离子液与蛋白质和核酸相互作用的研究
简述核酸和蛋白质代谢的相互关系

简述核酸和蛋白质代谢的相互关系全文共四篇示例,供读者参考第一篇示例:核酸是细胞内的一种重要有机物质,它由核苷酸构成,是构成核酸的基本单元。
核酸分为DNA(脱氧核糖核酸)和RNA(核糖核酸)两种。
核酸在细胞内具有非常重要的功能,它们可以携带遗传信息,参与蛋白质的合成,调控细胞的生长和分化等过程。
蛋白质则是细胞内最重要的有机物质之一,是生命体内各种生物学功能和生命活动不可或缺的组成部分。
蛋白质合成是一个复杂的生物化学过程,需要核酸的介入才能完成。
在细胞内,RNA起着传递DNA信息的作用,RNA通过转录过程将DNA上的遗传信息转换成RNA信息,然后RNA将这些信息传递给细胞内的核蛋白合成机器,进而合成蛋白质。
核酸代谢和蛋白质代谢是密切相关的,两者之间存在着相互关系。
在细胞内,核酸和蛋白质代谢之间的相互关系主要体现在以下几个方面:核酸还可以调控蛋白质的合成。
在细胞内,存在着一些特殊类型的RNA,如miRNA和siRNA等,它们能够通过靶向特定基因的mRNA,抑制或促进这些基因的表达,从而影响蛋白质的合成。
这种核酸介导的蛋白质合成调控,使得核酸和蛋白质代谢之间形成了一种复杂的调控网络。
核酸代谢和蛋白质代谢还存在着其他相互关系。
核酸可以通过调节细胞内mRNA的降解速率,影响蛋白质的合成水平;而蛋白质也可以参与核酸的合成和修复过程。
这些相互关系构成了细胞内核酸和蛋白质代谢的相互调节机制,维持了细胞内生物学功能的正常运行。
第二篇示例:核酸和蛋白质是生物体内两种重要的生物大分子,它们在生物体内的代谢过程中密不可分。
核酸是生物体内的遗传物质,负责信息的传递和储存,而蛋白质则是生物体内的最重要的功能分子,承担着多种生物过程中的功能。
核酸和蛋白质之间通过一系列生物化学反应相互转化,相互影响,共同维持着生物体内的代谢平衡和生物功能的正常进行。
核酸的合成过程称为核酸代谢,蛋白质的合成过程称为蛋白质代谢。
核酸和蛋白质的代谢密切相关,二者之间的相互关系主要体现在以下几个方面:核酸和蛋白质的合成过程相互依赖。
质谱技术在生物大分子研究中的应用

质谱技术在生物大分子研究中的应用随着生物技术和生命科学的发展,质谱技术开始在生物大分子研究中得到广泛应用。
质谱技术是一种分析方法,可以通过检测并分析样品中的化合物来确定其分子结构和组成。
在生物大分子研究中,质谱技术可以用来研究蛋白质、核酸和糖类等生物大分子的结构、功能和相互作用等。
一、蛋白质质谱蛋白质是生物体内最基本的分子,其功能包括酶催化、信号转导和结构支撑等方面。
蛋白质的性质和功能由其结构决定,因此研究蛋白质的结构及其相互作用对生物学和医学领域具有重要的意义。
质谱技术可以用来研究蛋白质的结构和功能,如质谱分析可以确定蛋白质的分子量和氨基酸序列。
基质辅助激光解离/飞行时间质谱(MALDI-TOF MS)技术可以分析蛋白质和肽片段的分子量,而痕量MS技术可以精确测量蛋白质的分子质量。
液相色谱-串联质谱法(LC-MS/MS)可以鉴定蛋白质中的氨基酸序列和修饰信息,如磷酸化和甘油化等,从而揭示蛋白质的功能和相互作用。
二、核酸质谱核酸是生物体内遗传信息传递的媒介,包括DNA和RNA。
核酸的结构和功能也是其生物学意义的重要方面。
质谱技术可以用来研究核酸的结构和分子量等信息。
质谱法可以通过测定核酸片段的分子量来确定其序列和修饰。
电喷雾质谱(ESI-MS)和MALDI-TOF MS可以用来对DNA和RNA分子进行分析。
这些技术通常需要在较高的离子流量下分析样品,并产生高分辨率质谱图。
此外,质谱技术也可以用于检测DNA和RNA中的化学修饰,如磷酸化、甲基化和糖基化等。
这些化学修饰对核酸的空间结构和功能有很大的影响。
三、糖类质谱糖类是生物体内最多样化的大分子之一,包括各种多糖和糖蛋白等。
质谱技术可以用来研究糖类的结构和分子量等信息。
常用的糖类质谱技术包括ESI-MS和MALDI-TOF MS。
这些技术可以用来测定糖类的分子量、组成和链接信息,并指导糖类结构的确定和构建。
液相色谱-串联质谱法可以定量分析有关糖类结构的信息,如糖链结构和糖蛋白的修饰等。
2010化学科学部国家自然科学基金项目

