参与植物防御反应的拮抗物质,阻止病害的传播和病原菌的进一步侵入。将这些参与植物防御反应的相关基因导入植物,使其在植物体内表达,可以提高植物的抗病能力。植物抗病基因在进化中形成了几种共有的进化形式。植物祖先抗病基因的复制创造了新基因座。基因间和基因内重组导致了变异,也导致了新特异性抗病基因的产生;另外,与特异性识别相关的富含亮氨酸重复区顺应于适应性选择;同样,类转座元件在抗病基因座中的插入加速了抗病基因的进化(庄军等,2004)。 1.1 植物抗病基因的分类 植物中许多抗病基因已被克隆,根据抗病蛋白(R蛋白)将抗病基因(R基因)分为以下几类。第一类,玉米抗圆斑病的基因Hml,其编码的解毒酶能钝化病原真菌所产生的HC 毒素,代表着抗病基因中与病原物亲和性因子作用的一类基因。 第二类,番茄抗细菌叶斑病的基因pto,其编码蛋白Pto是一种丝氨酸/苏氨酸激酶。AvrPto 蛋白是病原菌假单胞杆菌Pseudomonas进入植物细胞中通过Ⅲ型分泌系统分泌的,现已证实Pto激酶噜噗结构域中204位苏氨酸决定着Pto对AvrPto的特异性识别。具有自动磷酸化能力的Pto激酶与AvrPto相互作用从而产生了过敏性反应。 第三类抗病基因所编码的蛋白显示出与细胞间信号转导蛋白具有结构相似性。这些蛋白所共有的基元是富含亮氨酸重复序列(Leucine-rich repeat,LRR),一般由24个氨基酸残基组成,其共同蛋白序列是LXXLXXLXXLXLXXNXLSGXIPXX(氨基酸的单字符号,X代表任何一种氨基酸)。这一类型基因的共同结构是LRR-TM,它们编码的蛋白包括胞外N端LRR 重复区、膜锚定蛋白和胞质内C末端部分(如图1所示)。 第四类是水稻抗白叶枯病Xanthomonas oryzae pv.oryzae,Xoo的基因xa27。这一基因所编码的Xa21蛋白具有3个受体激酶特征的主要结构域:胞外LRRs结构、跨膜结构域及胞内激
植物抗病性的分子机制和信号传导
第42卷2006年第2期 西 北 师 范 大 学 学 报(自然科学版) Vol 142 2006 No 12 Journal of Northwest Normal University (Natural Science ) 收稿日期:2005Ο03Ο23;修改稿收到日期:2005Ο05Ο23 作者简介:李淮(1959— ),男,甘肃临洮人,馆员.主要研究方向为生物信息学.?科研综述? 植物抗病性的分子机制和信号传导 李 淮1,王 莱1,武国凡1,于 玲2 (11西北师范大学生命科学学院,甘肃兰州 730070; 21南京农业大学作物遗传育种国家重点实验室,江苏南京 210000) 摘 要:植物抗病性的分子机制一直是植物病理学关注的焦点.近年来,国内外不少学者和实验室正在大量分离和培 养与抗病有关的突变体,并且寻找和研究与抗病有关的基因和抗病机制.研究表明,在病原物与植物的相互作用、病原信号的传导和抗病性激发的过程中存在着一系列的调节因子和基因,并形成复杂的调控网络.综述了近年来国内外植物抗病性的分子研究进展,阐述了植物抗病性分子机制和信号传导.关键词:植物抗病机制;信号传导;水杨酸中图分类号:Q 7:Q 94518 文献标识码:A 文章编号:10012988Ⅹ(2006)022******* The mechanism and signal t ransduction of plant disease resistance L I Huai 1,WAN G Lai 1,WU Guo Οfan 1,YU Ling 2 (11College of Life Science ,Northwest Normal University ,Lanzhou 730070,G ansu ,China ; 21National Key Laboratory of G enetics and Breeding ,Nanjing Agriculture University ,Nanjin 210000,Jiangsu ,China ) Abstract :This article clarifies t he advance of mechanism and