4遗传模型
高考生物考前梳理必修2遗传与进化实验4制作DNA双螺旋结构模型(含答案)

高考生物考前梳理:
4.制作DNA双螺旋结构模型
目的要求
通过制作DNA双螺旋结构模型,加深对DNA结构特点的认识和理解。
材料用具
曲别针、泡沫塑料、纸片、扭扭棒、牙签、橡皮泥、铁丝等常用物品,都可用作模型制作的材料。
模型设计
制作模型前首先应该进行设计,并考虑以下问题。
1.分别用哪几种材料来代表组成DNA的磷酸、脱氧核糖和碱基?这3种物质是在什么部位相互连接的?怎样将这几种材料正确地连接起来?
2.DNA中每个脱氧核苷酸之间是在什么部位相互连接的?怎样将脱氧核苷酸正确
地连接起来?
3.在模型中,如何体现DNA的两条链反向平行的?又怎样体现两条链的碱基之间互补配对?讨论
1.DNA只含有4种脱氧核苷酸,它为什么能够储存足够量的遗传信息?
提示:DNA虽然只含有4种脱氧核苷酸,但是碱基的排列顺序却是千变万化的。
碱基排列顺序的千变万化,使 DNA 储存了大量的遗传信息。
1.DNA是如何维系它的遗传稳定性的?
提示:(1)靠DNA碱基对之间的氢键维系两条链的偶联;(2)在DNA双螺旋结构中,由于碱基对平面之间相互靠近,形成了与破基对平面垂直方向的相互作用力(该点可不作为对学生的要求,教师可进行补充说明)。
2.你能够根据DNA的结构特点,设想DNA的复制方式吗?
提示:不一定要求学生答出半保留复制,可以引导学生从 DNA 的结构来进行推测,如从碱基互补配对原则出发去思考。
生物系统的数学模型

生物系统的数学模型引言生物系统是由生物体组成的复杂系统,其中包括生物体的结构、功能和相互作用。
为了更好地理解和解释生物系统的行为,科学家们使用数学模型来描述和研究这些系统。
生物系统的数学模型是一种用数学语言描述生物系统的方式,通过建立数学方程和模拟方法,可以预测和探索生物系统的行为和特性。
1. 动力系统模型动力系统模型是生物系统中常用的一种数学模型。
它描述了生物体在时间上随着不同因素的变化而变化的过程。
例如,人体的代谢过程可以用动力系统模型来描述。
该模型将人体的代谢活动分解为一系列的化学反应,并使用微分方程来描述反应速率和物质浓度的变化。
通过求解这些微分方程,可以预测不同条件下人体代谢的动态变化。
2. 群体动力学模型群体动力学模型是用来描述群体中个体之间相互作用的数学模型。
生物系统中的许多行为和特性是由群体中个体之间的相互作用所决定的,例如群体中的迁徙、繁殖和竞争等。
群体动力学模型使用微分方程或差分方程来描述个体之间的相互作用和群体的动态变化。
通过改变模型的参数和初始条件,可以预测群体行为的变化和演化。
3. 网络模型网络模型是一种描述生物系统中各个组成部分之间相互连接关系的数学模型。
生物系统中的许多结构和功能是通过各个组成部分之间的网络连接实现的,例如脑部神经元之间的连接和代谢途径之间的关联。
网络模型使用图论和复杂网络理论来描述和分析这些连接关系。
通过分析网络的拓扑结构和动态特性,可以揭示生物系统中的关键组成部分和功能模块,并预测系统的稳定性和鲁棒性。
4. 遗传算法模型遗传算法模型是一种基于遗传和进化原理的数学模型,用于解决生物系统中的优化和适应性问题。
生物系统中的许多特性和行为是通过进化过程中的适应性选择所形成的,例如生物体的形态和行为特征。
遗传算法模型使用遗传算法的原理来模拟和优化生物系统的演化过程。
通过不断迭代和优化,可以找到生物系统中的最优解或接近最优解的解决方案。
结论生物系统的数学模型是一种用数学语言描述和解释生物系统行为的方法。
遗传病发病率的数学模型高考生物计算题真题解析

遗传病发病率的数学模型高考生物计算题真题解析在生物学的学习中,数学模型的应用是必不可少的一部分。
遗传病发病率的数学模型是其中的一种重要的应用。
在高考生物考试中,经常会出现与遗传病发病率相关的计算题。
本文将对遗传病发病率的数学模型进行解析。
一、遗传病发病率的定义遗传病发病率是指在一定种群中,某种遗传病在一定时间内新发病的率。
通常以千分之几或者百分之几来度量。
二、遗传病发病率的数学模型在遗传病发病率的计算中,常用的数学模型有“剧毒遗传树”的模型、受体率模型和费歇卡模型等。
1. “剧毒遗传树”模型这个模型用于计算某种遗传病在家族中新发病的概率。
根据遗传病的类型,可以绘制出遗传病在家族中的传递路径,即“剧毒遗传树”。
通过剧毒遗传树,可以计算某一代或多代的发病率。
2. 受体率模型受体率模型适用于计算某种常染色体显性疾病的发病率。
该模型根据遗传病基因的传递方式,计算家族中某一世代的发病率。
3. 费歇卡模型费歇卡模型适用于计算某种常染色体隐性疾病的发病率。
该模型根据遗传病基因的传递方式,计算家族中某一世代的发病率。
三、数学模型的应用举例以下是一个实际的应用题,来看一下如何使用数学模型计算遗传病发病率。
某地需要对一种遗传病进行筛查,该遗传病为常染色体隐性疾病,发病率为1%。
假设筛查区域的人口为10万人,请计算该地区有该遗传病的携带者人数和患病儿童的数目。
解答:根据费歇卡模型,假设该地区该遗传病的携带者频率为p,正常基因的频率为q,则有p+q=1。
由题意可知,发病率为1%,即患病个体的频率为0.01,而携带者个体的频率为2pq。
根据人口数量,我们可以设定有N个个体,则正常的基因型频率为q^2,即q^2=N*q,从而可以得到q=N/(N+1)。
而遗传病的携带者频率为2pq,即2pq=N(N+1)。
根据题意可得:2pq=0.01N(N+1)2q(N-(N+1))=0.01N(N+1)q(N+1-N)=0.005(N^2+N)q=0.005(N+1)代入已知的人口数量N=10万,可得到q=0.005*1.01=0.00505。
生产计划与调度优化模型研究

