【国家自然科学基金】_1,5-二磷酸核酮糖羧化酶_期刊发文热词逐年推荐_20140731
【国家自然科学基金】_核酮糖-1_基金支持热词逐年推荐_【万方软件创新助手】_20140730
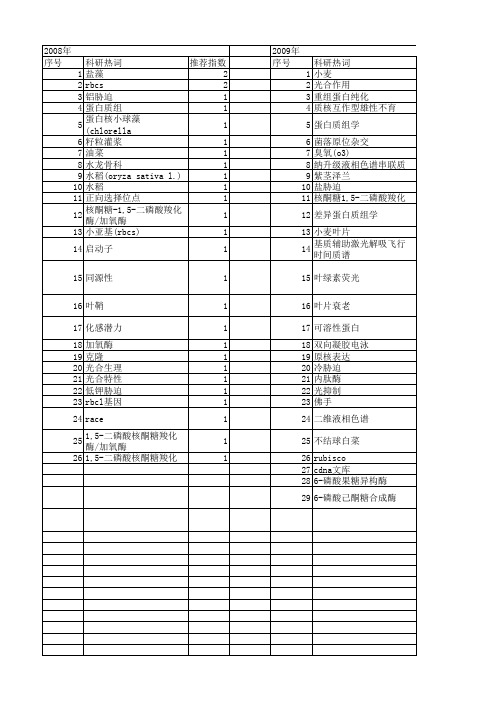
科研热词 推荐指数 盐藻 2 rbcs 2 铝胁迫 1 蛋白质组 1 蛋白核小球藻(chlorella pyrenoidosa) 1 籽粒灌浆 1 油菜 1 水龙骨科 1 水稻(oryza sativa l.) 1 水稻 1 正向选择位点 1 核酮糖-1,5-二磷酸羧化酶/加氧酶1 小亚基(rbcs) 1 启动子 1 同源性 1 叶鞘 1 化感潜力 1 加氧酶 1 克隆 1 光合生理 1 光合特性 1 低钾胁迫 1 rbcl基因 1 race 1 1,5-二磷酸核酮糖羧化酶/加氧酶 1 1,5-二磷酸核酮糖羧化酶 1
53 54
2011年 科研热词 推荐指数 基因表达 3 黄瓜 2 植物表达载体 2 拟南芥 2 弱光 2 叶绿体定位 2 光合作用 2 黄瓜叶片 1 银杏叶 1 野大麦 1 酶活性 1 酵母双杂交系统 1 通路克隆技术 1 通路克隆(gateway)技术 1 适应性进化 1 蛋白互作 1 聚乙二醇 1 种子重量 1 盐胁迫 1 溃疡病菌 1 温室效应 1 油菜 1 毛白杨 1 核酮糖-1,5-二磷酸羧化酶/加氧酶1 核酮糖-1,5-二磷酸羧化/加氧酶(rubisco) 1 微生物 1 序列分析 1 差异蛋白质 1 小麦 1 固碳 1 含油量 1 双向电泳 1 双向凝胶电泳 1 北京杨 1 分子生态 1 凤尾蕨科 1 冷胁迫 1 共进化 1 入门载体 1 光诱导型启动子 1 信号转导 1 亚适温 1 亚弱光 1 二磷酸核酮糖羧化酶小亚基3c启动子 1 rubisco相互作用蛋白质 1 rubisco活化酶(rca) 1 rbcl基因 1 pen-l4*-prbcs-5-二磷酸核酮糖羧化酶/加氧酶小亚基 1
核酮糖1,5-二磷酸羧化酶(RuBPCase)检测试剂盒(RuBP微板法)
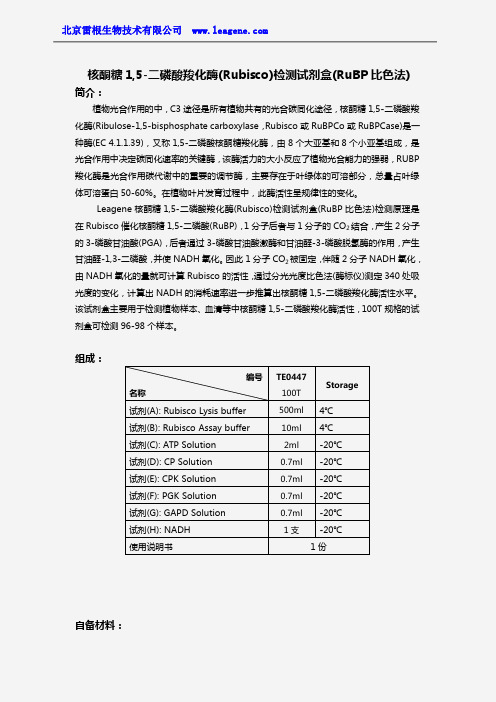
核酮糖1,5-二磷酸羧化酶(Rubisco)检测试剂盒(RuBP比色法)简介:植物光合作用的中,C3途径是所有植物共有的光合碳同化途径,核酮糖1,5-二磷酸羧化酶(Ribulose-1,5-bisphosphate carboxylase,Rubisco或RuBPCo或RuBPCase)是一种酶(EC4.1.1.39),又称1,5-二磷酸核酮糖羧化酶,由8个大亚基和8个小亚基组成,是光合作用中决定碳同化速率的关键酶,该酶活力的大小反应了植物光合能力的强弱,RUBP 羧化酶是光合作用碳代谢中的重要的调节酶,主要存在于叶绿体的可溶部分,总量占叶绿体可溶蛋白50-60%。
在植物叶片发育过程中,此酶活性呈规律性的变化。
Leagene核酮糖1,5-二磷酸羧化酶(Rubisco)检测试剂盒(RuBP比色法)检测原理是在Rubisco催化核酮糖1,5-二磷酸(RuBP),1分子后者与1分子的CO2结合,产生2分子的3-磷酸甘油酸(PGA),后者通过3-磷酸甘油酸激酶和甘油醛-3-磷酸脱氢酶的作用,产生甘油醛-1,3-二磷酸,并使NADH氧化。
因此1分子CO2被固定,伴随2分子NADH氧化,由NADH氧化的量就可计算Rubisco的活性,通过分光光度比色法(酶标仪)测定340处吸光度的变化,计算出NADH的消耗速率进一步推算出核酮糖1,5-二磷酸羧化酶活性水平。
该试剂盒主要用于检测植物样本、血清等中核酮糖1,5-二磷酸羧化酶活性,100T规格的试剂盒可检测96-98个样本。
组成:自备材料:编号名称TE0447100TStorage 试剂(A):Rubisco Lysis buffer500ml4℃试剂(B):Rubisco Assay buffer10ml4℃试剂(C):ATP Solution2ml-20℃试剂(D):CP Solution0.7ml-20℃试剂(E):CPK Solution0.7ml-20℃试剂(F):PGK Solution0.7ml-20℃试剂(G):GAPD Solution0.7ml-20℃试剂(H):NADH1支-20℃使用说明书1份1、研钵或匀浆器2、离心管3、低温离心机4、恒温箱或水浴锅5、96孔板6、酶标仪操作步骤(仅供参考):1、准备样品:①植物样品:取植物组织清洗干净,切碎,植物组织:Rubisco Lysis buffer按一定的比例,加入预冷的Rubisco Lysis buffer,冰浴情况下充分匀浆或研磨。
【国家自然科学基金】_rbcs_基金支持热词逐年推荐_【万方软件创新助手】_20140801
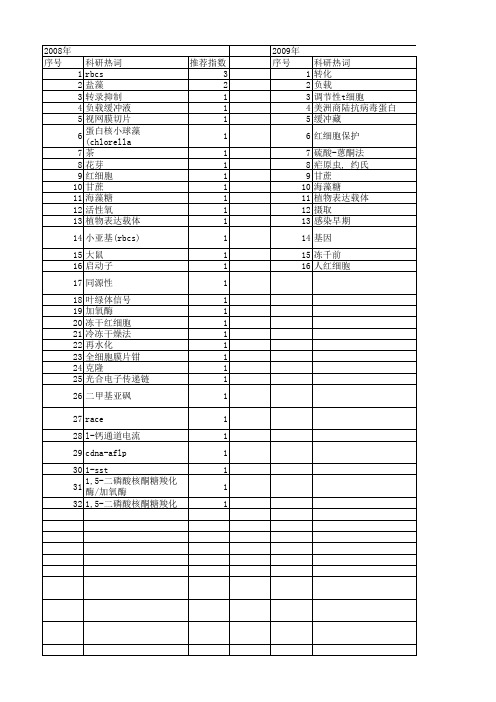
科研热词 推荐指数 边缘细胞 2 水稻 2 黑豆 1 黏液 1 黄瓜嫁接 1 颗粒 1 铝毒 1 铝形态 1 转录表达 1 超氧化物歧化酶活性 1 超冷铷铯分子 1 谷胱甘肽过氧化物酶 1 诱导系统耐性(ist) 1 血红蛋白 1 蜡质芽胞杆菌 1 蛋白核小球藻 1 耐旱性 1 网织红细胞 1 缺铁性贫血 1 细胞壁多糖 1 红细胞(rbc) 1 红细胞 1 相对重构贡献(rrbc) 1 盐胁迫 1 电力变压器 1 生防菌 1 生长周期 1 生理特性 1 牛巴贝斯虫 1 热工学 1 灰关联 1 溶解气体分析(dga) 1 渗透参数 1 氢损伤 1 气体交换 1 最佳冷却速率 1 故障诊断 1 振转光谱 1 抗氧化酶活性 1 患者 1 干旱胁迫 1 启动子 1 叶绿素荧光 1 原子力显微镜成像 1 光诱导表达载体 1 光缔合 1 光合作用 1 侵染性 1 体外培养 1 主元分析(pca) 1 yisui shengxue granule 1 ultracold rbcs molecules 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
科研热词 转化 负载 调节性t细胞 美洲商陆抗病毒蛋白 缓冲藏 红细胞保护 硫酸-蒽酮法 疟原虫, 约氏 甘蔗 海藻糖 植物表达载体 摄取 感染早期 基因 冻千前 人红细胞
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
abo基因 1 1,5-二磷酸核酮糖羧化酶/加氧酶小亚基 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68
rubisco活化酶基因 rubisco小亚基 rubisco rovibrational spectrum root border cells rbcs基因 physiological characteristic photoassociation mucilage hemoglobin h disease gsh-px活性 dsc drought stress chinese medicine black soybean antioxidative effect
【国家自然科学基金】_耐盐基因_基金支持热词逐年推荐_【万方软件创新助手】_20140729
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 5253 cdna构建 54 3'-race
1 1
推荐指数 3 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
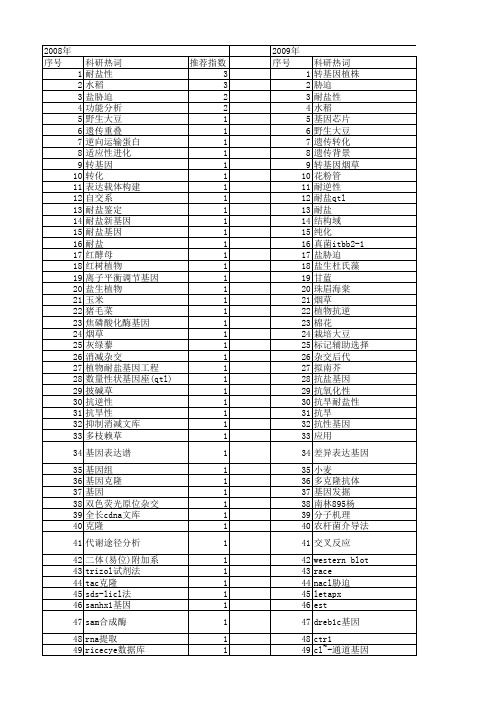
科研热词 转基因植株 胁迫 耐盐性 水稻 基因芯片 野生大豆 遗传转化 遗传背景 转基因烟草 花粉管 耐逆性 耐盐qtl 耐盐 结构域 纯化 真菌itbb2-1 盐胁迫 盐生杜氏藻 甘蓝 珠眉海棠 烟草 植物抗逆 棉花 栽培大豆 标记辅助选择 杂交后代 拟南芥 抗盐基因 抗氧化性 抗旱耐盐性 抗旱 抗性基因 应用 差异表达基因 小麦 多克隆抗体 基因发掘 南林895杨 分子机理 农杆菌介导法 交叉反应 western blot race nacl胁迫 letapx est dreb1c基因 ctr1 cl~-通道基因 cdna badh基因 3-磷酸甘油脱氢酶