项目批准号/申请代码1项目名称项目负责人21002081/B0201 Smiles重排应用于合成黄樟素衍生物研究血管内皮细胞凋亡的分子机制左华21021004/B0501 复杂体系的高效分离与表征 邹汉法21073071/B0301 高压下有机晶体的多晶型研究 邹勃21003047/B0305 表面活性素的定向结构改造、结构与性能研究 邹爱华21075076/B050306 小分子与蛋白质相互作用的表面增强拉曼散射检测方法研究宗瑞隆21072065/B020706 含深度共熔溶剂介质中醋酸菌Acetobacter sp.CCTCC M209061细胞催化手性醇不对称合成反应的研究宗敏华21074013/B0401 新型手性稀土金属络合物催化丙交酯立体选择性开环聚合反应研究自国甫21003117/B030203 紧密结合长程分子动力学计算机模拟和二维红外光谱技术以研究蛋白质折叠的动力学机理庄巍21010302022/B070201 东亚沙尘/气溶胶及其对全球气候变化的影响国际学术研讨会庄国顺21077060/B0704 典型全氟化合物在沉积物中的分配行为与微观机制 祝凌燕21077119/B070302 河流岸边带厌氧氨氧化反应的热区分布与过程效应 祝贵兵21076198/B060201 含固体颗粒的液态化工介质离心泵输送特性研究 朱祖超21072108/B020901 新型噻唑类除草剂的设计、合成与构效关系的研究 朱有全21077100/B070203 水稻土中藻对砷的甲基化作用及分子机制 朱永官21003046/B030201 丙烷脱氢-氧化耦合工艺中Pt基核壳双金属催化剂作用机制的第一性原理研究朱贻安21001095/B0104 金属核酸酶与DNA结合模式及切割活性的理论研究 朱艳艳21074082/B040101 含硒的功能性RAFT试剂的合成及其聚合研究 朱秀林21010302028/B04 第二届中加先进材料会议 朱秀林21006097/B060409 基于机械力活化理论的氯代芳烃固态Heck反应研究 朱兴一21024801/B01 Science China Chemistry 朱晓文21072104/B020507 黄素辅酶及其模型物负氢转移各基元步骤热力学研究朱晓晴21006104/B061201 多级孔分子筛催化剂上废塑料高效催化转化的定向调控朱向学21036006/B060203 分子筛及其膜材料的吸附、扩散与分离性能研究 朱伟东21076077/B060702 有害重金属离子高灵敏检测与高效分离一体化荧光传感器朱维平21077039/B070102 碳纳米管整体柱微萃取/全二维气相色谱法同时检测环境样品中超痕量二噁英和多氯联苯朱书奎21072190/B020101 通过分子内C-H键官能化合成几类杂环化合物的新方法研究朱强21072001/B0205 巯基作为配体的金纳米团簇参与有机反应的研究 朱满洲21073157/B0301 新颖贵金属磺基苯甲酸化合物结构与催化性能研究 朱龙观21001017/B0101 超声波辅助离子液体法合成稀土氟化物纳米晶及其光学性能研究朱玲21073062/B030702 具有荧光示踪功能的光控释放药物的量子点纳米复合物朱麟勇21006024/B060409 CO2-CH4干气重整NiMgO催化剂的极性(111)表面设计、制备和活性及抗积炭性能研究朱卡克21020102038/B05 功能纳米材料的组装与光电生物传感 朱俊杰21002028/B020601 Falcipain-2和DHFR双重抑制剂的设计、合成及其生物学评价朱进21072187/B020506 基于酰胺折叠物的新型螺旋状纳米管的设计、合成及性质研究朱槿21004025/B040606 嵌段共聚物乳液液滴的界面不稳定现象机理与微结构调控朱锦涛21006054/B060702 新型齐聚物糖基水凝胶因子设计合成及其凝胶行为 朱金丽21071014/B0101 氮化物以及氮氧化物可见光光催化剂的制备与性能研究朱鸿民21077137/B0704 SPME原位采样技术监测土壤-农作物系统的持久性有机污染物朱芳21076234/B060203 火灾下热功能含湿织物的干燥收缩分形分析及湿热传递模型朱方龙21072151/B020706 生物催化不对称羰基还原胺化反应的探索 朱敦明21066014/B0608 内生真菌石杉碱甲生物合成途径及代谢调控研究 朱笃21077049/B070203 土壤中煤源颗粒对有机污染物的吸附、解吸研究朱东强21074056/B040606 利用介电松弛谱研究酶电极中导电高聚物与生物大分子界面微结构及电荷传输朱丹21001033/B010701 介孔材料-核酸适体的组装及在药物控释技术中的应用研究朱春玲21061003/B010303 含氮、氧配位供体原子的有机配体及其配合物的合成、结构及性能研究朱必学21002069/B0202 双核金属配合物“协同”活化惰性C-H键朱柏林21002048/B0206 新型黄酮类肿瘤血管阻断剂的合成与生物活性研究 周中振21073152/B030606 电催化过程中低覆盖度吸附态中间体的原位红外光谱检测周志有21073096/B0302 无机纳米薄片/条带的计算设计与嵌锂性能周震21071051/B010401 扭曲度可调型类血红素铁卟啉的合成及其复合物性能研究周再春21074134/B040502 PEO树枝齐聚物嵌段共聚物的自组装与结晶行为研究周云春21071143/B010303 纳米尺度金属有机骨架材料的设计合成及其催化构效关系研究周有福21075114/B0511 膜保护配位聚合物微固相萃取技术在多溴联苯醚预富集和分析中的应用周友亚21006129/B060409 微乳液中纳米粒子定点负载构建新型钯整体式催化剂的研究周永华21074069/B040502 超支化聚合物的支化拓扑结构和性能关系研究周永丰21003075/B030606 DMFC电催化剂载体材料氮掺杂石墨烯的基础与应用研究周盈科21062003/B0207 中药桑白皮对HIV-1 LTR启动子活性的调控作用研究周英21076142/B060304 氧化铁/一氧化碳循环分解水制氢基础研究 周亚平21001065/B0103 芴基发光金属-有机骨架材料的设计合成和性能研究周馨慧21076036/B060702 砜和手性亚砜的选择性氧化合成及其机理研究 周新锐21003115/B030201 太阳能光催化制氢材料吸光机制的理论研究周新21073173/B030402 乙烯及衍生物分子的电离能、键能及解离动力学研究周晓国21003110/B030607 氧化-还原分子电子输运的STM裂结技术和电化学超快循环伏安法研究周小顺21072132/B020104 水溶性金属配合物催化水相偶联反应的研究 周向葛21072155/B020702 可诱导核酸交联剂的设计、合成及生物活性研究 周翔21002016/B020402 醉鱼草属植物杀虫活性成分及其作用机制研究周霞21003088/B030605 现场表面振动光谱法研究锂离子电池电极和离子液体的界面结构周尉21003005/B030502 介电弛豫法对电流变液动态界面多重极化特性的解析研究周威21010302016/B06反应与分离系统的耦合与集成周涛21007059/B0707 长三角毗邻海域有机氯农药随食物链(网)的迁移转化及生态风险周珊珊21037002/B070203 土壤污染微界面过程及其分子诊断与调控原理 周启星21032003/B020104 高效不对称催化反应及其在天然产物和手性药物合成中的应用研究周其林21074025/B0404 灵芝有效降糖天然大分子提取物的组成结构及在糖尿病治疗中的作用机理周平21077130/B0703 甲基对硫磷及4-硝基酚污染土壤的微生物修复机理研究周宁一21074080/B040101 多种拓扑结构的环型偶氮苯聚合物的设计、合成和性能研究周年琛21006036/B0612 碱木质素的溶液行为及其化学修饰模型物在固液界面的吸附机理研究周明松21072218/B021102 荧光编码标记分子(“分子条形码”)概念性验证 周明21004053/B040501 生物相容超分子聚合物的合成及其在磁共振造影剂的应用周密21001045/B0104 硒通过调节内质网应激发挥类胰岛素作用的研究 周军21002118/B0208 有机小分子催化的不对称环加成反应机理研究 周静21002005/B020702 应用新型质谱技术研究人类端粒G-四链体DNA的结构特性及其与小分子的结合位点周江21063004/B030105 在线灌注活细胞的P-31核磁共振波谱学特征研究 周建威21003034/B030105 基于表面等离子体结构的WGM/SERS生物传感特性研究周吉21006022/B060802 定向嵌段共聚制备温度和pH双重敏感聚氨酯基智能材料周虎21001038/B010403 稀土纳米材料促小鼠成骨细胞增殖作用的分子机制研究周国强21027006/B040608 与显微结构研究集成的冷热台型高速扫描高灵敏热分析仪的研制周东山21071052/B010801 β-环糊精衍生物萃取分离对映体过程中的手性识别机理及构效关系周从山21073150/B030302 氨基多羧酸高效高选择性络合催化降解研究 周朝晖21001032/B010303 基于肟类桥连配体的单分子磁体的合成与性质研究 周爱菊21081220312/B060104 第四届中美“能源与环境:化学工程师的机遇与挑战”化工研讨会仲崇立21064004/B0403 壳聚糖新型衍生物作为农业杀菌剂研究 钟志梅21002104/B020203 基于共轭环状金属配合物的线型分子导线的合成与研究钟羽武21076195/B0608 大肠杆菌K5产肝素前体heparosan代谢控制研究 钟卫鸿21076194/B060409 基于Baylis-Hillman反应的手性膦杯芳烃的合成及催化性能研究钟为慧21006103/B060903 燃料电池用掺杂型非铂催化剂制备及其构效关系研究钟和香21074012/B0402 基于聚对苯撑乙炔分子链内环化反应构筑新型共轭高分子支俊格21073110/B030204 量子相空间动力学:轨线——密度函数方法 郑雨军21072067/B020506 基于有机分子聚集诱导发光特性的手性识别研究 郑炎松21073095/B030106 离子液体对TiO2的成核、物相及形貌的影响研究 郑文君21071062/B010403 新型有机硒化合物协同TRAIL诱导肿瘤细胞凋亡的分子机制研究郑文杰21003053/B0301 螺旋体为前体的配位聚合物的组装与动态组合化学库的建立郑盛润21077011/B070302 苯二氮类镇静催眠药物在A2/O工艺中的强化净化 郑少奎21037003/B07 典型工业过程中持久性有机污染物生成机制与控制原理郑明辉21006073/B060407 基于扩散层原位生长纳米碳纤维的燃料电池膜电极组件研究郑俊生21073129/B030608 锂离子电池中正负电极间的相互作用与机理研究 郑洪河91022011/B0103 含[MCuxSy] (M = Mo, W)功能基元的簇合物的合成及高阶非线性光学性能研究郑和根21071033/B0111 枝状结构硅纳米线的合成与高灵敏度生物传感器的制备郑耿锋21075085/B0503 多模式薄膜化学蒸气发生:装置、反应体系及应用 郑成斌21073228/B030105 硼掺杂TiO2光催化活性增强机制的固体NMR和量子化学计算研究郑安民21007062/B0701 近海海洋环境中PBDEs迁移转化机制研究 赵宗山21073235/B030301 FCC汽油选择性加氢脱硫新型L沸石基催化剂及反应机理研究赵震21006031/B060306 新型的ZIFs晶体膜的制备及其分离CO2/N2机理 赵祯霞21062024/B020102 DNA聚合酶抑制剂(+)-Aphidicolin全合成研究 赵元鸿21074081/B040101 多组分星形和接枝聚合物的合成及性能研究 赵优良21073114/B030301 氧化铝载体在含水加氢体系中的水合脱结构研究 赵永祥21005059/B050105 基于多功能免疫磁珠和微流控芯片的痕量循环肿瘤细胞检测赵永席91022022/B0211 低维有机微晶材料的设计、合成与光电性能研究 赵永生21074091/B040308 分子印迹中空纤维膜的制备及其对手性分子吸附与拆分性能的智能化调控赵义平21073146/B030203 具有多桥的复杂分子体系中的电子转移理论和应用 赵仪21075101/B0503 以纳米颗粒为载体的分子药物控制释放和监测体系研究赵一兵21003039/B030802 离子液体与有机分子的相互作用及其对若干化学反应选择性的影响赵扬21062030/B020506 新型桥联环糊精/DABO类HIV逆转录酶抑制剂包结配合物的超分子体系研究赵焱21066005/B060903 新型稀土/金属离子掺杂Ta2O5纳米电极材料的制备及在燃料电池中的应用赵彦宏21073216/B030503 氢/氘分子在微孔碳材料上的动力学量子筛分离机理的研究赵学波21076059/B061103 基于甘油氢解和CO2醇解反应过程集成的碳酸丙烯酯新绿色合成反应研究赵新强21005060/B050102 固定化功能蛋白质取向及构象表征和调控色谱新方法赵新锋21076040/B060905 锌离子调节酵母菌乙醇耐性的分子机制和胁迫耐受酵母的构建赵心清21007063/B0701 有机基团修饰的磁性纳米材料在复杂环境水样品分析中的应用研究赵晓丽21005091/B050206 分子印迹阵列传感器的研制及其在食品安全快速检测中的应用赵晓娟21073033/B0305 基于光子晶体编码微球的蛋白质SERS检测及其应用 赵祥伟21001043/B010101 具有超顺磁性的小粒径大孔径介孔药物载体的制备及性能研究赵文茹21075065/B050206 维持酶蛋白自然构型的新型生物传感器电极的研制及应用赵文波21072029/B020601 以天然生物碱冬青生菌素H为先导的药物设计、合成及生物活性研究赵圣印21075125/B0509 粒径单分散双孔型PGMA微球基质及小分子药物配基高效亲和色谱体系的研究赵睿21007035/B070102 竹炭样品处理技术分离检测痕量环境污染物及其作用机理研究赵汝松21061016/B010303 异核低维分子基磁性材料的合成和相关性质研究 赵琦华21062018/B0207 樟叶越橘中熊果苷咖啡酰基转移酶的基因克隆与功能分析赵平21077094/B0707 高污染电子拆解区多氯联苯孕产妇污染负荷及对胎盘功能影响的分子机理赵美蓉21072056/B020104 新型双官能有机膦催化剂的合成及应用 赵梅欣21076140/B060203 化学-生物耦合膜反应器的构建及其对水体中多氯联苯原位去除机理研究赵林21002057/B021102 基于氮杂杯芳烃的配位自组装及性能研究 赵亮21003158/B0302 气相过渡金属离子与有机分子若干基本反应的理论研究赵联明21062028/B020405 嗜盐放线菌新物种抗肿瘤活性代谢产物研究 赵立兴21003011/B030204 常温质子迁移反应分子力场的设计开发与应用研究 赵立峰21072068/B020703 分子设计的藻胆蛋白的光动力学效应及其光疗作用探索赵开弘21072164/B020102 二苯乙烯-苯丙木脂素类似物的不对称合成及其生理活性研究赵静峰21003113/B0302 TiO2 表面湿电子态及光化学反应的理论研究 赵瑾21073028/B030706 具有超长激发态寿命的环铂室温磷光染料的分子设计、合成、光物理性质与应用研究赵建章21011130154/B020506 检测糖类分子的模块化手性荧光分子探针的研究 赵建章21077132/B070303 假单胞菌ND6菌株高效降解萘和萘胁迫应答的分子机理赵化冰21077001/B070302 耦合离子交换功能的高分子絮凝剂及其去除水中小分子溶解性有机物的研究赵华章21006002/B060204 分子层面多元体系内纳微结构药物颗粒的自聚体构建机理研究赵宏21074058/B040101 侧链或侧基可控断裂的梳型聚合物的制备 赵汉英21001089/B0113 基于单分子转动产生铁电性的有机铁电体的合成与性能测试赵海霞21077077/B0703 电吸附调控-光催化氧化降解不透光污染体系的方法与协同机制赵国华21072115/B0206 以AKT为靶标的新型三取代吲哚衍生物的设计、合成及抗肿瘤活性研究赵桂森21005087/B0501 卷烟烟气中主要杂环胺类化合物的体内代谢研究 赵阁21032006/B02 含氟有机化合物的合成、反应规律及应用研究 赵刚21003071/B030301 纳米-亚纳米复合结构贵金属催化剂的合成与表征 赵丹21072117/B021101 新型硼桥联梯形pi-共轭分子材料的设计合成及性能研究赵翠华21075122/B0501 基于代谢组学的转基因植物的安全性评价研究 赵春霞21076093/B060804 中国红豆杉细胞合成多乙酰基紫杉烷的代谢规律及调控机制赵春芳21076219/B061103 高纯纳米Al13溶胶制备大孔拟薄水铝石及其转化机制研究赵长伟21073035/B030201 负载型前过渡金属氧化物团簇模型催化剂构效关系的理论研究章永凡21072096/B020901 新型烯丙基香豆素类分子的设计与合成及其抗菌活性研究章维华21076052/B060702 糖分子内P、N手性配体的设计、制备及在不对称合成中的应用章鹏飞21072173/B020706 基于双AAO酶的手性汇聚法研究 张子张21073224/B0305 基于金属纳米颗粒-氧化石墨烯复合体系的表面增强拉曼基底的构筑及其用于细胞与药物相互作用研究 张智军21002051/B0201 炔丙基全碳1,3-偶极子在构建呋喃并杂环衍生物中的应用张志国21076126/B060301 强极性流体的气液相平衡行为及其应用 张志刚21077043/B070301 基于转化频率的碳烟催化燃烧机制张昭良21073162/B030602 纳米结构CeO2薄膜的光助阳极电沉积机制及其腐蚀电化学行为研究张昭21072042/B020101 新型环状过氧化物的合成方法研究张占辉21006075/B0603 微波辅助离子液体提取天然产物有效成分的传质机理研究张越非21076143/B060306 磷酸促进型掺锆二氧化硅/聚合物杂化膜及性能的研究张裕卿21071096/B010303 配合物嫁接的具有"二合一"功能的纳米复合材料 张玉良21072169/B0202 过渡金属催化的C-O键活化反应研究张玉红21074020/B040101 聚丙烯腈基碳纳米微球的可控制备及其基础问题研究张幼维21075102/B050303 金银纳米粒子增强室温磷光机理及其应用研究 张勇21001097/B010701 陀螺状磷酸亚铁锂/碳纳米管复合材料的结构调控及电化学特性研究张勇21066009/B061201 富氧增强褐煤燃烧性能及其在水泥熟料煅烧中的应用研究张永锋21005020/B0505 介孔纳米材料富集质谱鉴定糖肽新方法用于大肠癌血清糖肽组的研究张莹21076207/B060806 细胞间质体外模拟系统的构建及物质传递基本规律的研究张英21002092/B020405 白蚁共生放线菌的抗菌成分研究张应烙21073003/B030106 石墨烯表面化学修饰以及量子尺寸效应研究 张艳锋21005086/B050202 环介岛等温基因扩增过程的电化学信息研究张旭志21075132/B0504 14N核磁共振研究赖氨酸三甲基化蛋白质的相互作用动力学张许21074141/B040607 烯烃嵌段共聚物及其与聚烯烃共混体系的结构与性能关系研究张秀芹21010402027/B030201 过渡金属二茂夹心卤化物的国际合作研究 张秀辉21075029/B0502 基于分子印迹聚合物固相微萃取β-兴奋剂电化学传感器的研究张修华21001103/B0107 电沉积制备石墨烯/导电聚合物复合薄膜电极材料及其电容特性张熊21076030/B060306 优先渗透分离CO2的ZIF型金属有机骨架结构类分子筛膜的设计制备研究张雄福21076188/B061102 低温氮等离子体改性碳纳米管吸附二氧化碳的基础研究张兴旺21002117/B020601 靶向Mcl-1蛋白的抗肿瘤药物先导化合物的优化改造 张兴龙21074106/B0401 基于CO2/环氧化物可控共聚技术合成可降解的聚合物分子刷张兴宏21002070/B0201 钯催化多组分环化反应合成含硫杂环化合物的研究 张兴国21005010/B050304 LED诱导化学发光适配体传感器张信凤21077070/B070701 丁二烯代谢产物1-氯-3-丁烯-2-醇和1-氯-3-丁烯-2-酮与DNA碱基反应的研究张新宇21073061/B030606 纳米碳纤维的微结构及表面性质与其电催化活性的内在规律研究张新胜21027013/B0505 小型质谱仪器关键技术研究张新荣21072002/B020401植物甾醇单体氧化物的制备及其细胞毒性的研究张欣21007025/B070403 从芳香烃受体(AhR)的基因型研究卤代芳烃污染物的鸟类种间敏感差异性张效伟21010302032/B0509 2010 年第四届生物分析、生物医学工程与纳米技术国际会议(ISBBN 2010)张晓兵21072025/B0211 新型甲基酮桥连的给体-共轭-受体类分子电子开关的研究张晓安21003079/B0306 染料敏化太阳能电池中染料的分子设计、合成与光电性能研究张晓21073001/B030607 金属基复合材料电极的构建及其在肼类化合物中的检测应用张小俊21036007/B061201 离子液体及其在清洁化工中的应用基础研究 张香平21007066/B070403 六溴环十二烷3种异构体在鱼体内代谢过程研究 张娴21071030/B01 介电可调的有机-无机杂化包合物 张闻21076130/B060805 皮胶原-有机交联剂反应机理的计算化学研究 张文华21075041/B0502 半导体复合纳米材料光电分析方法在乙酰胆碱酯酶活性检测中的应用研究张文21076012/B060306 膜吸收近膜壁面处的传质行为及其传质强化手段研究张卫东21007023/B070302 树脂基纳米M/Fe(0)的研制及其催化脱除硝酸根的研究张炜铭21003097/B030106 高效可控制备微/纳米条形码的新方法研究 张伟明21074035/B040501 新型特定结构功能性β-环糊精包结络合超分子聚合物的构建、形态和应用张伟安21075039/B050106 磁性纳米固定相毛细管电色谱分离系统研究 张维冰21074059/B040106 疏水单体在水中的RAFT聚合:壳-冠型空心微球反应器中的RAFT聚合研究张望清21003119/B030301 IrMOx双活性位催化剂CO选择氧化微观动力学研究 张万生21077134/B070201 城市大气碳质颗粒物综合测定及七组分来源研究 张烃21071007/B010404 钙化结节中钙磷酸盐的结构及其转变——内、外源性金属离子的作用张天蓝21081260197/B06 第三届两岸化学工程与产品工程高峰研讨会 张锁江21072143/B020101 低价稀土金属试剂在合成多烯及多官能团化合物中的应用研究张松林21075021/B050901 微流控体系下DNA-蛋白质相互作用的单分子检测技术研究张松21077095/B070101 环境水体中超痕量溴酸根的在线富集、聚焦和毛细管电泳分离分析研究张书胜21007006/B070302 膜曝气分离单级自养脱氮生物反应器过程控制和功能菌群研究张寿通21004048/B040309 聚丁二酸丁二醇酯的仿生改性及其仿细胞外层膜结构纳米胶束的研究张世平21001036/B010701 具有多种聚阴离子基团的锂离子电池正极材料的结构调控及脱嵌锂性能研究张森21072156/B020601 新型激酶抑制剂:8-羟基-2-芳基-1-异喹啉酮类的合成和抗肿瘤活性研究张三奇21004021/B040308 利用非氟嵌段聚合物和超临界二氧化碳制备纳米孔径的高分子功能化薄膜张锐21003144/B030204 汽车尾气催化剂Pt掺杂CaTiO3的自再生机理的计算研究张秋菊21006108/B060402 转鼓式生物浸出反应器的传递特性和放大规律研究 张庆华21002107/B021102 新型“类离子液体”软功能材料制备与性能研究 张庆华21003081/B030802 离子液体与极性溶剂间相互作用规律研究 张庆国21001061/B010303 具有化学传感功能的多孔发光MOFs材料的设计构筑及其检测大气中POPs的研究张庆富21074123/B0403 单个偶氮聚合物囊泡的光致可逆形变的放大研究 张其锦21062013/B0210 四氯化硅催化氢化制备三氯氢硅工艺及机理研究 张宁21006023/B060304 扩张床吸附原位提取中药有效成分的方法研究 张敏21007083/B0705 环境渐变过程实验模拟及初始浓度效应研究 张美一21005006/B0502 基于扫描电化学显微镜的指纹采集技术的研究 张美芹21006069/B060802 “智能”溶栓策略探索——多尺度分子动力学模拟 张麟21076176/B060306 含无机纳米水通道反渗透复合膜的结构设计与制备 张林21076107/B061202 微小空间反应器中生物激发合成沸石与纳米金属粒子的研究张利雄21071032/B010303 新型季铵盐功能化的金属有机骨架材料的合成、结构与性能研究张丽娟21027002/B0501 蛋白质样品多级预处理系统的研制 张丽华21073177/B0307 甲醛分子离子电子激发态的振动分辨光解离动力学研究张立敏21073069/B030304 自掺杂光催化剂的设计、制备及其可见光光催化性能研究张礼知21074152/B0404 含树枝化结构基元的新型聚多糖衍生物研究 张黎明21001004/B0107 功能氧化物/碳纳米纤维复合材料的制备和电催化性能研究张莉21075016/B0501 多维毛细管液相色谱-质谱联用新技术用于甲型H1N1流感病毒的研究张兰21002115/B0203 环状手性含氟亚砜亚胺、亚磺酰胺和磺酰胺的立体专一性合成及应用研究张来俊21003050/B030301 限域纳米空间内酸碱有机官能团分子的可控组装及其协同催化机理的研究张坤21076162/B0608 油包水乳化体系中新型交联酶聚集体的构建及其结构与性能张峻21074048/B040605 具有不对称构筑基元的微结构阵列及其各向异性光学性质张俊虎21076063/B061201 基于离子液体的氯硅烷催化反应-相控耦合分离研究 张军21006110/B060901 炭黑和气体组分在煤气化过程中的作用机理研究 张聚伟21004001/B040601 停流光谱技术研究阳离子聚合物/DNA络合与解络合动力学张璟焱21076144/B060404 气液固三相高剪切反应器性能与模型放大研究 张金利21001120/B010303 多孔金属多氮唑框架 张杰鹏21073191/B0301 基于非手性源的单手性材料的催化不对称结晶 张健21002062/B020601 STAT3选择性抑制剂的设计、优化及其功能研究 张健21004077/B040303 基于主-客体相互作用的三重化学响应性聚合物组装体:设计、构建及其药物传输性能研究张建祥21074063/B040401 微量填充生物可降解高分子共混体系的微结构与性能研究张建明21073053/B030803 桥联型双核稀土多取代芳香羧酸配合物结构与热分解反应机理及性能研究张建军21071025/B010303 异金属团簇化合物的合成和性能研究 张建军21071019/B010601 二元金属氨硼烷的制备、释氢机理及其在推进剂中应用研究张建国21061004/B010701 含Fe钨青铜结构化合物的结构调控与电磁特性研究 张辉21071021/B010902 锝-99m、氟-18、碘-125-VEGF多肽肿瘤显像剂的制备、体外评价及生物分布研究张华北21073055/B030802 溶液中无机阴离子在纳米晶表面上的复合与稳定化作用研究张虎成21073077/B0304 有机光电材料激子态动力学研究 张厚玉21071140/B0105 新型稀土纳米复合材料的合成、发光和磁性能的研究张洪杰21074071/B040609 透明质酸多糖自聚集物理水凝胶的形成机理和流变学研究张洪斌21071027/B0101 功能化的多金属氧酸盐组装分子磁性晶态材料的可控合成与性能研究张宏21007069/B0704 T-2毒素生物转化及毒性的分子机制研究 张红霞21005067/B050105 集成化多功能可控细胞操纵及分析微流控芯片的研制张何21073079/B030702 高双光子吸收截面有机半导体材料的设计与性质研究张浩力21004032/B040102 新型烯烃复分解催化剂的设计、合成以及在制备结构可控的高性能聚合物材料中的应用张浩21077030/B0704 固氮蓝藻修复多氯联苯污染水稻土的机理研究 张杭君21077102/B0706 大辽河流域代表性卤代阻燃剂的污染特征与演变趋势张海军21006066/B060409 CH4/CO2重整高抗积碳金属/炭材料催化剂的制备及机理研究张国杰21007002/B070301 三维有序大孔-介孔复合氧化物原位担载贵金属纳米粒子的可控制备及同时消除NOx和碳烟的催化性能研究张桂臻21081260019/B020402 中国西北部中药资源开发国际研讨会 张桂珍21076215/B060802 层析过程中界面上蛋白质结构及动态变化 张贵锋21071146/B0107 多酸基多元复合光电催化材料的设计,制备及其性能研究张光晋21075126/B0509 基于聚集荧光增强机理的化学/生物传感的研究 张关心21076095/B0608 慢消化淀粉与茶多酚对餐后血糖反应的协同作用 张根义21073105/B030505 基于alpha-烷氧基锌酞菁J聚集机理的酞菁光控自组装张复实21077120/B070304 含溴电子废物在超临界甲醇中的催化脱溴机制研究 张付申21064002/B040705 介孔材料环境下原位乳液聚合稳定性及其聚合物复合材料热学和力学性能研究张发爱21072226/B020601 基于小檗碱抗耐药真菌作用的小分子探针研究 张大志21003077/B030301 以废轮胎热解炭为载体的脱氢催化剂在有机液体储氢中的研究张翠21075129/B050901 病原体的超灵敏高通量单分子检测研究 张春阳21027007/B0506 电化学发光成像分析仪的研制 张成孝21005030/B0511 基于碳纳米管表面印迹技术的猕猴桃根中抗肿瘤活性成分分离及活性研究张朝晖21004080/B040303 组织诱导型可生物降解聚谷氨酸水凝胶支架材料制备及其在骨组织工程中的应用研究张超21077117/B070301 Pt/TiO2催化剂室温氧化甲醛的高活性机制研究及非贵金属化探索张长斌21005065/B050102 基于液滴技术的蛋白质组分离分析新方法 张博21077126/B070502 典型羟基多溴联苯醚拟/抗激素效应的H12定位选择机制及构效关系研究张爱茜21073087/B0303 多壁碳纳米管的结构缺陷及其自发氧化还原性能在催化反应中的作用研究张爱民21034004/B040101 大尺度螺旋聚合物的可控合成及其结构分析 张阿方21075077/B0503 痕量多溴联苯醚的表面增强拉曼光谱检测 占金华21072159/B0201 过渡金属催化下各类杂环化合物的新合成方法研究 詹庄平21076184/B060702 持久低表面能、环境友好含短氟碳链聚合物的分子设计与合成詹晓力。
分子生物学试题试卷3