signal t ransduction of plant disease resistance.The mechanism of plant disease resistance is always t he focus of plant pat hology.Recently ,many mutant s related to plant disease resistance have been t rained and separated in many laboratory.Now new gene and mechanism about plant disease resistance are still seeked and researched. The result s indicate t hat series gene and regulation factors are involved in interaction between plant and pat hogen ,in signal t ransduction and in p rocess of stimulating disease resistance ,and t he complicated regulation net is established. K ey w ords :mechanism of plant disease resistance ;signal t ransduction ;salicylic acid 研究植物的抗病性不仅直接关系到作物产量和 质量的提高,而且对于植物保护和环境建设也具有同样重要的意义.研究发现,植物的抗病性不仅与植物的种类有关,而且与病原物有直接关系.目前,对于病原物致病、植物抗病的分子生物学基础和信号传导方面的研究已取得了一系列进展. 1 植物抗病性的分子机制 111 病原菌致病的分子基础 植物对病原物的反应有抗病和感病两大类:抗 病反应又叫非亲和反应,这一系统是以寄主抗病和病原物无毒为特征,寄主植物对病原物有抑制、排斥和减毒作用,使病害不发生或受到限制;感病反应又叫亲和反应,以寄主感病和病原物有毒为特征,造成植物严重发病[1].通常由几类物质被认为是病原物致病因子,即毒素、酶类、胞外多糖及其它毒性因子.植物病害的症状类型与致病因子的性质有密切的关系,如腐烂通常认为与病菌的胞壁降解酶有关;坏死与毒素有关;萎蔫可能与毒素有关,也可能与胞外多糖有关;生长畸形与激素失调 3 11
《植物免疫学》课程教学大纲 - 西北农林科技大学植物保护学院
《植物免疫学》课程教学大纲 课程编号:02013 英文名称:Plant immunology 一.课程说明 植物免疫学是专门研究植物抗病性机制及其利用方法的科学,是植物类专业的一门重要的选修课。 1. 课程类别 专业课程 2. 适应专业及课程性质 植物保护专业、农学专业、园艺专业和草业专业选修 3. 课程目的 通过本课程的学习可以让学生了解植物抗病性原理,系统掌握利用植物抗病性的基本理论与方法,达到科学利用植物抗病性控制植物病害的目的。 4. 学分与学时 学分为1.5.学时为32 5 .建议先修课程 植物学、植物生理学、植物生物化学、遗传学、作物遗传育种学、普通植物病理学和农业植物病理学 6. 推荐教材或参考书目 推荐教材: (1)植物免疫学(第一版).李振岐主编.中国农业出版社出版.1995年 (2)植物免疫学实验(第一版).商鸿生主编.中国农业出版社出版.1994年 参考书目: (1)植物病理过程的遗传学和分子基础.范德普朗克著(曾士迈等译).上海科学技术出版社.1982年 (2)植物分子遗传学(第二版).刘良式.科学出版社.2003年 (3)主要农作物抗病性遗传研究进展.朱立宏.江苏科技出版社.1990年 (4)抗病性的持久性.杨作民等.中国农业大学出版社.1997年 (5)植物生理与分子生物学(第三版).余叔文等.中国科学出版社.2005年 7. 教学方法与手段 本课程采用课堂教学与实验教学相结合的方法 8. 考核及成绩评定 考核方式:考试 成绩评定: (1)平时成绩占30%,形式有:实验课,平时测验 (2)考试成绩占70%,形式有:闭卷考试 9.课外自学要求 据教学进展情况,要求学生对中外文植物免疫学相关文献进行性选择性自学,扩大知识面,并对所学内容做好笔记,将自学笔记作为考核平时成绩的依据 二、课程教学基本内容及要求 第一章绪论