生产计划与调度优化模型研究随着工业化和市场化的发展,生产计划与调度越来越重要。
优化生产计划与调度模型的研究,不仅可以提高企业的生产效率,还可以降低生产成本,提高资源利用率。
本文将会重点介绍生产计划与调度优化模型的研究,包括模型的分类、应用场景以及最新发展趋势。
一、生产计划与调度优化模型的分类生产计划与调度模型可以分为几种:线性规划模型、动态规划模型、贪心算法模型、遗传算法模型等。
1.线性规划模型线性规划模型是指在线性条件下求解最优解的数学模型。
它可以用来解决一般的生产计划和调度问题,包括生产计划、物料订购、生产维修、员工排班、车辆调度等。
利用线性规划模型,可以使各种资源的使用达到最优化,实现最佳效益。
2.动态规划模型动态规划模型是一种优化问题的数学模型。
它是以最优解为目标,采用分步决策方式的算法,逐步解决问题。
这种模型适合于解决一些具有复杂性和不确定性的问题,例如库存控制、作业调度、排队论以及飞行管制等。
3.贪心算法模型贪心算法是指在每个阶段都能选择最优决策,从而达到全局最优状态的算法模型。
贪心算法具有一般性,适用于多种生产计划与调度问题。
贪心算法适合解决一些简单的问题,但对于那些复杂的问题,贪心算法得到的结果可能不是最优的。
4.遗传算法模型遗传算法是一种模仿自然界进化过程而发展起来的优化算法。
遗传算法模型擅长解决大规模复杂问题,例如车辆路径规划、员工排班等。
遗传算法模型通过数值计算和成本分析,可以找到最优的生产计划和调度方案。
二、生产计划与调度优化模型的应用场景生产计划与调度模型的应用场景非常广泛,这里只列举了一部分。
1.生产计划生产计划是制定生产过程的最初阶段。
在生产计划阶段,生产部门会预测销售量、确定生产资源和机器设备合理配置等。
这些预测和决策需要采用适当的数学模型和方法进行分析和解决。
2.车辆调度车辆调度是企业中非常重要的一个工作。
在车辆调度过程中,需要考虑路线选择、货物装载、车辆配备等多个方面的因素。
数学建模之遗传模型

.
X—链遗传模型
• (iv)如果第n-1代配偶的同胞对是(A, Aa)型,那么它们的雄性后代将等可能 地得到基因A或a,它们的雌性后代的基 因型将等可能地是AA或A。又由于第n 代雌雄结合是随机的,那么第n代配偶 的同胞对将等可能地为四种类型 (A, AA) , (A , Aa) , , (a , AA) , (a,Aa) 之 一。其他类似可分析
.
常染色体隐性病模型
• 假若我们能识别这些疾病的隐性患者, 并且规定隐性患者不能结合(因为两个隐 性病患者结合,他们的后代就可能成为 显性患者),那么未来的儿童,虽然有可 能是隐性患者,但绝不会出现显性特征, 不会受到疾病的折磨。现在,我们考虑 在控制结合的情况下,如何确定后代中 隐性患者的概率。
X—链遗传模型
父体一母体的基因型
(A,AA) (A,Aa) (A,aa) (a,AA) (a,Aa) (a,aa)
A
1
1/2
0
1
1/2
0
a
0
1/2
1
0
1/2
1
后
代
基 AA
1
1/2
0
0
0
0
因
型
Aa
0
1/2
1
1
1/2
0
aa
0
0
0
0
1/2
1
.
X—链遗传模型
• (iii)在每一代中,配偶的同胞对也是六 种类型之一,并有确定的概率。为计算 这些概率,设an,bn,cn,dn,en和fn分 别是第n代中,配偶的同胞对为(A,AA), (A,Aa),(A,aa),(a,AA),(a,Aa), (a,aa)型的概率,n=0,1,…。令
利用重组自交系(RILs)群体进行质量数量性状的遗传分析遗传模型和小麦产量性状遗传

照互补、抑制、重叠、积加、显性上位、隐性上位及协调作用等控制质量性状基因的互作方
式, 在微效基因背景下进行了建模, 探讨了两个位点主基因控制的质量 数量性状的遗传
模型和分析方法。但作为控制质量 数量性状的主基因, 这些作用方式只是一些特例。以
上模型大多是对分离世代个体 (或株系) 进行分析, 会受环境影响较大而导致不易准确鉴定其
R IL s 群体对由 1 对或 2 对主基因和微效基因共同控制的质量 析方法进行探讨。并采用该方法对小麦产量性状进行分析。
数量性状的遗传模型和分
1 遗传模型
根据遗传学原理, 主基因效应较大, 控制的性状呈现明显的不连续分布; 由大量的微效 基因控制的性状呈正态分布。某一主基因型在微效基因背景下, 呈现以该主基因型值为均数 的正态分布。同一群体若有多个不同的主基因型值, 微效基因对每个主基因型值的影响是相 同 的, 因此多个正态分布的方差 (Ρ2) 相同, 该群体的分布则是各个正态分布的联合分布。 R IL s 群体是由一系列相对纯合的系组成, 从理论上讲, 若某个 R IL s 群体涉及 1 对主基因 A —a (A , a 分别表示增减效) 差异, 则 R IL s 群体中有 2 种不同的主基因型: AA、aa, 各占 1 2; 若涉及 2 对主基因A —a、B —b 差异, 则有 4 种不同的主基因型: AABB、AA bb、aaBB、
经典观点认为生物的性状分为质量性状和数量性状, 质量性状受少数主基因控制, 而数
ቤተ መጻሕፍቲ ባይዱ
量性状受大量微效基因控制。事实上, 作物的许多经济性状既有可分组的趋势, 又呈现连续 变异, 同时表现出质量性状和数量性状两者的特征, 莫惠栋称之为质量 数量性状 (Q Q 性状) [1, 2], 这类性状是由主基因 多基因混合遗传体系[3]控制的。有些性状被认为属数 量性状的范畴, 越来越多的Q TL s (guan tita tive tra it lines) 分子标记定位结果表明其存在作用 较大的主基因[4~ 11]; 而有些被认为属质量性状的范畴, 实际上也表现出连续变异的特征, 又 有微效基因的作用。质量 数量性状是较广泛地存在的。不少学者[1~ 3, 12~ 19 ]对质量 数
常见的炎症模型的造模方法有哪些

