第三章内膜系统演示文稿
合集下载
内膜系统3作用机理PPT演示文稿

目录
返回
前进
退
4.4 .1. 溶酶体的形态结构
酸性水解酶是指在酸性条件下
具有活性的水解酶类。这些酶在酸 性溶液内都能分解重要的生物化合 物-------蛋白质、核酸、多糖及脂 类等,但在正常细胞内这些重要的 活性物质并不被这些酶所消化,而 且溶酶体自身的膜也不被消化,又 能保持溶酶体内的酸性环境,这与 溶酶体膜的结构和功能特征密切相 关。在溶酶体膜上具有质子泵 (proton pump),依赖ATP水解放出 能量将H+泵入溶酶体内维持其内 腔的酸性pH。构成溶酶体膜的蛋 白质是异乎寻常的高度糖基化。一 般认为这可保护膜免受溶酶体内蛋 白酶的消化。
14
目录
返回
前进
退
储积病是由于有关溶酶体酶缺乏,在患者的溶酶体内 累积了蛋白多糖(proteoglycan)所致,例如Hurler病。
Hurler病人肝组织 活检切片的肝细胞 电镜照片示溶酶体 (L)内充满了蛋白多 糖分子 X 5500
目录
返回
前进
退
在有的工矿企业中有一种职业病----矽肺(cilicosis), 其病因就与溶酶体有关。当肺部吸入矽尘后,矽粉末 (SiO2)被组织中的吞噬细胞吞噬。但是溶酶体酶不能破坏 矽粉末,而矽粉末能使溶酶体膜破裂,释放出其中的水解 酶,引起细胞死亡,放出的矽粉末,再被健康的吞噬细胞 吞噬可得到同样的结果。如此吞噬细胞相继地吞噬、死亡 ,最后刺激成纤维细胞导致胶原纤维结的沉积,结果是减 低了肺的弹性,损伤了肺的功能。已有实验证明,矽粉末 破坏溶酶体膜,是由于在矽颗粒表面形成矽酸,由酸的羟 基基团与溶酶体膜的受体分子之间形成氢键引起的。
目录
返回
前进
退
4.4 .1. 溶酶体的形态结构
内膜系统动画演示
↓
蛋白质合成完成,核糖体大小亚基分离,进入核糖体再循环
内质网、高尔基复合体与细胞的分泌活动 117分钟
3
H标记亮氨酸 17分钟
内质网、高尔基复合 体在细胞分泌活动中 起着重要的运输作用。
5分钟
分泌颗粒
转运小泡
内体
融合
糖基化 分选信号M6P 内体
运输 小泡
融合
脱Pi
大囊泡 膜厚:8nm; 泡内含物质:高电子密度物质,浓缩泡。 来 源:扁平囊周边或局部球状膨 突脱落形成。 扁平囊 呈盘状,3-10层称——————高尔基堆 扁平囊间距:20-30nm;囊腔宽:6-15nm 凸 面:形成(顺)面; 凹面:成熟(反)面 形成面膜厚: 6nm ; 成熟面膜厚:8nm 囊腔内含:中等电子密度的物质 来 源:小囊泡融合。 小囊泡 30-80nm球形小泡 膜厚:6nm; 泡内含物质:低电子密度物质,较透明。 来 源:由rER‘芽生’而来。 100-500nm;
SRP受体(SRP
receptor, SRPR):为ER膜上的跨膜蛋白,为异
二聚体蛋白,含有、两个亚基, 亚基含有SRP的结合部 位, 亚基以疏水区锚定在ER膜上。
信号肽 SRP 细胞质
ER腔
信号假说 mRNA AP 核糖体 信号肽
A
信号识别颗粒 (SRP)
tRNA
SRP受体 细胞质
membrane flow 跨膜流动
Heterophagy异体吞噬 Autophagy自噬 Autolysis自溶
Phagocytosal body
Phagosome
Myeline figure
Phagolysosome
Lipofusin
Lysosome
蛋白质合成完成,核糖体大小亚基分离,进入核糖体再循环