植物诱导抗病性的结构抗性机制
万方数据
万方数据
万方数据
植物诱导抗病性的结构抗性机制 作者:刁毅, Diao Yi 作者单位:攀枝花学院生物与化学工程系,四川,攀枝花,617000 刊名: 攀枝花学院学报(综合版) 英文刊名:JOURNAL OF PANZHIHUA UNIVERSITY 年,卷(期):2006,23(1) 被引用次数:7次 参考文献(23条) 1.董合忠;李维江植物诱导抗病性及其利用[期刊论文]-莱阳农学院学报 2001(04) 2.张元恩植物诱导抗病性研究进展 1987(02) 3.张晓燕;武爱兵Ca2+在植物诱导抗病性中的作用[期刊论文]-河北林果研究 2002(02) 4.刘西钊;陈长永棉花枯萎病抗病性诱导物的筛选和有效间隔期 1990(02) 5.李冠植物诱导抗病性 1990(06) 6.J.G.Fuchs;Y.M.oenee-loccoz;G.Defago Nonphathogenic Fusarium oxysporum Strain Fo47 indudces Resisitance to Fusarium Wilt in Tomato 7.T.Reglinski;P.R.Poole Induced resistance against Sclerotinia Sclerotiorum in kiwifruit leaves[外文期刊] 1997(46) 8.Richard Karban;Rodney Adamchak Induced Resistance and interspecific competition between spider mites and a vascular wilt Fungis[外文期刊] 1987 9.L.sticher;B.Mauch-Mani;metraus Systemic Acquired resistance 1997 10.Induction of systemic acquired disease Resistance in plants by chemicals 1994 11.Vance C.P;Kirk T.K Lignification as a mechanism of disease resistance 1980 12.Campbell M.M;Ellis B.E Fungal elicitor mediated response in pine cell cultures 1992 13.梁永超;孙万春硅和诱导接种对黄瓜碳疽病的抗性研究[期刊论文]-中国农业科学 2002(03) 14.骆桂芬;孙俊涛黄瓜叶片中糖和木质素的含量与霜霉病诱导抗性的关系 1997(01) 15.胡景江;朱玮溃疡菌杨树细胞壁中HRGP和木质素的诱导作用 1997(03) 16.宾金华;潘瑞织茉莉酸甲酯诱导烟草幼苗抗病与过氧化物酶活性和木质素含量的关系[期刊论文]-应用与环境生物学报 1999(02) 17.石雪晖;王淑英葡萄叶片中单宁、木质素、PPO活性与抗黑豆病的关系 1997(04) 18.黄雪梅;张昭其果蔬采后诱导抗病性[期刊论文]-植物学通报 2002(04) 19.李淼;檀根甲植物抗病性研究现状与前景展望[期刊论文]-江西农业大学学报(自然科学版) 2002(05) 20.毛爱军水杨酸诱导辣椒抗疫病生化机制的研究 2006(05) 21.夏启中植物抗病的物质代谢基础[期刊论文]-黄冈职业技术学院学报 2004(03) 22.胡东维白粉菌初生芽管对小麦诱导抗性的细胞学研究 1997(02) 23.马青寡聚糖诱导黄瓜对白粉菌抗病反应的超微结构研究[期刊论文]-植物病理学报 2004(06) 本文读者也读过(10条) 1.王瑜.吴丽芳.余增亮茉莉酸及其甲酯在植物诱导抗病性中的作用[期刊论文]-生物学杂志2000,17(1) 2.王海华.康健植物诱导抗病性的应用研究与展望[期刊论文]-湘潭师范学院学报(自然科学版)2003,25(3) 3.王海华.康健植物诱导抗病性及应用前景[期刊论文]-生物学通报2001,36(6)