核酮糖1,5-二磷酸羧化酶(RuBPCase)检测试剂盒(RuBP比色法)
核酮糖1,5-二磷酸羧化酶(Rubisco)检测试剂盒(RuBP比色法)简介:植物光合作用的中,C3途径是所有植物共有的光合碳同化途径,核酮糖1,5-二磷酸羧化酶(Ribulose-1,5-bisphosphate carboxylase,Rubisco或RuBPCo或RuBPCase)是一种酶(EC4.1.1.39),又称1,5-二磷酸核酮糖羧化酶,分子量约为53kD,由8个大亚基和8个小亚基组成,是光合作用中决定碳同化速率的关键酶,该酶活力的大小反应了植物光合能力的强弱,RUBP羧化酶是光合作用碳代谢中的重要的调节酶,主要存在于叶绿体的可溶部分,总量占叶绿体可溶蛋白50-60%。
在植物叶片发育过程中,此酶活性呈规律性的变化。
Leagene核酮糖1,5-二磷酸羧化酶(Rubisco)检测试剂盒(RuBP比色法)检测原理是在Rubisco催化核酮糖1,5-二磷酸(RuBP),1分子后者与1分子的CO2结合,产生2分子的3-磷酸甘油酸(PGA),后者通过3-磷酸甘油酸激酶和甘油醛-3-磷酸脱氢酶的作用,产生甘油醛-1,3-二磷酸,并使NADH氧化。
因此1分子CO2被固定,伴随2分子NADH氧化,由NADH氧化的量就可计算Rubisco的活性,通过分光光度比色法(分光光度计)测定340处吸光度的变化,计算出NADH的消耗速率进一步推算出核酮糖1,5-二磷酸羧化酶活性水平。
该试剂盒主要用于检测植物样本、血清等中核酮糖1,5-二磷酸羧化酶活性,25T规格的试剂盒可检测23-24个样本。
组成:编号名称TE044925TStorage试剂(A):Rubisco Lysis buffer250ml4℃试剂(B):Rubisco Assay buffer15ml4℃试剂(C):ATP Solution3ml-20℃试剂(D):CP Solution 1.5ml-20℃试剂(E):CPK Solution1ml-20℃试剂(F):PGK Solution1ml-20℃试剂(G):GAPD Solution1ml-20℃试剂(H):NADH1支-20℃试剂(I):碱性基液3ml RT 使用说明书1份自备材料:1、研钵或匀浆器2、离心管3、低温离心机4、恒温箱或水浴锅5、比色杯6、分光光度计操作步骤(仅供参考):1、准备样品:①植物样品:取植物组织清洗干净,切碎,按植物组织:Rubisco Lysis buffer按一定比例,加入预冷的Rubisco Lysis buffer,冰浴情况下充分匀浆或研磨。
【国家自然科学基金】_黄瓜(cucumis sativus l.)_期刊发文热词逐年推荐_20140802
53 54 55 56 57 58 59 60 61 62 63 64
ph 1 nacl胁迫 1 dna序列 1 dna 1 cdna克隆 1 cdna-aflp 1 cdna 1 biozol 1 atpase 1 6-磷酸葡萄糖酸脱氢酶 1 5–二磷酸核酮糖羧化/加氧酶(rubisco) 1 1 1
推荐指数 6 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
推荐指数 16 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31
科研热词 黄瓜 黄瓜黑星病 雌雄异花同株 逆转录酶 诱导抗病性 致 热处理 杂种优势 抗热性 抗氧化系统 性型 异质性 幼苗 分子标记 几丁质酶 全雌性 保护酶 低氧胁迫 乙烯 β -1,3-葡聚糖酶 ty1-copia类逆转座子 pcr检测 24-表油菜素内酯
1,5-二磷酸羧化酶/加氧酶(Rubisco)

1,5-二磷酸羧化酶/加氧酶(Rubisco)植物生理学通讯第43卷第2期,2007年4月363核酮糖.1,5.二磷酸羧化酶/加氧酶(Rubisco)梅杨,李海蓝,谢晋,罗红艺华中师范大学生命科学学院,武汉430079Ribulose-I,5-bisphosphateCarboxylase/oxygenase(Rubisco)MEIY ang,LIHai—Lan,XIEJin,LUOHong—YiCollegeofLifeSciences,CentralChinaNormaliVPl魄Wuhan430079,China提要:文章就核酮糖一1,5.二磷酸羧化酶,加氧酶(Rubisco)的分布,结构,性质,分类与功能的研究进展作了介绍.关键词:核酮糖一1,5一二磷酸羧化酶/加氧酶(Rubisco);Rubisco类似蛋白(RLP);羧化/氧化值;热稳定性;分类核酮糖一1,5一二磷酸羧化酶/加氧酶(ribulose一1,5一bisphosphatecarboxylase/oxygenase,Rubisco)是植物光合作用过程中固定CO,的关键酶,同时也参与植物的光呼吸代谢途径,消耗植物光合作用合成的有机物,由此造成的净光合效率损失高达50%(Lundqvist和Schneider1991;熊晓然等2003;Ashida等2005).因此,研究Rubisco对提高植物的光合作用效率有重要意义.自从1953年Calvin等在研究光合碳循环过程中证实其存在至今,有关Rubisco的一系列问题不断得到阐明,人们对其性质,结构,类型和功能等诸多方面有了更深入的了解,这方面的研究己取得了较大进展,本文就此作介绍.1Rubisco的分布与定位Rubisco广泛分布于具光合功能的细胞器中.它是一个含量很丰富的酶,据估计全世界Rubisco 的量约有4×10吨.Rubisco在C植物中主要定位于叶肉细胞叶绿体问质中,在基粒片层上也有少量分布;在C植物中则定位于维管束鞘细胞中的叶绿体问质内.低等藻类的Rubisco主要定位在淀粉核上.许多化能自养细菌和蓝藻中存在多角形细胞内含物,其中含有大量的羧化酶,称之为羧基化小体.羧基化小体最初是从氧化硫细菌中分离出来的,并且含有大量的Rubisco.有研究表明,羧基化小体是原核生物体内储存Rubisco的重要场所,能够协助Rubisco在低浓度的CO,条件下完成羧化.那不勒斯硫杆菌(Thi0baci,,U neapolitanus)的Rubisco缺陷菌株无羧基化小体的形成,且必须在高浓度的CO,下才能生长(Baker等1998).海洋氢弧菌(Hydrogenovibriomarinus)MH一110在CO,浓度低于0.15%时就会形成羧基化小体(Y oshizawa等2004).2Rubisco的结构采用x射线晶体衍射技术,人们解析了不同来源的Rubisco的晶体结构,包括烟草(Nicotiana tabacum),菠菜(Spinaciaoleracea),深红红螺菌(Rhodospirillumrubrum),莱茵衣藻(Chlamydomonas reinhardtii),蓝藻聚球藻(y,zcDcDcc"PCC 6301),喜温红藻(Galdieriapartite),绿色硫细菌(chlorobiumtepidum),超耐热原始菌(菌, ThermococcuskodakaraensisKOD1)等(Kitano等2001;Parry等2003;Li等2005).Rubisco一般由多个大亚基(LSU)和小亚基(ssu)组成,其中大亚基的分子量为50~55kDa,小亚基为1218kDa(Ashida等2005).大亚基具有催化功能,小亚基仅具有调节作用.迄今的研究认为,Rubisco的大亚基由N,C两个结构域组成.N结构域从N末端开始,包括137个氨基酸,其中含有5股D折叠.C结构域中含有丰富的0c螺旋,其中以0c/D桶状结构域(al Dbarreldomain)最引人注目.它包括8个0c螺旋和8个D折叠,彼此连接成8个环.一个大亚基收稿2006.11—20修定2007—03一l2资助华中师范大学精品课程建设项目.通讯作者(E—mail:******************.ca:Tel:************).364植物生理学通讯第43卷第2期,2007年4月的羧基末端为另一个大亚基氨基末端部分覆盖,形成漏滴状的活性中心,Mg参与其中.Mg与亚基中的3个氨基酸残基所含有的氧原子发生作用,它们分别是氨甲酰化的Lys,侧链Asp3和Glulq4(Lundqvist和Schneider1991).C端结构域同时含有一个特征性的突环(Lo0p6).Loop6影响酶与气体分子(包括CO和O)的亲和性,一系列发生在环内的突变均能影响酶的羧化/氧化值(Q 值);在形成烯醇化中间产物的过程中,Loop6也起关键性的作用(Parry等2003).Loop6同时还影响酶活性状态的形成与维持.