一、名词解释1.SD序列(Shine-Dalgarno sequence):mRNA中用于结合原核生物核糖体的序列。
SD序列在细菌mRNA起始密码子AUG上游7-12个核苷酸处,有一段富含嘌呤的碱基序列,能与细菌16SrRNA3’端识别,帮助从起始AUG处开始翻译。
2.E位点:是脱氨酰tRNA(deaminoacyl-tRNA)离开A位点到完全从核糖体释放出来的一个中间停靠点,只是作暂时的停留。
当E位点被占据之后,A位点同氨酰tRNA的亲和力降低,防止了氨酰tRNA的结合,直到核糖体准备就绪,E位点腾空,才会接受下一个氨酰tRNA3.P位点:即肽酰tRNA位点(peptidyl-tRNA site), 又叫供位(donor site), 或肽酰基位点,主要位于大亚基, 是肽基tRNA移交肽链后肽酰tRNA所占据的位置, 即与延伸中的肽酰tRNA结合位点。
4.A位点:即氨酰基位点,是与新掺入的氨酰tRNA(aminoacyl-tRNA )结合的位点,又叫受位(entry site),主要位于大亚基,是接受氨酰tRNA的部位5.Kozak序列:是位于真核生物mRNA 5’端帽子结构后面的一段核酸序列,通常是ACCACCAUGG,它可以与翻译起始因子结合而介导含有5’帽子结构的mRNA翻译起始。
对应于原核生物的SD序列。
6.内部核糖体进入位点:缩写IRES,是一段核酸序列,它的存在能够使蛋白质翻译起始不依赖于5‘帽结构,从而使直接从信使RNA(mRNA)中间起始翻译成为可能。
通常来讲,真核生物翻译只能从mRNA的5‘端开始,因为翻译起始必须依赖于5’端的帽子结构。
一般来讲,内部核糖体进入位点通常位于RNA病毒基因组的5’非翻译区(UTR),这样病毒蛋白的翻译就可以不依赖于5‘帽子结构。
7.上游可读框:mRNA前导序列中的可读框,可作为顺式调节元件将mRNA的翻译速率与氨基酸水平相偶联8.分子伴侣:是细胞中一大类蛋白质,是由不相关的蛋白质组成的一个家系,它们介导其它蛋白质的正确装配,但自己不成为最后功能结构中的组分9.核定位序列(Nuclear localization sequence):蛋白质的一个结构域,通常为一短的氨基酸序列,它能与入核载体相互作用,使蛋白能被运进细胞核。
核酸提取常见试剂地作用原理

