第三节噬菌体的遗传分析
噬菌体的遗传分析

表5-3
琥珀型抑 制基因
su1+ su2+ su3+ su4+ su5+
5种琥珀抑制基因的性质
插入的 氨基酸
合成的蛋白质 占野生型%
丝氨酸
28
谷氨酰胺
14
酪氨酸
55
酪氨酸
16
赖氨酸
5
赭石型抑 制基因
+ +
第二节 噬菌体突变型的互补试验
一X174条件致死突变型的互补测验 (一) X174 DNA结构复制 图5-1 (二) X174的突变型与互补测验
B am14,am16,och5,ts9,tsl16,och1,och8,och11,
C och6
D am10,amH81,
E am3,am6,am27,
F am87,am88,am89, amH57,op6, op9,tsh6,ts41D
G am9,am32,ts,ts79
H amN1,am23,am80,am90,ts4
半透明,大 h+r- 亲组合
透明,大 h-r- 重组合
Ecoli B
半透明,小 h+r+ 重组合
(二) 噬菌体重组值的计算
重组噬菌斑数
重组值 =
X100%
总噬菌斑数
三 T4突变型的三点试验 (m:小;r:快速;tu:混浊)
表5-5 T4的 m r tu x + + + 三点试验结果
亲本类型 单交换型 单交换型 双交换型
合计
类 型 噬菌斑数 %
重组频率%
m-r r-tu m-tu
m r tu +++
《噬菌体遗传变异》课件

复杂的调控系统
噬菌体通过操纵宿主细菌的基因表达来确保自 身复制和装配。
寄生策略
噬菌体利用细菌的代谢和复制机制来增殖和传 播。
噬菌体的变异现象
1
细胞感染噬菌体的基本过程
噬菌体通2
噬菌体的遗传变异分类
突变、基因重排和顺式转移是噬菌体发生遗传变异的重要方式。
2. (2) Rostocha E, Briers Y, Noben JP, Lavigne R. Tracking the Genetic Stability of Bacteriophage Therapy Products. In Phage Therapy: A Practical Approach. Humana Press, New York, NY. 2019. pp. 41-53.
噬菌体变异的生物学意义
1 增加生存能力
噬菌体的变异使其能适应不同环境和宿主,提高生存率和传播能力。
2 收集有益的基因
噬菌体通过变异可以获取和传递有益的基因,如耐药基因。
噬菌体变异的应用前景
抗生素研究
噬菌体变异为开发新型抗生素提供了来源和启示。
新型抗菌药物研发
噬菌体变异可以为寻找新的抗菌机制和药物靶点提 供线索。
噬菌体遗传变异
介绍噬菌体以及其遗传特征,探究噬菌体遗传变异的现象、生物学意义和应 用前景,展示其在医学领域中的重要性。
什么是噬菌体?
噬菌体是一种以细菌为宿主的寄生病毒,通过感染细菌来繁殖自身,并对细 菌进行遗传调控。
噬菌体的遗传特征
寄生病毒
噬菌体以细菌为宿主,并在细菌体内复制自身。
基因组结构
噬菌体基因组包含核酸序列,编码复制酶和结 构蛋白。
结论
重要的基础研究领域
细菌和病毒的遗传—噬菌体的遗传分析(遗传学课件)

寄主范围突变体:其吸附器能吸附在抗噬 菌体细菌的细胞壁上,侵染并使其裂解。
二、T2噬菌体的基因重组
r-型突变:噬菌斑比正 常大2倍,边缘清晰。 r+型:噬菌斑小,边缘 模糊。
二、T2噬菌体的基因重组
h+型:噬菌体只能利用E.coli B 菌株。 h-型突变:噬菌体能利用E.coli B和E.coli B/2菌株(对T2的抗 性菌株。
烈性噬菌体在侵染 宿主细胞后,能进行营 养增殖,并在短期内使 寄主细胞裂解。如T噬 菌体系列。