内质网、高尔基复合体与细胞的分泌活动 117分钟
3
H标记亮氨酸 17分钟
内质网、高尔基复合 体在细胞分泌活动中 起着重要的运输作用。
5分钟
分泌颗粒
转运小泡
内体
融合
糖基化 分选信号M6P 内体
运输 小泡
融合
脱Pi
大囊泡 膜厚:8nm; 泡内含物质:高电子密度物质,浓缩泡。 来 源:扁平囊周边或局部球状膨 突脱落形成。 扁平囊 呈盘状,3-10层称——————高尔基堆 扁平囊间距:20-30nm;囊腔宽:6-15nm 凸 面:形成(顺)面; 凹面:成熟(反)面 形成面膜厚: 6nm ; 成熟面膜厚:8nm 囊腔内含:中等电子密度的物质 来 源:小囊泡融合。 小囊泡 30-80nm球形小泡 膜厚:6nm; 泡内含物质:低电子密度物质,较透明。 来 源:由rER‘芽生’而来。 100-500nm;
SRP受体(SRP
receptor, SRPR):为ER膜上的跨膜蛋白,为异
二聚体蛋白,含有、两个亚基, 亚基含有SRP的结合部 位, 亚基以疏水区锚定在ER膜上。
信号肽 SRP 细胞质
ER腔
信号假说 mRNA AP 核糖体 信号肽
A
信号识别颗粒 (SRP)
tRNA
SRP受体 细胞质
membrane flow 跨膜流动
Heterophagy异体吞噬 Autophagy自噬 Autolysis自溶
Phagocytosal body
Phagosome
Myeline figure
Phagolysosome
Lipofusin
Lysosome
第三节细胞质内膜系统演示文稿

功 能——中间膜囊、成熟面含酶较多, 利于物质的浓缩和分泌。
当前第39页\共有63页\编于星期五\21点
当前第40页\共有63页\编于星期五\21点
二、高尔基复合体的化学组成
高尔基体膜:蛋白质 60%、脂类 40% ,有ER共同的蛋白成分。膜脂中磷脂酰胆碱含量介于ER和质膜之间。 高尔基体中的酶:主要有糖基转移酶、磺基-糖基转移酶、氧化还原酶、磷酸酶、蛋白激酶、甘露糖苷酶、转移酶和磷脂酶等不同的类型。 标志酶—糖基转移酶。
信号肽介导分泌蛋白在粗面内质网的合成
当前第24页\共有63页\编于星期五\21点
SRP
信号识别颗粒 (SRP) : signal recognition particle
存在于细胞质基质中,由6条多肽链和一个RNA分子组成。能够识别信号肽,与核糖体结合,形成SRP-核糖体复合体;复合体可以与粗面内质网上的SRP受体结合。
反面
分泌蛋白
粗面内质网
高尔基复合体
切除甘露糖
反面分选蛋白质示意图
膜蛋白
当前第51页\共有63页\编于星期五\21点
分选的结果
顺面分选:
反面分选:
驻留蛋白
溶酶体蛋白 分泌蛋白 膜蛋白
当前第52页\共有63页\编于星期五\21点
内质网
高尔基复合体
细胞膜
运输小泡
分泌泡
核膜
细胞核 内质网 高尔基复合体 细胞膜
当前第35页\共有63页\编于星期五\21点
扁平囊泡
小囊泡
大囊泡
顺面高尔基 网状结构
电镜下高尔基复合体横切面模式图
反面高尔基 网状结构
当前第36页\共有63页\编于星期五\21点
The Golgi Apparatus
当前第39页\共有63页\编于星期五\21点
当前第40页\共有63页\编于星期五\21点
二、高尔基复合体的化学组成
高尔基体膜:蛋白质 60%、脂类 40% ,有ER共同的蛋白成分。膜脂中磷脂酰胆碱含量介于ER和质膜之间。 高尔基体中的酶:主要有糖基转移酶、磺基-糖基转移酶、氧化还原酶、磷酸酶、蛋白激酶、甘露糖苷酶、转移酶和磷脂酶等不同的类型。 标志酶—糖基转移酶。
