海洋学(二)--耿晨曦
海洋科学专业本科课程设置

海洋科学专业本科课程设置引言海洋科学是研究海洋及其相关的自然和人文科学问题的学科。
本科教育是培养海洋科学人才的重要环节,合理设置的课程体系是保证学生全面学习和掌握相关知识的基础。
本文将介绍海洋科学专业本科课程设置的1200字概述。
一、基础课程1.自然科学基础课程:–高等数学–高等物理学(海洋物理学)–概率论与数理统计–计算机科学基础2.海洋科学基础课程:–海洋学导论–海洋地质学–海洋化学–海洋生物学二、专业课程1.海洋物理学专业课程:–海洋动力学–海洋气候学–海洋遥感与海洋气象学–海洋波浪与潮汐学2.海洋地质学专业课程:–海洋地球化学–海洋地球物理学–海洋沉积学–海洋地质资源与勘探3.海洋化学专业课程:–海洋化学动力学–海洋生物地球化学–海洋环境化学–海洋有机地球化学4.海洋生物学专业课程:–海洋生态学–海洋生物多样性–海洋生物进化与分类学–海洋生物遗传学三、专业实践1.实习:–专业实习1(海洋采样与观测)–专业实习2(海洋研究船实习)–专业实习3(海洋实验室操作)2.实验室:–海洋科学实验室课程3.毕业论文:–学生需选择海洋科学专业相关研究课题,并完成毕业论文四、选修课程1.海洋保护与管理2.鱼类学3.海洋能源学4.海洋遗址考古学结论以上是海洋科学专业本科课程设置的1200字概述。
这些课程涵盖了海洋科学各个专业的基础知识和专业知识,旨在培养具备科学素养、创新能力和实践能力的海洋科学人才。
我国深海Argo区域观测网成功完成“深海玄武”浮标首次批量布放

我国深海Argo区域观测网成功完成“深海玄武”浮标首次批
量布放
陈朝晖;任翀;刘增宏;熊海霞;张林林;高志远
【期刊名称】《海洋学研究》
【年(卷),期】2024(42)1
【摘要】2023年12月,由中国海洋大学牵头,崂山实验室、自然资源部第二海洋研究所、中国科学院海洋研究所共同参与的“深海Argo区域观测网建设”在西太平洋菲律宾海盆顺利完成首批6000 m级国产“深海玄武”浮标的布放。
自12月9日起,经质量控制的深海温度和盐度剖面观测数据实时提交至全球Argo资料中心(GDAC),参与国际共享与交换。
这是我国首次批量布放国产深海Argo浮标,标志着我国在深海Argo区域观测网建设上迈出了重要的一步。
【总页数】2页(P117-118)
【作者】陈朝晖;任翀;刘增宏;熊海霞;张林林;高志远
【作者单位】中国海洋大学物理海洋教育部重点实验室;崂山实验室;自然资源部第二海洋研究所;卫星海洋环境动力学国家重点实验室;中国科学院海洋研究所
【正文语种】中文
【中图分类】P71
【相关文献】
1.我国首次完成深海富钴结壳采掘试验
2.我国首次在西太平洋布放深海微生物原位富集系统
3.我国首次完成万米级深海科考
4.一句话新闻
5.《干旱环境监测》2020年第36卷总目次
因版权原因,仅展示原文概要,查看原文内容请购买。
物理海洋学系
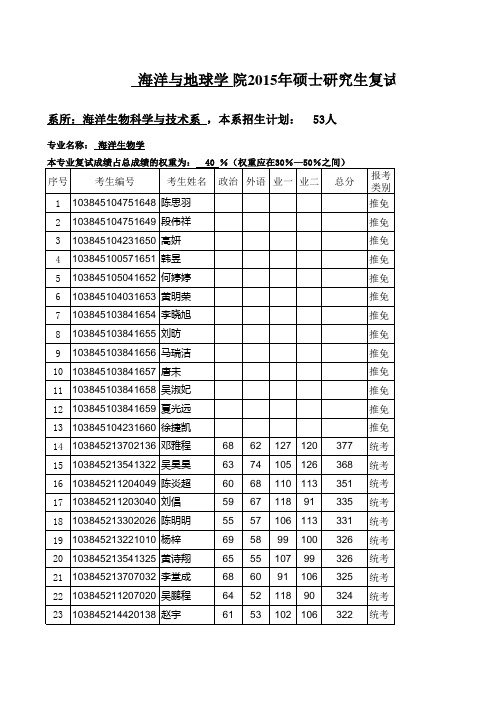
107 107 102 100 91 96 99 105 103 97 96 93 90 92
40 %(权重应在30%—50%之间)
序号 1 2 3 4
考生编号
考生姓名
政治 外语 业一 业二
总分
报考 类别 推免 推免
103845103861666 李积山 103845103841667 沈雅威 103845213222017 叶燕如 103845213311008 金梦迪 66 70 50 64 110 100 88 103 326 325
103845104751648 陈思羽 103845104751649 段伟祥 103845104231650 高妍 103845100571651 韩昱 103845105041652 何婷婷 103845104031653 黄明荣 103845103841654 李晓旭 103845103841655 刘昉 103845103841656 马瑞洁
10 103845103841657 唐未 11 103845103841658 吴淑妃 12 103845103841659 夏光远 13 103845104231660 徐捷凯 14 103845213702136 邓雅程 15 103845213541322 吴昊昊 16 103845211204049 陈炎超 17 103845211203040 刘倡 18 103845213302026 陈明明 19 103845213221010 杨梓 20 103845213541325 黄诗翔 21 103845213707032 李堂成 22 103845211207020 吴鹏程 23 103845214420138 赵宇 68 63 60 59 55 69 65 68 64 61 62 74 68 67 57 58 55 60 52 53 127 120 105 126 110 113 118 91 377 368 351 335 331 326 326 325 324 322
中国海洋大学研究生录取情况综合
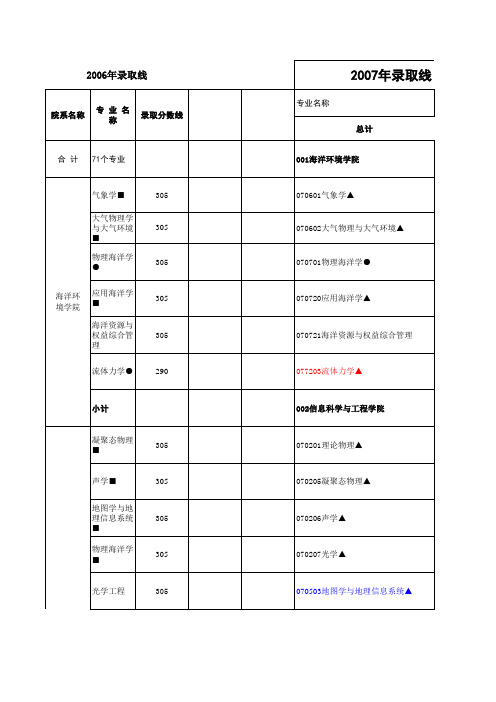
305
305 305 305 305 312(7) 305 290 300 290 290 306(16) 314(24) 290
070207 光学 地图学与地理 070503 信息系统 海洋信息探测 070722 与处理 080300 光学工程 通信与信息系 081001 统 信号与信息处 081002 理 计算机系统结 081201 构 计算机软件与 081202 理论 计算机应用技 081203 术 摄影测量与遥 081602 感 003化学化工学院 化学化工学院 070301 无机化学
305 170
020209数量经济学 010外国语学院 010外国语学院 050201英语语言文学 050203法语语言文学 050205日语语言文学 050210亚非语言文学 050211外国语言学及应用语言学 011文学院 011文学院 050103汉语言文字学 050105中国古代文学 050106中国现当代文学 060103历史地理学 012法学院 012法学院 030103宪法学与行政法学 030107经济法学 030108环境与资源保护法学 030109国际法学(含国际公法等) 030201政治学理论 030206国际政治 030301社会学 120401行政管理 013 数学系 070101基础数学▲ 070102计算数学▲ 070104应用数学▲ 070105运筹学与控制论▲ 014基础教学中心 014基础教学中心
环境地质方向(考试科目为工程地质 或水文地质)
321(+21) 363(+63) 300
环境规划与管 083020 理 011管理学院 011管理学院 管理科学与工 120100 程 120201 会计学 120202 企业管理 120203 旅游管理 技术经济及管 120204 理
海洋研究所2021年秋季入学博士报名通过资格审核考生名单
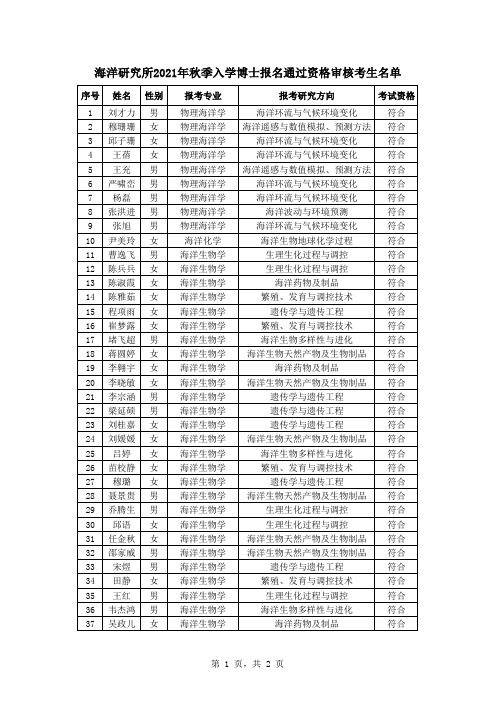
海洋地球物理与构造地质 海洋生态安全 海洋生态安全 海洋生态安全 生物资源生态学 养殖生态学 生物资源生态学 海洋生态安全 海洋生态安全 生物资源生态学 海洋生产力 生物资源生态学 养殖生态学
海洋金属腐蚀与防护 海洋生物腐蚀与污损 海洋金属腐蚀与防护 海洋生物腐蚀与污损 海洋金属腐蚀与防护 海洋金属腐蚀与防护 海洋生物腐蚀与污损
海洋研究所2021年秋季入学博士报名通过资格审核考生名单
序号 姓名 性别
1 刘才力 男 2 穆珊珊 女 3 邱子珊 女 4 王蓓 女 5 王充 男 6 严啸峦 男 7 杨磊 男 8 张洪进 男 9 张旭 男 10 尹美玲 女 11 曹逸飞 男 12 陈兵兵 女 13 陈淑霞 女 14 陈雅茹 女 15 程项雨 女 16 崔梦露 女 17 堵飞超 男 18 蒋圆婷 女 19 李翱宇 女 20 李晓敏 女 21 李宗涵 男 22 梁延硕 男 23 刘桂嘉 女 24 刘媛媛 女 25 吕婷 女 26 苗校静 女 27 穆璐 女 28 聂景贵 男 29 乔腾生 男 30 邱语 女 31 任金秋 女 32 邵家威 男 33 宋煜 男 34 田静 女 35 王红 男 36 韦杰鸿 男 37 吴政儿 女
海洋环境化学 海洋环境化学 海洋环境化学 海洋环境化学 海洋环境化学 养殖遗传学与育种生物技术 养殖遗传学与育种生物技术 养殖遗传学与育种生物技术 水产养殖生物病害防治 养殖遗传学与育种生物技术
考试资格
符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合
符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合 符合
海洋学基础知识(大连海事大学海洋科学专业课海洋生物学)PPT课件

3、What is the factors effect salinity?