温和性噬菌体在侵染宿主细胞后,可以将 其基因组插入寄主染色体,随寄主染色体同步 复制,且具有潜在的合成噬菌体粒子的能力。 如P噬菌体和λ噬菌体。
二、T4噬菌体的生活周期
T4噬菌体是烈性噬 菌体,DNA通过尾部 结构注入细胞,在宿主 内转录启动,合成噬菌 体DNA、外壳蛋白, 组装成噬菌体释放。
在物理或化学条件 诱导下,溶源细菌可进 入裂解途径,最终产生 大量噬菌体。
用hr+和h+r同时感染 E.coli B株,称为双重感 染。将释放出来的子代噬 菌体接种在混合生长有B 和B/2菌株上,结果在培 养基上出现了四种噬菌体 菌斑。
hr+×h+r混合感染产生的噬菌斑类型
根据噬菌体的表型特征推导它们的基因型
噬菌体表现型
透明
小
半透明
大
半透明
小
透明
大
推导的基因型
hr+ 亲本型
《遗传学》
一、噬菌体的性状
(一)噬菌斑的形态
不同基因型的噬菌 体形成不同形态的噬菌 斑,有的大,有的小, 有的边缘清楚,有的边 缘模糊等。
7、细菌和噬菌体的遗传分析2

第三节 噬菌体的遗传分析 三、烈性噬菌体与基因定位
双重感染(混合感染、复感染):是指用两种噬菌体同时感染某一菌株。 双重感染(混合感染、复感染):是指用两种噬菌体同时感染某一菌株。 ):是指用两种噬菌体同时感染某一菌株
例如: 例如: 噬菌体Ⅰ ):能感染 能感染B B/2菌株 噬菌斑透明;产生噬菌斑小且边缘模糊; 菌株, 噬菌体Ⅰ(hr+):能感染B和B/2菌株,噬菌斑透明;产生噬菌斑小且边缘模糊; 噬菌体Ⅱ(h+r):只能感染B菌株产生噬菌斑;噬菌斑生长较快(约两倍大)且 ):只能感染 菌株产生噬菌斑;噬菌斑生长较快(约两倍大) 只能感染B 噬菌体Ⅱ 边缘清晰; 边缘清晰; 两种噬菌体同时感染B菌株(双重感染) 用 hr+ 和 h+r 两种噬菌体同时感染B菌株(双重感染)。 在双重感染( 的过程中, 在双重感染(相当于 hr+ ×h+r)的过程中,共同生存在同一个宿主细胞中的 两个噬菌体DNA也可以发生交换,产生基因重组。 hr、 两个噬菌体DNA也可以发生交换,产生基因重组。在其子代中可以得到 hr、h+r+ 两 DNA也可以发生交换 种重组体以及 两种亲本类型, 种噬菌体。 种重组体以及 hr+、h+r 两种亲本类型,共4种噬菌体。
再做杂交: 再做杂交:rc rb+ × rc+ rb
结果表明: 结果表明:
rc—rb的重组值 ﹥ rb—h
∴ h位于rb及rc之间,排列顺序 rc—h—rb。 位于r 之间, 由于T2 噬菌体的连锁图是环状的,所以2 排列都对。 由于T2 噬菌体的连锁图是环状的,所以2、3排列都对。
第三节 噬菌体的遗传分析 四、温和噬菌体 与溶源性周期和溶菌周期
《噬菌体遗传与变异》课件

新合成的噬菌体粒子在宿主细胞 内成熟,并释放出宿主细胞外。
噬菌体对宿主的致病作用
破坏宿主细胞结构
噬菌体感染导致宿主细胞 破裂,释放出有毒物质, 引发感染症状。
引发免疫反应
噬菌体感染可引发宿主免 疫反应,导致炎症、发热 等症状。
传播与流行
噬菌体可通过感染一个宿 主,再传播给其他宿主, 引发流行性疾病。
调控机制
噬菌体DNA的复制与转录受到严格的调控机制,以确保遗传信息 的正确传递。
噬菌体DNA的突变与重组
突变类型
噬菌体DNA的突变类型包括点突变、插入和缺 失等。
重组机制
噬菌体DNA的重组机制涉及到同源重组和非同 源重组两种方式。
Hale Waihona Puke 生物学意义突变与重组在噬菌体的进化、变异和适应性方面具有重要意义。
03
利用噬菌体的特异性感染和裂解细菌的能力,治疗细菌感染性疾 病。
噬菌体疫苗
利用噬菌体作为载体,表达和展示病原体抗原,用于预防传染病 。
噬菌体基因检测
利用噬菌体感染和扩增特定病原体的基因,用于快速检测和诊断 疾病。
噬菌体在环境治理中的应用
污水处理