信号肽介导分泌蛋白在粗面内质网的合成
当前第24页\共有63页\编于星期五\21点
SRP
信号识别颗粒 (SRP) : signal recognition particle
存在于细胞质基质中,由6条多肽链和一个RNA分子组成。能够识别信号肽,与核糖体结合,形成SRP-核糖体复合体;复合体可以与粗面内质网上的SRP受体结合。
反面
分泌蛋白
粗面内质网
高尔基复合体
切除甘露糖
反面分选蛋白质示意图
膜蛋白
当前第51页\共有63页\编于星期五\21点
分选的结果
顺面分选:
反面分选:
驻留蛋白
溶酶体蛋白 分泌蛋白 膜蛋白
当前第52页\共有63页\编于星期五\21点
内质网
高尔基复合体
细胞膜
运输小泡
分泌泡
核膜
细胞核 内质网 高尔基复合体 细胞膜
当前第35页\共有63页\编于星期五\21点
扁平囊泡
小囊泡
大囊泡
顺面高尔基 网状结构
电镜下高尔基复合体横切面模式图
反面高尔基 网状结构
当前第36页\共有63页\编于星期五\21点
The Golgi Apparatus
《细胞的内膜系统》课件
内膜系统对细胞功能的影响
分子运输
内膜系统是细胞界面的重要组成 部分,可影响大部分分子在细胞 内和细胞外的转运和分布。
信号转导
内膜系统是细胞内外各种化学和 物理信号传递的关键路径,影响 着细胞各种功能的调节和整合。
能量代谢
内膜系统包括线粒体、内质网等 参与了膜上的氧化脱氧核苷酸反 应和电子传递,对细胞能量代谢 发挥不可替代的作用。
作用
线粒体参与ATP的合成、脂 质代谢、离子调节,以及凋 亡、离体器械复合、细胞老 化等诸多重要的仅在维 持细胞的代谢平衡,也对细 胞内各种不同功能的交集具 有重要的作用。
葡萄糖转运蛋白家族
1
分类
2
这个家族的小分子蛋白质一般下分A、B、
C、D、E、F、G、H8种亚家族,其中A、
溶酶体的组成和功能
组成
溶酶体是由高尔基体形成的膜结 构,内部充满多种水解酶。
功能
溶酶体主要负责内外源性膜蛋白 的降解、核苷酸、蛋白质等的水 解及一些小分子的储存等作用。
酸性环境
溶酶体内具有较低的pH值,成为 细胞唯一的酸性环境。
线粒体的结构和作用
结构
线粒体是细胞能量代谢最为 重要的场所之一,它具有双 层膜的结构,有内、外两个 膜。
内质网也承担一些重要的信号转 导功能。
高尔基体的构成和作用
1
构成
高尔基体通常由若干个扁平的、袋状的、同心排列的、但大小和形态不同的囊泡 组成,被称为高尔基小体。
2
作用
高尔基体不仅参与各种分子的加工、转运、储存等作用,同时也是重要的溶酶体 和内质网信号传递过程中的必要途径。
3
分类
根据不同的生理作用和形态,高尔基体可分为早期、中期和晚期高尔基体。
[课件]内膜系统-1PPT
![[课件]内膜系统-1PPT](https://img.taocdn.com/s3/m/5379ddfaad51f01dc281f146.png)
Endomembrane System
Endoplasmic
Golgi
reticulum 内质网
complex 高尔基复合体 溶酶体 过氧化物酶体
lysosome
peroxisome
Endoplasmic (ER)
内质网
内质网的形态结构和类型 内质网的功能
粗面内质网(RER)的功能 滑面内质网(SER)的功能
放射性示踪实验
3
H标记亮氨酸
117分钟
分 泌 蛋 白 运 输 途 径