Water removed by Evaporation Freezing And added by precipitation (rain & snow) Melting (iceberg)
分泌物、 排泄物、 分解物、 代谢物、 陆源排海
2、Salinity(盐度)
• Salinity is defined as the total amount of salt dissolved in seawater.
• Salinity is usually expressed as the number of grams of salt left behind when 1,000 grams of seawater are evaporated.
海水结冰吗?
请你思考
➢ 海水含盐度很高,降低了海水的冰点。淡水结冰是在0度, 含10‰盐度的水冰点为0.5℃,而含35‰盐度的水冰点是-1.9℃。 地球上各大洋海水平均
盐度为34.48‰,因此, 海水的冰点在-1.9℃左右。 ➢ 海洋受洋流、波浪、风暴 和潮汐影响很大,这些因
素一方面加强了海水混合
•海洋上空的气温比陆地 上空的气温变化慢,海 水对大气温度起着调节 作用。
海水温度变化及其影响因素
热量的 收支
空间变化
水平 方向
垂直 方向
时间变化
季节 年际 变化 变化
对大气 温度的 调节作 用
收入:太 阳辐射
支出:海 水蒸发
由低纬 向高纬 递减; 同纬度 暖流附 近水温 高,寒 流附近 水温低
西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示

第53卷 第7期 2023年7月中国海洋大学学报P E R I O D I C A LO FO C E A N U N I V E R S I T YO FC H I N A53(7):094~103J u l y,2023西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示❋陈文文1,胡敏章2❋❋,张锦昌3,4,5❋❋,林婧雪4,6,张旭博3,4,5(1.中国海洋大学,山东青岛266100;2.中国地震局地震研究所,地震大地测量重点实验室,湖北武汉430071;3.南方海洋科学与工程广东省实验室(广州),广东广州511458;4.中国科学院南海海洋研究所,南海生态环境工程创新研究院,中国科学院边缘海与大洋地质重点实验室,广东广州511458;5.中国-巴基斯坦地球科学研究中心,中国科学院-巴基斯坦高等教育委员会,巴基斯坦伊斯兰堡45320;6.中国科学院大学,北京100049)摘 要: 本文利用卫星重力数据和海底地形数据对大塔穆火山开展详细的重力导纳分析㊂结果显示大塔穆火山的岩石圈有效弹性厚度是1~3k m ,指示火山形成于洋中脊之上,符合低重力异常和洋中脊三联点的构造背景㊂A i r y 均衡模型和岩石圈挠曲均衡模型推算的大塔穆火山的平均地壳厚度是11~17k m ,最厚处拥有一个约30k m 的地壳根,与实际地震观测结果基本一致㊂大塔穆火山的超厚洋壳不同于正常洋中脊,目前地幔柱与洋中脊相互作用是比较合理的成因模式㊂研究还发现大塔穆火山底下存在一个质量缺失的低密度区,这个区域可能是残留岩浆房造成的结果,与火山中心的地球化学特征㊁地震波速异常以及广泛的后期火山活动相吻合㊂另外,这个低密度区提供浮力支撑火山中心隆起,可能导致火山侧翼因差异性沉降而产生正断层㊂关键词: 西太平洋;洋底高原;海底火山;重力均衡;导纳分析;有效弹性厚度;质量载荷中图法分类号: P 738;P 67 文献标志码: A 文章编号: 1672-5174(2023)07-094-10D O I : 10.16441/j.c n k i .h d x b .20220219引用格式: 陈文文,胡敏章,张锦昌,等.西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示[J ].中国海洋大学学报(自然科学版),2023,53(7):94-103.C h e nW e n w e n ,H uM i n z h a n g ,Z h a n g J i n c h a n g ,e t a l .G r a v i t y a d m i t t a n c e a n a l y s i s a n d i t s i m pl i c a t i o n s f o r t h e f o r m a t i o n o f t h eT a m uM a s s i f i n t h ew e s t P a c i f i cO c e a n [J ].P e r i o d i c a l o fO c e a nU n i v e r s i t y of C h i n a ,2023,53(7):94-103. ❋ 基金项目:广东省自然科学基金杰出青年项目(2021B 1515020098);南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项项目(G M L 2019Z D 0205);国家自然科学基金重大项目(41890813);中国科学院项目(133244K Y S B 20180029,131551K Y S B 20200021,Y 4S L 021,Q Y Z D Y -S S W -D Q C 005,I S E E 2021P Y 03);国家重点研究发展计划项目(2017Y F C 1500403-03);广东省重点领域研究发展计划项目(2020B 1111520001);海南省自然科学基金项目(421Q N 381)资助S u p p o r t e db y t h eN a t u r a l S c i e n c e F o u n d a t i o n f o r D i s t i n g u i s h e dY o u n g S c h o l a r s o f G u a n g d o n g P r o v i n c e (2021B 1515020098);t h eK e y S p e c i a l P r o j e c t f o rI n t r o d u c e d T a l e n t sT e a m o fS o u t h e r n M a r i n eS c i e n c ea n d E n g i n e e r i n g G u a n g d o n g L a b o r a t o r y (G u a n g z h o u )(G M L 2019Z D 0205);t h eK e y P r o g r a mo fN a t i o n a lN a t u r a l S c i e n c eF o u n d a t i o no fC h i n a (41890813);t h eC h i n e s eA c a d e m y of S c i -e n c e s P r o je c t (133244K Y S B 20180029,131551K Y S B 20200021,Y 4S L 021,Q Y Z D Y -S S W -D Q C 005,I S E E 2021P Y 03);t h eN a t i o n a l K e y R e s e a r c h a n dD e v e l o p m e n t P r o g r a mof C h i n a (2017Y F C 1500403-03);t h eG u a ng d o n g P r o v i n c i a l R e s e a r ch a n dD e v e l o p m e n t P r o -g r a mi nK e y Ar e a s (2020B 1111520001);t h eH a i n a nP r o v i n c i a l N a t u r a l S c i e n c e F o u n d a t i o n o f C h i n a (421Q N 381)收稿日期:2022-04-13;修订日期:2022-05-13作者简介:陈文文(1981 ),女,博士㊂E -m a i l :w e n w e n c h e n @o u c .e d u .c n❋❋ 通讯作者:E -m a i l :h u m i n z h a n g @126.c o m ;j z h a n g@s c s i o .a c .c n 1 背景火山作用将地球内部物质输送到岩石圈,塑造了地表的构造形态㊂地球上的火山作用主要分布在扩张型板块边界(洋中脊)㊁汇聚型板块边界(岛弧)以及与热点有关的地区㊂强烈的热点作用喷出大量岩浆到地表,形成面积超过十万平方千米㊁地壳异常厚的火山区域,称为大火成岩省(L a r g e I gn e o u s P r o v i n c e )[1]㊂洋底高原(O c e a n i cP l a t e a u )是一种常见的海底地质构造单元,多为宽广的平顶海山,高几千米,面积可达数十到数百万平方千米,单个大火成岩省的体积达数百万立方千米,具有异常厚的地壳[1]㊂据统计,目前已知的洋底高原总面积约占全球海洋地壳(洋壳)面积的5.11%[2]㊂作为大陆溢流玄武岩在海洋中的对应物,洋底高原被认为是由短时间内大量的地幔岩浆喷发而成,在深海盆地中洋底高原是最显著的大火成岩省[3-4]㊂然而,不同于大陆溢流玄武岩,洋底高原隐藏于大洋深处,避免了风化侵蚀等对原始地质现象的改造和破坏;并且,其受陆源岩石的污染较少,更能代表地幔源区的岩性特质㊂因此,洋底高原是地球构造演Copyright ©博看网. All Rights Reserved.7期陈文文,等:西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示化历史进程中大规模岩浆活动的忠实记录者,对研究地幔动力学甚至地球演化历史具有重要意义㊂洋底高原的形成机制至今仍没有定论㊂过去几十年来,前人已经提出了几种洋底高原的成因假说,如地幔柱模型㊁板块模型㊁陨石撞击等,但没有一种假说可以完全解释所有的主要观测结果[5-12]㊂地幔柱模型(P l u m eM o d e l)提出大型海底火山是由于深部地幔热柱上升到岩石圈底部喷发岩浆而形成;而板块模型(P l a t eM o d e l)主张在地表板块的扩张边界处(即洋中脊)浅部地幔减压熔融喷发岩浆而促使大型海底火山形成㊂陨石碰撞成因由于缺乏证据而很少被学者接受㊂最新的全球三大洋底高原对比研究指出它们具有显著的地形隆起㊁异常厚的地壳㊁负地幔重力异常,以及形成于洋中脊之上或附近等共性特征,将地幔柱与洋中脊过程结合起来对已有成因模型进行修正,提出了地幔柱与洋中脊相互作用这个替代模型,是目前对洋底高原形成机制较为合理的解释[13-14];但其具体动力学模式还停留在定性描述和概念模型的层面上㊂沙茨基海隆(S h a t s k y R i s e)位于西太平洋海底,在日本以东约1600k m的公海区,是一座典型的海洋大火成岩省[3-4],是现存地球上第三大的洋底高原,仅次于太平洋的翁通爪哇高原(O n t o n g J a v aP l a t e a u)和印度洋的凯尔盖朗高原(K e r g u e l e nP l a t e a u)㊂沙茨基海隆是由晚侏罗-早白垩期间(约147~126M a)大规模火山活动所形成[15-16],高约4k m,面积约5ˑ105k m2,体积近7ˑ106k m3㊂沙茨基海隆主要由几座已停止活动的大型海底火山组成(见图1),其中最大的那座名叫大塔穆火山(T a m u M a s s i f),被誉为地球上最大的单体火山[15]㊂大塔穆火山是沙茨基海隆最早形成的部分,顶部较为平坦,水深约2k m,侧翼坡度缓和,面积约3ˑ105k m2㊂(500m等深线间距的背景水深数据来自于联合船测水深和卫星测高重力异常数据反演的海底地形模型[25]㊂大塔穆火山是文中提到的 地球上最大的单体火山 [15],黑色虚线方框指示本文的大塔穆火山研究区㊂红色实线表示磁异常条带和断裂带,展示了区内为洋中脊三联点的构造特征[19]㊂黑色实线表示跨越大塔穆火山的代表性人工地震测线[22,26]㊂红色圆圈表示大洋钻探取得火山基底岩石的钻孔位置㊂左上角的小图指示海隆在西太平洋的位置,位于日本㊁伊豆-小笠原-马里亚纳俯冲带的东边㊂B a t h y m e t r y d a t aw i t h500mc o n t o u r a r e f r o ms a t e l l i t e a l t i m e t r y[25].T a m uM a s s i f i s t h eE a r t h s l a r g e s t s i n g l e v o l c a n om e n t i o n e d i n t h e t e x t[15].B l a c k d a s h e d b o x h i g h l i g h t s t h e s t u d y a r e a o f T a m uM a s s i f.R e d l i n e s d e n o t em a g n e t i c a n o m a l y l i n e a t i o n s a n d f r a c t u r e z o n e s,i m p l y i n g a t r i p l e j u n c t i o n o fm i d-o c e a n r i d g e s[19].B l a c k l i n e r e p r e s e n t s s e i s m i c s u r v e y a c r o s sT a m uM a s s i f[22,26].R e d c i r c l e s s h o w t h eO D P a n d I O D P s i t e s t h a t c o r e i g n e o u s b a s e m e n t r o c k s.I n s e t i n t h e u p p e r l e f t s h o w s t h e l o c a t i o n o f S h a t s k y R i s e i n t h e w e s t P a c i f i c O c e a n,a n d t o t h e e a s t o f J a p a n a n d I B Ms u b d u c t i o n z o n e s.)