夏培元耐甲氧西林金黄色葡萄球菌的耐药机制及其治疗药
耐甲氧西林金黄色葡萄球菌研究进展

耐甲氧西林金黄色葡萄球菌研究进展本文从网络收集而来,上传到平台为了帮到更多的人,如果您需要使用本文档,请点击下载按钮下载本文档(有偿下载),另外祝您生活愉快,工作顺利,万事如意!金黄色葡萄球菌是造成医院和社区获得血流感染、皮肤及软组织感染的主要致病菌。
自耐甲氧西林金黄色葡萄球菌(methicillin-resistant Staphylococcus aureus,MRSA)检出以来,其耐药范围的扩大和耐药程度的曰益加重,给临床治疗带来较大困难,被称为“超级细菌”。
了解MRSA现状可更好治疗和避免传播。
本文对MRSA的流行病学、危险因素、耐药机制等研究进展综述如下。
1MRSA概念根据2014年CLSI诊断标准,“耐甲氧西林”或“耐苯唑西林”指表达mec或对青霉素酶耐药,当头孢西丁对金黄色葡萄球菌抑菌圈直径4 mg/L时可将其判定对苯唑西林耐药。
近年来,随着抗生素的广泛应用,逐渐出现了对万古霉素中介或耐药MRSA,分别称为VISA(vancomycirr intermediate Staphylococcus aureus )和VRSA (vancomycin—resistant Staphylococcus aureus )。
VISA和VRSA定义为金黄色葡萄球菌对万古霉素的最低抑菌浓度分别为4〜8mg/L和> 16 mg/L。
MRSA具有毒性强、耐药谱广和耐药机制复杂及传播速度快的特点,可分为社区获得性MRSA (community-acquired MRSA,CA-MRSA)和医院获得性MRSA(hospital-acquired MRSA,HA-MRSA)。
CA-MRSA是患者院外无感染MRSA 危险因素和入院72 h 内获得MRSA[1]。
CA-MRSA 是与HA- MRSA不同的表型和基因型一种病原体[2]。
HA- MRSA指在接触过医疗机构的个体间相互传播的MRSA菌株,HA-MRSA感染可在医院内发病,也可在社区内发病。
常见致病菌耐药机制与应对措施
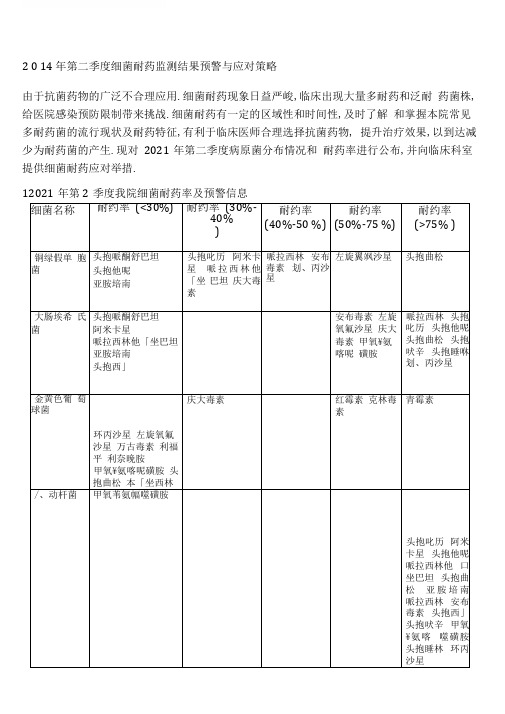
2 0 14 年第二季度细菌耐药监测结果预警与应对策略由于抗菌药物的广泛不合理应用.细菌耐药现象日益严峻,临床出现大量多耐药和泛耐药菌株,给医院感染预防限制带来挑战.细菌耐药有一定的区域性和时间性,及时了解和掌握本院常见多耐药菌的流行现状及耐药特征,有利于临床医师合理选择抗菌药物, 提升治疗效果,以到达减少为耐药菌的产生.现对2021年第二季度病原菌分布情况和耐药率进行公布,并向临床科室提供细菌耐药应对举措.12021年第2季度我院细菌耐药率及预警信息细菌名称耐约率(<30%)耐约率(30%-40%)耐约率(40%-50 %)耐约率(50%-75 %)耐约率(>75% )铜绿假单胞菌头抱哌酮舒巴坦头抱他呢亚胺培南头抱叱历阿米卡星哌拉西林他「坐巴坦庆大毒素哌拉西林安布毒素划、丙沙星左旋翼飒沙星头抱曲松大肠埃希氏菌头抱哌酮舒巴坦阿米卡星哌拉西林他「坐巴坦亚胺培南头抱西」安布毒素左旋氧氟沙星庆大毒素甲氧¥氨喀呢磺胺哌拉西林头抱叱历头抱他呢头抱曲松头抱吠辛头抱睡咻划、丙沙星金黄色葡萄球菌环丙沙星左旋氧氟沙星万古毒素利福平利奈晚胺甲氧¥氨喀呢磺胺头抱曲松本「坐西林庆大毒素红霉素克林毒素青霉素/、动杆菌甲氧苇氨幅噬磺胺头抱叱历阿米卡星头抱他呢哌拉西林他口坐巴坦头抱曲松亚胺培南哌拉西林安布毒素头抱西」头抱吠辛甲氧¥氨喀噬磺胺头抱睡林环丙沙星左旋氧氟沙星庆大毒素肺炎克'番伯菌头抱哌酮舒巴坦阿米卡星哌拉西林他「坐巴坦亚胺培南划、丙沙星左旋氧氟沙星头抱西」庆大毒素安布毒素头抱叱历头抱他呢头抱曲松头抱吠辛头抱睡咻甲氧平氮喀噬磺胺哌拉西林肺炎链球菌头抱叱历左旋氧氟沙星青霉素万古毒素头抱曲松头抱吠辛红霉素克林毒素甲氧¥氨喀呢磺胺表皮葡锢球菌万古霉素利福平利奈晚胺左旋氧氟沙星环丙沙星克林毒素甲氧平氮喀呢磺胺红霉素本「坐西林头抱曲松庆大毒素卡他布兰汉菌头抱哌酮舒巴坦哌拉西林他「坐巴坦左旋氧氟沙星头抱叱历头抱曲松头抱吠辛头抱西」头抱睡林甲氧平氮喀呢磺胺庆大毒素青霉素屎肠球菌万古霉素利奈晚胺红霉素青霉素左旋氧氟沙星利福平环丙沙星流感嗜血杆菌头抱叱历头抱哌酮舒巴坦头抱曲松左旋氧氟沙星利福平克林毒素红毒素头抱吠辛庆大毒素甲氧平氮喀呢磺胺青霉素备注:耐药率超过30%的抗菌药物,提示“预警抗菌药物〞;耐药率超过40%的抗菌药物,提示“慎用抗菌药物〞;耐药率超过50%的抗菌药物,提示“参照药敏试验结果用药〞;耐药率超过75%的抗菌药物,提示“暂停该类抗菌药物的临床应用〞.2细菌产生耐药性机制2.1铜绿假单胞菌耐药机制铜绿假单胞菌对生存环境和营养条件要求很低,在自然界分布广泛,甚至在医院内环境经常可见,其具有多药耐药性及耐药机制:〔1〕该菌能够产生破坏抗菌药物活性的多种灭活酶、钝化酶和修饰酶.〔2〕基因突变,作用靶位变异.〔3〕细胞膜通透性降低.〔4〕主动泵出机制将进入的药物排到体外.〔5〕产生生物膜,阻隔白细胞、多种抗体及抗菌药物进入细菌细胞内吞噬细菌.由于铜绿假单胞菌复杂的耐药机制导致其感染具有难治性和迁延性.2.2大肠埃希氏菌耐药机制大肠埃希菌是G-杆菌中别离率较高的时机致病菌,可引起人体所有部位的感染并且呈多重耐药性.(1)&内酰胺酶的产生①大肠埃希菌对B -内酰胺类抗菌药物耐药主要是由超广谱B -内酰胺酶〔ESBLs〕引起的, 对头霉素类及碳青霉烯类药物敏感. ESBLs可分为五大类:TEM型、SHV型、CTX-M 型、OXA 型和其他型,大肠埃希菌ESBLs酶以TEM型最常见.TEM型ESBLs呈酸性, 可水解头抱他咤、头抱曝历.SHV型ESBLs呈碱性,有水解头抱嚷吩的航基. CTX-M 型ESBLs呈碱性,对头抱曝肠水解水平强于头抱他咤.OXA型ESBLs呈弱酸性或弱碱性,主要水解底物是苯嘎西林,OXA型酶主要见于铜绿假单胞菌中,在大肠埃希菌中的别离率较低.②AmpC&内酰胺酶AmpC酶主要作用于头抱菌素类抗菌药物,且不能被克拉维酸抑制. 它是水解酶,与&内酰胺环竣基局部共价结合,在水分子作用下导致&内酰胺环开环,破坏&内酰胺类抗菌药物抗菌活性.③对酶抑制剂药的耐药的 &内酰胺酶对酶抑制剂药的耐药的 &内酰胺酶〔IRT〕主要有TEM 系列衍变而来,又称为耐酶抑制剂TEM系列酶.〔2〕药物作用靶位的改变〔3〕主动外排〔4〕外膜通透性的下降2.3肺炎克雷伯杆菌耐药机制肺炎克雷伯杆菌属于阴性杆菌,通常存在于人类肠道、呼吸道,是除大肠埃希氏菌外导致医源性感染的最重要的条件致病菌.由于抗菌药物的大量使用,在选择性压力下多药耐药肺炎克雷伯杆菌〔KPN〕菌株不断出现,耐药率日益上升,KPN耐药机制包括:〔1〕产抗菌药物灭活酶①&内酰胺酶包括产超广谱 &内酰胺酶〔ESBLs〕、AmpC酶、耐酶抑制剂&内酰胺酶、碳青霉烯酶〔KPC酶〕及金属&内酰胺酶〔MBLs〕等.ESBLs是耐药KPN产生的最主要的一类酶,由质粒介导,产ESBLsKPN对青霉素类、头抱菌素类及单环类药物耐药,但对头霉素类和碳青霉烯类及酶抑制剂敏感.KPN中AmpC酶由质粒介导,AmpC酶使KPN对临床上广谱&内酰胺类药物耐药, 且不能被&内酰胺酶抑制剂克拉维酸、舒巴坦抑制,但对碳青霉烯类敏感.产耐酶抑制剂&内酰胺酶菌株对青霉素类及青霉素与 &内酰胺酶抑制剂克拉维酸、舒巴坦的复合制剂均不敏感,可被他噪巴坦抑制.碳青霉烯酶〔KPC酶〕不能被EDTA抑制,由质粒介导.产KPC酶KPN对广谱头抱菌素类、单环类及碳青霉烯类耐药.2021年外国学者Eleman等发现2例携带KPC酶的泛耐药KPN甚至对多粘菌素和替加环素耐药.金属&内酰胺酶〔MBLs〕能被EDTA抑制,包括IMP、VIM、NDM-1等类型,前两者最常见且分布广泛.产MBLsKPN对头抱菌素类、单环类以及碳青霉烯类耐药.由于其具有较强耐药性和传播性,被媒体称为“超级细菌〞.②氨基糖昔修饰酶〔AMEs〕 KPN对氨基糖昔类耐药的主要机制是产AMEs,该酶可修饰抗菌药物分子中某些保持抗菌活性所必需的基团,使其与作用靶位核糖体的亲和力大为降低而导致耐药.修饰酶包括N-乙酰转移酶〔AAC〕、腺昔转移酶〔ANT〕、磷酸转移酶〔APH〕等.多药耐药KPNAMEs阳性率高达86.9%.〔2〕抗菌药物渗透障碍①细菌生物被摸〔BF〕形成BF是细菌适应环境形成的一种保护性生存方式,是细菌吸附于生物医学材料或机体黏膜外表,分泌多糖蛋白复合物后缠绕并聚集不同细菌隐藏其内部而形成的膜状物.由于BF具有屏障作用,其内部细菌得以被保护,从而逃脱免疫和抗菌药物的杀伤作用,其内部细菌得以被保护,从而逃脱免疫和抗菌药物的杀伤作用, 使其失效.BF菌的特殊结构和生理特性使菌体内抗菌药物浓度显着降低,而低于致死量的抗菌药物更易诱导细菌产生 &内酰胺酶而导致耐药.外膜孔蛋白的改变革兰阴性菌细胞外膜上存在由许多微孔蛋白组成的孔道,溶质可通过孔道进入外周间隙.假设微孔蛋白改变或缺失,那么抗菌药物难以渗入细菌细胞内,导致耐药.孔蛋白的改变能提升细菌耐药性,与灭活酶并存时的耐药程度较灭活酶单独作用时高,可降低KPN 对碳青霉烯类药物的敏感性.〔3〕主动外排机制主动外排以ATP供能,主动将渗入细菌细胞内的抗菌药物泵出胞外, 外排底物包括&内酰胺类、大环内酯类、唾诺酮类等抗菌药物.〔4〕抗菌药物作用靶位改变①DNA旋转酶和拓扑异构酶构象变异DNA旋转酶和拓扑异构酶构象发生变化,使抗菌药物不能与酶-DNA复合物结合,不能阻止细菌DNA的合成.引起KPN对曜诺酮类耐药.另一种机制是细菌产生由质粒介导的耐药基因qnr,其编码蛋白与拓扑异构酶IV特异性结合,从而减少了唾诺酮类药物的作用靶点,导致细菌耐药.②16srRNA甲基化酶细菌通过产16srRNA基因甲基化酶使药物作用靶位(16srDNA ) 甲基化,导致甲基化的16srDNA与氨基糖昔类药物的亲和力下降而耐药.产16srRNA 甲基化酶的细菌对氨基糖甘类泛耐药.(5)基因盒-整合子系统与细菌耐药性的获得和传播相关的基因元件,即整合子.它是能捕获外源基因使其转变为功能性基因的移动性DNA分子.常位于染色体、质粒或转座子上,并能在其间移动.I、II和田类整合子为耐药整合子.KPN中的整合子多为I、出类,II类整合子携带率降低.整合子可以在菌种间流动,在KPN多药耐药性的介导及传递中具有重要意义,促进KPN不断出现新的耐药方式,对临床治疗提出了严峻挑战.2.4鲍曼不动杆菌耐药机制鲍曼不动杆菌广泛分布于外界环境,主要存在水和土壤中,易在潮湿环境中生存, 如浴盆、肥皂盒等处.该菌粘附力极强,易在各类医用材料上粘附,成为潜在贮菌源. 此外,该菌还存在于健康人皮肤、咽部,也存在于结膜、唾液、胃肠道及阴道分泌物中. 感染源可以是患者自身(内源性感染),亦可以是鲍曼不动杆菌感染者或带菌者,尤其是双手带菌的医务人员.(1)产生抗菌药物灭活酶:①&内酰胺酶:最主要的是D组的OXA-23酶,局部菌株还携带超广谱&内酰胺酶(ESBLs)、头抱菌素酶(AmpC)和B类的金属&内酰胺酶;②氨基糖甘类修饰酶:由于各种修饰酶的底物不同,可导致一种和几种氨基糖甘类抗生素耐药.(2)药物作用靶位改变:拓扑异构酶gyrA、parC基因突变导致的唾诺酮类抗菌药物耐药;armA等16SrRNA甲基化酶导致几乎所有氨基糖甘类抗生素耐药.(3)药物到达作用靶位量的减少:包括外膜孔蛋白通透性的下降及外排泵的过度表达. 鲍曼不动杆菌基因组显示,其富含外排泵基因,外排泵高表达在鲍曼不动杆菌多重耐药中发挥重要作用.鲍曼不动杆菌能泵出的药物包括(3内酰胺类,氨基糖甘类,红霉素,氯霉素,四环素类,氟唾诺酮类,甲氧节咤.主动外排机制在鲍曼不动杆菌对碳青酶烯类耐药的机制中也有一定的作用.鲍曼不动杆菌的耐药相当复杂,其本身具有的多重耐药机制造成它的多重耐药性,从其他菌株获得耐药基因并能表达,抗菌药物的强大选择压力,是造成它对药物耐药性不断上升的主要原因.2.5金黄色葡萄球菌耐药机制(1)青霉素耐药的金黄色葡萄球菌:产生 &内酰胺酶,水解青霉素中有效基团.(2)甲氧西林耐药金黄色葡萄球菌(MRSA):获得MeeA基团,编码产生PBP2a, 对&内酰胺类抗菌药物敏感性减低.〔3〕万古霉素耐药的金黄色葡萄球菌〔VRSA〕:获得万古霉素耐药肠球菌的耐药基因, 使万古霉素失去作用位点;或是细胞壁增厚,使万古霉素不能到达作用靶位.2.6肺炎链球菌耐药机制〔1〕伊内酰胺类抗生素的耐药机制①青霉素结合蛋白〔PBPS〕的改变;②非青霉素结合蛋白因素:非PBPs基因相关的肺炎链球菌耐药菌株都是感受态缺陷菌株.在哌拉西林耐药菌株中存在一个非PBPs的耐药基因cpoA〔糖基转移酶〕,其变异使肺炎链球菌在〔3 -内酰胺类抗菌药物破坏了细胞壁肽聚糖合成的情况下仍能生存,从而导致耐药;murM基因变异与肺炎链球菌青霉素高水平耐药及头抱菌素耐药相关;肺炎链球菌获得性TEM基因可能是其对〔3-内酰胺类药物耐药的原因之一.〔2〕肺炎链球菌对大环内酯类的耐药①erm基因介导核糖体靶位修饰:耐药肺炎链球菌可合成Erm酶〔erm基因编码〕,将核糖体23SrRNA的2058位的腺喋吟残基N26位二甲基化,从而使大环内酯类抗生素与核糖体作用位点的亲和力下降,可引起大环内酯类抗生素高水平耐药;②mef基因介导的主动外排机制:大环内脂类外排基因〔mef〕编码的产物是一种外排泵,能以大环内脂类抗生素为底物,通过消耗能量将药物排出菌体外,从而降低细菌对药物的敏感性.MefA外排泵的特异性底物是十四元环和十五元环大环内酯类,mef基因介导的耐药都发生在M型耐药细菌,且一般是低水平耐药〔1〜32止g/mL〕临床上可以通过增加大环内脂类抗生素的剂量来克服此耐药;③50S核糖体突变:50S核糖体中23SrRNA任一位点的突变都会引起大环内酯类抗菌药物与核糖体结合亲和力的改变,从而导致肺炎链球菌的耐药,突变的核糖体数量与耐药水平呈正相关.〔3〕肺炎链球菌对唾诺酮类药物的耐药机制:肺炎链球菌对氟曜诺酮类药物耐药主要由两个机制介导:一是DNA促旋酶及拓扑异构酶IV唾诺酮耐药决定区域〔QRDR〕染色体变异;二是外排泵作用增加.3应对举措3.1铜绿假单胞菌〔PAE〕耐药应对举措3.1.1抗菌药物不能盲目使用,应根据药敏试验的结果选择应用.3.1.2铜绿假单胞菌〔PAE〕对碳青霉烯类抗菌药物美罗培南、亚胺培南耐药率最低,其次对含酶抑制剂的复合抗菌药物头抱哌酮/舒巴坦、哌拉西林/他嘎巴坦耐药率较低,对头抱他咤耐药率也较低,临床应从其中选择敏感性强且性价比高的药物来治疗PAE感染;虽然PAE对阿米卡星和氯霉素耐药率较低,但由于前者的肾毒性和后者对造血系统的抑制作用,临床医师很少选择应用.3.2大肠埃希菌〔ECO〕耐药应对举措3.2.1氟唾诺类药物已不适用于对大肠埃希菌感染的经验治疗,用药需慎重.3.2.2对头抱毗肠与碳青霉烯类药物的使用应严格掌握适应症,碳青霉烯类药物仅适用于大肠埃希菌所致的严重感染.3.2.3氨基糖甘类药物与〔3内酰胺酶抑制药是治疗大肠埃希菌感染较好的选择.3.3肺炎克雷伯杆菌〔KPN〕耐药的治疗策略3.3.1轻、中度感染:首选复方〔3 -内酰胺类/ %内酰胺酶抑制剂.次选氨基糖昔类与头霉素类抗菌药物联合治疗.3.3.2对上述治疗效果不佳者,可选用碳青霉烯类抗菌药物.3.3.3对严重的产ESBLs肠杆菌科细菌感染者,首选碳青霉烯类抗菌药物或联合治疗方案. 3.4针对鲍曼不动杆菌耐药的治疗策略3.4.1碳青霉烯类抗生素可通过细菌外膜上有通透性功能的孔蛋白,进入胞质间隙后与靶蛋白PBP结合,通过干扰细胞壁的合成从而杀菌,因其对大多数&内酰胺酶具有高度稳定性,所以在临床治疗中能取得较好的疗效,故临床医生可将碳青霉烯类抗生素作为经验治疗鲍曼不动杆菌引起的感染的一线药物,同时应高度注意微生物实验室的实验结果,结合药敏结果合理的调整使用有效抗菌药物.3.4.2舒巴坦可以抑制细菌产生的多种 &内酰胺酶和多数超广谱 &内酰胺酶的同时,它还可以直接作用于细菌的青霉素结合蛋白PBP2,从而显示出它对不动杆菌的独特杀菌作用.3.4.3由于外膜通透性下降及主动外排机制也是细菌的耐药机制,这使得联合用药治疗成为可能.多西环素和粘菌素联合可能是最正确选择,但临床可操作性及真正疗效报道甚少. 我们在选择联合用药方案时,一方面要考虑到药物的MIC值,另一方面应尽可能了解当地的主要耐药表型,便于针对不同的耐药机制,合理联合用药.3.4.4对于泛耐药鲍曼不动杆菌,也不一定就是无计可施,分析目前的泛耐药鲍曼不动杆菌,发现在全耐药中含舒巴坦的制剂有相当的比例为中介,选用相对大剂量的舒巴坦〔4〜6g/d〕有效,这说明,如果有中介药物,加大其剂量对泛耐药鲍曼不动杆菌也是有望取得疗效的.另一方面是新近研发的新型四环素类 -替加环素,因其可以克服所有的泵出机制,虽然应用的经验不多,却可以作为一种很具有潜力的新药.3.5金黄色葡萄球菌耐药应对策略3.5.1在MRSA高度流行的医院或科室,对疑似重度MRSA感染患者,首先经验性选择抗MRSA药物治疗,再根据药敏结果调整到相应敏感的药物.3.5.2不同器官的MRSA感染,疾病的严重程度不同,选择的药物、剂量、疗程、给药途径有较大差异:①皮肤感染:社区获得性MRSA所致的脓疱病,建议局部应用夫西地酸或莫匹罗星进行治疗.周围无蜂窝组织炎的小脓肿在切开引流后一般不需要抗菌药物治疗.②皮肤溃疡并感染:皮肤MRSA定植较感染更为常见,单纯的皮肤溃疡只需局部应用夫西地酸或莫匹罗星治疗.并发蜂窝组织炎、邻近部位骨髓炎或菌血症的患者,或糖尿病患者足部溃疡部位有MRSA定植的患者,应考虑针对MRSA进行全身治疗.③蜂窝组织炎/外科伤口感染:轻症感染建议应用多西环素和克林霉素治疗.多西环素和克林霉素耐药菌株的感染,应选择糖肽类或利耐噪胺,无磺胺过敏者也可选择复方新诺明;对于较严重的感染或者有菌血症高度风险的患者,建议应用静脉输注糖肽类、利耐噪胺或达托霉素治疗.如果考虑存在混合感染〔例如糖尿病足感染〕,且MRSA为主要病原体, 可以考虑单独应用替加环素治疗;有关联合治疗的临床试验极少,且联合治疗有增加药物毒性的风险,不建议任何联合治疗方案.利福平联合夫西地酸治疗皮肤软组织感染的不良反响明显,不建议这种联合.插管部位的感染:对伴有明显硬结、蜂窝组织炎或菌血症的静脉输注部位的严重感染,推荐静脉应用糖肽类或利耐噪胺治疗,轻症感染可以口服药物治疗.③对于单纯的泌尿系感染,建议根据体外药敏结果来选用味喃妥因、甲氧节胺喀咤、复方新诺明等口服药物治疗.对于复杂的泌尿系感染,建议用糖肽类或达托霉素治疗.④MRSA骨和关节感染应以外科综合治疗为根底.建议静脉应用糖肽类单独治疗或联合经静脉应用利福平或夫西地酸钠作为首选方案.急性假体MRSA感染,早期〔病症出现2d内〕手术对保存假体很重要.对于慢性假体感染,应进行外科清创、取出假体.没有证据说明任何单药或联合用药更具优势.⑤应用糖肽类或利耐噪胺治疗MRSA菌血症,疗程至少14d.并发感染性心内膜炎或具有发生感染性心内膜炎高危因素者应延长疗程至6周.利耐噪胺疗程一般不超过4周, 如需延长疗程需注意其不良反响.达托霉素可以作为万古霉素的替代选择.⑥应用糖肽类或利耐噪胺治疗MRSA引起的肺部感染;不伴肺炎的慢性化脓性肺病或支气管扩张症患者,抗MRSA治疗的临床意义尚不明确;利耐噪胺有良好肺组织穿透力, 可用于此类患者的治疗.⑦对MRSA引起的深部眼睛感染和中枢神经系统感染, 建议应用万古霉素单独或联合利福平治疗,根据药敏也可选用利耐噪胺或复方新诺明;对静脉治疗无效的MRSA脑膜炎, 可以考虑万古霉素鞘内注射;庆大霉素、夫西地酸钠或氯霉素可以用于敏感细菌引起的眼睛浅部感染.⑧外科手术感染的预防性用药:对有MRSA定植史或感染史且未去除者,或有MRSA 带菌的高危风险者,在接受外科手术时需接受糖肽类预防感染.如估计患者有重新出现MRSA带菌的可能或患者来自MRSA高流行的机构,建议使用糖肽类治疗;对无MRSA 定植的患者,建议应用氨基糖甘类预防葡萄球菌感染.3.6肺炎链球菌耐药应对策略3.6.1一般PISP 〔中介水平耐药〕仍可选用青霉素治疗,但应加大剂量;PRSP所致的下呼吸道感染可选用阿莫西林500mg/克拉维酸钾125mg 〔675mg〕配方的方案治疗.3.6.2奎奴普丁30/达福普丁70,该药为复合制剂,属链阳菌素类抗菌药,协同抗菌活性增强为单药的8〜16倍,用于PISR PRSP 〔耐青霉素肺炎链球菌〕、MRSP 〔耐红霉素肺炎链球菌〕感染治疗.3.6.3利奈噪胺,该药为恶噪烷酮类新型化学合成抗菌药,对PISR PRSP均有高度抗菌活性.特点为有静脉与口服制剂,口服吸收完全,与静脉给药相比,其绝对生物利用度达100%,适合于序贯疗法.由于作用部位和方式独特,〔选择性结合于50s亚单位核糖体、抑制细菌蛋白质合成〕,不易与其他抑制细菌蛋白质合成的抗菌药发生交叉耐药, 体外也不易诱导产生耐药性,此为该药的优点.3.6.4用药动学/药效学参数标准抗菌药的使用行为PRSP/MRSP别离率不断上升,与头抱菌素与大环内酯类抗菌药的大量使用以及治疗方案不合理有很大关系.如头抱曲松钠对PRSP本有很强抗药作用,但单剂量治疗,使治疗后的血药浓度虽高于抗敏感的肺炎链球菌但低于抗耐药肺炎链球菌的浓度,助长了耐药的增长.而MRSP那么主要是广泛应用大环内酯类抗菌药所致.4.结语对免疫力低下的住院患者采取支持疗法,增强患者抵御感染的水平;早期诊断,快速进入手术,缩短住院时间,减少医院内交叉感染的时机;围手术期合理使用抗菌药物, 以免造成新的耐药菌株出现;严格执行消毒隔离制度和无菌技术,定时通风、净化室内空气;医护人员在接触患者时应戴工作帽、口罩、鞋套,标准洗手方法;医疗操作应做到准确细致,可减少组织损伤;外科患者手术前备皮手法应轻柔细致,保护患者皮肤的自身免疫屏障;重视引流管的清洁,及时冲洗和更换引流管;严格执行家属探视制度, 减少家属逗留时间等.我院应继续增强抗菌药物治理,各个部门增强合作,标准抗菌药物使用,杜绝滥用抗菌药物.增强医务人员对院内感染的熟悉和防控,增强对医院的工作人员培训,标准操作,减少耐药菌的传播流行.另外,还应监测医院的细菌感染谱及耐药情况.通过各方面的综合举措,相信在对耐多药细菌的防治会获得更多的经验和更新的进展. 药剂科医院感染科微生物室2021年07月。
甲氧西林耐药金黄色葡萄球菌肺炎的诊断与治疗