常见的炎症模型的造模方法有哪些炎症是人体对于外部刺激或内部损伤的一种自我保护反应,通常会伴随一系列的症状,例如红肿、疼痛、发热等。
炎症模型是用来研究这个生理过程的工具,可以有助于我们理解炎症的机制和寻找治疗炎症相关疾病的方法。
下面将介绍一些常见的炎症模型的造模方法。
1. 细菌或病毒感染模型:这是最常见的炎症模型之一。
通过给动物注射或感染细菌、病毒或其他微生物,可以模拟真实感染情况下的炎症反应。
例如,在小鼠中注射细菌,可以观察到炎性细胞浸润、组织纤维化等反应,并通过分析炎症介质的表达来进一步研究炎症的机制。
2. 激素介导的炎症模型:某些激素(如肾上腺皮质激素)可以诱导炎症反应。
通过给动物注射这些激素,可以在短时间内引发炎症反应,研究其发生机制和治疗方法。
例如,给小鼠注射肾上腺素可以引发全身性炎症反应,并观察其炎症介质的变化和症状的表现。
3. 实体和组织损伤模型:通过物理或化学方法对动物的组织或器官进行损伤,可以模拟外伤引发的炎症反应。
例如,通过切割或压迫动物的皮肤或组织,可以引起局部炎症反应,观察炎性细胞的浸润和炎症介质的表达。
4. 致敏模型:在动物中引入过敏原,例如在小鼠中注射抗原,可以引发过敏反应并诱导炎症反应。
通过分析过敏反应的机制和研究炎症介质的变化,可以帮助我们了解过敏和炎症之间的关系。
5. 自身免疫模型:自身免疫性疾病与炎症有密切关系。
通过给动物注射自身抗原,例如在小鼠中注射胸腺提取物,可以触发自身免疫反应并引起炎症。
通过研究这些模型,可以深入了解自身免疫疾病的发生机制和炎症调节的方法。
6. 遗传模型:研究表明,一些基因的突变或缺陷与炎症相关的疾病有关。
通过培育具有特定基因突变的动物模型,可以模拟这些疾病,并研究其炎症反应的变化,以及借此寻找治疗方法。
7. 让步模型:炎症调控的研究中,为了便于实验操作,常常使用让步模型。
通过给动物注射低剂量的炎症介质,例如注射低剂量的细菌成分(如脂多糖),来模拟炎症反应,并研究其机制。
数量遗传学4-交配设计

第四章交配设计
第一节北卡诺林那设计I(NCI)
•交配方案:在一个群体中,选择n1个父本,每个父本与n2个不同的母本交配,产生n
1
×n2个全同胞家系,每家系含r个个
体,共有n
1
个半同胞家系群。
123
123456789
1 2 3 41
2
3
4
1
2
3
4
1
2
3
4
1
2
3
4
1
2
3
4
1
2
3
4
1
2
3
4
1
2
3
4
父本母本全同胞家系
第二节北卡诺林那设计II (CNII )
交配方案:n 1父本与n 2母本完全交配,共n 1×n 2个全同胞家
系,每系r 个个体。
1
2
3
1
231234
1234
1234
1234
1234
1234
1234
1234
1234
父本母本全同胞家系
123123
CNIII 优点:
•通过MS S 和MS TS 分别估计出V A 和V D ,以相同精度独立地测验和估计V A 和V D
•可以统计测验超显性:F = MS TS / MS S
第五节双列杂交
•交配方案:任一群体中,选一组(n1个)父本和一组(n2个)母本相互交配。
当父、母本不同时,相当于NCII。
–完全双列杂交:父母本相同,后代包括自交。
–不完全双列杂交:父母本不同(NCII)。
•方差分析及组成(略)。
回交自交系(BIL)群体4对主基因加多基因混合遗传模型分离分析方法的建立

作物学报ACTA AGRONOMICA SINICA 2013, 39(2): 198−206 /zwxb/ ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@DOI: 10.3724/SP.J.1006.2013.00198回交自交系(BIL)群体4对主基因加多基因混合遗传模型分离分析方法的建立王金社赵团结盖钧镒*南京农业大学大豆研究所 / 国家大豆改良中心 / 农业部大豆生物学与遗传育种重点实验室(综合) / 作物遗传与种质创新国家重点实验室, 江苏南京 210095摘要: 主基因加多基因混合遗传模型是用于分析数量性状表型数据的统计分析方法, 该方法便于育种工作者利用杂种分离世代的数据对育种性状的遗传组成初步判断,制定相应的育种策略,也可用于校验QTL定位所揭示的数量性状的性状遗传组成。
回交自交系(BIL)群体是永久性群体, 可以进行有重复的比较试验, 适用于受环境影响较大的复杂性状的遗传研究。
本研究以BIL群体为对象构建了4对主基因、主基因加多基因分离分析方法的遗传模型, 包括2类11个遗传模型。
利用基于IECM (iterative expectation conditional maximization)算法的极大似然分析方法估算各个混合遗传模型中的分布参数, 用AIC值和一组适合性测验结果选取最优模型, 并从入选模型的分布参数通过最小二乘法估计遗传参数。
由1个模拟的随机区组试验对模型进行验证, 模拟群体中遗传参数的估计值与设定值之间具有很好的一致性。
利用本文建立的模型重新分析大豆回交自交系群体(Essex×ZDD2315)及其亲本对胞囊线虫(Hetero-dera glycines Ichinohe) 1号生理小种的抗性数据后发现4对主基因模型优于原报道的3对主基因模型, 说明本方法的有效性和正确性。
关键词:回交自交家系群体(BIL); 主基因加多基因混合遗传; 分离分析Establishment of Segregation Analysis of Mixed Inheritance Model with Four Major Genes Plus Polygenes in Backcross Inbred Lines (BIL) Populations WANG Jin-She, ZHAO Tuan-Jie, and GAI Jun-Yi*Soybean Research Institute of Nanjing Agricultural University / National Center for Soybean Improvement / Key Laboratory for Biology and Genetic Improvement of Soybean (General), Minister of Agriculture / National Key Laboratory for Crop Genetics and Germplasm Enhancement, Nanjing 210095, ChinaAbstract: The segregation analysis of major genes plus polygenes is a statistical method for genetic analysis of quantitative traits. The method is particularly valuable for plant breeders to use their data accumulated from segregation populations to estimate the genetic system of target traits, which is necessary for designing breeding strategies and also useful for validating the results of QTL mapping. The backcross inbred line (BIL) population is one of the permanent populations, which is suitable for genetic analysis of complex traits and can be used in replicated experiments. For BIL population, the analytical procedures of three and less major genes plus polygenes mixed inheritance models have been established. The objective of the present study was to estab-lish the analytical procedures of segregation analysis for four major genes plus polygenes mixed inheritance models in BIL popu-lation. Eleven genetic models with four additive and (or) epistatic major genes including those without and with polygenes were established. The component distribution parameters were solved and estimated by using maximum likelihood method based on IECM (Iterative Expectation Conditional Maximization) algorithm. Among the possible models, the best one was chosen accord-ing to Akaike’s Information Criterion (AIC) and a set of tests for goodness of fit. Then the genetic parameters of the optimal model were estimated through the least square method. For demonstration of the established procedures, a simulated data set of a本研究由国家重点基础研究发展计划(973计划)项目(2009CB1184, 2010CB1259, 2011CB1093), 国家高新技术研究发展计划(863计划)项目(2011AA10A105, 2012AA101106), 国家自然科学基金资助项目(31071442, 32671266), 农业部公益性行业专项(200803060), 江苏省优势学科建设工程专项和国家重点实验室自主课题项目资助。
遗传算法模型公式