图1沙茨基海隆的测深与构造图F i g.1S h a t s k y R i s e b a t h y m e t r y a n d t e c t o n i cm a p59Copyright©博看网. All Rights Reserved.中 国 海 洋 大 学 学 报2023年沙茨基海隆具有时空分布连续的火山结构㊁较薄的火山盖顶沉积㊁良好的地磁场分布以及成熟的洋壳等特征,同时具有洋中脊三联点(属于一个古太平洋三联点,由伊泽奈崎板块㊁法拉龙板块和太平洋板块交汇而成)的构造背景[9,10,13,15-24],是研究大型海底火山㊁洋底高原㊁海洋大火成岩省等形成机制的天然实验室㊂本文聚焦在沙茨基海隆内最大的海底火山-大塔穆火山,在卫星测高重力与海底地形观测数据基础上,采用重力导纳分析技术研究该火山的重力均衡特征,计算区内洋壳厚度㊁岩石圈有效弹性厚度,并揭示火山底部载荷,从而探讨其成因与动力学过程㊂通过这座誉为地球上最大火山的研究,有望加深对全球大型海底火山形成演化的基本规律的认识和理解㊂2 方法与数据2.1重力导纳分析方法根据A i r y 均衡模型和岩石圈挠曲均衡模型[27-28],本文将基于理论模型计算的重力导纳与基于实测海底地形与重力异常数据的观测重力导纳进行对比分析,以研究海底构造的均衡状况㊂参考文献[29],理论重力导纳计算公式为:Z (k )f l e x =2πG (ρc -ρw )e -k d (1-Φe (k )e -k t )㊂(1)式中:G 为万有引力常数;ρc ㊁ρw 分别为地壳和海水密度;d 为区域平均水深;t 为平均地壳厚度;k =2π/λ为波数,λ为地形波长,Φe (k )为岩石圈挠曲响应函数:Φe (k )=D k 4(ρm -ρc )g +1éëêêùûúú-1㊂(2)式中:ρm 为地幔密度;D =E T 3e /[12(1-υ2)]为岩石圈抗挠刚度,E 为杨氏模量,T e 为岩石圈有效弹性厚度,υ为泊松比㊂观测重力导纳的计算公式为:Z (k )=<ΔG (k )H *(k )><H (k )H *(k )>㊂(3)式中:ΔG (k )为重力异常的傅里叶变换;H (k )为地形的傅里叶变换;符号 * 表示复共轭;符号 <> 表示在频率域内求周期平均㊂重力导纳分析过程中,主要涉及岩石圈有效弹性厚度T e ,平均水深d ,地壳密度三个参数的调整㊂胡敏章等[30]研究指出,在频率域内波长小于50k m 的情况下,由于海底地形被岩石圈强度所支撑,不产生均衡补偿,因此短波段(<50k m )理论重力导纳不随着岩石圈有效弹性厚度(T e )的变化而变化,但依然受地壳密度和平均水深参数影响,可在20~50k m 波段中通过重力导纳分析来先行测试模型最佳地壳密度和水深参数㊂在比较大塔穆火山理论重力导纳与观测重力导纳的过程中,测试了不同的地壳密度(2500~3000k g/m 3)和水深(平均模型深度ʃ500m )㊂通过不断调整地壳密度㊁水深参数,计算理论重力导纳,并将其与观测重力导纳进行比较,当两者之差的均方根最小时,获得相关参数的最佳估值㊂从图2看出,当理论重力导纳与观测重力导纳差值均方根最小的时候(1.475m G a l /k m ),最佳地壳密度和水深的数值分别是2500k g /m 3和4600m ㊂同样,地壳密度参数和水深参数确定后,通过岩石圈取不同值时的理论重力导纳与观测重力导纳进行对比,当理论重力导纳与观测重力导纳之差的均方根达到最小时,可以确定最佳的岩石圈有效弹性厚度[29]㊂从图3看出,当理论重力导纳与观测重力导纳差值均方根最小的时候(3.318m G a l /k m ),大塔穆火山的最佳岩石圈有效弹性厚度是3k m㊂(在海底地形20~50k m 波段中,通过理论重力导纳与观测重力导纳的比较,确定模型最佳的地壳密度和水深㊂D e t e r m i n a t i o n o f t h em o d e l o p-t i m u mc r u s t a l d e n s i t y a n dw a t e r d e p t h i s b y f i t t i n g t h e o b s e r v e d a n d t h e -o r e t i c a l a d m i t t a n c e s a t 20~50k m w a v e b a n d s i n s e a f l o o r t o p o g r a p h y.)图2 模型最佳地壳密度和水深示意图F i g .2 M o d e l o p t i m a l v a l u e s o f c r u s t d e n s i t y a n dw a t e r d e pth 图3 大塔穆火山岩石圈有效弹性厚度误差分析图F i g .3 E r r o r a n a l y s i s o f l i t h o s ph e r i c e f f e c t i v e e l a s t i c t h i c k n e s s o f T a m uM a s s i f69Copyright ©博看网. All Rights Reserved.7期陈文文,等:西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示2.2实验使用数据与参数本次研究使用到卫星测高重力异常数据和海底地形数据,数据分析范围见图1中黑色虚线方框,包含整个大塔穆火山的主体部分㊂卫星测高重力异常数据来自斯克利普斯海洋研究所(S c r i p p s I n s t i t u t i o n o f O c e a-n o g r a p h y),版本V23.1㊂海底地形数据是通过重力垂直梯度异常和船测水深联合求得[31-32],数据空间分辨率为1'ˑ1'㊂在计算岩石圈有效弹性厚度过程中,胡敏章等[30]通过测试不同的海底地形模型,发现联合重力垂直梯度异常和船测水深数据构建的海底地形模型最适用于洋壳重力均衡分析㊂表1大塔穆火山重力均衡分析地壳模型参数T a b l e1C r u s t a l P a r a m e t e r s f o r I s o s t a s y a n a l y s i s o f T a m um a s s i f参数P a r a m e t e r符号N o t a t i o n数值V a l u e海水密度①ρw1030k g/m3地壳密度②ρc2500k g/m3地幔密度③ρm3350k g/m3平均水深④d4600m平均地壳厚度⑤t14k m杨氏模量⑥E1011N/m2泊松比⑦υ0.25底部载荷深度⑧d L14k m 注:平均地壳厚度来自于A i r y均衡模型和地震观测的联合约束[26]㊂T h em e a n c r u s t a l t h i c k n e s s(t)i s e s t i m a t e d b a s e d o nA i r y i s o s t a t i cm o d-e l a n ds e i s m i c p r of i l e s[26].①D e n s i t y o f s e a w a t e r;②D e n s i t y o f c r u s t;③D e n s i t y o fm a n t l e;④M e a nw a t e r d e p t h;⑤M e a nc r u s t a l t h i c k n e s s;⑥Y o u n g sm o d u l u s;⑦P o i s s o n s r a t i o;⑧I n n e r l o a d i n g d e p t h.3结果3.1大塔穆火山A i r y均衡模型理论重力导纳与观测重力导纳对比大塔穆火山(长宽约650k m乘以450k m)是沙茨基海隆中最大㊁最老的海底火山[10,15,17,18,22,26]㊂尽管大塔穆火山巨大,但是它的重力异常值很低(小于30 m G a l)[14],这意味着火山大体达到了重力均衡状态[33]㊂基于A i r y均衡模型(公式(1)中T e=0的情况),计算得到的理论重力导纳与观测重力导纳的对比分析见图4㊂导纳结果显示大塔穆火山的平均地壳厚度是11~ 17k m,这与实际地震观测结果相符合[26]㊂火山海底地形波长(λ)大于160k m或者小于60k m(即波数1/λ,小于0.00625或者大于0.01667)的时候,处于重力均衡状态㊂当火山海底地形波长在60~160k m(波数在0.00625~0.01667)之间时,结果显示观测重力导纳值明显低于理论重力导纳值,这意味着相对于A i r y 均衡模型该波段存在质量缺失或者低密度区;同时,也暗示着该地区的海底地形隆起可由低密度重力异常产生的浮力所支撑㊂(黑色圆点表示观测重力导纳数据(带误差)㊂红色虚线㊁点线㊁点虚线表示基于不同平均地壳厚度(t)的理论重力导纳曲线㊂F i l l e db l a c k c i r c l e s w i t h e r r o r b a r s a r e o b s e r v e d a d m i t t a n c e s.R e d d a s h e d,d o t t e d,a n d d a s h-d o t t e d l i n e s a r e t h e o r e t i c a l a d m i t t a n c e s f o r d i f f e r e n tm e a n c r u s t a l t h i c k-n e s s(t).)图4大塔穆火山A i r y均衡模型理论重力导纳与观测重力导纳对比图F i g.4C o m p a r i s o n o f g r a v i t y a d m i t t a n c e s f r o mA i r yi s o s t a t i cm o d e l,a n d a l t i m e t r y a n d b a t h y m e t r yo b s e r v a t i o n s o v e r T a m u M a s s i f3.2大塔穆火山挠曲均衡模型理论重力导纳与观测重力导纳对比图5结果显示大塔穆火山的岩石圈有效弹性厚度是1~3k m㊂这个低数值的岩石圈有效弹性厚度,意味着大塔穆火山形成于强度很弱的岩石圈上;考虑到其磁异常数据显示的洋中脊三联点构造特征[18-19,24],进一步指示了大塔穆火山形成于洋中脊之上㊂(黑色圆点表示观测重力导纳数据(带误差)㊂红色虚线㊁点线㊁点虚线表示基于不同岩石圈有效弹性厚度(T e)的理论重力导纳曲线㊂F i l l e d b l a c k c i r c l e sw i t h e r r o r b a r s a r e o b s e r v e d a d m i t t a n c e s.R e d d a s h e d,d o t-t e d,a n d d a s h-d o t t e d l i n e s a r e t h e o r e t i c a l a d m i t t a n c e s f o r d i f f e r e n t l i t h o s-p h e r i c e f f e c t i v e e l a s t i c t h i c k n e s s(T e).)