甲氧西林耐药金黄色葡萄球菌肺炎的诊断与治疗作者:刘又宁、施毅来源:中华医学院呼吸病学分会感染学组耐甲氧西林金葡菌(methicillin-resistantmethicillin-resistant Staphylococcus aureus,MRSA)是引起医院获得性和社区相关性感染的重要病原菌,自1961年首次发现MRSA以来,其在金葡萄中分离率不断上升,据2009年中国CHINET细菌耐药性监测报告,我国10省市14所教学医院临床分离出的4 114株金葡菌中MRSA比率高达52.7% (2010年的情况类似,4 452株金葡菌中MRSA的比率为51.7%)。
已知MRSA是医院获得性感染最重要的革兰阳性球菌,国外已有报道对万古霉素耐药的金葡菌(vancomycin-resistant Staphylococcus aureus,VRSA),而更为令人震惊的是近年来世界各地发现了致病力极强的社区获得性MRSA,防治形势极为严峻。
MRSA 肺炎是临床最为常见的感染类型之一,其诊治面临极大的挑战,需要引起大家的重视。
一、定义呼吸系统MRSA感染主要有社区相关MRSA肺炎(community-associated MRSA pneumonia)和医院相关MRSA肺炎(hospital associated MRSA pneumonia),后者包括呼吸机相关肺炎(ventilator-associated pneumonia,VAP)、卫生保健相关肺炎(healthcare associated pneumonia,HCAP)。
社区相关MRSA肺炎(CA-MRSA肺炎):又称为社区获得性MRSA肺炎(community-acquired MRSA pneumonia),是指肺炎患者在门诊或入院48 h内分离出MRSA菌株,并且在1年内无住院或与医疗机构接触史,无MRSA感染或定植史,无留置导管和其他经皮医用装置使用史。
耐甲氧西林金黄色葡萄球菌感染抗菌药物作用机制、分子结构、药物选择、治疗措施及联合用药注意事项

耐甲氧西林金黄色葡萄球菌感染抗菌药物作用机制、分子结构、药物选择、治疗措施及联合用药常用抗MRSA药物万古霉素临床常用糖肽类抗菌药物,主要用于治疗革兰阳性球菌感染,尤其是 MRSA 感染一线治疗。
抗 MRSA 作用主要通过抑制 MRSA 细胞壁合成,抑制 MRSA 细胞质内 RNA 合成及影响 MRSA 细胞膜通透性实现。
常见不良反应包括肾毒性、耳毒性、红人综合征、变态反应等。
监测万古霉素药物谷浓度和药时曲线下面积以提高疗效和降低肾毒性。
对普通感染成人万古霉素目标谷浓度维持在10|—15 mg/L,AUC 0—24 h 的目标范围在 400—650mg·h·L-1。
去甲万古霉素药物结构比万古霉素少一个甲基,抗菌谱和抗菌活性与万古霉素相同,不良反应也相似,抗感染成本明显低于万古霉素。
替考拉宁与万古霉素同属糖肽类,是常用重要的抗革兰阳性菌感染药物,广泛用于治疗耐药革兰阳性菌所致的各类感染。
分子结构与万古霉素相似,主要作用机制为特异性与细胞壁前体肽聚糖结合,阻断细胞壁合成,引起细菌细胞壁缺陷,导致细菌死亡。
常见不良反应一般轻微且短暂,严重不良反应较罕见。
药物蛋白结合率为90%—95%,消除半衰期长达47—100 h,肾功能不全者消除半衰期进一步延长,血液透析和腹膜透析不能清除药物。
利奈唑胺药物是噁唑烷酮类化学合成抗菌药物,主要用于治疗耐青霉素和耐多药肺炎链球菌肺炎、MRSA 所致复杂和单纯皮肤和软组织感染或耐万古霉素肠球菌感染。
作用机制为选择性与核糖体50s亚单位结合,阻碍起始复合物形成,干扰蛋白质合成,起到抑菌作用。
独特作用机制使利奈唑胺与其他抗菌药物之间不易产生交叉耐药,主要不良反应是味觉改变、血小板减少及白细胞减少,长期服用还可能引发骨髓抑制。
常用治疗MRSA 药物中,仅利奈唑胺有口服剂型。
达托霉素具有环状结构的脂肽类化合物,临床主要用于治疗革兰阳性球菌引起的复杂皮肤软组织感染,以及葡萄球菌引起的菌血症、心内膜炎。
金黄色葡萄球菌耐药的现状及临床治疗对策

金黄色葡萄球菌耐药的现状及临床治疗对策孙宏莉徐英春单位:中国医学科学院中国协和医科大学北京协和医院摘要:金黄色葡萄球菌,尤其是甲氧西林或苯唑西林耐药的金黄色葡萄球菌(MRSA)是引起院内感染的多重耐药菌。
目前MRSA已逐渐成为全世界院内感染的主要病原菌。
目前在许多国家仍在增长,MRSA几乎对所有?-内酰胺类抗生素耐药,甚至累及到红霉素,环丙沙星和庆大霉素。
本文对金黄色葡萄球菌的耐药现状、耐药机制以及金黄色葡萄球菌感染的危险因素和治疗对策进行了简要介绍。
关键词:金黄色葡萄球菌耐药现状耐药机制危险因素治疗对策答1. 甲氧西林或苯唑西林耐药的金黄色葡萄球菌( MRSA )是引起院内感染的多重耐药菌。
是吗?A.是B.不是2. 根据 NCCLS 判定标准,对于金黄色葡萄球菌,万古霉素的MIC≤4ug/ml 时为敏感。
对吗?A.不对B.对金黄色葡萄球菌,尤其是甲氧西林或苯唑西林耐药的金黄色葡萄球菌( MRSA )是引起院内感染的多重耐药菌。
在第一个耐青霉素酶的β-内酰胺类抗生素甲氧西林用于临床治疗葡萄球菌感染不久, 1961 年在英国发现了世界首例 MRSA 。
从此, MRSA 逐渐成为全世界院内感染的主要病原菌。
目前在许多国家仍在增长, MRSA 几乎对所有β- 内酰胺类抗生素耐药,甚至累及到红霉素,环丙沙星和庆大霉素。
1996年在日本首次发现了MRSA对万古霉素敏感性下降的菌株,即万古霉素中介的金黄色葡萄球菌(VISA)。
VISA 的出现预示着万古霉素治疗葡萄球菌感染的临床疗效下降,随之万古霉素耐药的金黄色葡萄球菌(VRSA)临床株是否会分离到,成为人们监测和关注的热点。
如果葡萄球菌一旦对万古霉素耐药,临床将如何治疗?所以连续监测金黄色葡萄球菌的耐药现状,了解葡萄球菌的耐药机制,感染的危险因素及其治疗对策具有重要的临床意义。
金黄色葡萄球菌的耐药现状:我院细菌室从 1988~2003 年连续监测金黄色葡萄球菌对常用抗生素的耐药性变迁,结果见表 1 。
耐甲氧西林金黄色葡萄球菌耐药机制分析