利用噬菌体的抑菌和裂解细菌的能力,处理污水中的有害细菌, 提高水质。
噬菌体变异的机制
1 2
DNA复制错误
在DNA复制过程中,由于碱基配对错误或DNA 聚合酶的缺陷,导致基因突变的发生。
重组酶介导的基因重组
在同源或异源DNA之间,重组酶能够识别并催化 DNA的交换和重排,导致基因重组。
3
基因水平转移
通过噬菌体感染或细菌之间直接接触,可实现基 因在不同细菌或噬菌体之间的转移和重组。
《噬菌体遗传与变异》PPT课件
噬菌体遗传分析
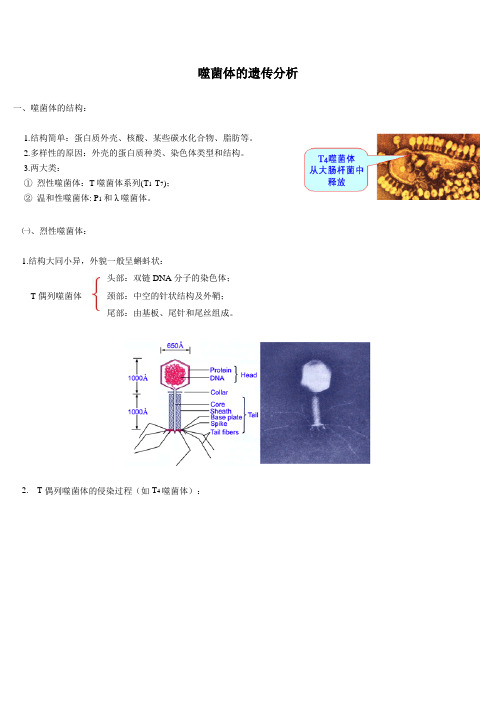
噬菌体的遗传分析一、噬菌体的结构:1.结构简单:蛋白质外壳、核酸、某些碳水化合物、脂肪等。
2.多样性的原因:外壳的蛋白质种类、染色体类型和结构。
3.两大类:①烈性噬菌体:T噬菌体系列(T1-T7);②温和性噬菌体: P1和λ噬菌体。
㈠、烈性噬菌体:1.结构大同小异,外貌一般呈蝌蚪状:T偶列噬菌体头部:双链DNA分子的染色体;颈部:中空的针状结构及外鞘;尾部:由基板、尾针和尾丝组成。
2.T偶列噬菌体的侵染过程(如T4噬菌体):尾丝固定于大肠杆菌,遗传物质注入破坏寄主细胞原有的遗传物质合成大量的噬菌体遗传物质和蛋白质组装许多新的子噬菌体溶菌酶裂解细菌释放出大量噬菌体。
右图为T4噬菌体侵染大肠杆菌的生活周期㈡、温和性噬菌体:例如λ和P1噬菌体,λ和P1各代表一种略有不同的溶源性类型。
1.溶源性噬菌体的生活周期:①.λ噬菌体:噬菌体侵入后,细菌不裂解附在E.coli染色体上的gal和bio位点间的attλ座位上通过交换整合到细菌染色体,并能阻止其它λ噬菌体的超数感染。
λ噬菌体特定位点的整合②P1噬菌体:不整合到细菌的染色体上,而是独立存在于细胞质内(见左下图)。
原噬菌体:整合到宿主基因组中的噬菌体。
仅少数基因活动,表达出阻碍物关闭其它基因。
原噬菌体经诱导可转变为烈性噬菌体裂解途径(见右下图)。
2.P1和λ噬菌体的特性:①P1和λ各代表不同的溶源性类型:P1噬菌体:侵入后并不整合到细菌的染色体上,独立存在于细胞质内;λ噬菌体:通过交换整合到细菌染色体上。
②溶源性细菌分裂两个子细胞:P1噬菌体复制则使每个子细胞中至少含有一个拷贝;λ噬菌体随细胞染色体复制而复制,细胞中有一个拷贝。
③共同特点:核酸既不大量复制,也不大量转录和翻译。
P1和λ噬菌体的生活周期特性二、T2噬菌体的基因重组与作图:1.噬菌体遗传性状分为两类:形成的噬菌斑形状:指噬菌斑大小、边缘清晰度、透明程度。
寄主范围:指噬菌体感染和裂解的菌株范围。
第三节噬菌体的遗传分析

2024/6/17
8
1 λ噬菌体
• 原噬菌体通过诱导(induction)可转变为 烈性噬菌体,进入裂解周期。
• 诱导可以通过不同的方式进行,如UV照射、 温度改变、与非溶原性细菌的接合等。
• 诱导使阻遏物失活,使噬菌体的其他基因 得以表达,促使噬菌体繁殖并进入裂解周 期。
2024/6/17
9
2 P1 噬菌体
10
二、噬菌体的基因重组
• 两个基因型不同的噬菌体同时感染一个宿 主细胞,叫做混合感染(mixed infection) 或双重感染(double infection)。
• 共同生存在同一个宿主细胞中的两个噬菌 体的DNA也可以发生交换,产生基因重组。
2024/6/17
11
二、噬菌体的基因重组
• 比如,一个噬菌体的基因型是a+b-,另一个 噬菌体的基因型是a-b+,同时感染同一个宿 主细胞,宿主细胞裂解以后,可能释放出基 因型为a+b+和a-b-的重组体来。 研究最深入的噬菌体突变体是T2 噬菌体的r(rapid lysis速溶性)突变体。一个正常的 T2噬菌体产生的噬菌斑小而边缘模糊,记为r +,突变体r-产生的噬菌斑大而边缘清晰。