图5大塔穆火山挠曲均衡模型理论重力导纳与观测重力导纳对比图F i g.5C o m p a r i s o n o f g r a v i t y a d m i t t a n c e s f r o mf l e x u r a li s o s t a t i cm o d e l,a n d a l t i m e t r y a n d b a t h y m e t r yo b s e r v a t i o n s o v e rT a m uM a s s i f79Copyright©博看网. All Rights Reserved.中 国 海 洋 大 学 学 报2023年3.3大塔穆火山考虑底部载荷时的理论重力导纳与观测重力导纳对比在上述均衡模型中考虑到底部载荷的影响,进一步分析大塔穆火山中质量缺失的问题㊂热地幔物质上升至地壳底部的残留,或者热异常等区域,形成局部的相对低密度区域,即为底部载荷,其浮力对海山有一定的支撑作用㊂以参数f 定义底部载荷与海底地形载荷之比,根据胡敏章等[29]的公式(6),计算f 取不同值时的理论重力导纳,将其与观测重力导纳相比较,可获得沙茨基海隆底部载荷规模的估值㊂根据3.1节的分析,将底部载荷深度设置在14k m ,也就是大塔穆火山平均地壳厚度11~17k m 的中间值,指示着火山底部地壳㊂图6显示大塔穆火山的底部载荷(f )在0.1~0.6之间㊂I t o 等[34]和S h i m i z u 等[35]的研究指出大塔穆火山的沉降要小于地幔柱模型的预测,暗示着相对于周边洋壳的均衡,火山地壳中存在质量缺失,火山中的相对低密度区产生浮力,从而支撑或者部分支撑了火山地形的隆起㊂另外,根据氧化镁成分的分析,H u s e n 等推测在大塔穆火山下可能存在一个浅部(小于6k m )和一个深部(18~24k m )连接起来的岩浆房系统㊂导纳分析结果显示的大塔穆火山底下这个质量缺失或者低密度区可能是由于这个岩浆房系统残留在地壳中所引起的[36]㊂(黑色圆点表示观测重力导纳数据(带误差)㊂红色虚线㊁点线㊁点虚线表示基于不同底部载荷(f )的理论重力导纳曲线㊂F i l l e db l a c k c i r c l e sw i t h e r r o r b a r s a r e o b s e r v e d a d m i t t a n c e s .R e dd a s h e d ,d o t t e d ,a n dd a s h -d o t -t e dl i n e sa r et h e o r e t i c a la d m i t t a n c e sf o rd i f f e r e n t i n n e r m a s sl o a d i n g (f ).)图6 大塔穆火山考虑底部载荷时的理论重力导纳与观测重力导纳对比图F i g .6 C o m p a r i s o n o f t h e o r e t i c a l a n d o b s e r v e d g r a v i t ya d m i t t a n c e s r e g a r d i n g i n n e rm a s s l o a d i n g ov e rT a m uM a s s i f 4 讨论4.1大塔穆火山地形与地壳厚度重力均衡导纳分析指出大塔穆火山的岩石圈有效弹性厚度小于3k m ,揭示了大塔穆火山形成于一个很弱的岩石圈之上,或者说形成于洋中脊之上,属于 o nr i d ge 海底构造[37-38],这点符合沙茨基海隆形成于洋中脊三联点的构造背景[18,19,24,39]㊂选取一条跨越大塔穆火山的代表性测线(A -B 测线,位置见图1),基于岩石圈挠曲均衡模型,利用第3节获得的岩石圈有效弹性厚度等参数和海底地形数据,可根据公式(2)计算莫霍面挠曲,进而推断莫霍面深度,推算结果见图7㊂图7同时给出了基于A i r y 均衡模型给出的莫霍面深度㊂总体来说,两个理论模型的莫霍面推算结果大致上与地震观测得到的莫霍面结果相吻合[26];揭示了大塔穆火山底下埋藏着一个深达约30k m 的地壳根㊂这个巨厚的洋壳意味着大量的岩浆通过洋中脊的海底扩张输出到地表㊂(地震莫霍面数据来自于张锦昌等[26]㊂红色圆圈表示大洋钻探钻孔U 1347取得火山基底岩石的位置㊂A -B 剖面位置见图1㊂S e i s m i cM o h od a t a a ref r o mZ h a ng e t a l .[26].R e d c i r c l e r e pr e s e n t s t h e I O D PU 1347s i t e t h a t c o r e s i g n e o u s b a s e m e n t r o c k .L o c a t i o n o f L i n eA -B i s s h o w n i nF i g .1.)图7 跨越大塔穆火山A -B 剖面的海底地形与莫霍面形态F i g.7 S e a f l o o r a n dM o h o u n d u l a t i o n s f r o mL i n e A -B a c r o s sT a m uM a s s i f4.2大塔穆火山与夏威夷-帝王海山链以及周边侏罗纪、白垩纪洋壳为了和大塔穆火山周边构造对比,采用移动窗口导纳分析技术[40]获取沙茨基海隆以及周边地区的岩石圈有效弹性厚度分布图(见图8)㊂结果显示沙茨基海隆与赫斯海隆都具有较低的岩石圈有效弹性厚度,符合两个海隆类似构造成因的认识[41]㊂沙茨基海隆的岩石圈有效弹性厚度要明显低于夏威夷-帝王海山链,这意味着洋底高原的形成与海山的形成具有不同的机理㊂夏威夷-帝王海山链是板内热点构造[42-43],是地幔热点刺穿先存岩石圈并加载在先存岩石圈上,所以岩石圈有效弹性厚度较大㊁岩石圈强度较大;而沙茨基海隆形成于洋中脊,没有先存的岩石圈,是新生的洋壳,所以岩石圈有效弹性厚度接近为零㊁岩石圈强度很弱㊂根据大洋岩石圈的冷却模型,大洋岩石圈随着海89Copyright ©博看网. All Rights Reserved.7期陈文文,等:西太平洋大塔穆火山的重力导纳分析以及对其形成机制的启示底扩张不断远离洋中脊会变老㊁变厚和变冷,导致岩石圈强度增大[44-45]㊂岩石圈有效弹性厚度记录了这一演变过程㊂图9整理了大塔穆火山周边的侏罗纪㊁白垩纪洋壳的岩石圈有效弹性厚度代表值,数据来自胡敏章等[30]㊂总体上,白垩纪洋壳的岩石圈有效弹性厚度比更老的侏罗纪洋壳要小,这符合上述冷却模型的推测㊂沙茨基海隆形成于晚侏罗-早白垩,跨越了侏罗纪到白垩纪两个地质时期,海隆的岩石圈有效弹性厚度比周边侏罗纪洋壳或者白垩纪洋壳都要低,这意味着沙茨基海隆虽然形成于洋中脊之上,但是其动力学机制区别于正常的洋中脊海底扩张模式㊂(黑色等深线勾勒出沙茨基海隆㊁赫斯海隆以及夏威夷-帝王火山链㊂红色星号代表大塔穆火山㊂B l a c k l i n e s a r e b a t h y m e t r y c o n t o u r s d e p i c t i n g S h a t s k yR i s e ,H e s sR i s e ,a n dH a w a i i a n -E m pe r o r S e a m o u n t C h a i n .R e d s t a r d e n o t e s t h e l o c a t i o n of T a m uM a s s i f .)图8 沙茨基海隆及周边地区的岩石圈有效弹性厚度F i g .8 L i t h o s p h e r i c e f f e c t i v e e l a s t i c t h i c k n e s s a r o u n d S h a t s k y Ri s e 4.3大塔穆火山形成机制的启示相对于周边洋盆,大塔穆火山具有超薄(1~3k m )的岩石圈有效弹性厚度;具有超厚(11~17k m )的平均地壳厚度;以及火山底下存在低密度区㊂超薄的岩石圈有效弹性厚度指示了大塔穆火山形成于洋中脊之上,这点符合磁异常数据揭示的洋中脊三联点构造背景[18-19,24,39],也符合火山形成于板块扩张边界,由于没有先存岩石圈(或强度极弱的岩石圈),岩浆喷出过程基本处于动力学均衡状态,展示出较低的重力异常值[14,33]㊂大塔穆火山展示出比周边侏罗纪或者白垩纪正常洋壳更低的岩石圈有效弹性厚度,加上它拥有比正常洋壳(6~7k m 厚度)厚一倍以上的地壳,都意味着大塔穆火山的形成机制是有区别于正常洋中脊的㊂无论A i r y 均衡和岩石圈挠曲均衡模型推算,还是人工地震实际观测结果[22,26],都揭示了大塔穆火山底下存在一个厚达约30k m 的地壳根,这样超厚的洋壳需要大量的岩浆从地幔输出到地表,正常洋中脊是没法满足的㊂尽管大塔穆火山形成于洋中脊之上,但是还需要额外的岩浆供给才能构建体量如此厚硕的洋壳㊂大塔穆火山的形成,在洋中脊过程基础上,还需要一个额外的机制㊂第一种可能就是地幔柱(M a n t l epl u m e )[1,3-4],地幔柱可以从地幔深部带来足够多的物质输送到洋中脊下面,产生地幔柱与洋中脊的相互作用,两者加成从而喷出大规模的岩浆㊂地幔柱模式在大塔穆火山上找到不少观测证据,包括大洋钻探航次在大塔穆火山上钻取到巨厚块状熔岩(厚达23m ),揭示了大规模熔岩流㊁快速喷发并扩散的特征[9,15];多道地震反射剖面显示,大塔穆火山是地球上最大的单体99Copyright ©博看网. All Rights Reserved.中国海洋大学学报2023年(红色星号代表大塔穆火山㊂黑色实线P&S是大洋岩石圈冷却模型数据来自于P a r s o n s a n d S c l a t e r[44]㊂绿色虚线G D H1是板块冷却模型数据来自于S t e i n a n d S t e i n[45]㊂蓝色三角(夏威夷-帝王海山链)㊁红色圆圈(侏罗纪洋壳)㊁紫色方形(白垩纪洋壳)数据来自于胡敏章等[30]㊂R e d s t a r d e n o t e s t h e l o c a t i o n o f T a m uM a s s i f.B l a c k l i n e l a b e l l e d P&S i s f r o m p l a t e c o o l i n g m o d e l i n t r o d u c e db y P a r s o n s a n dS c l a t e r[44].G r e e nd a s h e d l i n e l a b e l l e dG D H1i s f r o m p l a t e c o o l i n g m o d e l s u g g e s t e db y S t e i na n d S t e i n[45].D a t a o fH a w a i i a n-E m p e r o rS e a m o u n tC h a i n(b l u e t r i a n g l e s), J u r a s s i c(r e d c i r c l e s)a n dC r e t a c e o u s(p u r p l e s q u a r e s)o c e a n i c c r u s t s a r e f r o m H u e t a l.[30].)