4 分泌、释放于细胞外 3 转移到高尔基体
17分钟
2 进入rER
腔
3分钟
1 由rER上的核糖体合成
细胞核
高尔基体 含有粘液的囊泡
杯状细胞
三.高尔基复合体的功能
(一)参与细胞的分泌活动 (二)在分泌过程中的加工修饰作用
糖蛋白的合成和修饰 —— O-连接的糖基化
3.具有解毒作用(肝细胞) 4.参与骨骼肌的收缩
在肌细胞中肌质网(即SER)是Ca++贮存 场所,可通过释放和回收Ca++调节肌肉收
小结
1、内膜系统 2、内质网的形态特征 由一层单位膜围成的连续的网状膜系统 粗面内质网和滑面内质网 3、内质网的功能 RER:与蛋白质的合成、加工、运输有关 SER:多功能的细胞器 4、核糖体 游离核糖体 ——结合蛋白 膜结合核糖体 ——跨膜蛋白、分泌蛋白、 溶酶体酶蛋白、驻留蛋白
细胞质中核糖体在蛋白质合成启 动后, 根据mRNA上特定的信号 序列合成一段短肽,它可引导核糖 体结合到粗面内质网膜上 SRP receptor 通道蛋白移位子
rER function
1.蛋白质合成 信号肽假说的主要内容 指导因子
内膜系统Ippt-细胞生物学PPT课件

8
2020年10月2日 9
2020年10月2日 10
信号肽假说:Blobel&Dobbestein
2020年10月2日
在核糖体上合成信号肽
↓←SRP(在细胞质基质中)
SRP-核糖体复合体(蛋白质合成暂停)
↓←SRP受体(在rER上)
SRP受体-SRP-核糖体复合体
核糖体结合蛋白Ⅰ和Ⅱ(在rER)→↓→SRP 进入细胞质中再循环
核糖体结合蛋白Ⅰ和Ⅱ-核糖体(大亚基)复合体 (核糖体结合到rER)
↓
信号肽进入内质网腔,蛋白质合成继续
↓
蛋白质合成完成,核糖体大小亚基分离,进入核糖体再循环
11
2020年10月2日
N-连接的寡糖必需在ER膜上多萜醇(Dolichol)介导下才能转 移到蛋白质上。
12
2020年10月2日 13
膜
厚: 8nm;
泡内含物质:高电子密度物质,浓缩泡。
来
源:扁平囊周边或局部球
状膨突脱落形成。
小囊泡
直 径: 30-80nm 球形小泡
膜 厚: 6nm;
泡内含物质:低电子密度物质,较透明。
来
源:由rER‘芽生’而来。
凸面、顺面、未成熟面、形成面
26
扁平囊
2020年10月2日
凹 面:成熟(反)面
呈盘状,3-10层
• 高尔基体的发达程度与细 胞分化程度呈正相关。分 化好的细胞中,高尔基体 较发达;在未分化的细胞 中,高尔基体往往较同类 成熟型细胞少的多
24
一. 结构
光
网状、鳞片状结构
镜
大囊泡
电镜
扁平囊
小囊泡
25
2020年10月2日
2020年10月2日 9
2020年10月2日 10
信号肽假说:Blobel&Dobbestein
2020年10月2日
在核糖体上合成信号肽
↓←SRP(在细胞质基质中)
SRP-核糖体复合体(蛋白质合成暂停)
↓←SRP受体(在rER上)
SRP受体-SRP-核糖体复合体
核糖体结合蛋白Ⅰ和Ⅱ(在rER)→↓→SRP 进入细胞质中再循环
核糖体结合蛋白Ⅰ和Ⅱ-核糖体(大亚基)复合体 (核糖体结合到rER)
↓
信号肽进入内质网腔,蛋白质合成继续
↓
蛋白质合成完成,核糖体大小亚基分离,进入核糖体再循环
11
2020年10月2日
N-连接的寡糖必需在ER膜上多萜醇(Dolichol)介导下才能转 移到蛋白质上。
12
2020年10月2日 13
膜
厚: 8nm;