1.3 MRSA 检测:采用纸片扩散法(KB 法)与最低 抑菌浓 度 法 (MIC 法)两 种 方 法 进 行 MRSA 检 测。 KB法使 用 头 孢 西 丁(FOX)药 敏 纸 片 检 测 MRSA, 药敏结果 判 读 参 照 美 国 临 床 实 验 室 标 准 化 委 员 会
(CLSI)2017 版 抗 微 生 物 药 物 敏 感 试 验 执 行 标 准 ,放 在(35±2)℃ 的 专 门 培 养 箱 里 面 48h 之 后 ,测 出 纸 片 附近的抑菌 圈 直 径,比 如:FOX 纸 片 直 径 ≤21 mm, 即判 为 MRSA,FOX 纸 片 直 径 ≥22 mm 者 判 为 MSSA,介于二者之间可进行 MIC法筛选。 1.4 mec基 因 检 测:①DNA 模 板 的 提 取:在 高 压 灭 菌 的 试 管 内 将 待 测 菌 株 用0.9% 氯 化 钠 注 射 液 调 到2.0(1.8~2.2)麦 氏 单 位 菌 悬 液,沸 水 中 煮 15 min,吸取1000μL 菌悬液于编好号的35个 EP 管
为:80V90 mA、连 上 电 源,在 恒 压 情 况 下 电 泳,之 后,通过凝胶成像系统来得到成像,且将结果存留。 1.5 基 因 测 序:采 取 mec 基 因 且 阳 性 较 好 的 菌 株,将其 PCR 产 物 送 英 潍 捷 基 (上 海)贸 易 企 业 做 测序,之后,将得到 的 数 据 和 GenBank 里 面 的 相 关
1 材 料 与 方 法 1.1 菌株来 源:实 验 所 用 菌 株 来 源 山 西 医 学 科 学 院 、山 西 大 医 院 从 临 床 标 本 分 离 的 剔 除 重 复 与 污 染
文献[3],由 上 海 生 工 生 物 工 程 公 司 合 成。 引 物 序 列 见 表 1。
重要致病菌的耐药机制

重要致病菌的耐药机制公司标准化编码 [QQX96QT-XQQB89Q8-NQQJ6Q8-MQM9N]重要致病菌的耐药机制1.甲氧西林耐药葡萄球菌(MRS) 金葡菌可产生一种特殊的青霉素结合蛋白PBP2a,其与β内酰胺类抗生素的亲和力减低,因而产生耐药性(MRSA)。
PBP2a 由mecA 基因编码,由转座子携带并整合至葡萄球菌染色体的mec 部位。
每株MRSA 菌都有mecA 基因,而敏感株则无。
MecRI-mecl 是调节基因,通过抑制mecA 的转录决定PBP2a 的合成水平而调节细菌的耐药程度。
在细菌基因组中还存在着辅助基因femA、femB、femC、femD,与甲氧西林耐药性的表达有关。
这些辅助基因与mecA 基因协调作用,产生对β内酰胺类抗生素的高度耐药。
MecA 基因广泛分布于金葡菌及凝固酶阴性葡萄球菌。
带有mecA 基因的菌株对青霉素类、头孢菌素类、单环β内酰胺类抗生素均呈耐药。
由于其所在的转座子常带有对其他抗生素的耐药基因,使耐甲氧西林葡萄球菌常可对红霉素、四环素类、夫西地酸、磺胺药、链霉素等氨基苷类及氟喹诺酮类同时耐药,但上述抗生素的耐药机制则各不相同。
1996 年欧洲首次报道了对万古霉素不敏感金葡菌,MIC 为8~16ug/ml,称为VISA(Vancomycin intermediate suscepitble 或GISA(Glycopeptide intermediate susceptible S aureus),其机制尚未完全阐明,可能由于该菌产生过多的靶位,阻断了药物到达靶位,使之不能起抗菌作用。
2.耐万古霉素肠球菌肠球菌属对万古霉素的耐药性共有6 种基因型,即VanA、VanB、VanC1、VanC2/C3、VanD和VanE。
其中VanA、VanB、VanD、VanE为获得性耐药,VanC1和VanC2/C3为固有的耐药性。
万古霉素与细菌肽聚糖前体末端的D-丙氨酰-D-丙氨酸结合,抑制细胞壁肽聚糖的合成。
耐甲氧西林金葡菌

5 浓度梯度(Etest)法
是1988年AB Biodisk公司推出, 在含20 g/L NaCl的MH琼 脂平板上, 贴上苯唑西林的试条, 菌液调至0.5~1麦氏 浊度, 35°C孵育24 h, 直接读取MIC值。MIC<2 μg/ml 为敏感, >4 μg/ml为耐药。Etest法结合了纸片扩散法 和肉汤稀释法的优点, 长塑料条含有连续的呈指数梯度 变化的苯唑西林(0.016~256 μg/ml), 故在检测低水平 或中等程度耐药的MRSA时结果更为准确。Novak[11]等 报道, 用Alamar法和Etest法对127株MRSA的检测比较, 两者结果相关较高, 用Etest法检测127株MRSA, 其中93 株MIC>256 μg/ml, 28株在6~256 μg/ml, 检出率达 96%, Etest法具有精确、可靠、稳定性好的特点, 但缺 点是价格昂贵。
2 获得性耐药
是质粒介导的耐药。某些菌株通过耐药因子产生大量β内酰胺酶, 使耐酶青霉素缓慢失活, 表现出耐药性[7], 多为临界耐药。
耐甲氧西林金黄色葡萄球菌(MRSA)的分型 MRSA分型对追踪传染源,研究型别与感染种类,耐药性的关 系有重要作用。国外开展较早的噬菌体分型,将待测菌于肉 汤中,35°C孵育6 h,涂于分型琼脂平板上,待干后将23种 噬菌体注入琼脂平板中的小方格内,再置35°C孵育6 h后移 至室温过夜观察结果。用4组23种噬菌体,将MRSA分为4群, 一般以Ⅰ群为最多[8],也有报告以Ⅲ群为多。噬菌体分型 结果常不满意,日本小粟 子证实有29.3%菌株不能分型,且 重复性差,不宜用于流行病学调查。质粒图谱分型较为可靠, 可分为18个型,能准确地分析菌株之间的相关性,将流行菌 株与非流行菌株加以区别。国内MRSA广泛存在分子量为1.6 Md、1.8 Md及2.67 Md的质粒,不同地区和不同医院会有特殊 质粒带。免疫印迹分型法将MRSA分为9个型,以B.C型为最常 见,各型含有特征性的分子带,该法比较稳定。染色体限制 性内切酶分析可识别病原体DNA链上特异位点及核苷酸序列, 能从基因水平显示病原体特征,MRSA还可用血清学、凝固酶、 耐药谱等方法分型。现在Southern印迹法也逐渐运用于MRSA 的分型。
耐甲氧西林金葡菌(MRSA)耐药机制的研究进展

耐甲氧西林金葡菌(MRSA)耐药机制的研究进展
夏培元;张慧琳
【期刊名称】《四川生理科学杂志》
【年(卷),期】1997(000)003
【摘要】耐甲氧西林金黄色葡萄球菌(Methicillin-Resistant Staphylococcus aureas,MRSA)自1960年被发现报道以来,到80年代后期已成为一个重要的全球性病原体,为院内感染的最常见致病菌,在一些大型教学医院中可占所有S.aureus感染的60~80%。
特别是在氟喹诺酮类药物环丙沙星(Ciprofloxacin)用于临床后,迅速出现的S.aureus耐药株中MRSA的比例超过90%。
临床上MRSA表现为对所有的β-内酰胺类抗生素耐药,并可能通过从某些肠球菌处获得质粒而扩大其耐药谱或增强其耐药性,给临床上治疗其感染造成了极大的困难。
故对其
【总页数】2页(P17-18)
【作者】夏培元;张慧琳
【作者单位】[1]华西医科大学临床药研究所;[2]华西医科大学临床药研究所
【正文语种】中文
【中图分类】R96
【相关文献】
1.MRSA的耐药机制及防治研究进展 [J], 段思琪;崔莎莎;李月;孙丽媛;陈堃;李明成
2.多重耐药性金黄色葡萄球菌(MRSA)的临床药物治疗及耐药机制研究 [J], 陈方圆;马笑雪;蔡景钰;王斯;罗恩杰
3.MecA基因及非MecA基因在MRSA耐药机制中的研究进展 [J], 汤兰兰;林洁如
4.MRSA耐药机制与分子生物学检测方法研究新进展 [J], 杨长顺;刘文恩
5.细胞壁的厚度可能是MRSA对吖啶黄素的耐药机制 [J], 李文赟
因版权原因,仅展示原文概要,查看原文内容请购买。
耐甲氧西林金黄色葡萄球菌耐药性分析

耐甲氧西林金黄色葡萄球菌耐药性分析刘广丽【摘要】目的了解临床分离的耐甲氧西林金黄色葡萄球菌(MRSA)耐药性.方法对临床分离的72株MRSA用K-B纸片扩散法测定对青霉素等16种抗菌药物的敏感性.结果 72株MRSA对万古霉素、替考拉宁均敏感,对复方新诺明、呋喃妥因、利福平、四环素、左氧氟沙星、克林霉素、阿奇霉素的耐药率分别为18.1%、20.8%、80.6%、84.7%、88.9%、93.1%、95.8%.对红霉素、庆大霉素、阿米卡星、青霉素、氨苄西林/舒巴坦、头孢西丁、氨苄西林均耐药.结论临床分离的MRSA为多重耐药菌.【期刊名称】《泰山医学院学报》【年(卷),期】2008(029)007【总页数】2页(P518-519)【关键词】耐甲氧西林金黄色葡萄球菌;耐药性;医院感染【作者】刘广丽【作者单位】泰山医学院附属泰安医院检验科,山东,泰安,271000【正文语种】中文【中图分类】R730.43葡萄球菌属是引起医院感染的重要致病菌,特别是耐甲氧西林金黄色葡萄球菌(MRSA)比例的上升[1],使葡萄球菌属对各种抗菌药物产生多重耐药。
临床用于MRSA感染的首选药物为糖肽类抗菌药物,由于该类细菌对多种抗菌药物耐药,造成临床治疗困难,已引起临床普遍关注与重视[2]。
本实验对临床分离的MRSA 耐药性进行研究,现报道如下。
1 材料与方法1.1 菌株来源 72株MRSA来自我院2007年1月~2007年7月临床各类标本,主要来自重症监护病房。
1.2 菌株鉴定及药敏试验全部菌株使用法国梅里埃ATB细菌鉴定仪鉴定菌种。
万古霉素、替考拉宁、复方新诺明、呋喃妥因、利福平、四环素、左氧氟沙星、克林霉素、阿奇霉素、红霉素、庆大霉素、阿米卡星、青霉素、氨苄西林/舒巴坦、头孢西丁、氨苄西林纸片为英国Oxoid公司产品,血浆凝固酶胶乳试剂为生物梅里埃产品。
纸片扩散法药敏试验按NCCLS规程操作金黄色葡萄球菌;头孢西丁抑菌圈≤21 mm耐药,≥22 mm为敏感;并根据美国临床实验室标准化委员会(NCCLS)2004年版要求进行抗菌药物敏感性判断。
甲氧西林耐药金黄色葡萄球菌感染与对策

皮肤、鼻前庭、直肠等是MRSA的主要定植部
位;地面、洗手盆、止血带、血压计的袖带等医院工作 区均可发现MRSA。MRSA以接触为主要传播模 式,医务人员的手更是病人间传播的主要媒介。 MRSA的不均一耐药性,给检测带来困难。常
年7月至2005年12月,共发生MRSA侵袭性感染
8987例,病死率6.3/10万;其中,CA—MRSA占 58.4%,医院感染的MRSA(HA—MRSA)占26.6%,
intermediate
作者单位:100730中国医学科学院中国协和医科大学北京协 和医院感染科(马小军),检验科(徐英春)
通信作者:马小军,Email:drmaxiaojun@刚1.corn
resistant
S.aureus,VISA)感染病例,但截止到2006
MRSA已经给公众健康带来了严重威胁。医务 人员不仅应该重视感染病例的诊断、治疗,更应从 MRSA产生的源头加以控制,特别应避免抗菌药物 的滥用,减少MRSA产生的压力。卫生部已于2007 年11月27日签署并启动了以“清洁医疗是更安全 医疗”为主题的“全球患者安全倡议活动”。在以预 防为主的医疗模式大背景下,为患者提供清洁、优质 的医疗服务,从而预防和控制MRSA感染的发生与 播散至关重要。
20.
准防护和接触传播防护最为重要。
通常的预防与控制措施有LJ引:不与他人共用私 人物品、注意洗手、戴手套、清洁被服和环境等u1|。 做好手卫生是关键环节。床单、面巾、睡衣、餐具等
[7]SaM—Salim B,Mathema B,Kreiswirth
methiciHin—resistant Staphylococcus
生堡内整盘壶:嫂!至垒旦筮生!鲞筮垒翅£照垫』!!!!婴丝型,』!塑!嫂!:y生:笪,塑!:垒
耐甲氧西林金葡菌(MRSA)治疗药物