遗传算法模型公式遗传算法是一种模拟生物进化过程的优化算法,它模拟了自然界中生物进化的过程,通过选择、交叉和变异等操作来搜索最优解。
遗传算法的核心是遗传操作,其基本公式如下:1. 初始化种群遗传算法首先需要初始化一个种群,种群中的每个个体都代表了问题的一个解。
个体的表示方式可以是二进制、十进制或其他形式,具体根据问题的特点而定。
2. 评估适应度对每个个体进行适应度评估,以确定其在问题中的优劣程度。
适应度函数的选择很关键,它应能准确地评估个体的性能,使优秀个体具有较高的适应度值。
3. 选择操作根据适应度值选择优秀个体作为父代,采用轮盘赌选择、竞争选择或其他选择策略,使适应度较高的个体有更大的概率被选中。
4. 交叉操作选中的父代个体通过交叉操作产生新的个体。
交叉操作可以是单点交叉、多点交叉、均匀交叉等,不同的交叉方式会对个体的基因组合方式产生影响。
5. 变异操作对新个体进行变异操作,以增加种群的多样性。
变异操作可以是位变异、插入变异、倒位变异等,通过改变个体的某些基因值来引入新的解。
6. 新种群生成通过选择、交叉和变异操作,生成新的种群。
新种群中的个体包括父代个体和经过交叉变异产生的子代个体。
7. 重复迭代重复进行评估适应度、选择、交叉和变异操作,直到满足终止条件。
终止条件可以是达到最大迭代次数、找到满意解或达到一定的收敛条件。
遗传算法模型公式的应用范围广泛,可以用于解决各种优化问题,如旅行商问题、背包问题、排课问题等。
通过模拟生物进化的过程,遗传算法能够在解空间中搜索到全局最优解或接近最优解的解。
总结起来,遗传算法模型公式包括了初始化种群、评估适应度、选择操作、交叉操作、变异操作和新种群生成等步骤。
通过不断的迭代优化,遗传算法能够搜索到问题的最优解。
遗传算法作为一种启发式算法,在解决复杂问题时具有很高的效率和鲁棒性。
在实际应用中,可以根据问题的特点灵活选择适当的遗传算法参数和操作策略,以获得更好的优化结果。
动物模型的分类

动物模型的分类动物模型是指以动物作为实验对象来研究和验证科学问题的工具。
根据不同的研究目的和实验需求,动物模型可以分为多种分类。
本文将就动物模型的分类进行详细介绍。
一、按照研究目的分类1. 疾病模型:用于研究特定疾病的发生机制、诊断方法和治疗策略。
例如,使用小鼠模型研究癌症的发生和治疗方法。
2. 行为模型:用于研究动物的行为特征、学习记忆、情绪等。
例如,使用大鼠模型研究抑郁症的行为表现和机制。
3. 发育模型:用于研究动物的胚胎发育过程、器官形成和功能发育。
例如,使用斑马鱼模型研究胚胎发育的遗传机制。
4. 药效学模型:用于评估药物的安全性和有效性。
例如,使用猕猴模型评估新药的药代动力学和毒性。
5. 遗传模型:用于研究基因对生物特征和疾病的影响。
例如,使用果蝇模型研究遗传突变对生命活动的影响。
二、按照实验动物种类分类1. 哺乳动物模型:包括人类近缘动物和实验室动物。
人类近缘动物如猴类、猿类等,其遗传与生理特征较接近人类,可以更准确地预测人类的反应。
实验室动物如小鼠、大鼠等,其繁殖能力强,容易获取,成本较低。
2. 禽类模型:包括鸡、鸭、鹅等。
禽类具有独特的生理特征,如鸡蛋的孵化过程、鸟类的迁徙行为等,可以用于研究特定的生物学问题。
3. 水生动物模型:包括斑马鱼、鲤鱼、鳗鱼等。
水生动物具有透明胚胎和容易获取的特点,适用于观察胚胎发育和遗传变异。
4. 爬行动物模型:包括蜥蜴、蛇、乌龟等。
爬行动物的生理特征与哺乳动物有所不同,可以用于研究特定的生物学问题,如蜥蜴的再生能力。
三、按照动物模型的应用领域分类1. 医学研究:用于研究疾病的发生机制、诊断方法和治疗策略。
例如,使用小鼠模型研究心脏病的发生机制和新药的疗效。
2. 药物研发:用于评估药物的安全性和有效性。
例如,使用猕猴模型评估新药的药代动力学和毒性。
3. 农业科学:用于提高农作物的产量和品质。
例如,使用水稻模型研究抗虫性和耐旱性的改良。
4. 环境科学:用于评估环境污染对生物的影响。
肿瘤动物模型常用建立方法

肿瘤动物模型常用建立方法肿瘤动物模型是用于研究和测试肿瘤发生、发展和治疗的工具。
建立适当的肿瘤动物模型对于揭示肿瘤的生物学特性和评估各种治疗方法的有效性至关重要。
以下是常用的肿瘤动物模型建立方法。
1. 移植瘤模型:这是最常见和简化的动物模型建立方法之一。
它涉及在动物体内或体外移植人类或动物来源的肿瘤细胞株。
这些细胞可以从肿瘤组织中分离得到,并在实验室中培养。
移植瘤模型的优点是易于建立和控制,但它不能反映肿瘤的整个发展过程。
2. 转基因模型:转基因动物模型是通过将特定的基因突变导入小鼠或其他动物体内来模拟肿瘤。
这些基因突变可以是人类肿瘤相关基因的突变,也可以是具有肿瘤形成潜能的其他基因的突变。
转基因模型可以提供更真实的肿瘤发展和治疗反应,但其建立过程相对复杂和耗时。
3. 化学诱发模型:这种方法通过给动物暴露于化学物质,如化学致癌物质或腺病毒,来诱发肿瘤的发生。
这些化学物质可以引起DNA损伤或基因突变,从而促进肿瘤的形成。
化学诱导模型可以提供与人类肿瘤相似的病理特征,但其应用范围受到化学物质的选择和剂量的限制。
4. 遗传模型:遗传模型使用特定品系的小鼠或其他动物,这些动物因其自身的遗传缺陷而具有高发生肿瘤的风险。
这些遗传模型可以是自然突变品系,也可以是通过基因工程技术引入的遗传缺陷。
遗传模型可以提供对特定肿瘤类型和易感因素的研究,但其适用范围受到特定品系的限制。
以上是常见的肿瘤动物模型建立方法。
根据具体研究目的和研究条件的不同,选择合适的肿瘤动物模型对于取得可靠的研究结果至关重要。
不同模型的优劣势需要综合考虑,并根据研究的需要进行合理选择。
基于历史数据的股票预测模型研究