活性中心的模拟分析表明,Rubisco是否处于活性状态与活性中心(Lysl75,Lys201)和Loop6(Lys334)的3个Lys残基密切相关(熊晓然等2003).小亚基远离活性中心,其含有4个反向平行的p折叠和2个0c螺旋,p折叠和0c螺旋的核心含有疏水的氨基酸残基及与大小亚基相互作用有关的保守氨基酸残基.小亚基能促进CO,与Mg对酶的活化,维持和稳定酶的活化构象.Spreitzer (2003)认为,小亚基可能在进化过程中扮演聚集大亚基活性位点的角色,小亚基可能有更多特异性功能.3Rubisco的类型根据Rubisco大亚基氨基酸序列同源性及空间结构的相似性,可以将其分为4类,即I,II,III,IV型(表1).I型主宴存在于能够进行光合作用的有机体内,如高等植物,真核藻类,蓝藻,光能及化能自养细菌及其它一些原核生物,它由8个大亚基和8个小亚基组成,呈LS的结构.Tabita(1995)分析不同的I型Rubisco结构后,又将其分为"Green—like"和"Red—like"2类,并进一步将其划分为A,B,C及D4个亚类.II型Rubisco由2罐个大亚基组成,呈L的结构,其分子量为110-450kDa,主要存在于一些光能及化能合成细菌,海产甲藻如共生甲藻(SymbiodiniumSpp.),膝钩藻(Gonyaulaxpolyedra)等中(Rowan等1996;Nassoury等2001).尽管I型与II型Rubisco在活性位点上的氨基酸残基高度保守,但II型与I型的大亚基的同源性仍很低,仅为28%(Kitano等2001;Liao等2004).III型也仅由大亚基组成,呈L结构,存在于某些嗜热古生菌如菌,詹氏甲烷球菌等中(Klenk等1997;Watson和Tabita1999;Kitano等2001).有人研究Tk-Rubisco的结果表明,它是由大亚基组表1不同来源的Rubisco及其羧化/氧化值,结构和功能代表植物;一表示暂无相关数据.好热性硫磺细菌含有2类Rubisco:rbcL—l和rbcL.2(Karlin和Mrazek2000):超耐热原始菌先前报道称为PyrococcuskodakaraensisKODI(Kitano等2001).植物生理学通讯第43卷第2期,2007年4月365成的(L)的十聚体,与菠菜中的I型Rubisco有36%的同源性,与深红红螺菌中的Ⅱ型Rubisco有30%的同源性(Kitano等2001);分析詹氏甲烷球菌Rubisco的序列表明,它与蓝藻中的聚球藻I型Rubisco有41%的同源性,与深红红螺菌中的II型Rubisco有33%的同源性(Watson和Tabita1999).尽管以上三类Rubisco相互之间的同源性较低,但已知的Rubisco参与羧化或氧化核酮糖一1,5一二磷酸(ribulose一1,5一bisphosphate,RuBP)过程的所有保守氨基酸残基在I,II和III型中都存在,除了Ⅲ型中Phe.∞被其它氨基酸所取代(Ashida等2005).IV型又称为Rubisco类似蛋白(rubisco—like—protein,RLP),存在于非光合细菌,部分不依赖卡尔文循环的光合细菌和古生菌如绿色硫细菌,好热性硫磺细菌rbcL.,泥生绿菌,枯草芽孢杆菌等中(Klenk等1997;Watson和Tabita1999;Hanson和Tabita2001,2003Ashida等2003).它与I,II,III相比,许多活性位点保守氨基酸残基缺失,同源性甚低.如泥生绿菌,好热性硫磺细菌cL.,枯草芽孢杆菌中分别有11,5,9个活性位点的保守氨基酸残基被其它氨基酸所取代;枯草芽孢杆菌RLP与I,II,III型仅有23%,23%,30%的序列同源性(Ashida等2005).实际上,在许多细菌中同时存在编码I型和Ⅱ型Rubisco的结构基因,但在正常情况下两者并不同时表达(English等1992;Karlin和Mrazek 2000).那不勒斯硫杆菌在rbcL,突变的情况下, rbcL,,可表达生成II型Rubisco,但菌体必须在高浓度的CO,下才能生长良好(Baker等1998).海洋氢弧菌MH一110含有3个拷贝的Rubisco基因(CbbLS—和CbbLS-2属于I型,CbbM属于II型).Y oshizawa等(2004)研究证实海洋氢弧菌MH一110在不同浓度的CO,条件下,三者以不同的组合形式进行表达.脱氮硫杆菌在厌氧条件下以硝酸盐作为电子受体能同时表达生成I,II2种类型Rubisco.最近,Carr~一Mlouka等(2006)报道,在世界范围内广泛引起水华的铜绿微囊藻PCC7806 (MicrocystisaeruginosaPCC7806)细胞内同时存在I,IV型Rubisco.沼泽红假单胞菌(Rhodopseudo. monaspalustris)除了含有I,II型Rubisco,还同时存在2类不同的RLP(Larimer等2004).这些发现导致不同类型Rubisco的共存问题变得更加复杂,这对研究Rubisco的进化可能有意义.4Rubisco的性质与一般的酶相比,Rubisco具有2个显着的特征.一是非专一性,即Rubisco既能催化羧化反应,也能催化加氧反应,具有双功能性;二是低效性,即Rubisco的催化效率较一般酶低,是由于Rubisco酶转换数低(真核生物的Rubiscok约为3~5S~,来源于不同生长温度的植物Rubiscok略有不同),催化效率有限(Watson和Tabita1999;Sage2002~Ashida等2005).4.1热稳定性Rubisco有一定的热稳定性,但不同来源的Rubisco的热稳定性存在较大差异.水稻Rubisco在50℃保温7min达到最大活力,随后迅速下降,30min后酶活性下降至40%;烟草Rubisco在50℃保温20min达最大活力,30min后活力还维持在98%左右;菠菜Rubisco氨甲酰化后经60℃的10mmo1.LDTr处理1h活力还维持50%(陈为钧等1999).在嗜热古生菌如菌中,Rubisco的热稳定性极高.Tk.Rubisco在30—110℃范围内均能有效完成十聚体结构的组装(Maeda等2002);在40~100℃范围内能够保持有效的羧化酶活性,且活性随温度(40-90℃)的升高而上升;在80℃保温15h活力仍维持50%(Ezaki等1999).Maeda等(2002)采用定点突变(E63S,R66S,D69S)的方法,进一步研究证实Tk—RubiSCO耐热性依赖于特殊的五角形晶体结构(pentagonalstructure),此种结构的特点是二聚体相互接触紧密,接触面上的氨基酸残基之间存在离子间的相互作用,并形成8对离子键.同时,低聚状态对耐热性的维持也有影响.Tk.Rubisco的十聚体结构提高了蛋白质的变性温度,从而进一步提高了Tk菌适应高温的能力.4.2酶促反应动力学参数一般而言,C植物及景天科酸代谢植物Rubisco的Kin(CO)在12~26 ~tmol?L~,C4植物Rubisco的Kin(CO2)为28~63 ∞1.L-(Chen等2002).但不同类型植物Rubisco Kin(CO2)仍存在差异,如水稻RubiscoKin(CO2)一般为12lxmo1.L~,而喜温红藻(Galdieriapartita)的K(co2)仅为6.6~tmol?L一,这种喜温红藻366植物生理学通讯第43卷第2期,2007年4月RubiscoK~(CO)值在当时被认为是所有Rubisco中最小的(Uemura等1997).Ezaki等(1999)研究菌的结果表明,Tk—RubiSCO具有更强的羧化能力,在CO,饱和,90℃高温条件下,Tk.Rubisco同化CO,的速率可达19.8x10.nmo1.mg(酶蛋白)? min~.Rubisco羧化与氧化反应的活力比按/V o=()/(V oKc)([CO:】/【O2】)计算.其中,,分别表示羧化和氧化反应的最大反应速率;&,K. 分别代表羧化和氧化反应的米氏常数;【CO】和【O】为气体浓度;(c.)