耐药性监测与控制
定期进行耐药性监测了解MRS的耐药情况 合理使用抗生素避免过度使用和滥用 加强医院感染控制防止MRS的传播 推广使用新型抗生素提高治疗效果
不良反应及处理方法
过敏反应:可 能出现皮疹、 瘙痒、呼吸困 难等症状应立 即停药并就医。
胃肠道反应: 可能出现恶心、 呕吐、腹泻等 症状可适当调 整剂量或更换
耐甲氧西林金葡菌 (MRS)治疗药物的
疗效评估
临床疗效评价标准
治疗效果:观察药物对MRS感染的治疗效 果包括治愈率、好转率等
安全性:评估药物的安全性包括不良反应、 过敏反应等
耐药性:评估药物对MRS的耐药性包括耐 药率、耐药机制等
药物相互作用:评估药物与其他药物的相 互作用包括协同作用、拮抗作用等
用药建议
用药原则
遵循医嘱:根据医 生建议选择合适的 药物和剂量
合理用药:避免滥 用抗生素防止耐药 性产生
定期监测:定期检 查血常规、肝肾功 能等指标确保药物 疗效和安全性
联合用药:根据病 情需要与其他药物 联合使用提高疗效
联合用药方案
抗生素:选择敏感性高的抗生素如万古霉素、利奈唑胺等 抗真菌药:对于合并真菌感染的患者可考虑使用抗真菌药 免疫调节剂:如干扰素、白介素等提高机体免疫力 辅助用药:如止咳、退热、止痛等药物缓解患者症状
国外研究进展: 国外学者在耐甲 氧西林金葡菌 (MRS)治疗药物 方面也取得了重 要进展如发现新 型抗菌药物、开 发新型治疗方法 等。
合作研究进展: 国内外学者在耐 甲氧西林金葡菌 (MRS)治疗药物 方面开展了广泛 的合作研究共同 推进了该领域的 发展。
未来研究方向: 未来耐甲氧西林 金葡菌(MRS)治 疗药物的研究方 向将更加注重新 型抗菌药物的研 发、新型治疗方 法的开发以及合 作研究的开展。
耐甲氧西林金黄色葡萄球菌的耐药机制及研究现状

其 中 内 酰胺 酶 为 MRS 产 生 的 主 要 酶 类 。B内 酰 胺 酶 可 A 一 以 特 异 性 的 的打 开 药 物分 子 结 构 中 的 口内 酰 胺 环 , 其 完 全 一 使 失 去抗 菌 活 性 ; 可 与 某 些 抗 口内 酰 胺 酶 青 霉 素 结 合 , 药 也 一 使 物 停 在 浆 膜 外 隙 , 而 体 现 耐 药 性 。MR A 的 主要 耐 药 机 制 从 S 是 由产 生 低 亲 和 力 的 P P a 代 替 了 其 他 S 菌 株 产 生 的 B 2, A PP B 。P P a很 少 或 不 与 0内 酰 胺 类 抗 生 素 结 合 , 且 通 过 B2 一 并
1 MRS 的 流 行 特 点 A
金 黄 色 葡 萄 球 菌 由 于 获 得 了 携 带 meA 基 因 的葡 萄 球 c 菌 盒式 染 色 体 ( C me) 并 通 过 其 不 断 积 累 抗 生 素 耐 药 基 S C c,
因 。近 年 来 , 内 外 学 者 已 从 金 黄 色 葡 萄 球 菌 中检 测 出 耐 消 国 毒 剂 的 菌 株 。 _ 6 研 究 发 现 S C c可 分 为 I、 C me Ⅱ、 、 、 型 , C me Ⅲ Ⅳ V SC c
其 所 产 生 的 内 酰 胺 酶 水 解 药 物 , 使 在 p 内 酰 胺 类 抗 生 素 即 一
[] a esHF Th h n ige ie oo yo tp yooc sa — 3Chmbr . eca gn pdmilg fsa h lccu u ruE] megIfc Di,0 1 7 1 8 e sJ ?E r net s2 0 , :7 . [ ] 键 奇 , 龙 洙 . 葡 菌 调 节 基 因 meA 高 表 达 对 苯 锉 西 林 耐 药 4连 崔 金 c 性 的 影 响 [] 中 华 微 生 物 和 免疫 杂 志 ,0 6 2 ( ) 6 3 J. 2 0 ,6 7 :0 . [] 支密 , 5黄 仵 蕾. S MR A对 消毒 剂 盒 常 用 抗 菌 药 的 耐 药 性 及 相 关
耐甲氧西林金黄色葡萄球菌耐药机制及检测方法研究进展