植物抗病基因研究进展
植物抗病基因研究进展 摘要:植物抗病基因在植物抗病过程中起重要作用。综述了植物抗病基因的克隆方法,结构特点,作用机制以及未来的发展前景。 关键词:R-基因;克隆方法;结构特点;作用机制 Advance on Plant Resistance Gene Research Abstract:R gene played an important role in defending the disease. This paper reviewed the gene cloning,structure characteristic,function mechanism and the foreground of R gene. Key words:R gene,gene cloning,structure characteristic,function mechanism R基因在植物的病害防御过程中起着非常重要的作用。植物时刻受到周围各种潜在病原菌的侵染。一旦病原菌侵入植物表皮,将遇到R基因和avr基因的互作,即植物的非寄主抗病性[1]。随之所产生的现象包括局部细胞坏死(过敏性细胞死亡),木质素和胼胝质形成,以及所有与抑制病原菌生长和病害发生相关的抗菌物质产生。病原菌还有可能通过不同的途径赋予植物长期的系统抗性诱导系统免疫,使植物能抵抗其他相似病原菌的侵染[2]。随着分子生物学的发展,更多的植物抗病基因克隆方法及作用机制逐渐为人们所发现,也有更多的植物抗病基因的功能得到了证实。 1 R基因的克隆方法 1) 表型基因克隆法:利用表现型差异或组织器官特异表达产生的差异来克隆植物基因就是表型克隆。表型克隆在策略上试图将表型基因与结构基因表达的特征联系起来,从而分离特定的与表型相关的基因,力求不必事先知道基因的生化功能或图谱定位,根据基因的表达效应就能直接分离该基因。 2) 转座子标签技术:转座子标签技术是比较经典的方法,是将转座子或T-DNA插入到欲分离基因的内部,基因发生突变而被标识,然后利用插入2个和小麦(Triticum aestivum)1 个(表1)[5]。 尽管R基因之间的序列同源性很低,但是这些R基因编码的蛋白也具有一些相似的结构特征,根据这些结构特征,已经克隆的R基因可以分为5个大类:
植物基因工程
一从现代农业到基因工程 (一)粮食安全现状 1、食物总量供给已成为全球的焦点之一: 从2000年开始,全球出现了当年粮食生产量比消费量低的情况,2003年全世界粮食的消费量超过生产量0.93亿吨,世界粮食储备也降低到30年来的最低水平。 1999年以来,我国粮食连续四年减产。1999-2002年,我国粮食总产量累计减少800亿公斤左右。自2000年以来,我国粮食年消费需求大致在4.8-4.9亿吨之间,产需缺口约400亿公斤。 (二)农业发展的一个主要矛盾——科技支撑能力不强 农业生产的规模化、专业化和多样化对科技提出了更高的要求,大幅度提高农业劳动生产率需要通过先进适用技术的广泛应用,而目前我国科技进步贡献率只有45%左右,与发达国家的70-80%有很大的差距。 一个农业劳动力养活的人口数: 美国:70人; 日本:约25人; 中国:4-5人。 农业发展的根本出路是现代农业,而其核心支撑条件是现代农业科技的进步。 (三)现代农业的内涵 现代农业是以现代工业和科学技术为基础,重视加强农业基础设施建设,充分汲取中国传统农业的精华,根据国内外市场需要和WTO规则,建立起采用现代科学技术、运用现代工业装备、推行现代管理理念和方法的农业综合体系(引自卢良恕院士)。 (四)建设农业科技创新体系是现代农业的一个根本任务 国家级农业科研工作应具有较强的关键性、全局性、基础性、战略性和前瞻性的特点,为加快现代农业建设提供科技支撑。省级有关农业的科研机构应逐步实行联合,重点开展应用研究和开发研究(也可根据需要适当开展应用基础研究),重视科技成果转化,更好地为发展生产服务(引自卢良恕院士)。 