核酸提取常见试剂地作用原理核酸提取是分子生物学研究中的基础步骤之一,常用于从细胞或组织中提取和纯化核酸。
核酸提取试剂包括细胞裂解缓冲液、蛋白酶、盐溶液、有机溶剂等。
核酸提取试剂的作用原理主要涉及以下几个方面:1.细胞裂解缓冲液:细胞裂解缓冲液是为了打破细胞膜和核膜,使细胞内的核酸暴露出来。
细胞裂解缓冲液一般含有一些离子和试剂,如EDTA、SDS等。
EDTA可以螯合离子,使核酸不被酶降解。
SDS则可以破坏细胞膜和核膜的完整性,使核酸释放出来。
2. 蛋白酶:蛋白酶的作用是降解核酸分子上的蛋白质,使核酸与蛋白质分离。
在核酸提取的过程中,蛋白酶可以去除核酸表面的蛋白质,从而提高核酸的纯度。
常用的蛋白酶有蛋白酶K和蛋白酶ase。
3.盐溶液:盐溶液的作用是使DNA在溶液中凝集并沉淀。
盐的高浓度可以中和DNA的负电荷,使DNA之间的静电斥力减弱,从而导致DNA分子的凝集和沉淀。
4.有机溶剂:有机溶剂在核酸提取中起到萃取作用,使DNA从细胞裂解液中分离出来。
有机溶剂常用的有酚类、氯仿和异丙醇。
酚类溶剂可以破坏细胞和核膜的完整性,同时与DNA形成无水醇相互作用,促使DNA分离出来。
氯仿可以和DNA结合,并与水进行分配,从而加速DNA的沉淀。
异丙醇则可改变溶液的表面张力,使DNA分子凝胶化和沉淀。
5.离心:离心是核酸提取中的一个重要步骤。
通过离心,可以将细胞碎片、蛋白质、碎屑等杂质与纯净的核酸分离开来。
离心的原理是通过旋转离心机,利用离心力将混合液中的成分分离开来,使沉淀物和上清液分离。
总的来说,核酸提取试剂是通过改变细胞的环境和物理化学性质来实现核酸的分离纯化。
通过使用细胞裂解缓冲液打破细胞膜和核膜,加入蛋白酶降解蛋白质,使用盐溶液凝集和沉淀DNA,加入有机溶剂进行DNA的萃取,最后通过离心分离杂质和DNA。
这些步骤综合起来,能够从复杂的样品中纯化出相对纯净的核酸。
蛋白质与核酸的相互作用核酸结合蛋白模板
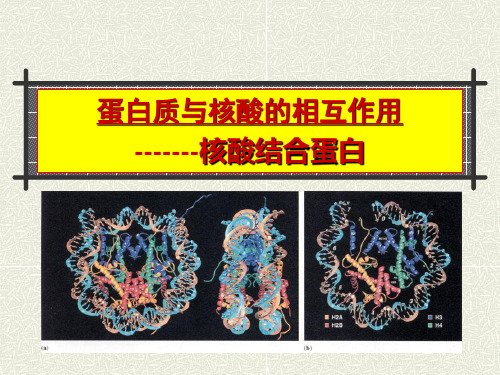
3.2.3 锌指结构的特点
Cys2His2锌 指蛋白与DNA 形成复合物的 X-射线晶体衍 射图谱。 三个锌指以 半环状排列于 DNA的大沟中。
3.2.3 锌指结构的特点
雌激素受体 (ER) DNA结 合结构域与 DNA识别因子 配位的同二聚 体。其中四个 圆代表二聚体 中的四个Zn 离子。
RNA结构的特点:胞内RNA一般呈单链结构,但往往 折叠成各种二级结构(突起、发夹、茎环等)。
RNA结合蛋白中的基本结构
结合结构 核糖核酸蛋白结 构域 dsRBD 结合部位 β-折叠 β-折叠 分布 真核生物 所有生物 举 例 U1A snRNP 果蝇的Staufen蛋白
K-同源蛋白
环区
真核生物
6.3 解读蛋白中的氨基酸
部分替换:用基因工程方法替换结构域中的某 些残基,研究其对与DNA结合的重要性。 结构分析:用X-射线、NMR方法研究发现,在 DNA和蛋白质结合过程中,蛋白质和DNA的构 象发生了适宜性的变化,水分子在蛋白和DNA 的相互作用中也发挥了特殊的作用。
6.4 假定的锌指蛋白DNA识别密码
目前还没有发现一套普遍的密码适用于所有的蛋白质 和氨基酸,但在锌指蛋白中发现了一个初步的规律。 锌指蛋白氨基酸残基与DNA碱基对应关系
3’
T A G
5’
与Zif268相关的锌指蛋白的部分DNA识别密码
三联体密码中碱基的位置
碱基
A C
5’
中部
3—Asn 3—Asn,Leu,Thr,Val
3’
-1—Gln+2--Ala
类固醇受体家 族 碱性结构域 带状-螺旋-螺旋 组蛋白-核心
α -螺旋
α -螺旋
真核生物
真核生物
生物大分子的纯化与鉴定技术