图9大塔穆火山与夏威夷-帝王海山链以及周边侏罗纪㊁白垩纪洋壳的对比图F i g.9C o m p a r i s o n o f T a m uM a s s i f,H a w a i i a n-E m p e r o rS e a m o u n t C h a i n a n d J u r a s s i c a n dC r e t a c e o u so c e a n i c c r u s t s n e a r b y火山,具有围绕单一火山中心呈现盾状㊁两翼坡度低的结构[15,22,26]㊂这些观测反应了熔岩流喷发时粘度低㊁喷出速率高且流动距离长,大量岩浆很快地从火山中心流出并扩散至周边海盆,与地幔柱头大规模㊁快速的岩浆喷发预测的特征一致㊂沙茨基海隆北部的两座火山(奥里火山和希尔绍夫火山,见图1)和帕帕宁海岭年龄逐渐年轻㊁规模逐步变小并呈现链状排列,反映了从地幔柱头过渡到地幔柱尾的过程㊂因此,地幔柱与洋中脊相互作用机制是能够较好地解释大塔穆火山上这些重要观测和模拟结果的,但其具体的动力学模式目前仍停留在定性认识层面上,还没有开展足够的定量研究[9-11,13-14]㊂另一种可能的额外机制是大面积㊁不均一的富集地幔减压熔融(M a s s i v eh e t e r o g e n e o u sf e r t i l e m a n-t l e)[46-48]㊂与地幔柱作用相比,由于不需要深部地幔物质参与,岩浆来源较浅,这种机制在时间上可以更长㊁在空间上可以更广㊂支持这种机制的观测证据有:地球化学与同位素分析表明,M O R B(洋中脊玄武岩)是大塔穆火山熔岩的主要岩石类型,这种岩浆与太平洋M O R B相似,但与热点相关的O I B(洋岛玄武岩)不同[49-50]㊂从地震折射数据计算得出的速度模型表明,地壳厚度与地震波速呈现负相关关系,即较厚地壳的火山中心呈现较低的地震波速,意味着大塔穆火山具有化学异常特征的岩浆来源,指示具有较冷的浅部地幔物质被动上涌的特征[51]㊂这种机制目前研究较少,很多学者认为难以解释大塔穆火山这样超级规模岩浆量的产生㊂导纳分析结果显示大塔穆火山底下14k m处存在一个低密度区,这个质量缺失的区域可能由于残留岩浆房造成㊂地球化学分析指出在大塔穆火山地壳浅部和深部均存在岩浆房,构建起从下而上的岩浆输送系统[36,52]㊂这个岩浆房系统残留在大塔穆火山下面,就会产生一个相对于周围的低密度区,而这个低密度区对应了火山中心部位较两翼更低的地震波速(假设地震波速与地壳密度是正相关关系)[51]㊂残留的岩浆房系统在大塔穆火山主体形成后,导致了广泛的后期火山活动,例如大塔穆火山顶峰的多伦多海岭(约15M a 晚于大塔穆火山,来自于拖网岩石定年数据)[41,53-55],以及成千上万㊁星罗密布在大塔穆火山表面上的次生海山椎[23,56]㊂另外,大塔穆火山底下这个低密度区的存在会产生浮力来支撑火山中心地形的隆起,导致整个火山出现差异性沉降[34-35],也就是火山中心比侧翼沉降得要少,这样会在火山翼部产生诸多正断层,这个推测也得到了多道地震剖面观测的证实[22-23]㊂5结论(1)大塔穆火山的岩石圈有效弹性厚度是1~ 3k m,指示火山形成于很弱的先存岩石圈之上,即形成于洋中脊之上㊂这个结果符合大塔穆火山处于低重力异常的均衡状态,以及处于洋中脊三联点的构造背景㊂(2)大塔穆火山平均地壳厚度是11~17k m,并拥有一个厚达约30k m的地壳根㊂A i r y均衡模型和岩石圈挠曲模型推算的结果与实际地震观测结果大体一致㊂超厚的洋壳需要大量岩浆喷发产生,指示大塔穆火山形成机制区别于正常洋中脊,需要额外的动力学过程参与㊂地幔柱与洋中脊相互作用是目前较好的成因解释模式,但需开展定量研究进一步证实㊂(3)大塔穆火山底下存在一个质量缺失的低密度区,这个区域可能是残留岩浆房造成的结果,对应了火山中心氧化镁的地球化学特征和较低地震波速的实际观测㊁以及火山表面诸多的次生海岭和海山椎的发现㊂这个低密度区还会产生浮力支撑火山中心隆起,导致火山侧翼沉降更多,从而在火山翼部产生一系列正断层㊂001Copyright©博看网. All Rights Reserved.。
扇贝养殖季烟台近海CH4_和DMS的时空分布及影响因素

第54卷 第1期 2024年1月中国海洋大学学报P E R I O D I C A L O F O C E A N U N I V E R S I T Y O F C H I N A54(1):086~094J a n .,2024扇贝养殖季烟台近海C H 4和D M S 的时空分布及影响因素❋张升辉1,孙 婧2,赵 淼3,4,胡晓珂1❋❋(1.中国科学院海岸带研究所,海岸带生物学与生物资源利用重点实验室,山东烟台264006;2.山东省海洋资源与环境研究院,山东省海洋生态修复重点实验室,山东烟台264006;3.江苏省海洋生物资源与环境重点实验室,江苏海洋大学,江苏连云港222005;4.江苏省海洋资源开发研究院,江苏连云港222005)摘 要: 甲烷(C H 4)和二甲基硫(D i m e t h y l s u l f i d e ,D M S )是重要的挥发性生源活性气体,在调节气候变化方面具有重要意义㊂本文为探究筏式扇贝养殖对C H 4和D M S 分布与释放通量的影响,分别于2019年5㊁6㊁7㊁8㊁9和11月对烟台近海进行现场调查㊂结果表明,贝类滤食可以间接将水中的有机物质输入到沉积物中,导致扇贝养殖区底层C H 4平均浓度比非养殖区高25.64%㊂贝类滤食也可以将浮游植物体内的二甲基巯基丙磷(D i m e t h y l s u l f o n i o p r o pi o n a t e ,D M S P )转化为海水中的颗粒态D M S P ,增加D M S P 向D M S 的转化,与非养殖区相比,养殖区表层D M S 浓度增加了24.13%㊂另外,表层海水C H 4浓度处于过饱和状态,而D M S P 可能是表层海水C H 4重要的潜在来源㊂整个扇贝养殖周期内,调查海域C H 4海-气通量约为9.69ˑ106g ,D M S 海-气通量约为3.33ˑ107g ,是大气C H 4和DM S 重要的源㊂关键词: C H 4;D M S ;D M S P ;扇贝养殖中图法分类号: P 734 文献标志码: A 文章编号: 1672-5174(2024)01-086-09D O I : 10.16441/j.c n k i .h d x b .20220395引用格式: 张升辉,孙婧,赵淼,等.扇贝养殖季烟台近海C H 4和DM S 的时空分布及影响因素[J ].中国海洋大学学报(自然科学版),2024,54(1):86-94.Z h a n g S h e n g h u i ,S u n J i n g ,Z h a o M i a o ,e t a l .T e m p o -s p a t i a l d i s t r i b u t i o n s a n d i n f l u e n c i n g f a c t o r s o f C H 4an d D M S d u r i n g s c a l l o p f a r m i n g i n Y a n t a i o f f s h o r e [J ].P e r i o d i c a l o f O c e a n U n i v e r s i t y of C h i n a ,2024,54(1):86-94. ❋ 基金项目:国家自然科学基金项目(42006047);国家自然基金委重大研究计划项目(92051119);泰山学者攀登计划项目(t s p d 20210317)资助S u p p o r t e d b y t h e N a t i o n a l N a t u r a l S c i e n c e F o u n d a t i o n o f C h i n a (42006047);t h e M a j o r R e s e a r c h P r o gr a m o f N a t i o n a l N a t u r a l S c i e n c e F o u n d a t i o n o f C h i n a (92051119);t h e T a i s h a n S c h o l a r s C l i m b i n g P r o g r a m (t s pd 20210317)收稿日期:2022-09-15;修订日期:2022-11-22作者简介:张升辉(1987 ),男,助理研究员㊂E -m a i l :s h z h a n g @yi c .a c .c n ❋❋ 通信作者:E -m a i l :x k h u @yi c .a c .c n 甲烷(C H 4)和二甲基硫(D M S )是具有重要环境意义的挥发性生源活性气体,在全球碳和硫循环过程中扮演着重要角色㊂甲烷可以通过吸收地球长波辐射增加地球表明温度,是二氧化碳以外最重要的温室气体,而D M S 扩散进入大气以后可以被氧化形成硫酸盐气溶胶颗粒,增加对太阳光的反射,降低地球表面温度,是重要的反温室气体㊂海洋是全球大气C H 4和D M S 的重要自然源,每年通过海-气扩散进入大气的C H 4约占全球大气C H 4来源的5%,而D M S 是海水中最重要,含量最丰富的挥发性生源硫化物,其海-气通量为15~33T g㊃a -1,占海洋硫通量的90%以上[1]㊂鉴于C H 4和D M S 在全球碳循环㊁硫循环及调控全球气候方面的重要作用,国际大型研究计划上层海洋-低层大气研究(S O L A S )将C H 4和DM S 列为重要研究内容㊂目前我国学者对于溶存C H 4㊁D M S 及其前体物质二甲基巯基丙酸(D M S P )的研究区域已经涵盖了东海[2-3]㊁黄海[4-5]㊁南海[6-7]及长江[8-9],黄河[10]等主要河口陆架区,但是对于近岸养殖海域溶存C H 4㊁D M S 和D M S P 的研究报道并不多见㊂虽然近岸海区只占全球海洋面积的一小部分,但是甲烷释放量却占到了全球海洋释放总量的68%[11]㊂近几十年来,由于海洋渔业资源的锐减使得近海养殖业得到迅猛发展,全球的海水养殖每年以10%的速度增加[12]㊂中国的海水养殖业在养殖面积,放养种类以及产量等方面也得到了很大的发展㊂同时,大规模发展海水养殖也导致近岸水体各种理化因子发生改变和底泥环境的恶化㊂研究表明,海水养殖区的悬浮物㊁总氮㊁总磷㊁生化需氧量(B O D )㊁化学需氧量(C O D )一般均高于对照区,并且海水养殖区的沉降量比非养殖区大得多,导致水产养殖区的底泥中,碳㊁氮㊁磷的含量比周围水体沉积物中要1期张升辉,等:扇贝养殖季烟台近海C H 4和DM S 的时空分布及影响因素高[13]㊂水产养殖引起的近岸水体和沉积物环境的改变必然会对C H 4和DM S 在海水中的迁移转化及排放通量产生影响㊂烟台近岸海域位于山东半岛东北部,毗邻黄海,是一个半封闭海域,其独特的地理环境使其成为中国北方重要的扇贝养殖基地,养殖面积达150k m2,年产量约为180万t,包括人工鱼礁㊁底播养殖和筏式养殖,其中筏式养殖面积占总养殖面积的2/3以上[12]㊂目前,烟台近海的筏式扇贝养殖品种主要为海湾扇贝,大都集中在养马岛以东海域,养殖周期为每年的4 11月,而养马岛以西海域由于政府规划已经禁止进行渔业养殖活动(见图1),因此烟台近海是研究扇贝养殖对C H 4和D M S 释放的理想场所,并于2019年5㊁6㊁7㊁8㊁9和11月分别对烟台近海进行现场调查(调查站位如图1所示),探究筏式扇贝养殖对烟台近海C H 4和D M S 分布与释放通量的影响㊂(蓝色为非养殖站位,红色为养殖站位㊂B l u e d o t s r e p r e s e n t n o n -f a r m i n gs t a t i o n s ,a n d r e d d o t s r e p r e s e n t f a r m i n g st a t i o n s .)图1 调查站位图F i g .1 L o c a t i o n o f t h e s u r v e y st a t i o n 1 样品采集㊁保存与测定利用5L N i s k i n R o s e t t e 采水器采集烟台近海表层与底层海水,采样站位的海水T ㊁S 等水文参数由C TD 设备在采集海水样品时同步测定,D O 采用碘量法测定,C h l a ㊁C H 4㊁D M S 和D M S P 的保存与测定方法如下:C h l a 的测定:取800m L 海水样品通过0.70μm玻璃纤维滤膜(W h a t m a n)过滤,用锡纸将滤膜包好,在-20ħ下避光冷冻保存㊂测定时,用10m L 90%的丙酮溶液在4ħ条件下避光萃取滤膜24h,之后离心10m i n ,取上清液使用F -4500荧光仪(日本日立)测定其荧光信号,方法检测限为0.01μg㊃L -1㊂ C H 4的测定:采用顶空平衡-气相色谱法,水样预处理时,通过气密性进样针定量注入30m L 高纯N 2(纯度优于99.999%),使同体积的水样从样品瓶中排出,顶空样品瓶内形成气液两相共存的密闭体系.