到2030年,我国人口的持续增长将要达到高峰期,预计达到16亿人口,解决这个庞大人口的口粮是一个新的挑战。 随着人民生活水平的提高,肉蛋奶和水产品的消费不断增加,粮食作为饲料的比重将越来越大,人均粮食占有量的标准应有所提高。 2、食品安全性也成为全球的焦点之一: 农业综合措施、现代农业技术尤其是转基因技术的应用,使老百姓对当前食品尤其是转基因食品安全性问题十分关心。 (五)农业科技创新的一个核心内容:良种创新 农业科技创新的核心:良种+良法。良种对增产的作用所占的比重越来越大,良种是一个先进技术的集合体。 良种创新:植物良种创新、动物良种创新。植物食品占总食品的93%,动物食品占7%,但也间接来自植物食品,所以良种创新的首要任务是植物良种创新。 (六)传统育种面临的挑战 以杂交育种为核心的传统育种技术取得了丰硕的成果,目前仍然是主要作物的主要育种手段。目前传统育种技术在改良作物性状方面遇到了一些挑战,如缺乏特别性状的种质资源,育种周期长,难以克服不良性状的连锁或负相关,易受杂交不亲和及杂种不育的限制,远缘物种间不能进行遗物物质交流和性状转移。 (七)基因工程带来的机遇与竞争 20世纪50年代以来,DNA双螺旋模型和基因操纵子学说的提出,以及DNA限制性内切酶的发现,导致了DNA体外重组技术?a?a基因工程技术的发展,推动了分子生物学和基因工程本身在广度和深度方面以空前的速度蓬勃发展,生物技术相关产业和生命科学已经出现划时代的
植物免疫的机制
植物免疫的机制 摘要:植物具有特异与动物的免疫机制,它表现在避病性、抗病性及耐病性三方面,是植物在于病原物对抗并协同进化的过程中获得的。植物在受到病原物入侵后,不亲和植株能通过在入侵部位识别诱导因子,再通过各种信号传递途径引起预存诱导防卫系统及诱导防卫系统的应答,依靠物理性及化学性的防御机制,阻止病原物的入侵,同时整株植株获得诱导性系统抗性。 关键词:植物;免疫机制 Mechanisms of Plants’ immunity Summary: Plants have immune mechanisms which are different from animals, they represent at three ways—preventing, resisting and enduring. These mechanisms are obtained by antagonizing and evolving with pathogens. After invaded by pathogens, antipathic plants can recognize inducing factors in invasive place, response by prestored inducible system and inducible system throught many kinds of signal ways, stop pathogens’ invading by physics and chemistry recovery mechanisms and obtain inductivity systemic fastness at the same time. Key words: plants; immune mechanisms. 在自然环境中,植物与各种病原物的接触无时无处不在,但在大多数情况下,植物却能正常生长发育并繁衍后代,这是因为植物在其长期演化过程中,形成了多种与病原物对抗的途径,获得了免疫性。由于植物营固着生活,不像动物般能通过运动来躲避病原物,也不具有动物般的神经系统及体液循环系统,因此,植物的免疫具有不同于动物的特点。 1.植物免疫性的表现 植物的免疫性表现在避病性、抗病性及耐病性三方面。 1.1.植物的避病性 自然界的各种病原物几乎都有一个最适宜的发生和传播期,这是因为病原物的生长、传播及繁殖对周围的自然环境有一定的要求,如温度、湿度、酸碱度等,使其生长周期与节气相关。比如马铃薯晚疫病的病原菌大量发生和传播的最适条件是低温和高湿,对应我国华北地区就是七、八月份的雨季。同时,植物对某些病害又有一个最易染病期,上述的马铃薯晚疫病的最易染病期就在现蕾之后。