综㊀㊀述耐甲氧西林金黄色葡萄球菌耐药机制及检测方法研究进展∗李雪寒综述,李一荣ә审校(武汉大学中南医院检验科,湖北武汉430071)㊀㊀摘㊀要:耐甲氧西林金黄色葡萄球菌(M R S A)广泛传播是全球重要的公共卫生问题.葡萄球菌染色体m e c 基因盒元件(S C C m e c)在金黄色葡萄球菌中转移是导致M R S A广泛传播的重要原因.S C C m e c中的m e c A和m e c C基因分别编码P B P2a和P B P2c,其结合βG内酰胺类抗菌药物能力低下,是导致M R S A发生的关键因素,而且m e c A和m e c C介导的耐药表型存在一定差异.本文就S C C m e c的基本结构与功能㊁m e c A与m e c C介导的耐药机制与表型以及M R S A的检测方法等方面做以下综述.关键词:耐甲氧西林金黄色葡萄球菌;㊀m e c A;㊀m e c C;㊀头孢西丁;㊀苯唑西林D O I:10.3969/j.i s s n.1673G4130.2019.05.020中图法分类号:R446.5文章编号:1673G4130(2019)05G0586G05文献标识码:AA d v a n c e s i n t h e r e s i s t a n c em e c h a n i s ma n dd e t e c t i o nm e t h o d s o fm e t h i c i l l i nGr e s i s t a n t S t a p h y l o c o c c u s a u r e u s∗L IX u e h a n,L IY i r o n gә(D e p a r t m e n t o f C l i n i c a lL a b o r a t o r y,Z h o n g n a nH o s p i t a l,W u h a nU n i v e r s i t y,W u h a n,H u b e i430071,C h i n a)A b s t r a c t:T h ew i d e s p r e a do fm e t h i c i l l i nGr e s i s t a n t S t a p h y l o c o c c u s a u r e u s(M R S A)i s a n i m p o r t a n t g l o b a l p u b l i c h e a l t h p r o b l e m.T h e t r a n s f e r o f S t a p h y l o c o c c a l C a s s e t t eC h r o m o s o m em e c(S C C m e c)a m o n g S t a p h y l oGc o c c u s a u r e u s i s a k e y r e a s o n t h a tM R S As p r e a dw i d e l y.T h em e c Aa n dm e c C l o c a t e d i nS C C m e c e n c o d e PB P2a a n dP B P2c r e s p e c t i v e l y.T h e l o wa f f i n i t y f o r b e t aGl a c t a m s o f P B P2a a n dP B P2c i s a p i v o t a l f a c t o r l e a d i n g t o t h e o c c u r r e n c e o fM R S A.I na d d i t i o n,t h e r e a r e s o m ed i f f e r e n c e sb e t w e e n t h em e c AGm e d i a t e dm e t h i c i l l i nr e s i s t a n t p h e n t y p e a n d t h em e c CGm e d i a t e d.T h i s p a p e r r e v i e w s t h eb a s i cs t r u c t u r ea n df u c t i o no fSC C m e c,t h em e c AGa n dm e c CGm e d i a t e dm e t h i c i l l i n r e s i s t a n t p h e n t y p e s a n dm e c h a n i s ma n d t h e d e t e c t i o nm e t h o d s o fM R S A.K e y w o r d s:m e t h i c i l l i nGr e s i s t a n t S t a p h y l o c o c c u s a u r e u s;㊀m e c A;㊀m e c C;㊀c e f o x i t i n;㊀o x a c i l l i n㊀㊀金黄色葡萄球菌常定植于皮肤和鼻腔,致病能力强,能导致皮肤软组织感染㊁中耳炎㊁心内膜炎及菌血症等多种感染.20世纪40年代初期,青霉素G应用于临床后显著改善了金黄色葡萄球菌感染的治疗疗效.但是,1942年,临床上就发现了耐青霉素G的金黄色葡萄球菌.1959年,为治疗耐青霉素G的金黄色葡萄球菌感染,临床上引入了甲氧西林,一种半合成的耐青霉素酶的βG内酰胺类抗菌药物,它对产青霉素酶金黄色葡萄球菌有良好的杀伤作用.1961年, J E V O N S在英国首次证实了耐甲氧西林金黄色葡萄球菌(M R S A)的存在,此后,临床上M R S A菌株的检出率逐年增长.到1985年,M R S A开始出现暴发性流行,成为全球院内感染的首要病原体.在2011年,仅在美国一个国家,M R S A就造成了超过8万例的侵袭性感染[1].M R S A感染的高发生率和死亡率已经导致巨大的社会和经济损失.现就M R S A的流行状况㊁耐药机制和检测方法等研究进展做一综述.1㊀M R S A的流行状况㊀㊀自1961年首株M R S A发现以来,临床上M R S A 的检出率逐年升高,到20世纪80年代后期,M R S A已成为全球发生率最高的院内感染病原菌之一.在印度,1999年M R S A的检出率已达80.8%,到2004-2006年,更是增长到了86.5%[2].在欧洲的罗马尼亚,2015年,M R S A的检出率达到57.2%[3].还有一些地区,M R S A的检出率虽然不高但呈持续增长趋势.例如澳大利亚,M R S A的检出率由2000年的12.0%上升到2013年的19.0%.在欧洲的塞浦洛斯和斯洛伐克,M R S A的检出率从2012年的35.2%和21.7%分别上升到了2015年的43.4%和28.1%[4].在我国,从2005年中国细菌耐药性监测开始以来,在检出的革兰阳性菌中,金黄色葡萄球菌所占比例一直都是第1位,在33.0%左右波动,而M R S A又是耐药性金黄色葡萄球菌中最常见的.虽然自2005年以来,我国的M R S A检出率处于下降趋势,但局部区域的685 国际检验医学杂志2019年3月第40卷第5期㊀I n t J L a bM e d,M a r c h2019,V o l.40,N o.5∗基金项目:湖北省自然科学基金面上项目(2016C F B672).ә㊀通信作者,EGm a i l:l i y i r o n g838@163.c o m.㊀㊀本文引用格式:李雪寒,李一荣.耐甲氧西林金黄色葡萄球菌耐药机制及检测方法研究进展[J].国际检验医学杂志,2019,40(5):586G590.M R S A检出率依然在70.0%以上[5].20世纪90年代以来,M R S A所致感染的流行病学发生了显著的变化,不仅局限于医院内,而且呈现出向院外蔓延的趋势,于是提出了M R S A的两种分型,即医院获得性M R S A(HAGM R S A)和社区获得性M R S A(C AGM R S A).H AGM R S A通常引起住院㊁免疫力低下或老年患者的感染,这类人群一般是有手术史㊁经常接触医院卫生设施以及接受抗菌药物治疗的患者.而C AGM R S A则在社区间相互传播,倾向于感染健康的年轻人,这些人在发病前与医疗机构无任何接触.美国德克萨斯州的研究表明,大约70.0%的社区获得性金黄色葡萄球菌的感染均是C AGM R S A[6].此外,C AGM R S A也可以引起医院感染的暴发,这可能是由于C AGM R S A可以产生杀白细胞素(P V L)造成的[7].细菌和患者在社区和医院间不断流动,相互传播,导致了复杂的M R S A流行状况.2㊀M R S A的耐药机制2.1㊀葡萄球菌染色体m e c基因盒元件(S C C m e c)的基本结构和功能㊀金黄色葡萄球菌通常是通过获得S C C m e c而产生对甲氧西林的耐药性.S C C m e c为可移动遗传元件,能在葡萄球菌属内菌株间转移.S C CGm e c的大小为21~67k b,具有高度多态性.目前,世界范围内共发现11种S C C m e c(Ⅰ~Ⅺ型).虽然不同类型的S C C m e c的大小和基因组成有所差异,但仍具有以下共同特征[1]:(1)携带一个m e c基因复合体,由m e c基因及其调控系统组成,与M R S A耐药相关的m e c基因为m e c A和m e c C.(2)携带一个c c r基因复合体,由位于复合体中央的c c r基因和相邻的7~8个开放阅读框(O R F s)组成,其中c c r基因包括c c r A㊁c c r B和c c r C[8].c c r编码的重组酶c c r A B或c c r C具有位点特异性,能识别相对应的S C C m e c元件,使之从葡萄球菌染色体中精确地分离,并通过转移整合到其他葡萄球菌的菌株中,从而实现S C C m e c在葡萄球菌属内菌株间转移[9].如c c r B结合a t t B和a t t S序列能完成S C C m e c的剪切或整合,而c c r A尽管不直接参与S C C m e c的剪切或整合,但起到促进c c r B对序列的识别作用,因而c c r A和c c r B总是成对出现.而c c r C单独出现即可完成S C C m e c的剪切或整合[1]. (3)S C C m e c在o r f X基因(编码r R N A甲基转移酶)3ᶄ端的a t t B序列处整合入金黄色葡萄球菌染色体组,且S C C m e c的插入对o r f X的表达没有影响.此外,S CGC m e c元件还含有3个J区域,主要起连接作用,也可能携带一些其他的抗菌药物耐药决定簇,如转座子T n554(编码对红霉素的耐药)㊁质粒p T181(编码对四环素的耐药)㊁质粒p U B110(编码对氨基糖苷类的耐药)等基因元件.J区域的结构差异性是S C C m e c分型的依据[10].2.2㊀m e c A基因介导的耐药机制㊀m e c A大小为2129b p,位于Ⅰ~Ⅹ型S C C m e c上,编码一种新的青霉素结合蛋白P B P2a.P B P2a与βG内酰胺类抗菌药物的亲和力很低.因此,P B P2a不受βG内酰胺类抗菌药物的影响,继续发挥细胞壁黏肽合成酶的功能,催化细胞壁的合成,从而导致M R S A的产生[11].m e c A 的表达受其相邻的m e c R1Gm e c I系统的调控,这个系统与金黄色葡萄球菌中调控βG内酰胺酶表达的b l a R1Gb l a I 系统同源.m e c R1编码感受器蛋白M e c R1,m e c I编码与M e c R1相耦联的抑制蛋白M e c I.M e c R1是一种金属内肽酶原,能通过胞外的青霉素结合区域感受到βG内酰胺类抗菌药物的存在.当βG内酰胺类抗菌药物结合到M e c R1的感受器结构域,M e c R1被激活,其构象发生改变,诱导胞内感受器区域自动裂解,直接或间接导致M e c I分裂,从而阻碍M e c I结合到m e c A启动子区域,去除M e c I对m e c A的抑制作用,m e c A开始表达,生成大量的P B P2a,使细菌产生耐药性[1].2.3㊀m e c C基因介导的耐药机制㊀2007年,在一项关于奶牛乳腺炎的研究中发现了金黄色葡萄球菌菌株L G A251,它具有MR S A的表型,对苯唑西林和头孢西丁耐药,但是对m e c A和P B P2a进行验证时却反复显示为阴性.随后对其作全基因组测序显示, L G A251菌株携带一种新型的m e c A同源基因,起初命名为m e c A L G A251,于2012年更名为m e c C[12].在此之后,在瑞典[13]㊁西班牙[14]㊁斯洛文尼亚[15]等地相继发现m e c C.m e c C大小为2198b p,位于Ⅺ型S C C m e c上,其核苷酸序列与m e c A的同源性为69%.B A L L H A U S E N等[16]用m e c C敲除菌株(W44646Δm e c C)进行实验,发现该菌株对头孢西丁的最小抑菌浓度(M I C)值由16.00μg/m L下降到4.00μg/m L,对苯唑西林的M I C值由8.00μg/m L 下降到0.25μg/m L.将敲除的m e c C恢复后,菌株恢复对头孢西丁和苯唑西林的耐药性(M I C值上升为64.00μg/m L),从而证明了m e c C与M R S A的形成有关.该研究还对m e c A和m e c C进行了对比研究,发现m e c A和m e c C的启动子和操纵子的核苷酸序列也有所不同.m e c A启动子G10区域的序列为T A T A C T,而m e c C该处序列为T A T T A T,该差异可能是导致m e c A启动子的活性在转录水平上高于m e c C启动子的原因.在M e c I的结合位点处,m e c A 操纵子上包含一个30b p的回文序列,由2个紧密相连的15b p序列组成.而m e c C操纵子上的回文结构则被一个8b p的连接肽分为两段14b p的序列.这些差异表明,m e c C的转录调控系统与m e c A的存在差异.m e c C的调控元件为m e c I S C C m e cⅪ和m e c R1S C C m e cⅪ,分别编码m e c C转录抑制蛋白和感受器蛋白.其编码的蛋白与M e c I和M e c R1同源性分别为66%和45%[16].与m e c A类似,m e c C也编码一种青霉素结合蛋白,称为P B P2c.P B P2c与m e c A 编码的P B P2a同源性为63%.K I M等[17]对P B P2c 的性质进行研究,发现P B P2c具有药物偏好性,相对头孢西丁而言,其对苯唑西林的亲和力更高,其对苯唑西林的亲和力是P B P2a的4倍,因此表现为对头孢785国际检验医学杂志2019年3月第40卷第5期㊀I n t J L a bM e d,M a r c h2019,V o l.40,N o.5西丁的高度耐药,对苯唑西林处于中介耐药甚至是敏感水平,而P B P2a则没有这种现象.此外,两者在热稳定性方面也表现出了差异,在37ħ时P B P2a的活性最高且稳定,而P B P2c则在25ħ时表现出稳定的高活性.2.4㊀f e m基因介导的耐药机制㊀除m e c A㊁m e c C之外,金黄色葡萄球菌染色体上还有一些基因也参与M R S A耐药性的产生.这些基因一般与肽聚糖的合成㊁细胞的分裂和细胞壁的代谢相关.f e m就是一种独立于m e c A和m e c C的基因.与m e c A和m e c C不同,f e m既存在于耐药菌中,也存在于敏感菌中.敏感菌中的f e m基因被灭活,可使菌株对βG内酰胺类抗菌药物敏感性提高.目前,已被确定的f e m有6种:f eGm A㊁f e m B㊁f e m C㊁f e m D㊁f e m E和f e m F.如f e m A和f e m B分别编码F e m A和F e m B两种胞质蛋白,分别添加第2个和第3个甘氨酸残基以及第4个和第5个甘氨酸残基到细胞壁上的五甘氨酸肽间桥,从而参与细胞壁五甘氨酸肽间桥的形成.当f e m A和f e m B失活时,五甘氨酸肽间桥被单甘氨酸取代,使细胞壁肽聚糖成分发生改变,细菌对甲氧西林耐药性降低[18].f e m C是谷氨酰胺合成酶抑制因子,当f e m C失活时,谷氨酰胺合成酶基因(g l n A)的转录减少,主干肽链的DG谷氨酸的酰胺化减少,从而使谷氨酰胺生成减少,使细菌细胞壁的正常结构遭到破坏,细菌对甲氧西林的耐药性降低.f e m D编码磷酸葡萄糖胺变位酶,催化葡萄糖G6G磷酸与葡萄糖G1G磷酸的相互转化,f e m D失活会影响肽聚糖前体物质的合成,使细菌对甲氧西林的耐药性降低[19].3㊀M R S A的检测方法㊀㊀快速精准地检测M R S A对临床抗感染治疗具有重要的作用.M R S A检测方法较多,主要包括M RGS A表型检测(苯唑西林以及头孢西丁纸片扩散法㊁苯唑西林琼脂筛选法㊁胶乳凝集法检测P B P2a㊁显色培养基法等)和M R S A耐药基因检测.2008年,美国临床实验室标准化协会(C L S I)将头孢西丁纸片扩散法列为MR S A的推荐检测方法.2011年,D A T T A 等[20]比较了5种M R S A表型检测方法的性能,证明头孢西丁纸片扩散法为一种非常好的M R S A表型检测方法,但他们认为还需要使用其他的方法作为补充来提高检测的灵敏度.实验数据显示,乳胶凝集法的灵敏度为100.0%,特异度为99.2%,因此,推荐使用头孢西丁纸片扩散法与乳胶凝集法联合检测.之后, F A R A H A N I等[21]也通过实验证明了头孢西丁纸片扩散法是临床首选的M R S A检测方法.P C R法检测m e c A基因一直以来被认为是检测M R S A的 金标准 .K O U P A H I等[22]对比了P C R法和4种表型检测方法,结果显示,头孢西丁纸片扩散法和P C R法的灵敏度和特异度均为100.0%,苯唑西林纸片扩散法㊁苯唑西林琼脂稀释法和显色培养基法的灵敏度分别为95.45%㊁97.22%和98.13%,并认为头孢西丁纸片扩散法可以在没有分子检测设备的情况下作为 金标准 的替代方法用于检测M R S A.临床上对于M R S A的检测主要是根据MR S A对苯唑西林和头孢西丁高度耐药的表型以及通过验证m e c A和P B P2a来判定的.但是,对于m e c C M R S A,由于其对头孢西丁高度耐药而对苯唑西林敏感的表型以及m e c A和P B P2a阴性,可能会将其误判为甲氧西林敏感金黄色葡萄球菌(M S S A).因此,在进行检测时,选用头孢西丁进行药敏试验比苯唑西林对m e c C M R S A的检测结果更准确[23].当然,仅根据药敏试验结果检测m e c C MR S A是不够可靠的.与传统M R S A的检测相似,应用P C R技术验证m e c C是检测m e c C M R S A的 金标准 .S T E G G E R等[24]应用多重P C R技术对185株金黄色葡萄球菌同时进行m e c A和m e c C的检测,结果显示灵敏度和特异度均为100%.此外,该项技术还能同时检测P V L毒力基因和进行s p a分型.P I C H O N等[25]应用四重实时P C R技术同时检测m e c A㊁m e c C㊁P V L毒力基因和n u c,此项技术不仅能精确检测出4种基因,并且能将检测时间缩短到1h以内,这为临床上检测MR S A提供了更为快速且全面的方法.B E C K E R等[26]利用X p e r tM R S A G e n3系统联合P C R技术同时检测m e c A㊁m e c C和S C C m e cGo r f X连接区域,这项技术大大增加了M R S A检测的特异度.S E I D E L等[27]将核酸侧流免疫分析(N A L F I A)技术与P C R技术相结合用于检测m e c A和m e c C基因来检测M R S A.