再将样品瓶固定于振荡器上剧烈振荡5m i n,避光环境下(25ħ)静置2h ,使C H 4在气液两相达到分配平衡㊂样品测定时,首先用针筒将预处理时排出的水样匀速注回样品瓶,迫使顶空气沿进气管路进入串联的定量环(1m L ),然后通过色谱柱(6i n c h ˑ3m m ,P a r o pa k -Q 填充柱)进入G C -F I D (2010P l u s ,S h i m a d z u,日本),测定顶空气中C H 4的浓度㊂柱箱恒温60ħ,进样口和F I D 的工作温度分别为150和300ħ,该方法检测C H 4的精密度和准确度均优于2%㊂D M S 的测定:采用吹扫捕集-气相色谱法[3],用玻璃注射器取2m L 海水样品注入到样品瓶中,通入40m L ㊃m i n -1的高纯N 2进行吹扫,吹扫出的D M S 通过N a f i o n 渗透管(P e r m a P u r e,美国)除去水分,并通过六通阀(V a l c o ,美国)富集于浸在液氮中的1/8T e f l o n 捕集管中;吹扫3m i n 后将捕集管放入热水(>90ħ)中进行加热解析,在载气(高纯氮气)携带下通过色谱柱(D B -S u l f u r S C D ,60mˑ0.32m m )进入到G C -F P D 气相色谱(2010P l u s ,S h i m a d z u,日本)中进行测定㊂柱箱恒温150ħ,进样口和F P D 的工作温度分别为240和250ħ,该方法D M S 的检出限约为0.2n m o l /L D M S,精密度优于5%㊂D M S P 的保存和测定:取10m L 海水样品置于离心管中,加入50μL 50%的浓硫酸,常温避光保存㊂D M S P 的测定采用D M S P 在强碱环境(p Hȡ13)中完全裂解为D M S 的原理进行间接分析测定㊂主要步骤为:取2m L 保存的海水样品,加入200μL 10m o l /L的N a O H 溶液,4ħ下密闭保存24h,待其碱解完全后测定D M S 含量,并等比例换算为D M S P 浓度㊂2 结果与讨论2.1扇贝养殖季烟台近海C H 4的时空分布特征及影响因素调查海域不同月份表层和底层海水中的C H 4浓度及其变化范围如表1所示㊂整体而言,烟台近海的C H 4浓度高于渤海[14]㊁黄海及东海,低于桑沟湾养殖区[15],5㊁6㊁7三个月份的C H 4浓度明显高于8㊁9㊁11三个月份,并且各个月份的底层海水C H 4浓度均高于表层㊂如图2所示,调查海域C H 4浓度的高值区主要集中在近岸海域,最高值通常出现在S 10站位和S 11站位㊂7㊁8月份表层和底层的C H 4分布情况略有不同,表层高值区主要集中在调查海域西侧的S 5㊁S 6站位㊂78中 国 海 洋 大 学 学 报2024年图2 扇贝养殖季烟台近海C H 4(n m o l /L )的时空分布F i g .2 T e m p o r a l a n d s p a t i a l d i s t r i b u t i o n s o f C H 4(n m o l /L )i n t h e c o a s t a l w a t e r s o f Y a n t a i d u r i n g t h e s c a l l o p f a r m i n g p e r i o d 表1 烟台近海不同月份表层和底层海水中C H 4、D M S 和D M S P 浓度T a b l e 1 C H 4DM S a n d D M S P c o n c e n t r a t i o n s i n t h e s u r f a c e a n d b o t t o m s e a w a t e r o f Y a n t a i o f f s h o r e i n d i f f e r e n t m o n t h sn m o l /L 采样时间T i m e 表层S u r f a c e 底层B o t t o mC H 4D M SD M S P C H 4D M SD M S P5月M a y 14.93(12.02~21.67)7.86(4.00~15.71)28.96(16.30~36.60)17.51(11.71~22.10)8.24(2.82~21.49)15.90(9.86~35.75)6月J u n e 15.32(9.51~33.68)18.80(9.68~32.12)55.84(26.27~95.58)23.71(11.21~44.35)6.91(2.26~23.51)24.21(5.62~66.24)7月J u l y 12.10(3.34~37.53)18.29(3.43~39.41)38.04(25.33~68.79)18.40(5.53~38.53)8.86(2.30~25.79)17.47(7.52~47.99)8月A u g .6.07(4.22~9.03)8.53(3.52~18.64)19.97(3.85~39.47)6.73(5.02~12.92)7.81(0.81~16.20)25.13(12.15~54.70)9月S e p t .6.41(3.27~12.17)3.56(2.65~5.24)11.75(7.75~15.39)7.50(4.17~12.54)2.44(1.60~4.13)8.81(3.25~18.31)11月N o v .9.21(5.25~24.96)1.24(0.94~1.71)9.81(3.64~15.39)11.64(5.48~26.06)1.09(0.64~1.46)8.97(3.27~13.86) 在整个扇贝养殖周期内,调查海域C H 4与DO ㊁C h l a 之间不存在明显的相关性,这与谭丹丹等[14]在黄渤海的调查结果一致,表明在烟台近海D O 和C h l a 可能不是控制海水中C H 4浓度的主要因素㊂通过对比养殖海域与非养殖海域发现,养殖区底层平均C H 4浓度为15.73n m o l /L ,非养殖区底层平均C H 4浓度为12.52n m o l /L ,养殖区底层C H 4平均浓度比非养殖区高25.64%(见图3A )㊂滤食性贝类可以将海水中的有881期张升辉,等:扇贝养殖季烟台近海C H 4和DM S 的时空分布及影响因素机物摄入体内,而其中大部分未被摄取,并以粪便的形式排出体外,间接将水中的大量有机物质输入沉积物中,同时,筏式养殖也可以在一定程度上减缓水体的流动,加快颗粒物质的沉降,导致水产养殖区沉积物中相对较高的碳㊁氮㊁磷含量,这可能是养殖区底层C H 4平均浓度明显高于非养殖区的原因㊂养殖区表层平均C H 4浓度为9.65n m o l /L ,非养殖区表层平均C H 4浓度为11.91n m o l /L ,与底层相反,养殖区表层C H 4平均浓度比非养殖区低23.42%㊂通过图3D 可以看出,养殖区表层C H 4浓度低于非养殖区的时间集中在海水温度比较高的6 9月㊂在此期间,养殖海域层化现象显著,并且多次出现季节性缺氧现象[16]㊂通过调查期间底层D O 的浓度也可以看出,7㊁8和9月扇贝养殖区底层海水的D O 浓度要低于非养殖区㊂因此,推断是由于养殖海域海水的层化减慢了C H 4由底层向表层的扩散,增加了C H 4在水体的保留时间,导致6 9月养殖区表层C H 4平均浓度反而低于非养殖区㊂张福绥等[17]通过对海湾扇贝肥满度的研究发现,扇贝生长一年有春㊁秋两个高峰,春季为5月中旬 7月中旬,盛期为6月,秋季为9 10月,盛期为9月,最佳收获期为养殖当年的11 12月㊂通过对比扇贝不同生长时期C H 4浓度可以看出,扇贝收获季(11月)养殖区底层C H 4浓度比非养殖区高66%,这一比例明显高于扇贝生长盛期(31%和7%),这表明成熟扇贝可以更高效的将水中的有机物质输入到沉积物中,更利于甲烷的生产释放㊂图3 烟台近海养殖区与非养殖区D M S ,D M S P 及C H 4浓度平均值对比F i g .3 C o m p a r i s o n o f a v e r a g e D M S ,D M S P ,a n d C H 4co n c e n t r a t i o n s b e t w e e n s c a l l o p f a r m i n g a r e a a n d n o n -s c a l l o p f a r m i n g a r e a 2.2扇贝养殖季烟台近海D M S 、D M S P 的时空分布特征及影响因素调查海域不同月份表层和底层海水中的D M S㊁D M S P 浓度及其变化范围如表1所示㊂与C H 4相同,6月份的D M S ㊁D M S P 平均浓度最高,春夏两季的D M S㊁D M S P 平均浓度明显高于秋季,并且不同月份不同水层的D M S ㊁D M S P 分布趋势不尽相同(见图4)㊂浮游植物生物量是影响海水D M S 和D M S P 浓度的关键因素之一,并且多个研究已表明我国渤海㊁黄海及东海海域D M S ㊁D M S P 与C h l a 存在显著的正相关关系[3,5]㊂98中 国 海 洋 大 学 学 报2024年然而,在烟台近岸海域,整个扇贝养殖周期内并未发现D M S ㊁D M S P 与C h l a 之间存在显著的正相关关系,这表明影响近岸水体中D M S 和D M S P 迁移转化过程的因素更为复杂㊂图4 扇贝养殖季烟台近海D M S (n m o l /L )㊁D M S P (n m o l /L)的时空分布F i g .4 T e m p o r a l a n d s pa t i a l d i s t r ib u t i o n s o f D M S (n m o l /L )a n d D M S P (n m o l /L )i n t h ec o a s t a l w a t e r s o f Y a n t a id u r i n g t he s c a l l o pf a r m i ng pe r i o d 091期张升辉,等:扇贝养殖季烟台近海C H4和D M S的时空分布及影响因素养殖区和非养殖区不同月份C h l a㊁D M S㊁D M S P 的平均浓度分别如图3B㊁图3C㊁图3D所示,整个扇贝养殖期间内养殖区表层水体C h l a㊁D M S㊁D M S P的平均浓度分别为1.00μg/L㊁10.70n m o l/L和25.99 n m o l/L,非养殖区C h l a㊁D M S㊁D M S P的平均浓度分别1.68μg/L㊁8.62n m o l/L和28.97n m o l/L㊂与非养殖区相比,养殖区的C h l a和D M S P浓度分别降低了68%和11%,而D M S浓度增加了24%㊂贝类的生物过滤作用对水体中浮游生物及颗粒物质有着巨大的影响,在胶州湾,养殖贝类对浮游植物的消耗量占该生态系统浮游植物总消耗量的90%[18]㊂同时,K a s a m a t s u 等[19]的研究表明,贝类摄取的浮游植物及有机颗粒物中D M S P大部分并未参与代谢过程,而是以粪便D M-S P的形式排除㊂因此虽然养殖区的C h l a浓度降低了68.00%,而D M S P浓度仅仅降低了11.47%㊂另外,侯兴等[20]的研究表明,贝类粪便中的D M S P可以扩散到海水中,并导致水体中悬浮颗粒态D M S P的增加㊂D M S P作为微生物食物网中的关键化合物[21],可以为细菌的生长提供所需的硫源和碳源,贝类粪便中的D M S P可以吸引附着细菌,增加D M S P向D M S的转化,进而导致养殖区表层水体的D M S浓度高于非养殖区㊂通过计算不同月份扇贝养殖区和非养殖区的D M S㊁D M S P浓度得出,5月养殖区的D M S浓度比非养殖区高99%,D M S P浓度低12%,6月养殖区的D M S浓度比非养殖区高12%,D M S P浓度低5%,7月养殖区的D M S浓度比非养殖区高26%,D M S P浓度低8%,9月养殖区的D M S浓度比非养殖区高28%,D M-S P浓度低13%,11月养殖区的D M S浓度比非养殖区低34%,D M S P浓度低36%㊂综合整个扇贝养殖周期和不同月份养殖区和非养殖区的D M S㊁D M S P浓度差异,可以看出扇贝养殖对D M S释放的促进作用主要集中在其生长繁殖期,扇贝成熟后,其滤食能力的增强会显著降低表层水体的D M S P浓度,反而不利于D M S的生产释放㊂2.3D M S、D M S P与C H4的相关关系海洋中大部分C H4是在厌氧环境条件下,氧气和无机氧化剂(N O-3,F e2+,S O2-4)耗尽环境中作为电子给予体而形成的氧化产物[22]㊂因此,对海洋C H4循环的研究主要集中于缺氧的沉积物中㊂然而,诸多调查研究发现,部分海域的大洋区㊁陆架区和海湾区真光层海水中也普遍存在不同程度的C H4过饱和状态,如中国的黄东海[4]㊁南海北部陆坡[23],智利中部大陆架[24],亚热带北太平洋[25],黑海[26],波罗的海[27],北极水域[28]等㊂近年来,关于导致表层水体 C H4悖论 的成因众说纷纭,而越来越多的证据表明有氧环境下的C H4生成可能与海洋中含硫的甲基化合物密切相关㊂A l t h o f f等[29]的研究表明有机硫化物可在高氧条件下被铁(Ⅱ/Ⅲ)转化为C H4;D a m m等[30]在斯瓦尔巴特岛南部陆架S t o r f j o r d e n地区的调查结果显示D M S P 可以作为甲基营养型产C H4过程中潜在的营养基质,本次调查研究发现一些站位表层海水中的D M S P与C H4浓度高值高度重合且其表层C H4浓度高于底层,例如7月份S6㊁S7站位,这表明研究海域表层水体中存在C H4的其他来源㊂通过对整个扇贝养殖周期内的D