/(.c)称特性因子(specificityfactor),即羧化/氧化值(Q值)(Uemura等1997;Parry等2003).一定种类Rubisco的Q值为一常数,但不同来源的RubiscoQ值有较大差异(表1).绿色高等植物Rubisco的Q值一般在90~95之间;蓝藻的Q值也有35~40;细菌的Q值一般在9~45之间.从喜温红藻中发现的I型RubiscoQ值高达238(25℃),是高等植物Q值的2.5倍,说明其具有极强的固定CO:的能力(Uemura等1997).更为重要的是喜温红藻本身就属于植物,这对从亚基水平上改变Rubisco动力学性质及Q值,提高植物光合效率有重大意义. Tk—Rubisco存在更高的Q值且随温度的升高而上升,在50℃时为70,70℃时上升至250,90℃时则达到最大值310(Ezaki等1999).与此相反,喜温红藻的RubiscoQ值随着温度的升高反而降低(Uemura等1997).Watson和Tabita(1999)测定詹氏甲烷球菌Q值的结果表明,其羧化能力极差,Q值只有0.5,是所有Rubisco中Q值最低的.5Rubisco的功能已经证实,O是羧化酶反应的竞争性抑制剂;同样CO是加氧酶反应的竞争性抑制剂.因此,Rubisco处于光合碳还原(光合作用)和光合碳氧化(光呼吸)2个方向相反但又相互连锁的循环反应的交叉点上.当co#o:的浓度比值较高时,促进Rubisco催化的羧化反应.羧化反应一般分为烯醇化,羧化,水合,C—C键断裂,质子化5个阶段(Lundqvist和Schneider1991;Li等2005). Rubisco催化游离的CO,共价结合到底物RuBP上,进而生成两分子的3一磷酸甘油酸(3一phosphoglyc—ericacid,PGA),推动C3-PCR循环.当CO2/O2的浓度比值较低时,促进Rubisco催化的加氧反应,RuBP即裂解产生一分子的磷酸乙醇酸和一分子的PGA,前者进一步分解成乙醇酸和磷酸,参与绿色植物的光呼吸循环.研究初期,人们认为Rubisco的功能主要集中在光合碳同化及光呼吸过程中.然而随着各种不同类型RubiSCO的陆续发现和研究的深入, Rubisco的功能呈现出多样化(表1).詹氏甲烷球菌及其它产甲烷古生菌中的III型RubiSCO参与PRPP—RuBP—PGA途径(Ashida等称之为RuPP通路) (Finn和Tabita2004;Ashida等2005).在这一途径中RuBP的合成前体是1一焦磷酸.5一磷酸.核糖(5一phospho—D—ribose一1一pyrophosphate,PRPP),脱磷酸后经NAD氧化生成RuBP,接着在Rubisco的催化下与CO,结合生成PGA.RLP一般无羧化酶活性,但它能催化2,3一二酮基一5一甲硫戊基.1一磷酸(2,3一diketo一5一methythio—pentyl一1一phosphate,DK—MTP.1一P)的烯醇化反应,与硫代谢密切相关(Ashida等2003;Li等2005;Carr6一Mlouka等2006).枯草芽孢杆菌RLP在腺苷蛋氨酸补救合成途径中作为DK—MTP一1一P烯醇化酶发挥作用(Ashida等2003,2005).在其r/p突变株中导入深红红螺菌的II型Rubisco后,该菌株竟然可恢复生长,说明光合类型的Rubisco可能仍然保持着作为一腺苷蛋氨酸补救合成途径中DK—MTP.1一P烯醇化酶的功能.也有人认为是由于RubiSCO对底物存在一定范围的适应(Li等2005).绿色硫细菌RLP与硫代谢和氧化应激(oxidativestress)有关(Hanson和Tabita2001).铜绿微囊藻PCC7806RLP在硫代谢过程中也起一定作用.半定量RT—PCR的结果显示,在硫缺乏的情况下,吐Ⅳ的转录是正常状况(硫充足)下的22倍(Carte—Mlouka等2006).Rubisco作为植物叶中的主要含氮有机物之一,必然与植物对氮的吸收,利用及循环有关.研究不同种类C植物氮利用的情况表明,较高的Rubisco,是NADP苹果酸酶类型比NAD苹果酸酶类型具有更高的氮利用率的主要原因(Ghannoum等2005).Rubisco与植物雄性不育也有一定的联系.刘祚昌等(1983)研究玉米,高粱,水稻,小植物生理学通讯第43卷第2期,2007年4月367 麦和烟草等作物的细胞质雄性不育系Rubisco的结果表明,其活性均高于相应的保持系.水稻光周期敏感核不育农垦58S经长光照及红光间断暗周期处理表现为雄性不育,其Rubisco的活性明显低于可育状态(夏凯等1989).Schwender等(2004)最新发现甘蓝型油菜(Brassicanapus)中Rubisco参与植物中碳转化为油的代谢通道,此通道可导致碳作为油贮存的效率达到最大.6结语有人曾将红藻中高效率Rubisco酶的小亚基基因转入植物叶绿体中并得到表达,但表达出的小亚基却不能与植物本身的大亚基组装成全酶(Whitney和Andrews2001).这可能是由于小亚基存在特异性的修饰,或者协助组装的分子伴侣无法识别异源亚基.要解决上述问题尚待深入研究Rubisco的组装机制.众多的研究表明,低等藻类和古生菌体内存在不同类型Rubisco.Lonsdale等(1983)从玉米(Zeamays)线粒体中分离到一个与Rubisco大亚基同源的基因,此基因在大肠杆菌中表达合成的分子量为2lkDa的蛋白能与小麦的Rubisco抗体反应.如何科学地解释Rubisco的共存现象,共存现象与Rubisco的分子进化是否存在某种内在联系,植物体内是否也存在多拷贝的Rubisco基因,均待进一步研究.微生物Rubisco的研究大大扩展了人们对Rubisco类型及功能的认识.序列同源性比较分析的结果显示,III型和Ⅳ型Rubisco比I型及II型更原始;枯草芽孢杆菌及铜绿微囊藻PCC7806RLP功能及突变株的研究结果表明,m型与Ⅳ型Rubisco中可能存在Rubisco的原始形式,或者说它们与Rubisco的原始形式在结构及功能上有更多的相似性.根据研究枯草芽孢杆菌RLP的结果,Ashida等(2003,2005)提出一条Rubisco可能的进化路线,认为光合类型Rubisco正是由枯草芽孢杆菌RLP演变而来的.以上研究和发现并未完全解决Rubisco的分子进化问题,但却为人们指明了方向,即通过研究和分析微生物Rubisco的结构和功能有可能部分或完全揭示Rubisco的分子进化历程.参考文献陈为钧,赵贵文,顾月华(1999).RubisCO的研究进展.生物化学与生物物理进展,26(5):433-.-436刘祚昌,李继耕,罗会馨,陈福太(1983).二磷酸核酮糖羧化酶与细胞质雄性不育性的研究.遗传,10(1):362吕红,周集体,王竞,安利佳(2003).原核生物Rubisco的研究进展.微生物学通报,30(2):82—85夏凯,肖翊华,刘文芳(1989).湖北光敏感核不育水稻光敏感期叶片中ATP含量与RuBPcase活力的分析.杂交水稻,(4): 412.30熊晓然,陈蔚梅,冯胜彦,郭明雄,艾建宇,吴斌(2003).植物Rubisco 活性中心的模拟分析.中国生物化学与分子生物,19(4):493--498AshidaH,DanchinA,Y okotaA(2005).Wasphotosynthetic RuBisCOrecruitedbyacquisitiveevolutionfromRuBisCO—likeproteinsinvolvedinsulfurmetabolism?ResMicrobiol,l56:6lll8AshidaH,SaitoY,KojimaC,KobayashiK,OgasawaraN,Y okotaA(2003).AfunctionallinkbetweenRuBisCO—likeproton ofBacillusandphot0syntheticRuBisCo.Science,302:286~290BakerSH,JinS,AldrichHC,HowardGT,ShivelyJM(1998). InsertionmutationoftheformIcbbLgeneencodingribulose bisphosphatecarboxylase,oxygenase(RuBisCO)in ThiobacillusneapolitanusresultsinexpressionofformII