相对于普通P C R和实时P C R技术来说,P C R D核酸横向流动免疫检测技术有更精密的检测限度,并可以区分m e c的不同等位基因,且耗时较短,这使M R S A的检测向着更精准快速的方向迈进.4㊀总㊀㊀结㊀㊀M R S A被世界卫生组织列入了2017年 重点致病菌 名单,其所造成的感染在今后相当长时间内依然是棘手的全球公共卫生问题.临床微生物室通常使用纸片扩散法或显色培养法来鉴定M R S A,但是由于m e c A和m e c C的基因序列与调控序列均存在一定差异,金黄色葡萄球菌体内m e c A和m e c C表达的P B P2a和P B P2c结合βG内酰胺类抗菌药物的能力不同,从而呈现不同的耐药表型.m e c A介导的耐药表型已引起高度关注,但m e c C介导的耐药表型(头孢西丁耐药/苯唑西林敏感或轻度耐药)易被忽视甚至误认为甲氧西林敏感,从而导致治疗失败.因此,了解M R S A的耐药机制及其检测方法,特别是m e c C基因介导的耐药机制及相应的检测方法,对于指导临床抗感染治疗具有重要意义.参考文献[1]P E A C O C KSJ,P A T E R S O N G K.M e c h a n i s m so fm e t h iGc i l l i nr e s i s t a n c ei ns t a p h y l o c o c c u sa u r e u s[J].A n n u R e vB i o c h e m,2015,84(1):577G601.885 国际检验医学杂志2019年3月第40卷第5期㊀I n t J L a bM e d,M a r c h2019,V o l.40,N o.5[2]C H E NCJ,HU A N GYC.N e we p i d e m i o l o g y o f s t a p h y l oGc o c c u s a u r e u s i n f e c t i o n i nA s i a[J].C l i n M i c r o b i o l I n f e c t,2014,20(7):605G623.[3]E u r o p e a nC e n t r e f o rD i s e a s eP r e v e n t i o na n dC o n t r o l.A nGt i m i c r o b i a l r e s i s t a n c e s u r v e i l l a n c e i nE u r o p e2015.A n n u a l r e p o r to ft h e E u r o p e a na n t i m i c r o b i a lr e s i s t a n c es u r v e i lGl a n c en e t w o r k(E A R SGN e t)[R].E u r o s u r v e i l l a n c e,2017.[4]D iR U S C I O F,B JØR N H O L TJV,L E E G A A R D T M,e t a l.M R S Ai n f e c t i o n s i nn o r w a y:a s t u d y o f t h e t e m p o r a l eGv o l u t i o n,2006G2015[J].P L o SO n e,2017,12(6):e179771.[5]HU FP,G U O Y,Z HU D M,e t a l.R e s i s t a n c e t r e n d saGm o n g c l i n i c a l i s o l a t e si n C h i n ar e p o r t e df r o m C H I N E T s u r v e i l l a n c eo fb a c t e r i a lr e s i s t a n c e,2005G2014[J].C l i n M i c r o b i o l I n f e c t,2016,22S u p p l1:S9G14.[6]K A L EP,D HAWA NB.T h e c h a n g i n g f a c e o f c o m m u n i t yGa c q u i r e d m e t h i c i l l i nGr e s i s t a n tS t a p h y l o c o c c u sa u r e u s[J].I n d i a n JM e d M i c r o b i o l,2016,34(3):275G285.[7]G R U N D MA N N H,A I R E SGD EGS O U S A M,B O Y C EJ,e t a l.E m e r g e n c e a n d r e s u r g e n c e o f m e t i c i l l i nGr e s i s t a n tS t a p h y l o c o c c u s a u r e u s a sa p u b l i cGh e a l t ht h r e a t[J].L a nGc e t,2006,368(9538):874G885.[8]I T O T,K UWA HA R AGA R A IK,K A T A Y AMA Y,e t a l.S t a p h y l o c o c c a l c a s s e t t e c h r o m o s o m em e c(S C C m e c)a n a lGy s i so f M R S A[J].M e t h o d s M o lB i o l,2014,1085:131G148.[9]WA N GL,A HM E D M H,S A F O M K,e t a l.A P l a s m i dGB o r n es y s t e m t oa s s e s st h ee x c i s i o na n di n t e g r a t i o n o f s t a p h y l o c o c c a lc a s s e t t e c h r o m o s o m e m e c m e d i a t e d b y C c r Aa n dC c r B[J].J B a c t e r i o l,2015,197(17):2754G2761.[10]K O N G H,Y U F,Z HA N G W,e t a l.M o l e c u l a r e p i d e m i o lGo g y a n da n t i b i o t i cr e s i s t a n c e p r o f i l e so f M e t h i c i l l i nGR eGs i s t a n t s t a p h y l o c o c c u s a u r e u s s t r a i n s i n a t e r t i a r y h o s p i t a l i n c h i n a[J].F r o n tM i c r o b i o l,2017,8:838.[11]H I R AMA T S U K,I T O T,T S U B A K I S H I T AS,e t a l.G eGn o m i cb a s i sf o r m e t h i c i l l i nr e s i s t a n c ei ns t a p h y l o c o c c u s a u r e u s[J].I n f e c tC h e m o t h e r,2013,45(2):117.[12]P A T E R S O N G K,HA R R I S O N E M,H O L M E S M A.T h ee m e r g e n c eo f m e c C m e t h i c i l l i nGr e s i s t a n tS t a p h y l oGc o c c u s a u r e u s[J].T r e nd sM i c r o b i o l,2014,22(1):42G47.[13]U N N E R S T A D H E,B E N G T S S O N B,H O R N A F R AGN T Z I E N M,e ta l.M e t h i c i l l i nGr e s i s t a n ts t a p h y l o c o c c u sa u r e u s c o n t a i n i n g m e c Ci nS w e d i s hd a i r y c o w s[J].A c t aV e t S c a n d,2013,55:6.[14]G OM E ZP,G O N Z A L E ZGB A R R I O D,B E N I T O D,e t a l.D e t e c t i o no f m e t h i c i l l i nGr e s i s t a n tS t a p h y l o c o c c u sa u r e u s(M R S A)c a r r y i n g t h em e c C g e n e i nw i l d s m a l lm a m m a l s i nS p a i n[J].JA n t i m i c r o bC h e m o t h e r,2014,69(8):2061G2064.[15]D E R MO T A U,Z D O V CI,S T R UM B E L JI,e ta l.D e t e cGt i o no fm e t h i c i l l i nGr e s i s t a n tS t a p h y l o c o c c u sa u r e u sc a r r yGi n g t h em e c C g e n e i nh u m a n s a m p l e s i nS l o v e n i a[J].E p iGd e m i o l I n f e c t,2015,143(5):1105G1108.[16]B A L L HA U S E N B,K R I E G E S K O R T E A,S C H L E I M E R N,e t a l.T h em e c A h o m o l o g m e c Cc o n f e r sr e s i s t a n c eaGg a i n s tβGl a c t a m s i nS t a p h y l o c o c c u sa u r e u s i r r e s p e c t i v eo ft h e g e n e t i cs t r a i n b a c k g r o u n d[J].A n t i m i c r o b A g e n t sC h e m o t h e r,2014,58(7):3791G3798.[17]K I M C,M I L H E I R IÇO C,G A R D E T ES,e t a l.P r o p e r t i e s o fan o v e lP B P2A p r o t e i nh o m o l o g f r o m S t a p h y l o c o c c u s a u r e u s s t r a i nL G A251a n d i t sc o n t r i b u t i o nt ot h eβGL a cGt a mGr e s i s t a n t p h e n o t y p e[J].JB i o lC h e m,2012,287(44):36854G36863.[18]L IX,X I O N G Y,F A N X,e t a l.T h e r o l eo f f e m Ar e g u l aGt i n gg e n eo n m e t h i c i l l i nGr e s i s t a n tS t a p h y l o c o c c u sa u r e u s c l i n i c a l i s o l a t e s[J].M e d M a lI n f e c t,2012,42(5):218G225.[19]B E R G E RGBÄC H I B,R O H R E R S.F a c t o r si n f l u e n c i n g m e t h i c i l l i n r e s i s t a n c e i n s t a p h y l o c o c c i[J].A r c h M i c r o b iGo l,2002,178(3):165G171.[20]D A T T A P,G U L A T IN,S I N G L A N,e t a l.E v a l u a t i o no f v a r i o u sm e t h o d sf o rt h ed e t e c t i o no f m e t i c i l l i nGr e s i s t a n t s t a p h y l o c o c c u sa u r e u ss t r a i n sa n ds u s c e p t i b i l i t yp a t t e r n s [J].JM e d M i c r o b i o l,2011,60(P t11):1613G1616.[21]F A R A HA N IA,MO HA J E R IP,G H O L AM I N EB,e t a l.C o m p a r i s o no fd i f f e r e n t p h e n o t y p i ca n d g e n o t y p i cm e t hGo d s f o r t h ed e t e c t i o no fm e t h i c i l l i nGr e s i s t a n tS t a p h y l o c o cGc u s a u r e u s[J].N A mJM e dS c i,2013,5(11):637G640.[22]K O U P A H I H,H O N A R MA ND J A H R OMY S,R A HGB A R M.E v a l u a t i o n o f d i f f e r e n t p h e n o t y p i c a n d g e n o t y p i cm e t h o d s f o rd e t e c t i o no f m e t h i c i l l i nr e s i s t a n ts t a p h y l oGc o c c u s a u r e u s(M R S A)[J].I r a nJP a t h o l,2016,11(4):370G376.[23]S K O V R,L A R S E N A R,K E A R N S A,e t a l.P h e n o t y p i cd e t e c t i o no fm e c CGM R S A:C e f o x i t i n i sm o r e r e l i a b l e t h a no x a c i l l i n[J].JA n t i m i c r o bC h e m o t h e r,2014,69(1):133G135.[24]S T E G G E R M,A N D E R S E NPS,K E A R N SA,e t a l.R a pGi dd e t e c t i o n,d i f f e r e n t i a t i o na n dt y p i n g o f m e t h i c i l l i nGr eGs i s t a n t S t a p h y l o c o c c u s a u r e u sh a r b o u r i n g e i t h e rm e c Ao r t h en e w m e c Ah o m o l o g u em e c AL G A251[J].C l i nM i c r oGb i o l I n f e c t,2012,18(4):395G400.[25]P I C H O N B,H I L L R,L A U R E N T F,e ta l.D e v e l o p m e n t o f a r e a lGt i m e q u a d r u p l e xP C Ra s s a y f o r s i m u l t a n e o u s d eGt e c t i o no f n u c,p a n t o nGv a l e n t i n e l e u c o c i d i n(P V L),m e c Aa n dh o m o l o g u e m e c A L G A251[J].J A n t i m i c r ob C h eGm o t h e r,2012,67(10):2338G2341.[26]B E C K E R K,D E N I S O,R O I S I N S,e ta l.D e t e c t i o no f m e c AGa n dm e c CGp o s i t i v em e t h i c i l l i nGr e s i s t a n t s t a p h y l oGc o c c u s a u r e u s(M R S A)I s o l a t e sb y t h en e w X p e r tM RGS A g e n3P C Ra s s a y:T A B L E1[J].JC l i n M i c r o b i o l,2015,54(1):180G184.[27]S E I D E L C,P E T E R SS,E S C H B A C H E,e ta l.D e v e l o pGm e n t o f an u c l e i ca c i dl a t e r a l f l o wi m m u n o a s s a y(N A LGF I A)f o r r e l i a b l e,s i m p l e a n d r a p i d d e t e c t i o n o f t h em e t h iGc i l l i n r e s i s t a n c e g e n e sm e c Aa n dm e c C[J].V e tM i c r o b i o l,2017,200:101G106.(收稿日期:2018G09G15㊀修回日期:2018G12G02)985国际检验医学杂志2019年3月第40卷第5期㊀I n t J L a bM e d,M a r c h2019,V o l.40,N o.5。
重要病原菌的耐药性机制及其治疗策略