6.4
0.9
• 根据表7-2的结果可以分别作出3个连锁图。
P155
有四种可能的排列顺序。P155
2024/6/17
16
• 四种顺序都是可能的,要确定到底是那一 种,还缺条件。若知道rb和 rc之间的距离, 就 为可此以,推需知作rrbb、 +rrc c×和hr的b 排rc列+ 。顺结序果。rb与rc之 间的距离大于rb与h之间的距离,可知h应位 于rb与rc之间,即rbhrc。 至于ra位于h的哪一边,是靠近rc还是靠近 r是b?正因确为的T。2 DNA是环状的,所以两种答案都
课件噬菌体遗传与变异.ppt

形态与结构
– 蝌蚪形、微球形和丝形
化学组成
– 蛋白质与核酸
抗原性 抵抗力:比细菌强
噬菌体
噬菌体的种类
毒性噬菌体(virulent phage)
温和噬菌体(temperate phage)/ 溶原性噬菌体(lysogenic phage)
第二节 毒性噬菌体
毒性噬菌体(virulent phage) –能在宿主菌细胞内复制增殖,产生许多
–致育质粒(F质粒):编码细菌性菌毛 –耐药质粒(R质粒):编码细菌耐药性
毒力质粒(Vi质粒):编码细菌毒力因子 –细菌素质粒(Col质粒):编码大肠埃希菌的大
肠菌素 –代谢质粒
质粒是基因工程中最常用的基因载体
转座因子 (transposable element)
是指基因组中能够改变自身位置的一段DNA片段。 转位(transposition):转位因子能从染色体 或质粒的一个位置转移到在另一个位置。 转位因子的转位行为,能使DNA发生插入突变和 广泛的基因重排。 包括:插入序列、转座子、转座噬菌体
抑制基因转录 ✓ 阻遏蛋白 ✓ 辅阻遏物
促进基因转录
细菌的变异机制
基因的转移和重组 基因的突变
第三节 基因的转移和重组
基因转移(gene transfer)
–外源性的遗传物质由供体菌转入某受体菌细胞的过 程称为基因转移。
重组(rebination)
–转移的基因与受体菌DNA整合在一起称为重组,使 受体菌获得供体菌的某些性状。
IS Resistance Gene(s) IS
Tn
转座子的特征
转座子 Tn1 Tn2 Tn3
Tn4 Tn5 Tn6 Tn7 Tn9 Tn10 Tn551 Tn971 Tn1681
5答案细菌和噬菌体的遗传分析

细菌和噬菌体的遗传分析习题一一、填空题1、Hfr,F因子2、整合或游离于细菌染色体上或之外附加体3.末端4.裂解重组体合子诱导5、一次单交换6、Hfr,F因子,细菌7、溶菌,r+斑、r斑8、高频重组,广泛性转导9、F+ F+二、解释下列名词:F-菌株:未携带F因子的大肠杆菌菌株。
F+菌株:包含一个游离状态F因子的大肠杆菌菌株。
Hfr菌株:包含一个整合到大肠杆菌染色体组内的F因子的菌株。
F因子:大肠杆菌中的一种附加体,控制大肠杆菌接合过程而使其成为供体菌的一种致育因子。
F'因子:整合在宿主细菌染色体上的F因子,在环出时不够准确而携带有染色体一些基因的一种致育因子。
烈性噬菌体:侵染宿主细胞后,进入裂解途径,破坏宿主细胞原有遗传物质,合成大量的自身遗传物质和蛋白质并组装成子噬菌体,最后使宿主裂解的一类噬菌体。
温和性噬菌体:侵染宿主细胞后,并不裂解宿主细胞,而是走溶原性生活周期的一类噬菌体。
溶原性细菌:含有温和噬菌体的遗传物质而又找不到噬菌体形态上可见的噬菌体粒子的宿主细菌。
部分二倍体:当F+和Hfr的细菌染色体进入F-后,在一个短时期内,F-细胞中对某些位点来说总有一段二倍体的DNA状态的细菌。
三、选择题1-5、d b d b c6-10、A C A B A四、问答题2.为什么说细菌和病毒是研究遗传学的好材料?答:与其他生物体相比,细菌和病毒能成为研究遗传学的好材料,具有以下7个方面的优越性:(1)世代周期短:每个世代以min或h计算,繁殖速度快,大大缩短了实验周期。