生物大分子的纯化与鉴定技术生物大分子是生命体内最基本的组成元素之一,包括蛋白质、核酸、多糖和脂质等。
它们的结构和功能对于生物体的发育、代谢、传递遗传信息等方方面面都有着非常重要的作用。
因此,对它们进行纯化和鉴定是生物学和生命科学研究中不可或缺的重要步骤。
一、蛋白质的纯化与鉴定技术1. 活性层析技术活性层析是从混合样品中纯化蛋白质的一种常用技术。
它基于蛋白质与特定配体之间的互相作用,利用这种相互作用把想要纯化的蛋白质从混合物中分离出来。
这种方法不仅可以分离出单一种类的蛋白质,还可以根据蛋白质与配体的亲和性进行分层次纯化。
同时,利用不同的配体也能够分离出不同功能的酶,从而进一步扩大了对蛋白质的纯化范围。
2. 离子交换层析技术离子交换层析是一种基于蛋白质电荷的分离方法。
它利用固定在树脂表面上的离子,通过与蛋白质表面的离子相互作用,将蛋白质从混合物中分离出来。
这种方法常常用于分离带有不同电荷的蛋白质,以及酸性和碱性细胞因子等物质。
3. 尺寸排除层析技术尺寸排除层析技术是一种基于蛋白质大小的分离方法。
它通过让大分子在固定相中的孔隙中滞留时间长,从而将大分子和小分子分离出来。
这种方法通常用于分离相对分子质量较大的蛋白质,如重组蛋白、抗体等。
4. 逆相高效液相色谱技术逆相高效液相色谱是一种基于蛋白质亲水性的分离方法。
它利用逆相柱的反相作用,将亲水性较小的蛋白质从混合物中分离出来。
这种方法常常被用于提纯高表达体系中的蛋白质。
5. SDS-PAGE和Western Blotting技术SDS-PAGE是一种基于蛋白质质量和电荷的分离技术,通过在凝胶中加入SDS(十二烷基硫酸钠)和还原剂,可以使不同电荷和大小的蛋白质变得相同,从而进行准确的大小分离。
Western Blotting是一种检测蛋白质表达的方法,它利用特异性抗体将蛋白质分子分离出来,并将其转移到膜上,然后通过特异性抗体进一步检测目标蛋白质的表达量。
二、核酸的纯化与鉴定技术1. 常规离心技术常规离心技术是一种对复杂混合物进行分离和预纯化的方法,通过调整离心速度和离心时间,将不同大小和形状的细胞组分分离出来。
质谱分析蛋白质的相互作用

蛋白质间的相互作用存在于生物体每个细胞的生命活动过程中,互交叉形成网络,成细胞中一系列重要生理活动的基础。
其中,多数蛋白质是通过与配体分子结合或者是作为1个大的生物复合体的一部分,与细胞完整性维持、遗传物质复制、基因表达调控、信号转导、免疫应答等一系列生命过程。
研究蛋白质间相互作用的方式和程度,将有助于蛋白质功能的分析、疾病致病机理的阐明和治疗和新型药物的开发等众多难题的解决。
因此,确定蛋白质间相互作用关系、绘制相互作用图谱已成为蛋白质组学研究的热点。
近年来有许多方法被用于蛋白质相互作用的研究,酵母双杂交技术,免疫共沉淀技术,串联亲和纯化技术,化学交联技术,蛋白质芯片技术,荧光共振能量转移技术,噬菌体展示技术等。
酵母双杂交技术Fields和song等首先在研究真核基因转录调控中建立起来的,是在真核细胞中检测蛋白质与蛋白质之间的相互作用的方法。
该系统是通过两个分别称之为“诱饵蛋白”和“捕获蛋白”的融合蛋白形成一个完整的转录激活因子,从而激活报告基因的表达,通过在营养缺陷型培养基上生长或呈现显色反应来检测系统的功能。
酵母双杂交系统可在全基因组规模上进行蛋白质一蛋白质相互作用高通量的研究。
免疫共沉淀技术免疫共沉淀是利用抗原和抗体的特异性结合以及细菌的Protein A或G特异性地结合到免疫球蛋白的Fc片段的现象开发出来的方法。
其基本原理是,在细胞裂解液中加入抗兴趣蛋白的抗体,孵育后再加入与抗体特异结合的结合于Agarose珠上的Protein A或G,若细胞中有与兴趣蛋白结合的目的蛋白,就可以形成这样一种复合物:“目的蛋白—兴趣蛋白—抗兴趣蛋白抗体—Protein A或G”,经变性聚丙烯酰胺凝胶电泳,复合物又被分开。
然后经免疫印迹或质谱检测目的蛋白。
这种方法得到的目的蛋白是在细胞内与兴趣蛋白天然结合的,符合体内实际情况,得到的结果可信度高。
这种方法常用于测定两种目标蛋白质是否在体内结合;也可用于确定一种特定蛋白质的新的作用搭档。
核酸提取液作用原理

核酸提取液作用原理
核酸提取液是一种用于从生物样本中提取核酸(DNA或RNA)的化学试剂。
它的作用原理是通过破坏细胞膜和核酸蛋白质的相互作用,使核酸从细胞中释放出来,并且保护核酸免受酶的降解。
核酸提取液通常包含离子表面活性剂(如SDS或CTAB)、蛋白酶K、EDTA、盐和缓冲液等成分。
这些成分共同发挥作用,实现核酸的高效提取。
核酸提取液中的离子表面活性剂会破坏细胞膜的完整性。
离子表面活性剂具有亲水和疏水两个部分,可以相互作用并与细胞膜脂质结合,破坏脂质屏障,使细胞膜破裂。
同时,离子表面活性剂还可以与细胞膜蛋白质结合,破坏蛋白质的三维结构,进一步破坏细胞膜。
核酸提取液中的蛋白酶K可以降解蛋白质。
蛋白酶K是一种嗜热性蛋白酶,在高温条件下能够高效降解细胞中的蛋白质。
通过加入蛋白酶K,可以迅速降解细胞中的蛋白质,将核酸从蛋白质中释放出来。
核酸提取液中的EDTA可以螯合金属离子,抑制核酸酶的活性。
核酸酶是一类能够降解核酸的酶,在细胞内广泛存在。
通过加入EDTA,可以使其与金属离子结合,降低核酸酶的活性,从而保护核酸不被降解。
核酸提取液中的盐和缓冲液可以调节提取液的pH值和离子强度,为
核酸的提取提供合适的环境。
适当的pH值和离子强度可以使核酸分子更容易溶解在提取液中,并且有助于去除细胞中的蛋白质和其他杂质。
核酸提取液通过破坏细胞膜和核酸蛋白质的相互作用,释放核酸,并且通过蛋白酶的降解和核酸酶的抑制,保护核酸不被降解。
核酸提取液的使用可以方便、高效地从生物样本中提取出纯度较高的核酸,为后续的分子生物学研究和应用提供了重要的基础。
emsa凝胶迁移实验原理

emsa凝胶迁移实验原理什么是emsa凝胶迁移实验原理EMSA(Electrophoretic Mobility Shift Assay)是一种常见的核酸-蛋白质相互作用研究技术。
它通过利用蛋白质与核酸结合形成复合物的特性,在电场下进行凝胶电泳迁移,从而分析和鉴定核酸与蛋白质之间的相互作用。
这个过程的实质是先将核酸与蛋白质按照一定比例混合反应,形成核酸-蛋白质复合物,然后将这个复合物与未结合的核酸分子一起进行凝胶电泳。
核酸-蛋白质复合物迁移速度较慢,而游离核酸迁移速度较快,因此可以通过观察凝胶中的带状图案来判断核酸与蛋白质是否发生相互作用。
EMSA凝胶迁移实验原理包括以下几个关键步骤:1. 核酸与蛋白质的结合:首先,需要获得待检测蛋白质所特异结合的核酸序列。
这个核酸序列可以是DNA、RNA或核酸伸展结构。
然后,将核酸与蛋白质按照一定比例混合,在适当的反应条件下进行结合。
常用的反应条件包括缓冲液pH值、离子浓度、温度等。
2. 核酸-蛋白质复合物的分离:混合物中的核酸-蛋白质复合物通常比未结合的核酸分子具有较大的分子量,迁移速度较慢。
因此,可以通过凝胶电泳来将复合物与游离的核酸分子分离开来。
常用的凝胶材料包括聚丙烯酰胺凝胶和琼脂糖凝胶。
3. 凝胶迁移:将经蛋白质和核酸反应后的混合物加载到凝胶中,然后通过应用电场使核酸与蛋白质复合物迁移。
电场的作用是带动带电的分子在凝胶中移动,移动速度与其电荷量和形状有关。
4. 分析结果:通过凝胶电泳的分离,可以得到带状图案。
观察不同带状图案的位置、强度和大小可以判断蛋白质与核酸的结合情况。
当蛋白质与核酸发生结合时,形成的复合物迁移速度较慢,带状图案位置上移或移动较慢;反之,当蛋白质与核酸未结合时,带状图案位置下移或迁移速度较快。
需要注意的是,EMSA凝胶迁移实验只能证明核酸与蛋白质之间是否存在相互作用,无法提供相互作用的具体结合位点信息。
因此,为了进一步研究相互作用的机制,通常需要结合其他技术手段,如DNase I酶切、ChIP-Seq等。
核酸在蛋白质生物合成中的作用