RuBisCO,lossofcarboxysomes,andanincreasedCO2re—quirementforgrowth.JBacteriol,l80(16):4133—4l39Carr6一MloukaA,M6jeanA,QuillardetP,AshidaH,SaitoY, Y okotaA,CallebautI,SekowskaA,DittmannE,BouchierC etal(2006).AnewRubisco—likeproteincoexistswithaphot0syntheticRubiscointheplanktoniccyanobacteria Microcystis.JBiolChem,281:24462~24471ChenZH.WalkerRP,AchesonRM,LeegoodRC(2002). Phosphoenolpyruvatecarboxykinaseassayedatphysiologi—calconcentrationsofmetalionshasahighaffinityforCO2. PlantPhysio1.128:l60~164EnglishRS,WilliamsCA,LorbachSC.ShivelyJM(1992).Two formsofribulose—l.5-bisphosphatecarb0xylase,oxygenase fromthiobacillusdenitrificans.FEMSMicrobiolLett,94: lll—ll9EzakiS,MaedaN,KishimotoT,AtomiH.ImanakaT(1999). Presenceofastructurallynoveltyperibulose—bisphosphate carboxylase,0xygenaseinthehyperthermophilicarchaeon, Pyrococcus七Dd口七口r口P,lfK0D1.JBiolChem.274: 5078~5082FinnMW,TabitaFR(2004).ModifiedpathwaytOsynthesize ribulosel,5-bisphosphateinmethanogenicarchaea.J Bacteriol,186"6360,45366Ghannoum0,EvansJR,ChowWS,AndrewsTJ,ConroyJP.yon CaemmererS(2005).FasterRubiscoisthekeytosuperior nitrogen?-useefficiencyinNADP?-malicenzymerelativetO NAD—malicenzymeC4grasses.PlantPhysiol,l37:638,650 HansonTE,TabitaFR(2001).Adbulose—l,5-bisphosphatecar—368植物生理学通讯第43卷第2期,2007年4月boxylase,oxygenase(RubisCO)-likeproteinfromchlorobium tepidumthatisinvolvedwithsulfurmetabolismandthe responsetooxidativestress.ProcNatlAcadSciUSA,98:4397-4402HansonTE,TabitaFR(2003).Insightsintothestressresponseand sulfurmetabolismrevealedbyproteomeanalysisofa chlorobiumtepidummutantlackingtheRubisco-likeprotein. PhotosynthRes,78:23l-248KarlinS,MrazekJ(2000).Predictedhighlyexpressedgenesof diverseprokaryoticgenomes.JBacteriol,l82:5238-5250 KitanoK,MaedaN,FukuiT,AtomiH,ImanakaT,MildK(2001). Crystalstructureofanovel??typearchaealRubiscowithpen-- tagonalsymmetry.Structure.9:473,48lKlenkHP,ClaytonRA,TombJF,WhiteO,NelsonKE,Ketchum KA,DodsonRJ,GwinnM,HickeyEK,PetersonJDetal (1997).Thecompletegenomesequenceofthehyperthermop- hilic,sulphfate-reducingarchaeonArchaeoglobuslgidus. Nature.390:364—370LarimerFW,ChainP,HauserL,LamerdinJ,MalfhttiS,DoL,Land ML,PelletierDA,BeattyJT,LangASetal(2004).Corn—pletegenomesequenceofthemetabolicallyversatilephoto—syntheticbacteriumRhodopseudomonaspalustris.Nat Biotechno1.22:55lLiH,SawayaMR,TabitaFR,EisenbergD(2005).Crystalstructure ofaRuBisCO—likeproteinfromthegreensulfurbacterium Chlorobiumtepidum.Structure.13:779-789LiaoH,ZhouJY,DuLF,ZhangNH,WuL(2004).Ribulosel,5一bisphosphatecarboxylase,oxygenaseinArchaea.Natural ProductResDevelop,16:569-574LonsdaleDM,HodgeTP,HoweCJ,StemDB(1983).Maize mitochondrialDNAcontainsasequencehomologoustothe ribulose—l,5-bisphosphatecarboxylaselargesubunitgeneof chloroplastDNA.Cel1.34:l007-10l4LundqvistT,SchneiderG(1991).Crystalstructureofactivated ribulose—l,5-bisphosphatecarboxylasecomplexedwithits substrate,ribulose—l,5-bisphosphate.JBiolChem.266:l2604-l26llMaedaN,KanaiT,AtomiH,ImanakaT(2002).Theunique pentagonalstructureofanarchaealRubiscoisessentialfor itshighthermostability.JBiolChem.277:3l656-3l662 NassouryN,FritzL,MorseD(2001).Circadianchangesin ribulose-l,5-bisphosphatecarboxylase,oxygenasedistribu—tioninsideindividualchloroplastscanaccountfortherhythm indinoflagellatecarbonfixation.PlantCell,13:923-934ParryMAJ,AndralojcPJ,MitchellRAC,MadgwickPJ,KeysAJ (2003).ManipulationofRubisco:theamount,activity,func—tionandregulation.JExpBut,54:1321-1333RowanR,WhitneySM,FowlerA,Y ellowleesD(1996).Rubisco inmarinesymbioticdinonagellates:formIIenzymesineu- karyoticoxygenicphototrophsencodedbyanuclear