M S㊁D M S P与C H4进行相关性分析,发现表层水体中D M S P与C H4呈现显著正相关关系(见图5),而D M S与C H4之间不存在显著的相关性,该结果与Z i n-d l e r[31]在日本至澳大利亚的一条南北通道表层水体中的调查结果一致㊂尽管D a m m等[30]指出D M S P降解过程中产生的D M S也可能是生成C H4的潜在前体物质,耿澜涛[32]也发现表层海水中可溶有机质和D M S在光照条件下能产生C H4,并且C H4的产量和生成速率与可光降解物质的光照时长和浓度密切相关,但调查结果显示在太阳辐射最强的7㊁8月份,烟台近海D M S 与C H4的高值区并不吻合,并且D M S与C H4之间不存在显著的正相关关系㊂这表明7月份S6㊁S7站位C H4的高值可能来源于D M S P而非D M S,但关于他们之间的转换机制㊁途径和影响因素还需要进一步的深入研究㊂图5 D M S P与C H4的相关关系F i g.5C o r r e l a t i o n b e t w e e n D M S P a n d C H42.4扇贝养殖季烟台近海C H4和D M S的海-气通量海水中溶存C H4的饱和度R和海-气通量F计算公式如下:R=C o b s/C e q㊂(1)F=K wˑ(C o b s-C e q)㊂(2)式中:C o b s为C H4在表层海水中的实测浓度;C e q为气体在表层海水与大气达到平衡时的浓度,根据W i e s e n-19中国海洋大学学报2024年b u r g公式计算得出;K w为气体交换速率㊂D M S的海-气通量计算公式为:F=K(c w-c g/H)ʈK c w㊂(3)式中:K为海-气交换速率;c w和c g分别为D M S在大气和海水中的浓度;H为亨利常数;c g/H远小于c w,可忽略不计㊂C H4和D M S的海-气交换速率均采用W2014法[33]计算㊂计算结果表明烟台近海5㊁6㊁7㊁8㊁9和11月C H4饱和度平均值分别为606.2%㊁691.2%㊁588.8%㊁284.4%㊁284.2%㊁331.8%,海-气通量平均值分别为10.80㊁9.26㊁5.65㊁2.15㊁2.60和5.60μm o l/(m2㊃d),D M S为5.76㊁11.33㊁7.87㊁4.07㊁1.89和1.01μm o l/ (m2㊃d)㊂通过B i g e m a p G I S O f f i c e软件估算调查海域面积约为560k m2,大致估算出烟台近海扇贝养殖季的甲烷海-气通量为9.69ˑ106g,D M S海-气通量为3.33ˑ107g㊂3结论(1)烟台近岸海域表层海水C H4浓度处于过饱和状态,D M S P可能是表层海水中C H4重要的潜在来源㊂尽管扇贝养殖区底层C H4平均浓度比非养殖区高25.64%,但是由于夏季养殖区海水层化较为严重,增加了C H4在海水中的氧化,导致养殖区表层C H4浓度反而低于养殖区,整个扇贝养殖周期内调查海域甲烷的海-气通量约为9.69ˑ106g㊂(2)扇贝养殖可以影响D M S和D M S P的迁移转化过程,贝类滤食过程可以将浮游植物体内的D M S P转化为海水中的颗粒态D M S P,增加D M S P向D M S的转化㊂与非养殖区相比,养殖区的D M S P浓度降低了11.47%,D M S浓度增加了24.13%㊂整个扇贝养殖周期内调查海域D M S的海-气通量约为3.33ˑ107g㊂致谢:本文所用的D O和C h l a数据由中科院牟平海岸带环境综合试验站提供,作者对此表示诚挚谢意㊂参考文献:[1] L a n a A,B e l l T G,S i móR,e t a l.A n u p d a t e d c l i m a t o l o g y o fs u r f a c e d i m e t h l y s u l f i d e c o n c e n t r a t i o n s a n d e m i s s i o n f l u x e s i n t h eg l o b a l o c e a n[J].G l o b a l B i o g e o c h e m i c a l C y c l e s,d o i:10.1029/2010G B003850.[2]臧家业.东海海水中的溶存甲烷[J].海洋学报,1998,20(2),52-59.Z a n g J Y.D i s s o l v e d m e t h a n e i n s e a w a t e r o f t h e E a s t C h i n a S e a[J].A c t a O c e a n o l o g i c a S i n i c a,1998,20(2):52-59. [3]Z h a n g S H,S u n J,L i u J L,e t a l.S p a t i a l d i s t r i b u t i o n s o f d i m e t h y ls u l f u r c o m p o u n d s,D M S P-l y a s e a c t i v i t y,a n d p h y t o p l a n k t o n c o m-m u n i t y i n t h e E a s t C h i n a S e a d u r i n g f a l l[J].B i o g e o c h e m i s t r y, 2017,133(1):59-72.[4]张桂玲.中国近海部分海域溶解甲烷和氧化亚氮的生物地球化学过程[D].青岛:中国海洋大学,2004.Z h a n g G L.S t u d y o n B i o g e o c h e m i s t r y o f D i s s o l v e d M e t h a n e a n d N i t r o u s O x i d e i n t h e C o a s t a l W a t e r o f C h i n a[D].Q i n g d a o:O c e a n U n i v e r s i t y o f C h i n a,2004.[5]Z h a n g S H,Y a n g G P,Z h a n g H H,e t a l.S p a t i a l v a r i a t i o n o f b i o-g e n i c s u l f u r i n t h e s o u t h Y e l l o w S e a a n d t h e E a s t C h i n a S e a d u r i n g s u m m e r a n d i t s c o n t r i b u t i o n t o a t m o s p h e r i c s u l f a t e a e r o s o l[J].S c i e n c e o f t h e T o t a l E n v i r o n m e n t,2014,488-489:157-167. [6]Z h a i X,Z h a n g H H,Y a n g G P,e t a l.D i s t r i b u t i o n a n d s e a-a i r f l u-x e s o f b i o g e n i c g a s e s a n d r e l a t i o n s h i p s w i t h p h y t o p l a n k t o n a n d n u-t r i e n t s i n t h e c e n t r a l b a s i n o f t h e S o u t h C h i n a S e a d u r i n g s u m m e r [J].M a r i n e C h e m i s t r y,2018,200:33-44.[7]马立杰,崔迎春.南海中部和北部上层海水中溶存甲烷浓度及海气交换通量[J].热带海洋学报,2013,32(2):94-101.M a L J,C u i Y C.D i s s o l v e d m e t h a n e c o n c e n t r a t i o n a n d s e a-t o-a i r t r a n s f e r f l u x o f d i s s o l v e d m e t h a n e i n t h e u p p e r s e a w a t e r o f t h e c e n-t r a l a n d n o r t h e r n S o u t h C h i n a S e a[J].J o u r n a l o f T r o p i c a l O c e a-n o g r a p h y,2013,32(2):94-101.[8]Z h a n g G,Z h a n g J,L i u S,e t a l.M e t h a n e i n t h e C h a n g j i a n g(Y a n-g t z e R i v e r)E s t u a r y a n d i t s a d j a c e n t m a r i n e a r e a:R i v e r i n e i n p u t, s e d i m e n t r e l e a s e a n d a t m o s p h e r i c f l u x e s[J].B i o g e o c h e m i s t r y, 2008,91(1):71-84.[9]J i a n S,Z h a n g H H,Y a n g G P,e t a l.V a r i a t i o n o f b i o g e n i c d i m e-t h y l a t e d s u l f u r c o m p o u n d s i n t h e C h a n g j i a n g R i v e r E s t u a r y a n d t h e c o a s t a l E a s t C h i n a S e a d u r i n g s p r i n g a n d s u m m e r[J].J o u r n a l o fM a r i n e S y s t e m s,2019,199:103222.[10]顾培培,张桂玲,李佩佩,等.调流调沙对黄河口及邻近海域溶存甲烷的影响[J].中国环境科学,2011,31(11):1821-1828.G u P P,Z h a n g G L,L i P P,e t a l.E f f e c t o f t h e w a t e r-s e d i m e n tr e g u l a t i o n o n d i s s o l v e d m e t h a n e i n t h e l o w e r Y e l l o w R i v e r e s t u a r ya n d i t s a d j a c e n t m a r i n e a r e a[J].C h i n a E n v i r o n m e n t a l S c i e n c e,2011,31(11):1821-1828.[11]B a n g e H W,B a r t e l l U H,R a p s o m a n i k i s S,e t a l.M e t h a n e i n t h eB a l t i c a n d N o r t h S e a s a n d a r e a s s e s s m e n t o f t h e m a r i n e e m i s s i o n so f m e t h a n e[J].G l o b a l B i o g e o c h e m i c a l C y c l e s,1994,8(4):465-480.[12] Y a n g B,G a o X,Z h a o J,e t a l.T h e i n f l u e n c e o f s u m m e r h y p o x i ao n s e d i m e n t a r y p h o s p h o r u s b i o g e o c h e m i s t r y i n a c o a s t a l s c a l l o pf a r m i ng a r e a,N o r th Y e l l o w S e a[J].S ci e n c e o f t h e T o t a l E n v i-r o n m e n t,2020,759(1):143486.[13]陈慧娟.海水养殖对海洋生态环境的影响及其对策[J].