重要病原菌的耐药性机制及其治疗策略医学上,病原菌指能引起疾病的微生物。
随着抗生素使用的增加和滥用,很多病原菌出现了耐药性,这使得治疗感染疾病变得更加困难。
本文将探讨一些重要病原菌的耐药性机制以及治疗策略。
一、耐药性机制1.金黄色葡萄球菌金黄色葡萄球菌(S. aureus)是最常见的病原菌之一,在医院中也是常见的致病菌。
该菌引起的疾病包括皮肤和软组织感染、肺炎、败血症等。
耐甲氧西林金黄色葡萄球菌(MRSA)是一种对常见抗生素甲氧西林产生耐药性的S. aureus。
MRSA耐药性主要是由于其产生了一种酶称为β-内酰胺酶(β-lactamase),该酶能分解甲氧西林和其他β-内酰胺类抗生素分子,使其不能起到杀菌作用。
此外,MRSA具有改变其表面蛋白的能力,从而使人体免疫系统对其失去识别能力,也是其耐药性的重要机制。
2.大肠杆菌大肠杆菌(E. coli)是人体内正常微生物群落的一种成员,同时也是一种常见的致病菌。
大肠杆菌引起的疾病包括尿路感染、腹泻、败血症等。
耐氨基糖苷大肠杆菌是迄今发现的对氨基糖苷类抗生素产生耐药性的大肠杆菌。
大肠杆菌耐药性主要是由于其产生了一种酶称为氨基糖苷酶(aminoglycoside-modifying enzymes),该酶能改变氨基糖苷类抗生素的结构,使其不能起到杀菌作用。
此外,大肠杆菌也可以通过改变其细胞膜通道的结构减少抗生素的进入,从而产生耐药性。
3.肺炎链球菌肺炎链球菌(S. pneumoniae)是引起肺炎、中耳炎和鼻窦炎等疾病的常见病原菌。
耐青霉素肺炎链球菌(PRSP)是一种对青霉素类抗生素产生耐药性的S. pneumoniae。
PRSP耐药性主要是由于其改变了青霉素结构中与链球菌交互作用的目标蛋白——青霉素结合蛋白(penicillin-binding protein),使青霉素无法与目标蛋白结合起作用。
此外,PRSP还可以减少细胞膜中笔形草菌素的通道,从而减少药物进入细胞的速度,产生耐药性。
耐甲氧西林金黄色葡萄球菌感染及其耐药性研究