multigenefamily.PlantCell,8:539-553SageRF(2002).V ariationinthetofRubiscoinC3andC4plantsandsomeimplicationsforphotosyntheticperformanceat highandlowtemperature.JExpBut,53:609-620 SchwenderJ,GoffmanF,OhlroggeJB,Shachar-HillY(2004). RubiscowithouttheCalvincycleimprovesthecarboneffi- ciencyofdevelopinggreenseeds.Nature,432:779-782 SpreitzerRJ(2003).Roleofthesmallsubunitofribulose—l,5- bisphosphatecarboxylaseloxygenase.ArchBiochemBiophys, 4l4:l4l~l49TabitaFR(1995).Thebiochemistryandmetabolicregulationof carbonmetabolismandCO2fixationinpurplebacteria.In: BlankenshipRE,MadiganMT,BauerCE(eds).Anoxygenic PhotosyntheticBacteria.Amsterdam:KluwerAcademic Publishers,885--914UemuraK,Anwaruzzaman,MiyachiS,Y okotaA(1997).Ribu—lose一1,5-bisphosphatecarboxylase/oxygenasefromthermo- philicredalgaewithastrongspecificityforCO2fixation. BiochemBiophysResCommun,232:568-571 WatsonGMF,YuJP,TabitaFR(1999).Unusualribulose1.5一bisphosphatecarboxylase,oxygenaseofanoxicArchaea.J Bacterio1.18l:l569-l575WhitneySM,AndrewsTJ(2001).Thegenefortheribulose一1,5. bisphosphatecarboxylaseloxygenase(Rubisco)smallsubunit relocatedtotheplastidgenomeoftobaccodirectsthesyn—thesisofsmallsubunitsthatassembleintoRubisco.PlantCell,13:193-205Y oshizawaY,ToyodaK,AraiH,IshiiM,IgarashiY(2004).CO2一responsiveexpressionandgeneorganizationofthreeribu-lose-1,5-bisphosphatecarboxylase,oxygenaseenzymesand carboxysomesinhydrogenovibriomarinusstrainMH—ll0.JBacterio1.186:5685—,569l。
【国家自然科学基金】_羧化效率_基金支持热词逐年推荐_【万方软件创新助手】_20140801
科研热词 推荐指数 光合特性 3 水分胁迫 2 co2浓度 2 黄瓜 1 高温 1 高浓度臭氧 1 高大气co2浓度 1 非气孔限制 1 金花茶 1 荷叶白 1 胡桃楸 1 耐热性 1 羊草草原 1 生理生态因子 1 生理特性 1 玉米 1 温度胁迫 1 油松 1 水分利用率 1 氮素 1 气孔导度 1 次生林 1 桑树 1 日动态 1 抗虫棉 1 弱光胁迫 1 弱光 1 幼苗 1 小麦 1 季节动态 1 可塑性 1 光适应 1 光能传递分配 1 光强 1 光响应 1 光合速率 1 光合气体交换 1 光合效率 1 光合参数 1 光合作用 1 丁香 1 co_2响应 1 5–二磷酸核酮糖羧化/加氧酶(rubisco) 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40
科研热词 推荐指数 光合作用 5 叶绿素荧光 3 光合特性 2 黄瓜幼苗 1 高浓度co2 1 频率分布 1 雾凉季 1 长期 1 辣椒 1 薇甘菊(mikania micrantha) 1 蒸腾速率 1 耐旱性 1 稳定碳同位素 1 种间比较 1 碳同位素分辨率 1 玉米 1 热带经济植物 1 水分胁迫 1 水分利用效率 1 气孔导度 1 植物生长调节剂dcpta和dta-6 1 植物生长调节剂 1 植物 1 林线 1 摩西球囊霉 1 抗氧化酶 1 干旱 1 大豆 1 复合群落 1 呼吸速率 1 净光合速率 1 光合速率 1 光合参数 1 低温 1 优势种 1 亚高山 1 三叶鬼针草 1 no3-胁迫 1 dta-6 1 dcpta 1
53 54 55 56 57 58 59 60 61 62 63 64
2016届高三下学期第二次全国大联考(浙江卷)化学试卷(DOC)

2016年第二次全国大联考【浙江卷】理科综合化学试题绝密★启用前注意事项:1 .本试卷分第I卷(选择题)和第n卷(非选择题)两部分。
满分300分,考试时间150分钟。
2 •答题前考生务必用0.5毫米黑色墨水签字笔填写好自己的姓名、班级、考号等信息3.考试作答时,请将答案正确填写在答题卡上。
第一卷每小题选出答案后,用2B铅笔把答题卡上对应题目的答案标号涂黑;第H卷请用直径0.5毫米的黑色墨水签字笔在答题卡上各题的答题区域内作答,超出答题区域书写的答案无效,在试题卷、草稿纸上作答无效。
4.以下数据可供解题时参考:可能用到的相对原子质量:H 1 C 12 N 14 O 16 S 32 Cl 35.5 Na 23 Mg 24 Al 27第I卷选择题(共42 分)、单项选择题:本题包括7小题,每小题6分,共计42分。
每小题只有一个选项符合题1•下列物质运输的过程中不需要消耗ATP的是()A. 胰岛素的分泌过程B.神经纤维受到刺激后Na+的内流C . 蛙的红细胞吸收葡萄糖D.神经纤维未受刺激时,K+运进细胞意。
2•下列有关人体免疫的说法中,正确的是()A .当机体免疫功能不足或缺乏时,可引起过敏反应B •初次接触花粉时可能发生过敏反应C.特异性免疫中发挥作用的主要细胞是淋巴细胞,衰老和破损细胞的清除需免疫系统参与D •一种抗原只能与一种抗体或浆细胞结合,发生特异性免疫3.下列说法正确的是()A.转录的时候RNA聚合酶是沿着整条DNA长链进行B.RNA聚合酶与DNA分子某一启动部位结合后只导致一个基因的双螺旋片段解开C.基因表达的产物是RNA和蛋白质B. 生物甲与生物乙之间的关系是竞争C. 无生物乙时,生物甲的种群数量呈 J 型增长D. 无生物乙时,1〜3周生物甲种群出生率大于死亡率D.生物界的遗传密码是完全统一的4•某科研小组利用赤霉素和乙烯利开展了杜鹃开花数量和花期的研究,实验结果如下图,下 列 叙述错误的是( ) 4302010♦ GA 处理’花期 亠处理.开花数最 o ET 魁理,花期 ° ET 处理,开花数量 -160 :120IV ' 80 f -40 » 0 50100 200 300 400GA.巩浓度(W g • r 1)A. 赤霉素和乙烯利都属于植物生长调节物质B. 乙烯利不能延长杜鹃的花期C. 如果继续增大赤霉素的浓度后导致杜鹃的开花数量下降, 开花数量的作用说明高浓度赤霉素具有抑制 D.浓度为200mg/L 时,赤霉素对杜鹃开花数量的影响效果比乙烯利明显 5•已知常染色体隐性遗传病 -白化病由基因A 和a 控制,色盲症由基因 B 和b 控制。
【国家自然科学基金】_光合作用的光抑制_基金支持热词逐年推荐_【万方软件创新助手】_20140801
科研热词 贝壳堤岛 光合作用 阈值效应 水分利用效率 气体交换参数 叶绿素荧光参数 光合生产力 遮荫 迁地 荔枝 环剥 海拔 枝梢生长 宽刺蔷薇 土壤水分 叶形态 功能组 净光合速率 光诱导 光环境 光响应模型 光合参数 光保护
推荐指数 3 3 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