泡内含物质:高电子密度物质,浓缩泡。
来
源:扁平囊周边或局部球
状膨突脱落形成。
小囊泡
直 径: 30-80nm 球形小泡
膜 厚: 6nm;
泡内含物质:低电子密度物质,较透明。
来
源:由rER‘芽生’而来。
凸面、顺面、未成熟面、形成面
26
扁平囊
2020年10月2日
凹 面:成熟(反)面
呈盘状,3-10层
• 高尔基体的发达程度与细 胞分化程度呈正相关。分 化好的细胞中,高尔基体 较发达;在未分化的细胞 中,高尔基体往往较同类 成熟型细胞少的多
24
一. 结构
光
网状、鳞片状结构
镜
大囊泡
电镜
扁平囊
小囊泡
25
2020年10月2日
内膜系统-幻灯片(1)

研究发现:TGN在蛋白质与脂类的转运过程中还起到 “瓣膜”的作用,保证这些物质单向转运。
(二) GC的化学组成
大鼠肝细胞GC含有60%的蛋白质和40%的脂类。
1. 脂类成分:介于膜和ER膜之间
RER含有大量的不饱和磷脂,而胆固醇含量很低;细胞膜 中含有丰富的饱和磷脂和胆固醇;GC膜则介于两者之间。
又称为颗粒内质网(Granular Endoplasmic Reticulum GER) 光面内质网(Smooth Endoplsmic Reticulum, SER)
又称为无颗粒内质网(Agranular Endoplasmic
Reticulum AER)
微粒体(microsome):应用蔗糖密度梯度离心法将 ERE和SRE分离开来,离心后ER断裂成的许多,直 径在100nm的小泡。表面有核糖体的,来源于RER, 无则来源于SER。
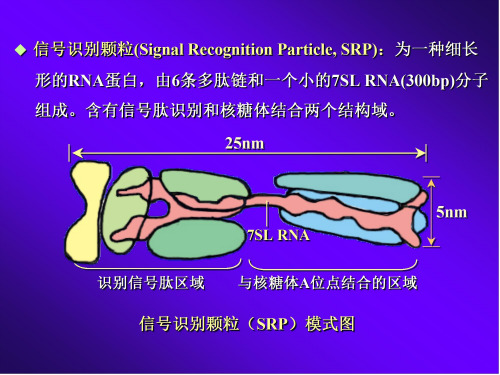
25nm
7SL RNA
5nm
识别信号肽区域 与核糖体A位点结合的区域
信号识别颗粒(SRP)模式图
SRP受体(SRP receptor, SRPR):为ER膜上的跨膜蛋白, 含有、两个亚基。 亚基含有SRP的结合部位, 亚 基以疏水区锚定在ER膜上。
信号肽
SRP
细胞质
ER腔
跨膜通道(Transmembrane channel):又称为蛋白转移器 (Translocon),使新生的肽链通过ER膜进入到ER腔中。
基
化 O-连接的糖蛋白:是蛋白质的Tyr、Ser和Thr残基侧链的
作 用
-OH与寡糖链共价相连。其糖基化过程全部或主要在高 尔基复合体内完成。
ER腔中N-连接的糖蛋白的形成
寡糖的组成:2个N-乙酰氨基葡萄糖 (N-Acetylglucosamine,
(二) GC的化学组成
大鼠肝细胞GC含有60%的蛋白质和40%的脂类。
1. 脂类成分:介于膜和ER膜之间
RER含有大量的不饱和磷脂,而胆固醇含量很低;细胞膜 中含有丰富的饱和磷脂和胆固醇;GC膜则介于两者之间。
又称为颗粒内质网(Granular Endoplasmic Reticulum GER) 光面内质网(Smooth Endoplsmic Reticulum, SER)
又称为无颗粒内质网(Agranular Endoplasmic
Reticulum AER)
微粒体(microsome):应用蔗糖密度梯度离心法将 ERE和SRE分离开来,离心后ER断裂成的许多,直 径在100nm的小泡。表面有核糖体的,来源于RER, 无则来源于SER。
25nm
7SL RNA
5nm
识别信号肽区域 与核糖体A位点结合的区域
信号识别颗粒(SRP)模式图