(2)易于管理和进行化学分析个体小,繁殖方便,可以大量节省人力、物力和财力;且代谢旺盛,繁殖又快,累积大量的代谢产物。
(3)便于研究基因的突变细菌和病毒均属于单倍体,所有突变都能立即表现出来,不存在显性掩盖隐性的问题。
(4)便于研究基因的作用通过基本培养基和选择培养基的影印培养,很容易筛选出营养缺陷型,利于生化[研究。
(5)便于基因重组的研究通过细菌的转化、转导和接合作用,在一支试管中可以产生遗传性状不相同的后代。
第六章细菌和噬菌体的遗传分析

18
五、细菌的遗传作图
(一)细菌的遗传重组 原核生物的遗传重组实质上是指受体中插入来自供体的遗传性不同 的DNA片段,并把这种DNA片段或它的复本整合为受体基因组的一部分。 受体的遗传重组可以通过三种途径来实现: 1、接合 供体细菌的DNA通过接合管进入受体细菌,实现基因重组。 2、转化 游离的细菌DNA片段被不同的细菌细胞(受体)吸收. 3、转导 一种细菌的DNA片段经过温和的或有缺陷的噬菌体传递给另一种细菌
2
霉菌菌落
3
大肠杆菌
4
二、细菌的突变型
(一)营养缺陷型 生理特性的突变包括:丧失合成某种营养物质能力的营养缺陷型。 (二)抗药突变型 抗性突变包括:抗药性或抗感染性。
5
三、细菌有性杂交
1964年Ledeberg和Tatum用大肠杆菌K,证明细菌有性杂交存在。 (一)大肠杆菌杂交试验
A bio-(生物素) met-(甲硫氨酸) thr+(苏) leu+(亮) thi+(VB1) B bio+(生物素) met+(甲硫氨酸) thr-(苏) leu-(亮) thi-(VB1) A、B都为营养缺陷型,在基本培养基上不能生长。
h+r × hr+
子代噬菌体基因型 h+r hr+ h+r+ hr 含B/B2培养基上噬菌斑 大半透明 小透明 小半透明 大透明 交换值=重组型噬菌体数/总噬菌斑数×100%= h+r++hr/总×100% =小半透明+大透明/总×100% (二)连锁图 T2快速溶菌突变型有多种,如ra、rb、rc都形成大噬菌斑,用不同类 型快速溶菌突变型与宿主范围突变型杂交,结果如下: 杂交组合 h+r hr+ h+r+ hr 重组值 h-r图距 h+ra×hr+ 34.0% 42% 12% 12% 24% 24 h+rb×hr+ 32.0% 56% 5.9% 6.4% 12.3% 12.3 h+rc×hr+ 39.0% 59% 0.7% 0.9% 1.6% 1.6 按图距h、ra、rb、rc有4种排列
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2020/2/29
24
1、 普遍性转导
当转导颗粒将内含物注入受体细胞后, 受体细胞就变成了部分二倍体。导入的基 因通过重组,再整合到受体细胞的染色体 上。
由于这一类噬菌体能够转移细菌染色体 的任何部位,因而被称为普遍性转导。
2020/2/29
25
1、 普遍性转导
• 两个基因被包装在一个转导颗粒中转导, 称为共转导(cotransduction)。
• 转导分为两种:普遍性转导和特异性转导。
2020/2/29
20
1、 普遍性转导
• E.Coli可以通过细胞与细胞的接合而交换 遗传物质,那么,鼠伤寒沙门氏菌是否也 有同样的现象存在呢? 用沙门氏菌的两个突变株杂交,一个是 phe—try—tyr—,另一个是met—his—。两菌 株分别培养时,没有发现野生菌落。将两 菌株在基本培养基上混合培养时,大约10— 5个细胞可以得到一个原养型菌落。这似乎 与E.coli的重组没有什么不同。
2020/2/29
17
二、噬菌体的基因重组