推荐指数 3 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 光抑制 光合作用 高温胁迫 非光化学猝灭 花青素 热带雨林树苗 形态 太阳辐射 叶绿素荧光 冬小麦 光饱和项 光适应 光系统ii 光破坏防御 光抑制项 光响应修正模型 光合特性
推荐指数 3 3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
科研热词 光合作用 叶绿素荧光 光抑制 陆地棉 铝胁迫 螺旋藻 藻类植物 藓类植物(土生扭口藓) 落叶阔叶树 荞麦 红毛菜 磷 生物结皮 环境因子 猫儿山 温度效应 海岛棉 森林生物学 日变化 悬钩子属 干出 常绿阔叶树 哀牢山 呼吸速率 叶绿素a荧光 叶片运动 净光合速率 光合能力 光合作用季节变化
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42
【国家自然科学基金】_条件真度_基金支持热词逐年推荐_【万方软件创新助手】_20140801
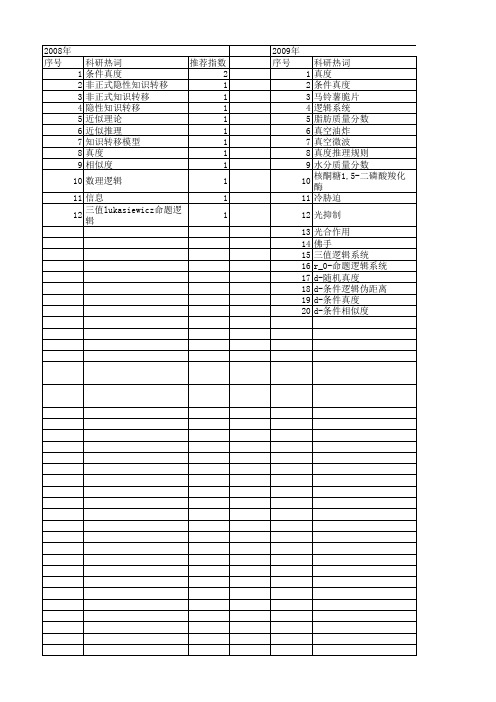
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 蕴涵算子 公式的条件真度 伴随对 三角模算子 d-条件真度 逻辑系统g3 近似推理 浮游动物
推荐指数 2 2 2 2 2 1 1 1 1 模糊命题系统g(o)del和l* 1 小黄鱼 1 吕泗渔场 1 d-真度 1 d-条件逻辑度量空间 1 d-条件发散度 1 d-条件伪距离 1 d-发散度 1
科研热词 真度 条件真度 马铃薯脆片 逻辑系统 脂肪质量分数 真空油炸 真空微波 真度推理规则 水分质量分数 核酮糖1,5-二磷酸羧化酶 冷胁迫 光抑制 光合作用 佛手 三值逻辑系统 r_0-命题逻辑系统 d-随机真度 d-条件逻辑伪距离 d-条件真度 d-条件相似度
推荐指数 3 3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 11 12
科研热词 推荐指数 条件真度 2 非正式隐性知识转移 1 非正式知识转移 1 隐性知识转移 1 近似理论 1 近似推理 1 知识转移模型 1 真度 1 相似度 1 数理逻辑 1 信息 1 三值lukasiewicz命题逻辑 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
科研热词 推荐指数 近似推理 2 高羊茅 1 计量逻辑 1 草坪质量 1 育种 1 综合抗性 1 磁化率 1 真度 1 理论和谐度 1 沉积物 1 条件随机逻辑度量空间 1 条件随机真度 1 条件随机相似度 1 条件真度 1 条件实验 1 有机碳同位素 1 无性系 1 孢粉 1 子一代 1 地球化学 1 古气候 1 华南 1 二叠纪-三叠纪之交 1 sedimentary rock 1 organic carbon isotopes 1 conditional experiments 1
【国家自然科学基金】_生态分化_基金支持热词逐年推荐_【万方软件创新助手】_20140801
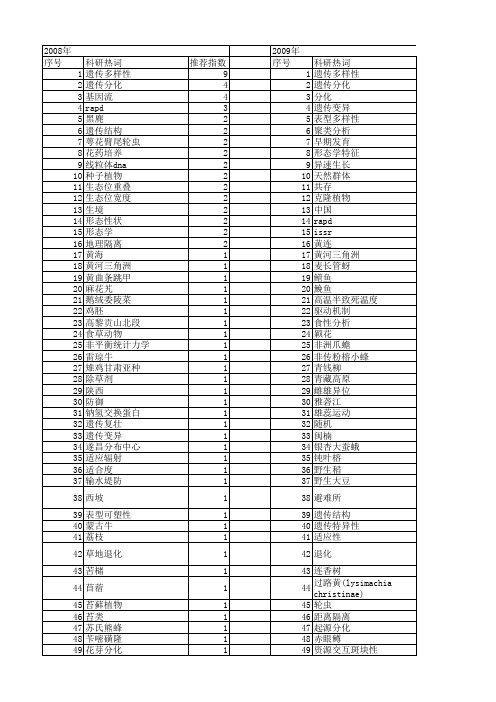
推荐指数 9 4 4 3 2 2 2 2 2 2 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
表达模式 表达序列标签 表型可塑性 蜜蜂 蚯蚓 藓类 蒙古羊系统 萼花臂尾轮虫 萍乡肉红鲫 菜粉蝶 菊头蝠 荔枝花芽分化 芳香化酶 花粉呈现理论 自交亲和性 臂尾轮属 能量投资 胡杨 群落结构 群落 群体分化 维管束植物 经济昆虫 细胞色素b基因 细胞色素b 繁殖分配 紫茎泽兰 紫外辐射 系统发育分析 系统关系 精巢 籼-粳稻鉴定 等位酶变异 竞争等级 竞争的重要性 空间自相关 空间结构 稀有鮈鲫 种群遗传分化 种群结构 种群增长 种群 种子植物 种子性状 神农架 社会性行为 短花针茅草原 省际边缘区域 盆西平原麦区 生长 生物结皮 生物学特征 生物学 生物型
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106
107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160
【国家自然科学基金】_附生植物_基金支持热词逐年推荐_【万方软件创新助手】_20140730
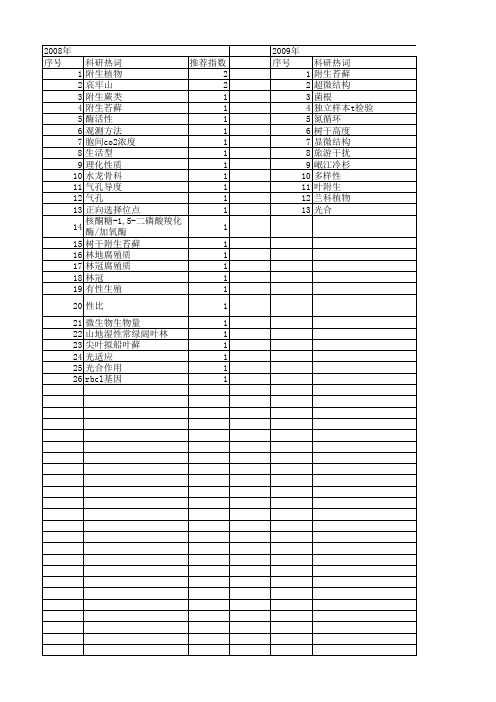
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
Байду номын сангаас
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
科研热词 推荐指数 附生植物 2 哀牢山 2 附生蕨类 1 附生苔藓 1 酶活性 1 观测方法 1 胞间co2浓度 1 生活型 1 理化性质 1 水龙骨科 1 气孔导度 1 气孔 1 正向选择位点 1 核酮糖-1,5-二磷酸羧化酶/加氧酶1 树干附生苔藓 1 林地腐殖质 1 林冠腐殖质 1 林冠 1 有性生殖 1 性比 1 微生物生物量 1 山地湿性常绿阔叶林 1 尖叶拟船叶藓 1 光适应 1 光合作用 1 rbcl基因 1
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13
科研热词 附生苔藓 超微结构 菌根 独立样本t检验 氮循环 树干高度 显微结构 旅游干扰 岷江冷杉 多样性 叶附生 兰科植物 光合
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42
花魔芋核酮糖-1,5-二磷酸羧化酶活化酶RCA家族基因的全基因组鉴定和功能分析

花魔芋核酮糖-1,5-二磷酸羧化酶活化酶RCA家族基因的全基因组鉴定和功能分析褚洪龙;郑文广;李竹梅【期刊名称】《曲靖师范学院学报》【年(卷),期】2024(43)3【摘要】花魔芋(Amorphophallus konjac)是中国目前主要种植的魔芋品种之一,具有重要经济价值.核酮糖-1,5-二磷酸羧化酶(Rubisco)是绿色植物进行光合作用暗反应中碳同化的关键酶.研究表明,Rubisco必须经Rubisco活化酶(Rubisco activase,RCA)激活,才能保持高催化活性,在植物光合作用中起着重要作用.利用生物信息学方法,在全基因组水平鉴定了花魔芋RCA家族基因,并对其理化性质、基因结构特征、蛋白保守性、亚细胞定位和系统进化等进行了初步分析.结果表明,该家族基因具有4个成员,等电点(Isoelectric point,PI)范围为5.38至8.72,亲水性平均值为负值.基因结构分析该家族基因成员含5~12个外显子.亚细胞定位预测显示,4个成员均定位在叶绿体.染色体定位分析发现,该家族基因成员分布在花魔芋26条染色体中的2条上,其中AkRCA1和AkRCA2位于同一条染色体上.启动子区域顺式作用元件分析表明,该家族基因成员启动子区域含有参与光响应、茉莉酮酸甲酯反应、干旱诱导、缺氧特异性诱导以及胁迫相关的作用元件等.进一步进行蛋白多序列比对发现,该家族基因成员均含有AAA+家族保守区以及2个ATP结合域,保守性较高.本研究为后续RCA功能研究及魔芋种质改良奠定基础.【总页数】10页(P31-40)【作者】褚洪龙;郑文广;李竹梅【作者单位】曲靖师范学院生物资源与食品工程学院【正文语种】中文【中图分类】Q943.2;S632.3【相关文献】1.沼泽红假单胞菌(Rhodopseudomonas palustris)核酮糖-1,5-二磷酸羧化酶/氧化酶基因克隆及其在大肠杆菌中的表达2.巴夫杜氏藻1,5-二磷酸核酮糖羧化酶/加氧酶小亚基基因的克隆和分析3.糠椴核酮糖-1,5-二磷酸羧化酶/加氧酶基因生物信息学分析4.多能硫杆菌1,5-二磷酸核酮糖羧化酶/加氧酶的酶学特征及基因检测5.桑树1,5-二磷酸核酮糖羧化酶活化酶基因cDNA片段的克隆及原核表达与植物反义表达载体的构建因版权原因,仅展示原文概要,查看原文内容请购买。
不同叶位鸡蛋果叶片中核酮糖—1,5—二磷酸羧化酶...