基于历史数据的股票预测模型研究随着经济全球化的发展,股票市场成为了每个国家的重要组成部分。
越来越多的人投资股票,希望获取利益。
然而,股票市场不稳定,波动大,风险也非常高。
因此,越来越多的人希望通过股票预测模型来降低风险,提高收益。
本文将从历史数据出发,探究一些常见的基于历史数据的股票预测模型,帮助投资者更好地进行股票投资。
1. 趋势预测模型趋势预测模型是一种基于历史股票价格数据的较为简单的预测模型,它通过分析历史价格的趋势,来预测未来股票价格变化的方向。
这种模型非常适用于那些价格波动不大,股票价格的趋势明显的股票。
对于这类股票,我们可以使用线性回归模型,通过对历史数据的拟合,预测未来价格的趋势。
然而,这种模型的缺点也很明显,它并不能准确地预测价格的具体数值。
因为股票市场是非常复杂的,价格受到多方面因素的影响,仅仅凭借历史价格数据进行预测,很难达到较高的精确度。
2. 时间序列模型时间序列模型是一种基于历史数据的经典股票预测方法。
这种模型通过历史价格数据找到股票价格的周期性变化规律,从而预测未来价格的变化趋势。
常见的时间序列模型包括AR模型、MA模型、ARMA模型、ARIMA模型等。
以ARIMA模型为例,它是将AR模型和MA模型的优点结合起来,进一步拓展出来的一种方法。
ARIMA模型的核心思想是通过“自回归”、“差分”、“移动平均”等手段,在历史股票价格数据的基础上,找到价格变化的周期性规律,进而预测未来价格的变化。
预测过程中涉及到的自回归项、差分次数、移动平均项等参数,需要通过数据分析来确定。
这种方法准确性比较高,但是需要更多的数据样本支撑,且计算过程较为复杂。
3. 人工神经网络模型人工神经网络模型是近年来发展非常快的一种股票预测方法。
它是模拟人脑神经系统,通过训练网络中的神经元,来实现对未来股票价格的预测。
在神经网络模型中,输入层接受历史股票价格数据,隐层和输出层分别对数据进行处理和预测。
在训练模型时,需要大量的历史数据支持,不断调整网络中的参数和结构,最终得到合适的预测效果。
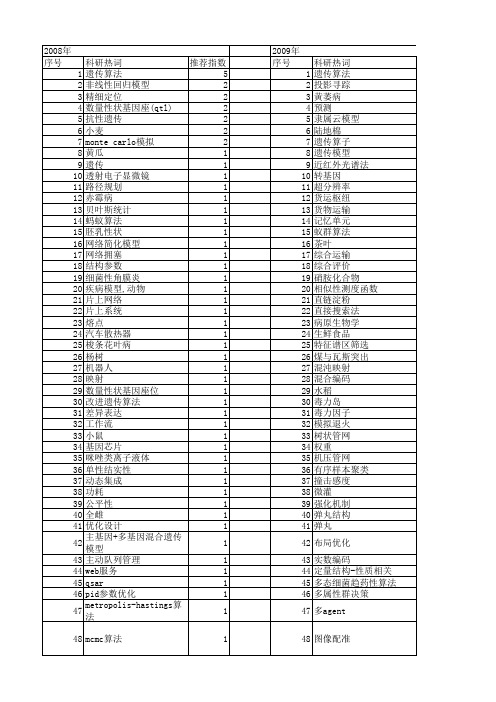
【江苏省自然科学基金】_遗传模型_期刊发文热词逐年推荐_20140815
1 1 1 1 1 1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
53 54 55 56 57 58 59法 内弹道 免疫遗传算法(iga) 免疫算法 偏最小二乘法 优化设计 优化模型 优化 任务分配 不可行度 rva谱 qtl作图 bp网络 2型猪链球菌
推荐指数 7 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49
遗传信息翻译的模型制作

遗传信息翻译的模型制作《普通高中生物课程标准》将模型知识列为课程的知识目标之一。
积极开展生物学模型的构建能很好地培养学生的科学素养。
高中生物必修2中遗传信息的表达抽象复杂,学生难以理解,如教师采用自制模型进行演示即可将抽象复杂转化为直观形象,并能收到良好的效果。
一、模型教具的制作1.材料硬纸板、木直尺、彩色磁钉、细铁丝、小铁钉、空心的圆珠2.制作方法(1)用硬纸板做成核糖体形状在大小亚基间开出两个小框(显示遗传密码)。
框上分别注明1、2两个位置。
每个小框内显示一个遗传密码,在该模板背面1、2两个位置的下方小亚基部位各钉上一个小铁钉。
小铁钉外套上两个空心的圆珠。
(2)mrna模型用长为1米木直尺。
在木尺面上连续写下碱基每相连的3个字母所占的长度和高度与核糖体模型上的小方框的长度一致。
同时用铁钉在木尺下方钉上一个与该木尺面垂直的同样长度的木尺,利于核糖体背后的空心圆珠在该垂直的木尺上沿着遗传密码移动。
(3)氨基酸模型用彩色磁钉的不同颜色代表不同的氨基酸。
并在磁钉上写出氨基酸名称和相对应的遗传密码。
(4)trna模型用细铁丝做成三叶草叶形状的rna模型若干个,并在每个trna 反密码子端写上各自的反密码子。
二、演示方法1.trna与氨基酸的结合任取一个trna模型并显示其反密码子,(如反密码子是uac的trna)让学生回答该trna携带的氨基酸的遗传密码和该氨基酸的名称(应为甲硫氨酸、密码为aug),然后将该氨基酸(磁钉)与trna(细铁丝)连接起来。
2.二肽的形成将核糖体模型放在标有遗传密码的木尺前面,同时使核糖体模型后的空心圆珠紧靠在与标有遗传密码的木尺垂直的木尺上。
让学生读出核糖体模型的框内显示的遗传密码并思考:应是携带什么氨基酸的trna进入。
如框内显示aug则携带甲硫氨酸的trna的(反密码子为uac)首先进入位点1,位点2显示cac则携带组氨酸的trna反密码子gug进入位点2。
甲硫氨酸与组氨酸间由于是磁钉就吸在一起形成二肽。
方差分析的遗传模型