核酸在蛋白质生物合成中的作用1.引言在细胞内,核酸和蛋白质是两种重要的生物分子,它们在生物体内具有各种不可替代的功能。
本文将探讨核酸在蛋白质生物合成中的作用。
2.核酸与蛋白质的功能2.1核酸的基本结构和功能核酸是由核苷酸组成的,核苷酸由糖分子、碱基和磷酸组成。
核酸分为D NA(脱氧核酸)和R NA(核糖核酸)两种类型。
D NA具有存储遗传信息的功能,而RN A则参与转录和翻译等生物合成过程。
2.2蛋白质的基本结构和功能蛋白质是由氨基酸组成的,氨基酸通过肽键连接形成多肽链,进而折叠成特定的三维结构。
蛋白质在生物体内具有结构支持、催化酶、运输、抗体等多种重要功能。
3.核酸在蛋白质生物合成中的作用核酸在蛋白质生物合成过程中发挥着关键的作用,主要包括转录和翻译两个过程。
3.1转录转录是指在细胞核内,D NA的信息通过RN A的合成被复制到R N A分子上的过程。
在这一过程中,核酸D NA作为模板被RN A聚合酶酶原识别并逐个配对与合成核苷酸的RN A链。
3.2翻译翻译是指根据RN A上的遗传信息,将氨基酸按照特定的顺序组装成蛋白质的过程。
这一过程由核糖体催化完成,核酸m RN A作为模板被tR NA 识别并配对与合成相应的氨基酸。
4.核酸在蛋白质生物合成中的调控核酸在调控蛋白质合成过程中发挥了重要的作用。
4.1转录调控转录调控是指在转录过程中,通过调节DN A和R NA聚合酶或其他蛋白质的相互作用,从而控制基因表达水平的一系列过程。
这一过程可以通过核酸结构的改变或与特定蛋白质的结合来实现。
4.2翻译调控翻译调控是指在翻译过程中,通过调节核糖体和t RN A或其他蛋白质的相互作用,从而影响蛋白质的合成速率和选择性的一系列过程。
这一过程可以通过核酸序列的特殊性或与特定蛋白质的结合来实现。
5.结论核酸在蛋白质生物合成中起到了重要的角色,通过转录和翻译过程参与了蛋白质的合成和调控。
进一步的研究将有助于揭示核酸与蛋白质之间更为复杂的相互作用及其在生物体内的功能机制。
生物化学第五节 生物大分子相互作用研究技术

第五节生物大分子相互作用研究技术2015-07-16 70976 0生物大分子之间可相互作用并形成各种复合物,所有的重要生命活动,包括DNA的复制、转录、蛋白质的合成与分泌、信号转导和代谢等,都是由这些复合物所完成。
研究细胞内各种生物大分子的相互作用方式,分析各种蛋白质、蛋白质-DNA、蛋白质-RNA复合物的组成和作用方式是理解生命活动基本机制的基础。
有关研究技术发展迅速,本节选择性介绍部分方法的原理和用途。
一、蛋白质相互作用研究技术目前常用的研究蛋白质相互作用的技术包括酵母双杂交、各种亲和分离分析(亲和色谱、免疫共沉淀、标签蛋白沉淀等)、FRET效应分析、噬菌体显示系统筛选等。
本部分简要介绍标签蛋白(tagged protein)沉淀和酵母双杂交技术(yeast two-hybrid system)。
(一)标签蛋白沉淀标签融合蛋白结合实验是一个基于亲和色谱原理的、分析蛋白质体外直接相互作用的方法。
该方法利用一种带有特定标签( tag)的纯化融合蛋白作为钓饵,在体外与待检测的纯化蛋白或含有此待测蛋白的细胞裂解液温育,然后用可结合蛋白标签的琼脂糖珠将融合蛋白沉淀回收,洗脱液经电泳分离并染色。
如果两种蛋白有直接的结合,待检测蛋白将与融合蛋白同时被琼脂糖珠沉淀( pull-down),在电泳胶中见到相应条带(图20-6)。
图20-6 标签融合蛋白沉淀实验流程示意图目前最常用的标签是谷胱甘肽S-转移酶( GST),有各种商品化的载体用于构建GST融合基因,并在大肠杆菌中表达为GST融合蛋白。
利用GST与还原型谷胱甘肽(glutathione)的结合作用,可以用共价偶联了还原型谷胱甘肽的琼脂糖珠一步纯化GST融合蛋白。
另一个常用的易于用常规亲和色谱方法纯化的标签分子是可以与镍离子琼脂糖珠结合的6个连续排列组氨酸( 6xHis)标签。
标签融合蛋白结合实验主要用于证明两种蛋白分子是否存在直接物理结合、分析两种分子结合的具体结构部位及筛选细胞内与融合蛋白相结合的未知分子。
蛋白质与核酸的定性与定量实验方法资料

蛋白质与核酸的定性与定量一、实验目的1、学习和掌握纯化蛋白质的原理和方法、蛋白质等电点的测量原理和方法。
2、进一步掌握使用双缩脲法对蛋白质的定性测定、利用定糖法对核酸的定性与定量测定二、实验原理1、蛋白质的定性测定:双缩脲法,课本P992、蛋白质的定量测定:Folin-酚法,实验P193、核酸的定性与定量测定:定糖法,课本P131、4、蛋白质等电点的测量在IEF的电泳中,具有pH梯度的介质其分布是从阳极到阴极,pH值逐渐增大。
如前所述,蛋白质分子具有两性解离及等电点的特征,这样在碱性区域蛋白质分子带负电荷向阳极移动,直至某一pH位点时失去电荷而停止移动,此处介质的pH恰好等于聚焦蛋白质分子的等电点(pl)。
同理,位于酸性区域的蛋白质分子带正电荷向阴极移动,直到它们的等电点上聚焦为止。
可见在该方法中,等电点是蛋白质组分的特性量度,将等电点不同的蛋白质混合物加入有pH梯度的凝胶介质中,在电场内经过一定时间后,各组分将分别聚焦在各自等电点相应的pH位置上,形成分离的蛋白质区带pH梯度的组成pH梯度的组成方式有二种,一种是人工pH梯度,由于其不稳定,重复性差,现已不再使用。
另一种是天然pH梯度。
天然pH梯度的建立是在水平板或电泳管正负极间引入等电点彼此接近的一系列两性电解质的混合物,在正极端吸入酸液,如硫酸、磷酸或醋酸等,在负极端引入碱液,如氢氧化钠、氨水等。
电泳开始前两性电解质的混合物pH为一均值,即各段介质中的pH 相等,用pH0表示。
电泳开始后,混合物中pH最低的分子,带负电荷最多,pI1为其等电点,向正极移动速度最快,当移动到正极附近的酸液界面时,pH突然下降,甚至接近或稍低于PI1,这一分子不再向前移动而停留在此区域内。
由于两性电解质具有一定的缓冲能力,使其周围一定的区域内介质的pH保持在它的等电点范围。
pH稍高的第二种两性电解质,其等电点为pI2,也移向正极,由于pI2>pI1,因此定位于第一种两性电解质之后,这样,经过一定时间后,具有不同等电点的两性电解质按各自的等电点依次排列,形成了从正极到负极等电点递增,由低到高的线性pH梯度。
蛋白质与生物大分子的相互作用研究

蛋白质
01
是由氨基酸组成的大分子,具有复杂的空间结构和功能,是生
命活动中不可或缺的组成部分。
生物大分子
02
是指细胞内存在的除蛋白质以外的其他大分子物质,如核酸、
脂质和糖类等。
相互作用
03
是指两个或多个分子之间发生的相互影响和作用,这种作用力
可以是物理的、化学的或者是生物学上的。
蛋白质与生物大分子相互作用的重要性
详细描述
蒙特卡罗模拟通过随机抽样和统计分析,可以模拟蛋白质与生物大分子的结合和 相互作用过程。该方法可以预测蛋白质与生物大分子的结合常数、亲和力以及结 合过程中的能量变化,提供有关结合机制和动力学特性的信息。
粗粒化模型
总结词
粗粒化模型是一种简化模型,将分子体系中的原子或分子的复 杂行为简化为较少的“粗粒”或“组分”,以便于模拟计算。
总结词
研究蛋白质与生物大分子相互作用的调控机 制有助于发现新的药物靶点和治疗策略。
详细描述
蛋白质与生物大分子的相互作用在许多生物 学过程中发挥着关键作用,包括基因表达、 物质代谢等。研究这些相互作用的调控机制 可以帮助我们更好地理解这些过程,并发现 新的药物靶点和治疗策略,为疾病治疗提供 新的思路。
蛋白质与生物大分子相互作用的类型
共价结合
通过化学键将蛋白质与生物大分子永久性地结合在一起。例如,蛋白质与核酸 的磷酸二酯键就是共价结合。
非共价结合
通过非共价键将蛋白质与生物大分子暂时性地结合在一起,这种结合力可以在 一定条件下被打破。例如,蛋白质与核酸的氢键、范德华力和疏水相互作用等。
02
蛋白质与生物大分子相 互作用的实验研究方法
蛋白质折叠与稳定性研究
总结词
研究蛋白质的折叠方式和稳定性对于理解其功能和疾病发生机制具有重要意义。
蛋白质纯化技术研究进展