• 根据表7-6中的结果,可得四种可能的连 锁图(见图7-23)
• 要确定到底是那一种,还缺条件。若知道 r的b和排列rc顺之序间。的距离,就可以推知rb、rc和h
2020/2/29
18
二、噬菌体的基因重组
• 为此,需作rb+rc × rb rc+ 。结果rb与 rc之间的距离大于rb与h之间的距离,可 知h应位于rb与rc之间,即rbhrc。
T偶列噬菌体感染细菌时,尾丝附着在 E.coli表面时,通过尾鞘的收缩将DNA经中空 的尾部注入宿主细胞。DNA进入宿主细胞以后, 随即破坏宿主的遗传物质,并借助宿主细胞的 代谢系统,转而合成大量的噬菌体DNA 和蛋白 质,组装成许多许多新的小噬菌体。最后使宿 主细胞裂解(lysis),一下子释放出数百个 子代噬菌体,
11
二、噬菌体的基因重组
• 两个基因型不同的噬菌体同时感染一个宿 主细胞,叫做混合感染(mixed infection) 或双重感染(double infection)。
• 共同生存在同一个宿主细胞中的两个噬菌 体的DNA也可以发生交换,产生基因重组。
2020/2/29
12
二、噬菌体的基因重组
• 比如,一个噬菌体的基因型是a+b-,另一个噬 菌体的基因型是a-b+,同时感染同一个宿主细 胞,宿主细胞裂解以后,可能释放出基因型为 a+b+和a-b-的重组体来。
2020/2/29
14
二、噬菌体的基因重组
• h-和h+均能感染B株。用基因型为r+ h- 和r- h+的两种T2 噬菌体同时感染E.coli B株。这种现象称为双重感染(double infection)。 将双重感染后释放出来的子代噬菌体 接种在同时长有B株和B/2株的培养皿内, 记录噬菌斑的数目和形态。
2020/2/29
6
1 λ噬菌体
• λ噬菌体侵入细菌后,细菌并不裂解,λ噬 菌体的DNA附着于E.coli染色体的gal和bio 位点之间的att位点上(attachment site), 并通过交换而整合到细菌染色体上。整合 以后,就能阻止其它λ噬菌体的超数感染 (superinfection)。整合在寄主染色体中 的噬菌体称为原噬菌体(prophage)。
以后发现这种过滤因子就是噬菌体 P这2一2 。实P验22虽是然一没种有沙证门明氏沙菌门的氏噬菌菌中体有。接 合现象存在,但是却发现了噬菌体介导 的基因转移过程——转导。
2020/2/29
23
1、 普遍性转导
•P22感染细菌细胞时,细菌染色体断裂成许多 小片段。在形成子代噬菌体颗粒时,偶尔会产 生错误,把细菌染色体的片段组合到头部。因 为决定噬菌体感染能力的是外壳蛋白,并不是 外壳所包裹的遗传物质。所以,这种假噬菌体 照样可以吸附到细菌上,并注入其内含物(遗 传物质)。这种内含物并不是噬菌体的遗传物 质,而是细菌DNA的一部分,当然对受体细菌 是无害的。这种假噬菌体称为转导颗粒 (transducing particle)。
研究最深入的噬菌体突变体是T2 噬菌体的r-
(rapid lysis速溶性)突变体。一个正常的T2 噬菌体产生的噬菌斑小而边缘模糊,记为r+,
突变体r-产生的噬菌斑大而边缘清晰。
2020/2/29
13
二、噬菌体的基因重组
• 正常的T2 噬菌体能感染E.coli B株。 突变型E.coli B株能抗T2的感染,记为 B/2株,T2 噬菌体的突变型h-又能克 服B/2株的抗性,既能侵染B株又能侵 染B/2株,形成透明的噬菌斑。正常T2 噬菌体记为h+,只能侵染B株,形成 半透明的噬菌斑。
2
(一)烈性噬菌体(virulent phage)
遗传学上应用最广泛的烈性噬菌体 是大肠杆菌(E.coli)的T偶列噬菌体。 它们的结构大同小异,外貌一般呈蝌蚪 状。T偶列和T奇列有些不同,以T2的结 构最为典型 。