方差分析的遗传模型方差分析的遗传模型:揭示和利用遗传多样性的潜能。
概述方差分析的遗传模型是一种强大的统计分析方法,用于研究遗传因素,它被广泛应用于研究遗传变异的形成、遗传基因关系的研究以及遗传疾病的形成机制。
该模型综合分析了遗传因素、环境因素和它们之间的相互作用,以梳理遗传变量文献中建议的模型,可以分析个体间的亲缘变异和环境变异,进而探索深层次的遗传因素。
结构方差分析的遗传模型包括两个分析过程:单位变量分析和综合变量分析。
单位变量分析是指将研究对象分为几个类别,单独计算每个类别的方差,以及比较各个类别之间的方差情况,这样就可以推断出各个类别之间是否存在遗传变异。
综合变量分析是指将所有研究对象视为一个整体,计算所有数据集中变量的方差爱这些变量在综合变量分析中称之为因子,然后将每个变量的方差都拆分为两部分,一部分是多样性变量,另一部分是因子变量。
其中,多样性变量表示变量之间的因素间变量,而因子变量表示因素内部变量(单位变量)。
工作流程方差分析的遗传模型的工作流程如下:1.数据采集:获取相关性评价、实验数据、相关技术参数等数据;2.数据处理:对数据进行清洗、标准化和离散化等处理;3.单因素方差分析:对所获取的单独变量或因子变量进行方差分析,计算变量与变量之间的关系;4.多变量分析:根据单因素方差分析结果,进行多变量方差分析,研究不同变量之间的复杂关系;5.分析结果:对分析结果进行判断和解释,研究不同变量之间的因果关系。
应用方差分析的遗传模型在研究人口遗传学领域有着广泛的应用。
例如,可以应用该模型来研究遗传性疾病具有遗传特征,以及家族中共性遗传疾病的发病机制;另外,也可以用该模型对人口遗传变化进行模拟,以及在两个不同人口之间进行交叉研究,从而了解不同人群之间遗传变异的情况和差异。
此外,还可以应用该方法进行基因功能研究,以及植物及动物的育种研究。
优势方差分析的遗传模型具有多重优势,首先,其可以对具有遗传特征的变量进行综合性分析,从而更好地探索探究遗传变异;其次,它可以从多个变量之间的关系出发,更有效地展示变量之间的复杂关系;最后,其还可以建立模拟的模型,从而更好地计算和识别基因独特的功能,为未来的研究奠定良好的基础。
遗传模型的适应性检验

遗传模型的适应性检验
遗传模型的适应性检验包括多个步骤:
(1)设置实验条件和数据搜集:在实验中设定合理的控制条件、模型测试参数及假设,并收集模型预测结果和观察值。
(2)分析市场反馈数据:研究市场表现与消费者行为,确定模型的预测结果是否与实际的观察一致。
(3)对模型进行统计性检验:计算相关系数、均值标准差、R系数等指标,确定遗传模型的准确性和可靠性。
(4)优化模型:进行参数优化操作,改造遗传模型的结构,以提高其准确度和有效性。
(5)反复检验模型:定期对模型进行重新检验,以确保其准确性、可靠性以及在实际应用中的有效性。
生物学中的模型

生物学中的模型生物学中的模型生物学是研究生命的科学,它关注生命如何从简单的化学反应和分子间相互作用演化成了复杂的生命形态,以及生命如何在不同层次上运作和交互。
为了更好地理解和研究生命,生物学家们常常使用模型来描述和解释生命的过程及其相互作用。
本文将介绍几种在生物学中常用的模型。
1. 生态系统模型生态系统模型是研究生态系统结构和功能的一种方法。
它通过在计算机上构建生态系统,来模拟生态系统中不同种群之间的相互作用,以及环境因素对生态系统的影响。
这种模型可以用来预测生态系统的响应,例如生物多样性损失、自然灾害和气候变化等。
同样的,生态系统模型也可以用来研究人为干扰对生态系统的影响,例如城市化、农业和工业污染等。
2. 分子模拟模型分子模拟模型是用来描述生物分子结构和功能的一种方法。
它基于物理化学原理,通过计算机程序模拟生物分子间的相互作用和运动。
这种模型可以用来研究许多生物分子的结构和功能,例如蛋白质、核酸和脂质等。
通过分子模拟模型,生物学家可以研究许多生物过程,例如生物催化、药物设计和分子信号传导等。
3. 细胞模型细胞模型是描述和解释细胞生物学过程的一种方法。
它通过计算机程序模拟细胞内各种分子和结构的相互作用和运动。
这种模型可以用来研究细胞的生长、分裂、代谢和信号传导等过程。
细胞模型可以帮助生物学家更好地理解细胞的功能和机制,同时也可以用来研究人类疾病的发生和治疗。
4. 人口遗传模型人口遗传模型是研究人类遗传学的一种方法。
它基于人口遗传学的理论,把人类作为一个群体来研究不同基因型和表型之间的相互作用。
这种模型可以用来研究人类基因的分布和变异,以及不同群体之间的遗传差异。
人口遗传模型也可以用来研究人类基因和疾病之间的关系,例如遗传性疾病的传播和流行。
总的来说,生物学中的模型是用来描述和解释生命过程及其相互作用的方法。
这些模型可以帮助生物学家更好地理解生命的机制和规律,同时也可以应用于医学和生物工程等领域,为人类提供更好的生活质量和健康保障。
ai大数据模型种类

ai大数据模型种类AI大数据模型种类随着人工智能技术的不断发展,各种AI大数据模型也应运而生。
这些模型是基于大数据的分析和学习,可以应用于各个领域,为人们的工作和生活带来了巨大的改变。
本文将介绍几种常见的AI大数据模型,它们的原理和应用场景。
1. 机器学习模型机器学习模型是最常见的AI大数据模型之一。
它通过训练数据集来学习规律和模式,并利用这些规律和模式进行预测和决策。
常见的机器学习模型包括决策树、支持向量机、神经网络等。
它们广泛应用于图像识别、自然语言处理、推荐系统等领域。
2. 深度学习模型深度学习模型是一种机器学习模型,它模仿人脑神经网络的结构和工作原理。
深度学习模型通过多层神经网络进行训练和学习,可以自动提取特征和模式。
深度学习模型在图像识别、语音识别、自然语言处理等领域取得了重大突破,如AlphaGo战胜人类围棋大师。
3. 强化学习模型强化学习模型是一种通过试错学习来优化决策的模型。
它通过与环境进行交互,不断尝试新的行动并根据反馈进行调整,以获得最大的回报。
强化学习模型在自动驾驶、游戏智能等领域具有广泛应用。
4. 遗传算法模型遗传算法模型是一种模拟生物进化过程的优化算法。
它通过模拟遗传操作(交叉、变异、选择)来搜索最优解。
遗传算法模型可以应用于优化问题、机器学习模型参数调优等领域。
5. 聚类模型聚类模型是一种将数据分组为具有相似特征的集合的模型。
它通过计算数据之间的相似度来判断数据是否属于同一组,从而进行数据分类和分析。
聚类模型广泛应用于市场分析、用户行为分析等领域。
6. 预测模型预测模型是一种根据历史数据和趋势来预测未来发展的模型。
它通过分析历史数据的规律和趋势,利用统计学和机器学习算法进行预测。
预测模型在金融、天气预报等领域具有重要的应用价值。
7. 生成模型生成模型是一种根据已有数据生成新数据的模型。
它通过学习已有数据的分布和规律,生成与原始数据相似的新数据。
生成模型在图像生成、音乐生成等领域具有潜在的应用前景。
简单遗传算法模型