SRP受体(SRP receptor, SRPR):为ER膜上的跨膜蛋白, 含有、两个亚基。 亚基含有SRP的结合部位, 亚 基以疏水区锚定在ER膜上。
信号肽
SRP
细胞质
ER腔
跨膜通道(Transmembrane channel):又称为蛋白转移器 (Translocon),使新生的肽链通过ER膜进入到ER腔中。
基
化 O-连接的糖蛋白:是蛋白质的Tyr、Ser和Thr残基侧链的
作 用
-OH与寡糖链共价相连。其糖基化过程全部或主要在高 尔基复合体内完成。
ER腔中N-连接的糖蛋白的形成
寡糖的组成:2个N-乙酰氨基葡萄糖 (N-Acetylglucosamine,
第三章细胞的内膜系统

1972年,stein等在研究骨髓瘤细胞中 的发现。
1975年,G.Blobel和D.sabatini提出了信号假 说。
• 1999年,G.Blobel获得了诺贝尔医学和生理 学奖。 现已确定:蛋白质N端的信号序列,控制蛋白质
在细胞内的转移与定位,是蛋白质在粗面型 内质网上合成的决定因素。
第四节、溶酶体(Lysosome)
一、溶酶体的形态特 征:溶酶体为直径很 小的囊状小体,其外 包以一层厚约6nm 的单位膜,膜内含有 活性很高的酸性水解 酶60多种。
溶酶体的类型
内体性溶酶体: 由高尔基复合体成熟面的扁平囊 形成有被小泡,脱落后再与内体 合并即为内体性溶酶体。
Two possible models explaining the organization of the Golgi complex and the transport from one cisterna to the next.
高尔基复合体与疾病
高尔基体肥大 高尔基体萎缩、消失 高尔基体与癌症
第一节 核糖体
早在40年代初,就有人发现细胞内有一种颗粒状 结构与蛋白质合成有关,1955年,Palade研究 了这种颗粒的性质。故历史上曾有所谓“Palade 颗粒”之称。直到1958年,Roberts才根据这种 颗粒组成成分的性质,提出了核糖核蛋白体的概 念,简称核糖体或核蛋白体(ribosome),是细 胞内蛋白质合成的“工作台”。
1945年Porter K.R和 Claude A.D用电子显 微镜观察培养的小鼠 成纤维细胞发现细胞 质中有一些形状大小 略有不同的小管、小 囊连接成网状的结构 称内质网。
内质网的形态结构
三种结构单位:扁平囊、小管和小泡
1、扁平囊排列: 内质网膜之间为狭窄的腔, 形状扁而长,不封闭, 两膜间宽度40-50nm, 常常很多重叠在一起。 在纵切面上为一个个两端 封闭的扁长管道, 平行状排列并相互连通。
1975年,G.Blobel和D.sabatini提出了信号假 说。
• 1999年,G.Blobel获得了诺贝尔医学和生理 学奖。 现已确定:蛋白质N端的信号序列,控制蛋白质
在细胞内的转移与定位,是蛋白质在粗面型 内质网上合成的决定因素。
第四节、溶酶体(Lysosome)
一、溶酶体的形态特 征:溶酶体为直径很 小的囊状小体,其外 包以一层厚约6nm 的单位膜,膜内含有 活性很高的酸性水解 酶60多种。
溶酶体的类型
内体性溶酶体: 由高尔基复合体成熟面的扁平囊 形成有被小泡,脱落后再与内体 合并即为内体性溶酶体。
Two possible models explaining the organization of the Golgi complex and the transport from one cisterna to the next.