不难发现这是由病原物的入侵特性决定的,如病原物特定的入侵途径、特定的入侵部位等,使其入侵与植物的生长周期相关。 如此一来,有的植物就可以通过发展出使其最易染病期避开病原物大量发生和传播期的免疫的机制而免受或少受病原物入侵,即获得了避病性。 植物的避病性的获得相信是生物间协同进化的结果,即由于自然选择的作用,最易染病期与病原物大量发生和传播期相一致的植物因受病原物入侵而灭亡,不能通过繁殖而将其基因传给后代,而最易染病期与病原物大量发生和传播期不一致的植物却得以繁衍,使种群的基因频率发生改变,最终种群获得了避病性。 1.2.植物的抗病性 植物的抗病性是指植物直接抵抗病原物入侵的特性,包括抗侵入、抗寄生及抗再侵染三方面。 1.2.1.抗入侵 抗入侵是指植物在受到病原物通过机械力量或酶类溶解植物表层或植物的伤口的方式
植物抗病基因工程研究进展
植物抗病基因工程研究进展 摘要随着植物抗病基因的分离,植物抗病机制的分子生物学和植物抗病基因工程取得了重大研究进展。该文就植物抗病基因工程的原理、目的基因、转化方法等进行综述,并对植物抗病基因工程的应用前景做了展望。 关键词植物;抗病基因;基因工程;原理;目的基因;转化方法;前景 随着世界人口的迅速增长,粮食问题已成为人类生存的关键问题。有专家预测,到2050 年,全球人口总数将膨胀至90 亿。剧增的人口将给为人类提供粮食的农业生产带来严峻的挑战。众多学者为提高作物产量作了许多努力,也取得了很大成果。但是,长期以来,因病菌侵染而造成的作物产量损失也是巨大的。当前,防治病害的主要策略是改进栽培措施和施用化学杀菌剂。但这只能从一定程度上控制病害的流行而不能从根本上解决问题,而且化学药剂所带来的环境污染和病原抗药性生理小种的形成等问题也给病害防治造成了更大的障碍。 自上世纪90 年代以来,分子生物学理论和技术的不断发展完善,使人们能够从分子水平上研究植物与病原菌的相互作用机制,植物基因工程的兴起更是为病害的控制提供了更广泛的选择余地。基因工程被认为是一项能为人类提供以食用动物为基础的健康和充足粮食途径的关键技术,在应用中扮演着重要角色。在植物抗病基因工程的研究历程中,以植物抗病毒基因工程开展最早,发展也最为迅速,部分转基因植株已开始用于生产。随着植物抗病反应机制和病原菌致病机理研究的深入,近年来植物抗病基因工程的研究取得了很大的进展。 1 植物抗病基因工程的原理 人们对植病互作机制的认识,主要来源于对模式植物拟南芥的研究。并且已经从拟南芥中鉴定和克隆了许多抗病基因,给其他作物的抗病性遗传分析提供了理论基础。 病原菌对宿主植物成功的感染,包括接触识别、崩解植物理化防御系统、产生毒素、灭活整个植株或部分组织的代谢生理活性。病原菌往往含有致病基因和毒性基因,其表达 调控包含有复杂的信号传导。 在经典遗传学中,植物与病原物的互作被看作是由基因型控制的,植物抗病性常常是由来源于植物的抗病基因R 与相应的来源于病原物的无毒基因avr 互相作用所决定的,即“基因对基因”学说。Flor通过亚麻与亚麻锈病菌之间的相互关系的研究,发现真菌的显性avr 基因的产物
唐定中-植物抗病信号
植物抗病信号 唐定中 一、概况 1、研究植物抗病的原因:因为它既是重要的生物学问题也是重要的农业问题。每年稻 瘟病造成水稻损失50-60亿美元。所以作物病害对农作物生产造成的危害非常严重,直接影响粮食安全。另外,病原菌还具有变异迅速,扩散迅猛。例如Vg99引起的 小麦杆锈病,在世界范围内都有扩增。 2、研究较多的几种病原菌类型主要有:细菌(如:假单胞杆菌、黄单胞杆菌)、真菌 (如:白粉菌、稻瘟病)、卵菌(如:霜霉病菌、疫霉菌)、病毒(如:烟草花叶病 毒、黄瓜花叶病毒)。这些病菌可分为两类不同的植物病原菌,即a.死体营养型或 腐生型,b.活体营养型或寄生型。 3、病菌感染植物的一般过程或条件:a.锚定/进入宿主细胞,b.抑制宿主的抗性,c.改 变宿主的生理机能,如对营养和水的需求,d.在宿主细胞内扩增,e.最终导致疾病 的发生。 4、病菌感染植物后,植物会产生两种反应:即抗病(亦称非亲和性反应,此反应中病 菌是无毒的,植物产生抗性。)和感病(亦称亲和反应,病原菌有毒,植物是敏感 型的) 5、如何确定植物的病型?