2020/2/29
3
2020/2/29
4
(一)烈性噬菌体(virulent phage)
• P1 噬菌体感染E.coli以后,不整合到细菌 DNA上,而是独立存在于寄主细胞内。P1 DNA可以复制但不裂解宿主细胞,也不影 响宿主细胞的正常代谢,但P1 的复制可以 使宿主的子细胞中也会有P1 DNA,而且可 以多于一个拷贝。
• 受P1 噬菌体感染的细菌也可以因诱导而进 入裂解周期。
2020/2/29
第 二节噬菌体的遗传分析
一、噬菌体的结构 二、T2 噬菌体的基因重组与作图 三、转导
2020/2/29
1
一、噬菌体的结构
• 噬菌体(bacteriaphage or phage)是病 毒的一类,结构很简单,由一个蛋白质 外壳包裹着一些核酸组成的。
• 噬菌体分烈性噬菌体和温和性噬菌体。
2020/2/29
• 若两个基因共转导的频率越高,说明这两 个基因在染色体上的距离愈近,连锁关系 越密切,反之亦然。因此,利用普遍性转 导可以测定细菌基因间的连锁关系。
2020/2/29
26
2、特异性转导
• 特异性转导:噬菌体只转移细菌染色体 的特定部分。源自2020/2/2927
• 至于ra位于h的哪一边,是靠近rc还是靠 近rb?因为T2 DNA是环状的,所以两种答 案都是正确的。
2020/2/29
19
三、转导(Transduction)
• 以噬菌体为媒介,将细菌的小片段染色体 或基因从一个细菌细胞转移到另一个细菌 细胞的过程叫做转导。
• 转导是细菌遗传物质传递和交换的又一重 要方式,它与转化、转导、接合的主要不 同之处在于是以噬菌体为媒介的。
2020/2/29
9
1 λ噬菌体
• 原噬菌体通过诱导(induction)可转变为 烈性噬菌体,进入裂解周期。
• 诱导可以通过不同的方式进行,如UV照射、 温度改变、与非溶原性细菌的接合等。
• 诱导使阻遏物失活,使噬菌体的其他基因 得以表达,促使噬菌体繁殖并进入裂解周 期。
2020/2/29
10
2 P1 噬菌体
2020/2/29
15
2020/2/29
16
二、噬菌体的基因重组
• 重组值= (重组型噬菌斑数/总噬菌斑数) ×100% = (h+r+ + h-r-)/ ( h+r+ + h-r- + h+r
-
+ h-r+)×100% 不同速溶菌突变型的表现型不完全相同,分 别 结记 果为 见教ra、材r表b、7-rc6。。用r-xh+ × r+h-获得的试验
这样的噬菌体称为烈性噬菌体(virulent Phage)。
2020/2/29
5
(二)温和噬菌体 (temperate phage)
• 温和性噬菌体具有溶原性(lisogeny)的 生活周期。这类噬菌体侵入细菌以后,细 菌细胞并不马上裂解。
• 温和性噬菌体有两种类型: 1 λ噬菌体 2 P1 噬菌体
2020/2/29
21
1、 普遍性转导
然而将两菌株放入U型管的两臂,中 间用滤板隔开,以防止细胞接触,但可以 允许比细胞小的生物大分子通过,也获得 了野生型细胞。这说明沙门氏菌的基因重 组不是通过细胞接合,而是通过过滤因子 (filterable agent)而发生的。
2020/2/29
22
1、 普遍性转导
2020/2/29
7
1 λ噬菌体
• 超数感染:一个细菌受一个以上同种噬菌 体感染的现象。
2020/2/29
8
1 λ噬菌体
• λ噬菌体的DNA被整合以后,既不大量复制, 亦不大量转录和翻译。往往只有一两个基 因表达,表达产物作为阻遏物关闭其他基 因的表达。 被溶原性噬菌体感染了的细菌称为溶原性 细菌(lysogenic bacterium)。 当溶原性细菌分裂成两个子细胞时,λ噬菌 体DNA随细菌染色体的复制而复制,每个 细胞中有一个拷贝。