高尔基复合体与疾病
高尔基体肥大 高尔基体萎缩、消失 高尔基体与癌症
第一节 核糖体
早在40年代初,就有人发现细胞内有一种颗粒状 结构与蛋白质合成有关,1955年,Palade研究 了这种颗粒的性质。故历史上曾有所谓“Palade 颗粒”之称。直到1958年,Roberts才根据这种 颗粒组成成分的性质,提出了核糖核蛋白体的概 念,简称核糖体或核蛋白体(ribosome),是细 胞内蛋白质合成的“工作台”。
1945年Porter K.R和 Claude A.D用电子显 微镜观察培养的小鼠 成纤维细胞发现细胞 质中有一些形状大小 略有不同的小管、小 囊连接成网状的结构 称内质网。
内质网的形态结构
三种结构单位:扁平囊、小管和小泡
1、扁平囊排列: 内质网膜之间为狭窄的腔, 形状扁而长,不封闭, 两膜间宽度40-50nm, 常常很多重叠在一起。 在纵切面上为一个个两端 封闭的扁长管道, 平行状排列并相互连通。
相关主题
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
核糖体上进行分泌蛋白质合成的信号肽假说
核糖体的存在形式
①附着核糖体:主要合成分泌性蛋白质 ②游离核糖体:主要合成结构性蛋白质
③功能状态都为多聚核糖体,非功能状态为 大、小亚基分开
2.粗面内质网与蛋白质的糖基化
蛋白质的糖基化:是指单糖或寡糖与蛋白 质共价结合形成糖蛋白。
蛋白质糖基化的作用
使蛋白质能够抵抗消化酶的作用 赋予蛋白质传导信号的功能 某些蛋白只有在糖基化之后才能正确折叠 在细胞表面形成糖萼
蛋白质糖基化作用方式 N-连接的糖基化:发生在内质网腔 内,糖蛋白中最普遍的一种。 O-连接的糖基化:主要或全部在高 尔基复合体中进行。
N-连接的糖基化
在粗面内质网中,糖链被连接在多肽链中天冬酰 胺残基(Asn)的氨基基团上,故称之为N-连接糖基化。
在内质网腔中N-乙酰葡萄糖胺、甘露糖和葡萄糖 的多个分子按顺序先后被连接到内质网膜中叫多帖醇 的脂质分子上,成为寡聚糖,并使寡聚糖活化。
浓缩泡→分泌颗粒→ 排出; • 膜嵌入蛋白质,即膜蛋白; • 需要与其他细胞组分严格隔离的蛋白质; • 需要进行复杂修饰的蛋白质;
4.脂类的合成
• 合成大部分细胞膜和细胞内膜系统的膜脂 • 合成脂类的酶都位于内质网脂质双层,活性部
位朝向胞质面 • 翻转酶 • 磷脂酰胆碱的生物合成过程:酰基转移酶、磷
脂酸酶、胆碱磷酸转移酶
(二)滑面内质网的功能
1.脂类和类固醇激素的合成 2.脂类代谢 3.糖原代谢 4.解毒作用 5.其它作用
五、内质网与疾病
脂类的合成
• 肾上腺皮质细胞、睾丸间质细胞、卵巢共黄 体细胞等类固醇激素细胞
• SER含有合成胆固醇全套酶系和使胆固醇转 化为类固醇激素(肾上腺激素、性激素)的酶 类及与脂类合成有关的酶类
内质网(膜厚5~6nm)是由一层单位膜围 成的形状大小不同的小管,小泡,扁囊状 结构,相互连接形成一个连续的网状膜系 统。
内质网的分布状态和数量多少与细胞的类 型有关。在同种细胞的不同部位、不同发 育时期,甚至随着生理功能的不同,内质 网也有差异。
(一)粗面内质网(RER)
粗面内质网(rough ER,RER):膜表面附着核糖 体;形态多为扁囊状。 功能:蛋白质的合成与转运。
已活化的寡聚糖即由糖基转移酶催化。
O-连接的寡糖的糖基化
• O-连接的寡糖是指与蛋白质的丝氨酸、苏 氨酸和酪氨酸残基侧链上的羟基基团连接 的寡糖。
• O-连接的寡糖的糖链中又增加了半乳糖、 岩藻糖和唾液酸等糖残基。
3.粗面内质网与蛋白质的运输