蛋白质纯化技术研究进展蛋白质是生物体中重要的分子之一,它们扮演着维持生命的许多功能。
蛋白质纯化是研究蛋白质结构和功能的重要手段,也是蛋白质制药的必要步骤。
随着蛋白质研究的深入,蛋白质纯化技术得到了迅速发展。
一、传统分离技术传统的蛋白质分离技术主要是色谱技术、离心分离、电泳和沉淀分离。
这些方法需要依赖蛋白质的理化性质进行分离,如分子量、电荷、亲水性等。
虽然这些方法在某些情况下能够达到较好的效果,但它们都存在某些缺陷:分离效率低、耗时长、操作复杂、易失活。
二、亲和层析技术为了弥补传统方法的不足,研究人员不断寻求更加高效的蛋白质纯化方法。
亲和层析技术因其选择性强、分离效率高而成为了越来越受欢迎的方法。
亲和层析是一种介于化学亲和力与色谱法之间的分离方法。
亲和层析技术基于结构域或分子表面的分子配对进行径流分离,通过其他化学分离步骤无法分离。
可以通过单独的亲和层析胶体,也可以通过一系列不同的分离中介来实现。
亲和层析技术的选择取决于要纯化的蛋白质,可以选择适当的亲和序列或化学分离介体来保留目标蛋白质。
例如,大分子蛋白质可以在硫酸盐,硝酸盐或氢氧化钠pH4处理的大雀麦亲和层析树脂上纯化。
其他分子可以使用具有含灵长目蛋白质核酸相互作用的亲和树脂或利用酵母细胞结构中单链碱性氨基酸残基与标记的蛋白质交互作用的阳离子树脂来纯化。
亲和层析技术是纯化问题的强力工具,并且在高效液相色谱基础上易于实现。
有基于亲和铁柱螯合技术和基于镍柱螯合技术等多种亲和层析技术。
除了常见的亲和柱,还可以使用核壳颗粒亲和层析和温和亲和层析方法等。
尽管这些技术在结构域或分子表面上具有出色的配对性,从而实现了高选择性和高效率的径流分离,但是它们仍受到性能限制,例如TTP血友病因子的亲和结合为单精联系数,目前仍无法推广生产。
此外,许多技术需要较长的开发时间和高成本的特异性重新定制,这也是大规模生产的主要障碍。
三、高通量纯化技术高通量纯化技术就是对传统方法和亲和层析技术的一个进一步创新和提高。
蛋白质与核酸的相互作用核酸结合蛋白模板
3.5 通过β链识别的蛋白
DNA识别 区域是由 1-2个β链 折叠结构 组成,它 们可以进 入DNA大 沟。
四、蛋白质中RNA结合motif
RNP(核糖核蛋白)结构域:
有两个保守motif-RNP1和RNP2,均为一
个大结构的一部分,β-α-β-β-α-β,其中motif
位于中心的一个β折叠处。通常与SnRNA结合,
3.2 半胱氨酸-组氨酸锌指
3.2.1 Cys2-His2锌指组件
锌指结构(Zinc finger) 是第一个被发现的真核细 胞中与DNA结合的蛋白质, 在真核基因组中广泛存在, 其约占基因组的0.5%, 在原核生物中虽有发现, 但相对较少。目前已发现 有六种类型,其经典结构 如右图。
3.2.2 常见的锌指结构三种类型
7.1 同蛋白质结合的核酸序列研究 7.1.1 凝胶阻滞分析:
原理:蛋白质可以与末端标记的核酸探针结合,电 泳时这种DNA/RNA蛋白质复合物比裸露的DNA或 RNA电泳迁移慢, 即表现为相对滞后。 用途: 鉴定和了解与蛋白质结合的DNA或RNA位点;
说明: 将蛋白质与标记的DNA或RNA保温或不保温, 电泳后比较迁移率。再用过量的特殊序列的寡聚核 苷酸替代试验, 以确定特殊序列与蛋白质结合位点。
研究表明:碱性结合结构域中,与DNA结合识
别的α螺旋可能是与DNA结合的结果。
如酵母bZIP蛋白的碱性结构域, 在溶液中为一
种无序的部分螺旋的结构; 当其同DNA结合时, 这种构象发生了变化,诱导了典型的α螺旋形 成。
3.4 类组蛋白结构motif
真核细胞的DNA分子,通过与组蛋白的作用, 将组蛋白包裹成核小体。每个核小体中含有两 个负超螺旋的DNA结构,与组蛋白相结合。
液相分离和蛋白互作

液相分离和蛋白互作1. 引言液相分离是一种广泛应用于化学、生物化学和生物技术等领域的重要技术。
而蛋白质作为生物体内最重要的功能分子之一,在细胞内发挥着关键的生物学功能。
液相分离技术与蛋白质互作研究有着密切的联系,可以帮助我们更好地了解蛋白质的性质和功能。
本文将重点介绍液相分离技术及其在蛋白质互作研究中的应用。
2. 液相分离技术的概述液相分离技术是指利用溶液中不同成分之间的分配系数差异,通过各种手段将它们分离开来的方法。
常见的液相分离技术包括凝胶过滤、凝胶电泳、层析技术等。
这些技术基于不同的原理和机制,可以实现对复杂混合物中目标分子的有效分离和纯化。
3. 蛋白质互作的重要性蛋白质互作是指蛋白质与其他分子(如蛋白质、核酸、小分子化合物等)之间的相互作用。
蛋白质互作在细胞内调控信号转导、细胞周期以及基因表达等重要生物学过程中起着关键作用。
通过研究蛋白质互作网络,可以揭示蛋白质的功能和调控机制,进而对相关疾病的发生和治疗提供理论依据。
4. 液相分离在蛋白质互作研究中的应用4.1 凝胶过滤技术凝胶过滤技术是利用凝胶过滤膜的孔隙大小选择性地分离不同分子量的蛋白质。
通过将混合物溶液滴在凝胶膜上,较大分子将被滞留在膜上,而较小分子则能通过膜孔进入收集器中。
这种技术广泛应用于蛋白质纯化和分析中,可用于分离互作蛋白质复合物中的组分。
4.2 凝胶电泳技术凝胶电泳技术是一种利用凝胶材料的孔隙结构和电泳作用力将蛋白质分离开来的方法。
根据凝胶的不同,可以将凝胶电泳分为聚丙烯酰胺凝胶电泳(SDS-PAGE)和聚丙烯酰胺凝胶电泳(PAGE)。
这些技术常被用于分离蛋白质复合物中的不同组分,并进一步检测和分析其相互作用。
4.3 层析技术层析技术是一种基于溶液中分子与固定相之间的相互作用,通过在固定相上进行吸附、解吸等过程实现分离的方法。
常见的层析技术包括大小排阻层析、亲和层析、离子交换层析等。
这些技术可用于从复杂混合物中高效地分离纯化特定蛋白质,进而研究其与其他分子的相互作用。
液相分离和蛋白互作

液相分离和蛋白互作引言:液相分离技术是一种广泛应用于生物化学和生物技术领域的方法,用于分离和纯化复杂混合物中的生物分子。
蛋白质是生物体内最重要的分子之一,其功能与其所处的细胞环境密切相关。
蛋白质在细胞内通过与其他分子的相互作用来实现其功能。
因此,研究蛋白质的互作关系对于理解细胞生物学和疾病机制具有重要意义。
一、液相分离技术1. 色谱技术色谱技术是一种基于物质在固定相和移动相之间的分配平衡特性进行分离的方法。
常见的色谱技术包括气相色谱、液相色谱和离子色谱等。
在液相色谱中,样品溶液经过一列填充有固定相的柱子,通过样品中各种成分在固定相和移动相之间的相互作用力差异来实现分离。
2. 凝胶电泳技术凝胶电泳是一种利用电场对带电生物分子进行分离的方法。
常见的凝胶电泳技术包括聚丙烯酰胺凝胶电泳、琼脂糖凝胶电泳和聚丙烯酰胺梯度凝胶电泳等。
在凝胶电泳中,样品溶液经过电场作用下在凝胶基质中移动,根据其电荷、大小和形状的差异来实现分离。
3. 膜分离技术膜分离技术是一种利用半透膜对混合物进行分离的方法。
常见的膜分离技术包括超滤、微滤和逆渗透等。
在膜分离中,混合物通过半透膜时,根据溶质的大小、形状和电荷等特性,使得不同组分通过膜的速率不同,从而实现分离。
二、蛋白质互作1. 蛋白质-蛋白质相互作用蛋白质-蛋白质相互作用是指两个或多个蛋白质之间的相互作用关系。
这些相互作用可以是非共价的,如蛋白质的结合、识别和调控等;也可以是共价的,如蛋白质的交联和修饰等。
蛋白质-蛋白质相互作用对于细胞信号传导、代谢调控和细胞周期等过程至关重要。
2. 蛋白质-核酸相互作用蛋白质-核酸相互作用是指蛋白质与核酸之间的相互作用关系。
这些相互作用可以是蛋白质与DNA/RNA的结合、识别和调控等。
蛋白质-核酸相互作用在基因表达、转录调控和DNA修复等生物学过程中发挥着重要作用。
3. 蛋白质-小分子相互作用蛋白质-小分子相互作用是指蛋白质与小分子化合物之间的相互作用关系。
三联体结合实验原理