海 峡药 学 2018年 第 3O卷 第 8期
情况 ,抗 菌药物 的不合理 使用 和抗 菌药 物大量使 用易增 加细 菌 耐药 性 ,使 医院感 染控制难度增加 ,使用微生 物检验方式实 施药敏 试验 ,能 根据相关试验结果选择抗 菌药物 ,能最大程度 减少 患者的耐药性 ,将治疗有效性提 高 ,再通过使用 微生物学 检查方 式对环境 消毒灭 菌情况 、医疗 器械灭 菌情况 实施全 面 监测 ,能从 根源上对 医院感染情 况进 行控 制 。临 床研究 中 显示 ,由于免疫抑制剂 、耐药菌株增多 、患者 因素等 影响 ,导致 医 院感染发 生率增加 ,而通 过进行微生物检验 工作 ,加强科 室 环境和 医疗器 械的消毒灭菌监测 ,指导抗生 素合理应用 ,能显 著 降低 医院的感染发生率 ,减轻每位住 院治疗患者感染程 度 , 使 患者能早期康复 。
验医学杂志 ,2015,36(9):1317,后插 2. [3]孙应贤 .微生物检验在医院感染 控制 中的价值探讨 [J].今 日健
康 ,2016,15(12):350-350. [4]张伟 .微生物检验在感染控制 中的价值分析 [J].中国医药指南 ,
2015,09(24):351-352. (5]李美丽 .微生物检验在 医院感 染控制 中的临 床意义分 析 (J).临
综上所述 ,通过对住 院治疗患 者实施微生物检验 ,不仅 能 降低感染发生 率 、控 制 院 内感 染 情况 ,还 能 改善 患者 感 染程 度 ,值得在 医院感染控制 中进 一步推广和应用 。
耐甲氧西林金葡菌耐药分子机制及其研究进展

耐甲氧西林金葡菌耐药分子机制及其研究进展摘要】耐甲氧西林金葡菌已成为医院及社区感染的主要病原菌之一。
经重组获得的mec复合体是其产生耐药性的结构基础,并通过产生PBP2a表达抗药性,而金葡菌耐药性表达及表达程度还受其内在的固有基因影响,本文重点讨论MRSA耐B内酰胺类抗生素的分子机制,包括其耐药决定性基因mecA基因,以及fem基因等其他辅助基因进行重点介绍。
【关键词】耐甲氧西林金葡菌耐药机制进展【中图分类号】R943 【文献标识码】A 【文章编号】2095-1752(2014)08-0295-02自20世纪60年代在欧美首先发现了耐甲氧西林金葡菌以来,MRsA在临床分离的葡萄球菌中的比例不断增加,其已成为医院感染的重要病原菌,现已证实MRSA与其他葡萄球菌具有相同的毒力和致病力[1]并常见其流行、暴发的报道。
重症监护病房(ICU)是引起院内MRSA感染的主要场所。
此外社区获得性MRsA亦逐年增多。
由于MRsA往往具有多药耐药(multidmg resistance,MDR)特征,甚至出现了对万古霉素耐药的临床分离株(VRSA),当氟喹诺酮类药物应用于临床后,迅速出现的金葡菌耐药株中MRSA的比例已超过90%。
[2]1、耐药机制MRSA的耐药性主要有2个来源:其一,是由于质粒介导的DNA转导、转化、或其他类型的DNA插入,导致p-内酰胺酶产生,属获得性耐药;其二,是由染色体DNA介导的固有耐药性,主要是mec基因编码的PBP2a的耐药性。
近年来对该方面研究较多。
1.1PBP2a由染色体介导(主要是对p-内酰胺类的耐药性),MRSA特有的mecA基因大量表达一种特殊的青霉素结合蛋白PBP2a,PBP2a与B一内酰胺类抗生素结合活性很低,可替代高亲和力的PBPs行使功能,是引起B一内酰胺类抗生素耐药的主要原因。
1.1.1青霉素结合蛋白细菌细胞壁的主要化学成分是肽聚糖,又称粘肽。
在粘肽合成过程中,相邻糖链中的两个五肽侧链的交联由转肽酶催化。
耐甲氧西林金黄色葡萄球菌耐药机制及基因分型的研究进展

・132・・综述・耐甲氧西林金黄色葡萄球菌耐药机制及基因分型的研究进展山西大学生物技术研究所(030006)王慧山西省人民医院张爱珍随着一种半合成青霉素——甲氧西林在1960年被首次应用于临床以来,仅仅1年之后在英国就发现了首例耐甲氧西林金黄色葡萄球菌(methicillin-r鹤istanceStaphylococcusaureus,MRSA),之后以惊人的速度在世界范围内蔓延。
成为医院内最常见的病原菌…。
MRSR感染最大危害在于它是医院感染的重要病源,院内一旦发生流行,往往迅速传播,较难控制,此外社区内获得性MRSA亦逐年增多【2J。
由于MRSA往往具有多重耐药(muhidrugresistance,MDR)特征,甚至出现了对万古霉索耐药的临床分离株(VRSA)[3],再加上其致病性强,己成为临床抗菌感染治疗的难题之一,与艾滋病、乙型肝炎并列为世界三大严重感染性疾患【3],并成为各国研究的热点。
因此。
探究MRSA耐药分子机制及其基因分型,寻找治疗MRSA感染的新药已非常必要。
1耐药机制MRSA耐药机制复杂,大致分为2类:①由质粒介导的DNA转导、转化、或其他类型的DNA插入,导致8-内酰胺酶产生,属获得性耐药;②由染色体DNA介导的固有耐药性,主要是mec基因编码的青霉素结合蛋白(PBP)2a的耐药性。
1.1获得性耐药由质粒介导,金黄色葡萄球菌过度产生B-内酰胺酶或甲氧西林酶。
高浓度的青霉素酶能缓慢水解甲氧两林或其他耐舻内酰胺酶抗生素,但过量产生p内酰胺酶尚不足以提供甲氧西林抗性,还需具有特定的宿主遗传背景。
1.2固有耐药性1.2.1PBP2a:对甲氧西林耐药和敏感的金黄色葡萄球菌产生5种PBPs:PBPI、PBP2、PBP3、PBP3’和PBP4,它们的功能主要是以转肽酶的形式在肽聚糖上四肽侧链与甘氨酸五肽交联的过程中发挥催化作用,从而参与细胞壁黏肽层的合成。
而哥内酰胺类抗生素通过与PBP结合,形成稳定的酯附着在其活性位点上。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
1 夏培元.耐甲氧西林金黄色葡萄球菌的耐药机制及其治疗药物的研究进展[J.国外医药抗生素分册,1999,20(4):1782 180.
2 Cook N.M eth icillin2resistant S tap hy lococcus au reus versus the burn patient[J.Burns,1998,24(2):91298.
3 Stranden AM,Eh lert K,L abisch insk i H,et al.Cell w all monoglycine cro ss2bridge and m eth icillin hypersuscep tibility
in a femAB nullm utant of m eth icillin2resistant S tap hy lococcus au reus[J.J Bacteri o l,1997,179(1):9213.
4 P iriz2D uran S,Kayser FH,Bergere2Bach i B.I mpact of sar and agr on m eth icillin resistance in S tap hy lococcus au reus[J.
FE M S M icrobi o l L ett,1996,141(223):255.
5 Kom atsuzaw a H,O h ta K,L abisch insk i H,et al.Characteri2 zati on of fm tA,a gene that modulates the exp ressi on of m eth i cillin resistance in S tap hy lococcus au reus[J.A nti m icrob
A gents Chemo ther,1999,43(9):212122125.
6 T am bic A,Pow er EG,T am bic T,et al.Ep idem i o logical anal2
ysis of m eth icillin2resistant S tap hy lococcus au reus in a Zagreb T raum a Ho sp ital using a random ly amp lified po lymo rph ic DNA2typ ing m ethod[J.Eur J C lin M icrobi o l Infect D is, 1999,18(5):3352340.
7 M c M anus A T,Goodw in CW,P ruitt BA J r,et al.O bserva2 ti ons on the risk of resistance w ith the extended use of van2 com ycin[J.A rch Surg,1998,133(11):120721211.
8 R eardon CM,B row n T PL aH,Stephenson A J,et al.M eth i2 cillin2resistant S tap hy lococcus au reus in burns patients2w hy all the fuss[J.Burns,1998,24(5):3932397.
9 府伟灵,肖光夏.抗内毒素类脂A单克隆和基因工程单链抗体的研究[J.第三军医大学学报,1994,16(5):3472348.
10 L o renz U,O h lsen K,Karch H,et al.H um an antibody re2 sponse during sep sis against targets exp ressed by m eth icillin
resistant S tap hy lococcus au reus[J.FE M S I mm uno lM ed M i2
crobi o l,2000,29(2):1452153.
122株甲型副伤寒沙门菌鉴定及药敏结果
俞连琴,骆 勇
(临安市人民医院,浙江临安311300)
关键词:甲型副伤寒沙门菌;生物学性状;耐药性
中图分类号:R378.2+2 文献标识码:B 文章编号:100524529(2003)0120006201
本地区发生一起甲型副伤寒沙门菌暴发流行。
现将我院来自发热患者的血、骨髓标本中分离的122株甲型副伤寒沙门菌的生物学性状及药敏结果分析如下。
1 材料与方法
菌株来源:122株菌株来源于本院门诊与住院患者的血、骨髓培养中分离鉴定所得。
仪器与方法:法国梅里埃A TB细菌鉴定仪及血液双向培养瓶。
2 结 果
122株甲型副伤寒沙门菌的生化反应结果较一致,除精氨酸双水解酶阳性率为89%、山梨醇阳性率为82%外,余均为100%。
抗生素敏感试验结果比较,见表1。
3 讨 论
3.1 本次流行的122株甲型副伤寒沙门菌生化反应和血清学特性基本一致,有100株生化谱为10560543000,与甲型副伤寒沙门菌生化反应完全一致。
推测本次暴发流行的甲型副伤寒沙门菌为单一流行株。
3.2 该菌株对大多数抗生素高度敏感,对喹诺酮二代培氟沙星耐药率3
4.4%,对第一代头孢菌素耐药率8%,对头孢
收稿日期:2002207224
表1 122株甲型副伤寒沙门菌药敏结果
抗生素试验株数敏感株数敏感率(%)
阿莫西林 64 64 100
哌拉西林122122100
替卡西林122122100
头孢噻吩645992
第三代头孢6464100
头孢曲松6464100
复达欣122122100
氨曲南12212098
亚胺培南 西司他丁12211796
复方新诺明122122100
妥布霉素122122100
阿米卡星122122100
庆大霉素122122100
奈替米星122122100
培氟沙星644266
环丙沙星12211796
多粘菌素E5858100
头孢磺吡苄5800
磷霉素5800
磺吡苄(第二代)、磷霉素耐药率100%,对第三代头孢菌素、三代喹诺酮类高度敏感。
・
6
・Ch in J N o socom i o l V o l.13N o.12003。