• 向细胞外分泌的蛋白质: RER合成→糖基化作用→小泡(含分泌蛋白)→经GC→
(二)滑面内质网(SER)
滑面内质网(smooth ER,SER):膜表面无核糖 体附着;形态多为分枝小管或小泡;多分布在 一些特化的细胞中。 功能:脂类合成的场所;解毒作用;物质运输 和交换等。
ห้องสมุดไป่ตู้
电镜下显示内质网(兰色),核糖体颗粒(绿色)
类
电镜下显示内质网腔和表面附着核糖体
结类
类
三、内质网的化学组成
光面内质网在糖原分解中的作用
SER的解毒作用
• 肝细胞的解毒作用------SER通过氧化、甲基化、结 合等方式,降低或排除毒性物质
• 药物、毒物─→(SER膜上的氧化酶系)→解毒或转化 如:氨基酸代谢→氨→尿素(无毒); 苯巴比妥类药物+葡萄糖醛酸→水溶性物质
盐酸的分泌、渗透压调节
哺乳动物胃底臂细胞的胞质中,SER 能将血浆中的CI-传递到细胞内分泌小管 的膜上, CI-可与胞质中由碳酸解离的H+ 在膜上结合产生HCI,排出细胞外。
微粒体:用蔗糖密度梯度离心方法,从细胞 匀浆中分离出的内质网 碎片。是研究ER化 学组成和功能的好材料。
内质网膜有脂类(30 % -40%)和蛋白质 (60 % -70 % )组成。
• 葡萄糖-6-磷酸酶(G-6-P酶)被视 为内质网膜的标志酶。
• 细胞色素P-450主要位于滑面内质网 的标志酶。
• 信号识别颗粒(signal recognition particle,SRP),它是 由6条不同的肽链结合在1分子的7SrRNA上组成的核 糖体蛋白,它能特异识别结合信号肽。SRP占据了 核糖体的A位,阻挡了携带氨基酸的tRNA进入核糖 体,使蛋白质的合成暂时中止。
• SRP受体(SRP receptor),它是结合在内质网膜上的镶 嵌蛋白,分子量约72000,它可以识别结合SRP。
四、内质网的功能
(一)粗面内质网的功能
1.粗面内质网与蛋白质合成 2.粗面内质网与蛋白质的糖基化 3.粗面内质网与蛋白质的运输 4.粗面内质网与膜脂的合成
1.粗面内质网与蛋白质合成 信号假说
• 信号密码:位于起始密码(AUG)之后,有一组编码 特殊氨基酸序列的密码子。
• 信号肽:蛋白质合成时,首先在游离核糖体上由 信号密码翻译出一段肽链,这一小段额外的肽链 为疏水的氨基酸序列,约有18~30个氨基酸组成。 信号肽是个信号,是附着核糖体的标记。
第三章内膜系统演示文稿
5内-6周质雄网性昆明小鼠 高尔基复合体 溶酶体 人过肾氧小化管上物皮酶细体胞HK-2
第一节 内质网
一、内质网(ER)的来源
1945年,K.R.Poter----电子显微镜-----小鼠 成纤维细胞
一般认为,在内膜系统中,内质网处于中心 位置,为内膜系统的发源地。
二、内质网的类型和形态结构
第二节 高尔基复合体
1898 意大利---高尔基--光镜---猫头鹰、猫--神经细胞
一、高尔基复合体的形态结构 成熟面
反面高 尔基网
电 高尔基中 镜 间膜囊
• 信号肽酶:水解信号肽的酶。
信号假说
信号识别颗粒模式图
附着核糖体与蛋白质的合成过程: 1.游离核糖体上信号肽合成; 2.胞质中SRP识别信号肽,形成SRP-核糖
体复合体,从而使多肽链合成暂停。 3.核糖体与粗面内质网结合,形成SRP-
SRP受体-核糖体复合体;
4. SRP脱离并参加再循环,核糖体上的多肽 链合成继续进行。 5.信号肽被ER膜上的信号肽酶切掉,合成后 的多肽链落入内质网腔中; 6.附着核糖体脱离内质网膜,大小亚基分开, 回到胞质基质中参加再循环,内质网膜上的 受体小管消失 。