答:一是测定病原菌的生长情况。对于白粉菌来说要看其孢 子数,对于细菌来说,要测定其生长曲线。二是评价植物的损伤程度。 6、植物的抗病层次主要有:非寄主抗性、基础抗性(PTI)、R基因介导的抗性(ETI)。 二、非寄主性抗性 1、大部分植物都能对大多数病菌产生抗性。多数情况下从某一种植物中分离到的病 原菌不能再次感染或引起其它植物感染,也不能繁殖。非寄主性抗性就是植物能 够抵抗大部分潜在致病菌的原因,是最常见的植物抗病方式。 2、非寄主性抗性的机理:(1)消极防御也可以说是天然的防御系统,只要是植物表 面的物理障碍(如角质层、细胞壁等)以及一些代谢产物,大部分为抗菌素。(2) 诱导型的植物抗病机制。如植物抗毒素,它是植物在受到病原菌的侵害后应答从 头合成的一种抗菌物 3、非寄主性抗性所涉及的基因:多数情况下,植物针对真菌产生的非寄主性抗性是 在其侵入的过程中进行。拟南芥的PEN1, PEN2和PEN3基因可用于对大麦白粉病 产生抗性。(PEN1是一种突触融合蛋白,它在细胞膜的囊泡运输过程中具有重要 作用。PEN2是一种糖基水解酶。PEN3是ABC转运蛋白) 4、病原菌可通过多种方式破坏植物的非寄主性抗性系统。从病原菌进入植物的方式 看,可通过植物体表已有的缺口进入,如气孔、伤口;也可痛过分解酶分解角质; 或是通过纤维素酶、果胶酶、纤维素内切酶等破坏细胞壁。 三、基础抗性(PAMP triggered immunity, PTI) PAMPs:病原相关的分子特征。MAMPs:细菌相关分子特征 PAMPs对于病原菌的生活方式来说是非常重要的,它在大部分微生物中都高度保守,但在宿主中却不存在。植物通过识别PAMPs来激活其基础抗性。 PAMPs主要有:鞭毛蛋白、脂多糖、几丁质以及麦角脂醇等。 1、PAMP的识别及基础免疫 人们对植物对PAMPs的应答的认识大部分来自于鞭毛蛋白。鞭毛蛋白是细菌运动的 重要组成部分,其N端和C端都高度保守,这使其成为非常重要的PAMP,且其不
植物病害调查与抗病性鉴定
普通植物病理学实验报告 植物病害调查与抗病性鉴定 专业:植物保护 班级:2010级1班 姓名:______尹全飞____ 学号:_____20100333____ 二零一二年四月一十五日
一、实验目的 1、掌握小麦抗性鉴定方法。 2、识别不同种对同一病原物的抗病反应类型; 3、熟悉病害严重度调查方法; 二、实验材料 田间正常生长小麦,包括品种:B21-F6-5-4、BL-21-13-7-5-3、苏麦-3 B44-F6-1-5 三、实验方法 1、随机抽取100株同种小麦植株观察其旗叶; 2、观察鉴定并记录其抗病感病类型及严重度; 3、用同样的方法观察记录四种不同的小麦品种; 4、记录、整理和分析数据,并制作出统计图表; 5、根据数据及表格,分析讨论,并且最终得出结论; 四、实验数据及处理 病级:免疫(0)、近免疫(0;)、高抗(1)、中抗(2)、中感(3)、高感(4)、混合型(X) 严重度:0—7级 一、品种:B21-F6-5-4 生育期:乳熟期 数量:100株 调查时间:2012年4月6日17:00 调查人:尹全飞、杨平、徐伯驹 编号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 病级0 0;0; 2 0 0; 3 3 4 4 X 3 0; 0; 0; 1 4 3 0 0; 严重 度0 1 2 2 0 0 4 3 6 6 7 4 1 0 2 1 6 4 1 2 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 0; 0; 0; 2 0 0 0 0; 0 0 0; 0; 0 0; 0; 0 0 0 0; 0; 0 0 1 2 2 0 0 1 0 0 1 2 0 1 2 0 0 0 1 0 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 0 0 2 0; 0 0 0; 0; 0 0; 0; 0; 0 0 0; 0; 0; 0; 0 0; 0 0 1 0 0 0 1 0 0 0 1 0 0 1 1 1 1 1 1 0