三联体结合实验原理三联体结合实验原理引言:三联体结合实验是一种常用的生物学实验技术,用于研究蛋白质与DNA或RNA之间的相互作用。
该实验原理基于三联体的形成,即蛋白质与核酸分子之间的特异性结合。
本文将详细介绍三联体结合实验的原理。
一、蛋白质与DNA/RNA相互作用蛋白质是生物体内重要的功能分子,它们能够与DNA或RNA相互作用并发挥调控基因表达等生物学功能。
这种相互作用通常通过特定区域上的氨基酸残基与核酸分子上的碱基进行结合来实现。
二、三联体结合实验原理三联体结合实验是一种常用的方法,用于研究蛋白质与DNA/RNA之间的相互作用。
该实验通过以下步骤进行:1. 准备试样需要准备纯化得到的目标蛋白质和DNA/RNA样品。
目标蛋白质可以通过多种方法纯化得到,如亲和层析、离心沉淀等。
而DNA/RNA样品则可以通过PCR扩增、合成等方法得到。
2. 三联体形成将目标蛋白质与DNA/RNA样品混合,并在一定的条件下进行反应,使其形成三联体。
这些条件通常包括适当的缓冲液、温度和离子浓度等。
在反应过程中,蛋白质的特异性结合区域与DNA/RNA上的特定序列进行结合,形成稳定的三联体。
3. 分离和检测将形成的三联体分离出来,并进行检测。
常用的分离方法包括凝胶电泳、超速离心等。
检测方法可以根据实验需要选择,如荧光染色、放射性标记等。
通过分离和检测,可以确定蛋白质与DNA/RNA之间是否发生了特异性结合。
4. 数据分析根据实验结果,可以对蛋白质与DNA/RNA之间的相互作用进行进一步分析。
可以确定特异性结合区域、结合强度等信息。
同时,还可以利用其他技术手段进一步验证实验结果,如核磁共振、X射线晶体学等。
三、应用领域三联体结合实验在生物学研究中具有广泛的应用。
它可以用于研究蛋白质与DNA/RNA之间的相互作用机制,揭示蛋白质的功能和调控机理。
三联体结合实验还可以用于筛选潜在的药物靶点,发现新型药物,并评估药物与靶点之间的相互作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第23卷第10期2011年10月化 学 进 展PROGRESS IN CHEMISTRYVol.23No.10 Oct.2011 收稿:2011年1月,收修改稿:2011年3月 †共同第一作者 ∗国家重点基础研究发展计划(973)项目(No.2007CB914101)和国家自然科学基金项目(No.20875009)资助∗∗Corresponding author e⁃mail:qufengqu@离子液与蛋白质和核酸相互作用的研究∗张 涛†1 陈 凡†1 盖青青1 屈 锋1∗∗ 张玉奎2(1.北京理工大学生命学院 北京100081;2.中国科学院大连化学物理研究所 大连116023)摘 要 离子液因其具有良好的生物兼容性和独特的理化性质,近年来在生物催化和生物大分子蛋白质与核酸的分离分析领域得到广泛应用。
离子液与生物大分子相互作用的研究是离子液相关理论与应用研究的基础,有关离子液与蛋白质和核酸相互作用的机理研究受到关注。
本文简要介绍了常用离子液的分类,离子液与蛋白质分子作用的机理,离子液与核酸分子作用的机理,以及离子液在酶催化反应、生物分子分离、生物分子电化学分析和毛细管电泳分析中的应用,并主要综述了近年的相关研究和应用进展。
关键词 离子液 蛋白质 核酸 相互作用中图分类号:O657.1;O629.7 文献标识码:A 文章编号:1005⁃281X(2011)10⁃2132⁃08Ionic Liquids and Protein /Nucleic Acid InteractionZhang Tao †1 Chen Fan †1 Gai Qingqing 1 Qu Feng 1∗∗ Zhang Yukui 2(1.School of Life Science,Beijing Institute of Technology,Beijing 100081,China;2.Dalian Institute of Chemical Physics,Chinese Academy of Sciences,Dalian 116023,China)Abstract Room⁃temperature ionic liquids(ILs)have aroused considerable interest recent years.They havebeen widely applied in biological catalysis and proteins and nucleic acids separation due to their good compatibilityand unique properties.The interaction of ionic liquids and biological macromolecules is the foundation of theirtheory and application study.In this paper,the types of common used ILs,the principle of interaction between ionic liquids and protein,and ionic liquids and nucleic acid are introduced respectively.Furthermore,the recent application of ILs in biological catalysis,biological molecules separation electrochemical analysis as well ascapillary electrophoresis analysis is reviewed.Key words ionic liquids;protein;nucleic acids;interactionContents1 Introduction 2 Type of ILs3 Interaction between ILs and protein 3.1 Electrostatic interaction 3.2 Hydrophobic interaction 3.3 Hydrogen bond interaction3.4 Coordination interaction 4 Interaction between ILs and nucleic acid4.1 Electrostatic interaction4.2 Hydrophobic interaction4.3 Hydrogen bond interaction 5 Application of ILs based on the interaction betweenprotein and nucleic acid 5.1 Enzyme⁃catalyzed reaction第10期张 涛等 离子液与蛋白质和核酸相互作用的研究·2133 ·5.2 Extraction separation5.3 Electrochemical analysis5.4 Separation and analysis in CE6 Outlook1 前言20世纪80—90年代以来,离子液(ionic liquids,ILs)的研究日益广泛。
离子液(又称室温离子液或室温熔融盐)是指在室温或接近室温下呈现液态的融盐,通常由特定的有机阳离子和无机阴离子构成[1,2]。
它不仅具有传统有机溶剂的优势,而且相比有机溶剂表现出很多独特的理化性质[3,4],如室温下稳定性好、蒸气压低、不可燃、溶解能力强和导电性好等。
更重要的是,可通过改变阳离子和阴离子种类而改变离子液的理化性质。
因此,离子液成为一种可人为设计的“绿色溶剂”[5,6]。
离子液作为有机溶剂的替代品一直应用于有机合成、电化学等领域。
自从Erbeidinger[7]首先报道离子液作为酶促反应介质可提高酶的稳定性和活性以来,研究发现许多酶都能在ILs及其形成的两相体系或单相体系中保持催化活性,因此ILs作为生物催化反应的溶剂或共溶剂的研究备受关注,ILs以其独特的优势成为生物催化反应研究的热点[8—10]。
此外,离子液在生物分子分离分析方面也取得新的应用进展,如其在酶催化、蛋白质和核酸的萃取分离[11—14]、及在色谱、电泳和电化学分析等生物分子分析检测中的应用[15—24]。
离子液在生物催化和分离分析领域的应用,涉及离子液与生物大分子相互作用的机理研究。
由于离子液的结构多样性和可设计性,它可承载多种功能基团,而生物大分子种类繁多、结构复杂,因此二者之间的相互作用机理比较复杂。
目前离子液与蛋白质、核酸等生物大分子相互作用机理的研究还相对较少,这可能与离子液几乎没有活性,很难直接产生光学、电学以及热力学信号,因此很难直接检测到其与生物大分子相互作用的物理化学信息有关[25,26]。
本文将对离子液与蛋白质和核酸的相互作用的机理研究,以及近年来离子液在生物大分子酶催化和分离分析中的应用进行综述。
2 离子液的分类离子液种类繁多,通过改变阴离子和阳离子的组合可产生成千上万种离子液,因而可设计和满足各种不同需求的离子液。
中国科学院张锁江课题组在2005年建立了离子液体数据库,当年已收集包含了276种阳离子、55种阴离子和588种离子液的物理化学性质,并且不断的更新和丰富[27]。
目前,用于蛋白质和核酸相关领域的离子液种类相对较少,根据阳离子组成的不同,主要分为4类[28,29]:(1)咪唑类离子液;(2)吡啶类离子液;(3)烷基季铵盐类离子液;(4)烷基季 盐类离子液。
其中,以烷基取代的咪唑基离子液和晕⁃烷基取代的吡啶类离子液的研究和应用较常见。
上述组成的离子液中,常见无机阴离子主要有Cl-、AlCl-4、BF-4、PF-6、NO-3、CF3SO-3、NTf-2、HSO-4和CF3CO-2等。
3 离子液与蛋白质的相互作用离子液与蛋白质相互作用时可以改变生物大分子理化性质,并通过离子液的功能基团直接或间接作用于生物大分子。
离子液种类繁多,其阳离子、阴离子和功能基团,都可与蛋白质产生作用,改变蛋白质的结晶行为[30—33]、变性温度[34]和溶解性能[35],以及改变酶的活性和稳定性等[36,37]。
例如离子液作为蛋白质结晶的添加剂,可改变蛋白质的晶体结构,提高结晶的均一性,促进蛋白质的结晶行为[30,32]。
某些特殊的离子液,如溴化1⁃丁基⁃3⁃甲基咪唑[33]或1⁃丁基⁃3⁃甲基咪唑四氟硼酸盐[31]能够控制蛋白质结晶的生长速度,形成均一稳定的结晶形态。
离子液作为酶催化的介质或者酶催化反应的助溶剂或添加剂,具有重大的应用前景。
某些离子液可以增强蛋白质的溶解性能,选择性地提高酶的催化活性,提高酶催化的效率[35—37]。
相反,某些离子液却使蛋白质的变性温度降低,破坏蛋白质的热稳定性,而且疏水作用越强,这种破坏作用越强[34]。
此外,离子液还可特异性地吸附或萃取蛋白质[38],甚至可通过形成离子液双水相体系对蛋白质进行选择性分配[39—41]。
蛋白质在某些离子液中通常不溶解[8],说明蛋白质与离子液之间没有明显的化学键合。
可溶解于离子液中的蛋白质,其在离子液中的紫外光谱最大吸收峰和红外光谱红外特征吸收峰没有发生明显位移和强度的变化,也说明离子液与蛋白质之间没有发生化学键合[39,42]。
然而离子液中的阴、阳离子通常以静电作用、疏水作用、氢键作用、范德华力、盐析效应和空间位阻作用等与蛋白质的一些特定功能基团相互作用。
因此离子液与蛋白质作用通常是由多种作用力共同导致,较难明确其主要作用力。
3.1 离子液阳离子与蛋白质的静电作用·2134 ·化 学 进 展第23卷离子液与蛋白质作用时,主要是离子液中阳离子起主要作用。
蛋白质作为两性分子其带电性质受介质pH影响,调节介质pH值可使蛋白质带负电荷,便于与离子液的阳离子部分发生静电作用。
溴化1⁃丁基⁃3⁃甲基咪唑/磷酸氢二甲离子液双水相体系([Bmim]Br/K2HPO4)中,蛋白质的分配随着体系pH的变化而改变,说明蛋白质的分配主要由静电作用决定,并可利用静电作用选择性地分离蛋白质[39]。
当蛋白质带正电时其在离子液相中的含量相对较低,而随着pH值减小,蛋白质带正电荷增加,与离子液正离子产生排斥作用增强,使萃取率降低;当溶液的pH值大于蛋白质的等电点时,因蛋白质带负电荷,其在离子液相中的含量增加。
Dreyer等[11]考察了4种模板蛋白在不同离子液/盐双水相体系中的分配主要是受静电作用和蛋白质分子量的影响。