简单遗传算法模型1. 概述遗传算法(GA)是一种启发式搜索算法,它受进化论中自然选择和遗传机制的启发而设计。
GA是一种非常强大的算法,它可以用来解决各种各样的优化问题。
2. 基本原理GA的基本原理是通过选择、交叉和变异操作,不断地优化一个种群的个体,最终使种群收敛到最优解。
选择:选择是GA中最关键的操作之一,它决定了哪些个体会进入下一代。
常用的选择方法有轮盘赌选择、锦标赛选择和精英选择等。
交叉:交叉是GA中另一个重要的操作,它通过交换两个个体的基因来生成新的个体。
交叉可以增加种群的多样性,提高GA的搜索能力。
常用的交叉方法有单点交叉、双点交叉和均匀交叉等。
变异:变异是GA中的一种随机操作,它通过改变个体的基因来产生新的个体。
变异可以防止种群过早收敛到局部最优解,提高GA的全局搜索能力。
常用的变异方法有比特翻转变异、均匀变异和高斯变异等。
3. 算法流程GA的算法流程如下:1. 初始化种群:随机生成一个种群,每个个体由一组基因组成。
2. 评估种群:计算每个个体的适应度值。
3. 选择:根据个体的适应度值,选择一部分个体进入下一代。
4. 交叉:对选出的个体进行交叉操作,生成新的个体。
5. 变异:对新的个体进行变异操作,生成最终的种群。
6. 重复步骤2-5,直到达到终止条件。
4. 优点和缺点GA具有以下优点:能够解决各种各样的优化问题。
不需要对问题做任何假设,也不需要任何先验知识。
能够找到全局最优解,而不是局部最优解。
GA也存在以下缺点:计算量大,尤其是当问题规模较大时。
容易陷入局部最优解,尤其是当问题具有多个局部最优解时。
难以确定合适的参数,例如种群规模、交叉率和变异率等。
5. 应用GA已被广泛应用于各种各样的领域,包括:组合优化:旅行商问题、背包问题、车辆路径问题等。
机器学习:神经网络训练、支持向量机训练、决策树学习等。
进化计算:遗传规划、进化策略、进化编程等。
6. 总结GA是一种非常强大的算法,它可以用来解决各种各样的优化问题。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
常染色体遗传模型
• 在常染色体遗传中,后代是从每个亲体 在常染色体遗传中, 的基因对中各继承一个基因, 的基因对中各继承一个基因,形成自己 的基因对,基因对也称为基因型。 的基因对,基因对也称为基因型。如果 我们所考虑的遗传特征是由两个基因A 我们所考虑的遗传特征是由两个基因A 控制的,那么就有三种基因对, 和a控制的,那么就有三种基因对,记 AA,Aa,aa。 为AA,Aa,aa。
0
0
Aa aa
0 0
1/2 0
1 0
1/2 1/4
1/2 1/2
0 1
常染色体遗传模型
• 农场的植物园中某种植物的基因型为 AA,Aa和aa。农场计划采用 型的植 , 和 。农场计划采用AA型的植 物与每种基因型植物相结合的方案培育 植物后代。那么经过若干年后, 植物后代。那么经过若干年后,这种植 物的任一代的三种基因型分布如何? 物的任一代的三种基因型分布如何?
x( n) an = bn c n
x(0)表示植物基因型的初始分布 即培育开 表示植物基因型的初始分布(即培育开 始时的分布), 始时的分布 ,显然有 an+bn+cn=1
常染色体遗传模型
• (ii)第 n-1代的分布与第 代的分布关系 第 代的分布与第n代的分布关系 代的分布与第 是通过上表确定的: 是通过上表确定的:
常染色体遗传模型
• 模型讨论: 模型讨论: • 若在上述问题中, 不选用因型 若在上述问题中 , 不选用因型AA植物 植物 与每一植物结合, 与每一植物结合,而是将具有相同基因 型植物相结合, 型植物相结合,那么后代所具有三种基 因型的概率由下表给出: 因型的概率由下表给出:
父体——母体的基因型 母体的基因型 父体 AA-AA - 后 代 AA 基 因 Aa 型 aa 1 0 0 Aa-Aa - 1/4 1/2 1/4 aa-aa - 0 0 1
x
(n)
an = bn
常染色体隐性病模型
• (iii)为使每个儿童至少有一个正常的父 为使每个儿童至少有一个正常的父 亲或母亲, 亲或母亲,因此正常人或隐性患者必须 与正常人结合, 与正常人结合,其后代的基因型概率由 表给出: 表给出:
父体——母体的基因型 母体的基因型 父体 AA-AA - 后 代 基 因 型 AA Aa aa 1 0 0 AA-Aa - 1/4 1/2 1/4
X—链遗传模型 链遗传模型
• (iv)如果第 - 1代配偶的同胞对是 , 如果第n- 代配偶的同胞对是 代配偶的同胞对是(A, 如果第 Aa)型 , 那么它们的雄性后代将等可能 型 地得到基因A或 , 地得到基因 或a,它们的雌性后代的基 因型将等可能地是AA或 A。 又由于第 因型将等可能地是 或 。 又由于第n 代雌雄结合是随机的, 那么第n代配偶 代雌雄结合是随机的 , 那么第 代配偶 的同胞对将等可能地为四种类型(A, 的同胞对将等可能地为四种类型 , AA), (A, Aa),, , AA), (a,Aa)之 ,,(a, , , ,, , 之 一。其他类似可分析
X—链遗传模型 链遗传模型
父体一母体的基因型 (A,AA) ) A a 后 代 基 因 型 AA 1 0 1 (A,Aa) 1/2 1/2 1/2 (A,aa) 0 1 0 (a,AA) 1 0 0 (a,Aa) 1/2 1/2 0 (a,aa) 0 1 0
Aa aa
0 0
1/2 0
1 0
1 0
1/2 1/2
遗传模型
1.常染色体遗传 常染色体遗传 2.X—链遗传 链遗传
遗传模型
• 随着人类的进化,人们为了揭示生命的 随着人类的进化, 奥妙,越来越注重遗传学的研究, 奥妙,越来越注重遗传学的研究,特别 是遗传特征的逐代传播, 是遗传特征的逐代传播,引起人们更多 的注意。无论是人,还是动、 的注意。无论是人,还是动、植物都会 将本身的特征遗传给下一代, 将本身的特征遗传给下一代,这主要是 因为后代继承了双亲的基因, 因为后代继承了双亲的基因,形成自己 的基因对, 的基因对,基因对确定了后代所表现的 特征。 特征。