不同叶位鸡蛋果叶片中核酮糖—1,5—二磷酸羧化酶...
宁正祥;李明启
【期刊名称】《植物生理学通讯》
【年(卷),期】1991(027)004
【摘要】随着叶龄的增加,鸡蛋果Ru BP羧化酶/加氧酶的V_(max)(CO_2)值明显减小,K_m(O_2)值提高;K_m(CO_2)和V_(max)(O_2)值则保持相对的稳定。
Ru BP 羧化酶活性和Ru BP加氧酶活性均随叶龄增加而下降,但前者下降的速度高于后者,致使羧化/加氧此值也随叶龄增加而减小。
【总页数】4页(P263-266)
【作者】宁正祥;李明启
【作者单位】不详;不详
【正文语种】中文
【中图分类】Q949.759.5
【相关文献】
1.芍药不同品种1,5-二磷酸核酮糖羧化酶活性差异研究 [J], 马丽茹;高健洲;刘燕
2.不同生境芦苇1,5-二磷酸核酮糖羧化酶活性与二氧化碳固定关系 [J],
3.不同核质组合水稻核酮糖—1,5—二磷酸羧化酶/加氧酶... [J], 宁正祥;万邦惠
4.草酸对氧化胁迫下水稻叶片中核酮糖-1,5-二磷酸羧化酶/加氧酶降解的保护作用(英文) [J], 彭新湘;刘拥海;李明启
5.水稻叶片中过氧化氢与核酮糖-1,5-二磷酸羧化酶/加氧酶降解的关系 [J], 刘拥海;彭新湘;李明启
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
科研热词 小麦 光合作用 质核互作型雄性不育 蛋白质组学 臭氧(o3) 纳升级液相色谱串联质谱 盐胁迫 核酮糖1,5-二磷酸羧化酶 差异蛋白质组学 小麦叶片 基质辅助激光解吸飞行时间质谱 叶绿素荧光 叶片衰老 可溶性蛋白 双向凝胶电泳 冷胁迫 内肽酶 光抑制 佛手 二维液相色谱 不结球白菜 rubisco
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
科研热词 推荐指数 诱导系统耐性(ist) 1 解剖结构 1 行距配置 1 蜡质芽胞杆菌 1 耐旱性 1 穗位叶 1 种植密度 1 生防菌 1 玉米 1 烟草叶片 1 水分胁迫 1 核酮糖1 1 抗氧化酶活性 1 应答蛋白 1 小麦 1 光质 1 光合特性 1 lc-ms/ms 1 5-二磷酸羧化酶/加氧酶 1
2014年 序号 1 2 3 4 5 6 7 8 9 10 11
2014年 科研热词 推荐指数 蛋白质 2 木薯 2 叶绿素荧光 2 稳定同位素探针技术 1 碳循环 1 核酮糖-1,5-二磷酸羧化酶 1 同化碳 1 加氧酶 1 光合效率 1 低温胁迫 1 co2同化 1
推荐指数 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 11年 科研热词 推荐指数 序号 野大麦 1 1 药用植物 1 2 缘管浒苔 1 3 盐胁迫 1 4 物种鉴定 1 5 正选择位点 1 6 核酮糖1,5-二磷酸羧化酶/加氧酶大亚基基因(rbcl基因) 1 7 核酮糖-1,5-二磷酸羧化酶/加氧酶1 8 放松分子钟模型 1 9 序列分析 1 10 前导序列 1 11 分类 1 12 凤尾蕨科 1 13 rbcl基因 1 14 dna条形码 1 15 cdna未端快速扩增 1 16 cdna 1 17 1,5-二磷酸核酮糖羧化酶/加氧酶小亚基 1 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
2011年 科研热词 推荐指数 植物表达载体 2 叶绿体定位 2 黄瓜叶片 1 野大麦 1 酵母双杂交系统 1 通路克隆技术 1 通路克隆(gateway)技术 1 适应性进化 1 蛋白互作 1 聚乙二醇 1 盐胁迫 1 温室效应 1 核酮糖-1,5-二磷酸羧化酶/加氧酶1 拟南芥 1 微生物 1 序列分析 1 小麦 1 固碳 1 双向电泳 1 分子生态 1 凤尾蕨科 1 冷胁迫 1 共进化 1 入门载体 1 光诱导型启动子 1 光合作用 1 信号转导 1 二磷酸核酮糖羧化酶小亚基3c启动子 1 rubisco相互作用蛋白质 1 rbcl基因 1 pen-l4*-prbcs-*t-gfp-l3*入门载体 1 erf 1 1,5-二磷酸核酮糖羧化酶/加氧酶小亚基 1 1,5-二磷酸核酮糖羧化酶/加氧酶 1 1,5 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
科研热词 推荐指数 花生 2 碳代谢酶 2 氮代谢酶 2 双向电泳 2 光合作用 2 蛋白质 1 葡萄卷叶病毒 1 葡萄 1 菌核病抗性 1 脂肪 1 肽质量指纹图谱 1 羧化效率 1 缘管浒苔 1 细胞周期 1 稻瘟菌激发子 1 直角双曲线修正模型 1 甲醇 1 油菜 1 水稻 1 棉花 1 核酮糖-1,5-二磷酸羧化酶/氧化酶1 果实品质 1 强电场辐射 1 多效唑 1 土壤含水量 1 品质类型 1 品质 1 光合有效辐射强度 1 光合日变化 1 产量 1 rubpcase 1 rubisco 1 ppdk 1 polarella glacialis 1 pcna 1 c4途径 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
科研热词 推荐指数 盐藻 2 rbcs 2 水龙骨科 1 水稻(oryza sativa l.) 1 正向选择位点 1 核酮糖-1,5-二磷酸羧化酶/加氧酶1 启动子 1 化感潜力 1 加氧酶 1 光合生理 1 低钾胁迫 1 rbcl基因 1 race 1 1,5-二磷酸核酮糖羧化酶/加氧酶 1 1,5-二磷酸核酮糖羧化酶 1