常染色体遗传模型
• 建模: 根据假设 ,先考虑第 代中的 建模: 根据假设(ii),先考虑第n代中的 AA型。由于第 -1代的 型与 型结 代的AA型与 型 由于第n- 代的 型与AA型结 后代全部是AA型: 第 n-l代的 型 代的Aa型 合 , 后代全部是 型 代的 型结合, 型的可能性为1、 与AA型结合,后代是 型的可能性为 、 型结合 后代是AA型的可能性为 2,而第 - l代的 型与 代的aa型与 型结合, , 而第n- 代的 型与AA型结合, 后 型结合 代 不 可 能 是 AA 型 。 因 此 当 n=1 , 2 , 3…….. • an=1an-1 +1/2bn-1+0cn-1 - - -
0 1
X—链遗传模型 链遗传模型
• (iii)在每一代中 , 配偶的同胞对也是六 在每一代中, 在每一代中 种类型之一,并有确定的概率。 种类型之一,并有确定的概率。为计算 这些概率, 这些概率,设an,bn,cn,dn,en和fn分 别是第n代中 配偶的同胞对为(A, 代中, 别是第 代中,配偶的同胞对为 ,AA), , (A,Aa),(A,aa),(a,AA),(a,Aa), , , , , , , , (a,aa)型的概率,n=0,1,…。令 型的概率, , 型的概率 , , 。 • x(n)=(an , bn , cn , dn , en , fn)T , n=0, , 1,… ,
常染色体遗传模型
• 下面给出双亲体基因型的所有可能的结 使其后代形成每种基因型的概率: 合,使其后代形成每种基因型的概率:
父体——母体的基因型 母体的基因型 父体 AA-AA - AA-Aa - AA-aa - Aa-Aa - Aa型
1
1/2
0
1/4
X—链遗传模型 链遗传模型
• X—链遗传是指雄性具有一个基因 或a, 链遗传是指雄性具有一个基因A或 , 链遗传是指雄性具有一个基因 雌性具有两个基因AA, 或 Aa, 或 aa。 雌性具有两个基因 , , 。 其遗传规律是雌性后代以相等概率得到 母体两个基因中的一个, 母体两个基因中的一个,雌性后代从父 体中得到一个基因, 体中得到一个基因,并从母体的两个基 因中等可能地得到一个。下面, 因中等可能地得到一个。下面,研究与 X—链遗传有关的近亲繁殖过程。 链遗传有关的近亲繁殖过程。 链遗传有关的近亲繁殖过程
常染色体遗传模型
• 假设:(1)设an,bn和cn。分别表示第 代 假设: 设 分别表示第n代 植物中, 基因型为AA, Aa和 aa的植物 植物中 , 基因型为 , 和 的植物 占植物总数的百分率。 为第n代植 占植物总数的百分率 。 令 x(n) 为第 代植 物的基因型分布: 物的基因型分布:
常染色体遗传模型
• 这里因为 和AA都表示了同一外部特 这里因为Aa和 都表示了同一外部特 我们认为基因A支配基因 支配基因a, 征,我们认为基因 支配基因 ,也可认 为基因a对于 来说是隐性的。 对于A来说是隐性的 为基因 对于 来说是隐性的。当一个亲 体的基因型为Aa, 体的基因型为 ,而另一个亲体的基因 型是aa,那么后代可以从aa型中得到基 型是 ,那么后代可以从 型中得到基 型中得到基因A, 因a,从Aa型中得到基因 ,或得到基 , 型中得到基因 因a。这样,后代基因型为 或aa的可 。这样,后代基因型为Aa或 的可 能性相等
常染色体隐性病模型
• 假设 • (i)常染色体遗传的正常基因记为 , 不 常染色体遗传的正常基因记为A, 常染色体遗传的正常基因记为 正常基因记为a,并以AA, , 分别 正常基因记为 ,并以 ,Aa,aa分别 表示正常人,隐性患者, 表示正常人,隐性患者,显性患者的基 因型 • (ii)设an , bn 分别表示第 代中基因型为 分别表示第n代中基因型为 设 AA,Aa 占总人数的百分数,记 , 占总人数的百分数,
X—链遗传模型 链遗传模型
• 假设 • (i)从一对雌雄结合开始 , 在它们的后代中 , 从一对雌雄结合开始, 在它们的后代中, 从一对雌雄结合开始 任选雌雄各一个成配偶, 然后在它们产生的 任选雌雄各一个成配偶 , 后代中任选两个结成配偶。如此继续下去。 后代中任选两个结成配偶。如此继续下去。 • (ii)父体与母体的基因型组成同胞对 , 同胞对 父体与母体的基因型组成同胞对, 父体与母体的基因型组成同胞对 的形式有(A, 的形式有 ,AA),(A,Aa),(A,aa),(a, , , , , , , AA),(a, Aa),(a,aa)。 初始一对雌雄的同 , , , , 。 胞对, 是这六种类型中的任一种, 胞对 , 是这六种类型中的任一种 , 其后代的 基因型由表给出。 基因型由表给出。
常染色体隐性病模型
• 现在世界上已经发现的遗传病将近 现在世界上已经发现的遗传病将近4000 在一般情况下, 种。在一般情况下,遗传疾病是与特殊 的种族、部落及群体有关。例如, 的种族、部落及群体有关。例如,遗传 病库利氏贫血症的患者以居住在地中海 沿岸为多, 沿岸为多,镰状网性贫血症一般流行在 黑人中, 黑人中,家族黑蒙性白痴症则流行在东 欧犹太人中间。 欧犹太人中间。患者经常未到成年就痛 苦地死去, 苦地死去,而他们的父母则是疾病的病 源。
常染色体隐性病模型
• 假若我们能识别这些疾病的隐性患者, 假若我们能识别这些疾病的隐性患者, 并且规定隐性患者不能结合(因为两个隐 并且规定隐性患者不能结合 因为两个隐 性病患者结合, 性病患者结合,他们的后代就可能成为 显性患者),那么未来的儿童, 显性患者 ,那么未来的儿童,虽然有可 能是隐性患者,但绝不会出现显性特征, 能是隐性患者,但绝不会出现显性特征, 不会受到疾病的折磨。现在, 不会受到疾病的折磨。现在,我们考虑 在控制结合的情况下, 在控制结合的情况下,如何确定后代中 隐性患者的概率。 隐性患者的概率。