福建水产,2007(3):47-50.C h e n H J.T h e i m p a c t o f m a r i c u l t u r e o n m a r i n e e c o l o g i c a l e n v i-r o n m e n t a n d i t s c o u n t e r m e a s u r e s[J].J o u r n a l o f F u j i a n F i s h e r i e s, 2007(3):47-50.[14]谭丹丹,张洪海,翟星,等.冬季黄渤海D M S,D M S P和C H4的分布及影响因素[J].中国环境科学,2019,39(5):2143-2153.T a n D D,Z h a n g H H,Z h a i X,e t a l.D i s t r i b u t i o n s o f d i m e t h y l-s u l f i d e,d i m e t h y l s u l f o n i o p r o p i o n a t e,m e t h a n e a n d i n f l u e n c i n g f a c-t o r s i n t h e Y e l l o w S e a a n d B o h a i S e a d u r i n g w i n t e r[J].C h i n a E n-v i r o n m e n t a l S c i e n c e,2019,39(5):2143-2153.[15]孙明爽.东㊁黄海及桑沟湾海域溶解甲烷的分布㊁释放和产生研究[D].青岛:中国海洋大学,2014.S u n M S.D i s t r i b u t i o n,E m i s s i o n a n d P r o d u c t i o n o f M e t h a n e i n291期张升辉,等:扇贝养殖季烟台近海C H4和D M S的时空分布及影响因素t h e Y e l l o w S e a,E a s t C h i n a S e a a n d S a n g g o u B a y[D].Q i n g d a o: O c e a n U n i v e r s i t y o f C h i n a,2014.[16]张福绥,马江虎,何义朝,等.胶州湾海湾扇贝肥满度的研究[J].海洋与湖沼,1991,22(2):97-103.Z h a n g F S,M a J H,H e Y C,e t a l.A s t u d y o n t h e m e a t c o n d i-t i o n o f t h e b a y s c a l l o p i n J i a o z h o u B a y[J].O c e a n o l o g i a e t L i m n o-l o g i a S i n i c a,1991,22(2):97-103.[17]张文静,董志军,孙西艳,等.2018年烟台四十里湾及其邻近海域网采浮游植物群落结构的时空变化[J].海洋与湖沼,2020,51(5):1151-1161.Z h a n g W J,D o n g Z J,S u n X Y,e t a l.S p a t i a l a n d t e m p o r a l d i s-t r i b u t i o n o f n e t-s a m p l e d p h y t o p l a n k t o n c o m m u n i t y i n S i s h i l i B a ya n d i t s a d j a c e n t w a t e r,Y a n t a i i n2018[J].O c e a n o l o g i a e t L i m n o-l o g i a S i n i c a,2020,51(5):1151-1161.[18] H a n D,C h e n Y,Z h a n g C,e t a l.E v a l u a t i n g i m p a c t s o f i n t e n s i v es h e l l f i s h a q u a c u l t u r e o n a s e m i-c l o s e d m a r i n e e c o s y s t e m[J].E c o-l o g i c a l M o d e l l i n g,2017,359:193-200.[19] K a s a m a t s u N,K a w a g u c h i S,W a t a n a b e S,e t a l.P o s s i b l e i m p a c t so f z o o p l a n k t o n g r a z i n g o n d i m e t h y l s u l f i d e p r o d u c t i o n i n t h e A n t-a r c t i c O c e a n[J].C a n a d i a n J o u r n a l o f F i s h e r i e s a n d A q u a t i c S c i-e n c e s,2004,61(5):736-743.[20]侯兴,王颖,刘天红,等.4种常见经济滤食性贝类摄食活动对球等鞭金藻产生二甲基硫化物的影响[J].渔业科学进展,2021,42(5):124-131.H o u X,W a n g Y,L i u T H,e t a l.E f f e c t s o f f e e d i n g a c t i v i t i e s b yf o u r c o m m o n e c o n o m i c b i v a l v e s o n t h e p r o d u c t i o n o f d i m e t h y l s u l-f u r c o m p o u n d s f r o m I s o c h r y s i sg a l b a n a[J].P r o g r e s s i n F i sh e r yS c i e n c e s,2021,42(5):124-131.[21]S t e f e l s J,S t e i n k e M,T u r n e r S,e t a l.E n v i r o n m e n t a l c o n s t r a i n t so n t h e p r o d u c t i o n a n d r e m o v a l o f t h e c l i m a t i c a l l y a c t i v e g a s d i m e-t h y l s u l p h i d e(D M S)a n d i m p l i c a t i o n s f o r e c o s y s t e m m o d e l l i n g[J].B i o g e o c h e m i s t r y,2007,83(1):245-275.[22]陈烨,孙治雷,吴能友,等.海洋沉积物中甲烷代谢微生物的研究进展[J].海洋地质与第四纪地质,2022,42(6):82-92.C h e n Y,S u n Z L,W u N Y,e t a l.A d v a n c e s i n t h e s t u d y o fm e t h a n e-m e t a b o l i z i n g m i c r o b i a l c o m m u n i t i e s i n m a r i n e s e d i-m e n t s[J].M a r i n e G e o l o g y&Q u a t e r n a r y G e o l o g y,2022,42(6):82-92.[23]Z h o u H,Y i n X,Y a n g Q,e t a l.D i s t r i b u t i o n,s o u r c e a n d f l u x o fm e t h a n e i n t h e w e s t e r n P e a r l R i v e r E s t u a r y a n d n o r t h e r n S o u t hC h i n a S e a[J].M a r i n e C h e m i s t r y,2009,117(1-4):21-31.[24]F l o r e z-L e i v a L,D a m m E,F a ría s L.M e t h a n e p r o d u c t i o n i n d u c e db y d i m e t h y l s u l f i d e i n s u r f ac e w a t e r o f a n u p w e l l i n g e c o s y s t e m[J].P r o g r e s s i n O c e a n o g r a p h y,2013,112-113:38-48. [25] H o l m e s M E,S a n s o n e F J,R u s t T M,e t a l.M e t h a n e p r o d u c-t i o n,c o n s u m p t i o n,a n d a i r-s e a e x c h a n g e i n t h e o p e n o c e a n:A ne v a l u a t i o n b a s e d o n c a r b o n i s o t o p i c r a t i o s[J].G l o b a l B i o g e o-c h e m i c a l C y c l e s,2000,14(1):1-10.[26]S c h m a l e O,B e a u b i e n S E,R e h d e r G,e t a l.G a s s e e p a g e i n t h eD n e p r p a l e o-d e l t a a r e a(N W-B l a c k S e a)a n d i t s r e g i o n a l i m p a c t o nt h e w a t e r c o l u m n m e t h a n e c y c l e[J].J o u r n a l o f M a r i n e S y s t e m s, 2010,80(1-2):90-100.[27]S c h m a l e O,Wäg e J,M o h r h o l z V,e t a l.T h e c o n t r i b u t i o n o f z o o-p l a n k t o n t o m e t h a n e s u p e r s a t u r a t i o n i n t h e o x y g e n a t e d u p p e r w a-t e r s o f t h e c e n t r a l B a l t i c S e a[J].L i m n o l o g y a n d O c e a n o g r a p h y, 2018,63(1):412-430.[28] S h a k h o v a N,S e m i l e t o v I,S a l y u k A,e t a l.E x t e n s i v e m e t h a n ev e n t i n g t o t h e a t m o s p h e r e f r o m s e d i m e n t s o f t h e E a s t S i b e r i a nA r c t i c S h e l f[J].S c i e n c e,2010,327(5970):1246-1250.[29] A l t h o f f F,B e n z i n g K,C o m b a P,e t a l.A b i o t i c m e t h a n o g e n e s i sf r o m o rg a n o s u l ph u r c o m p o u n d s u n d e r a m bi e n t c o n d i t i o n s[J].N a t u r e C o m m u n i c a t i o n s,2014,5(1):1-9.[30] D a m m E,K i e n e R P,S c h w a r z J,e t a l.M e t h a n e c y c l i n g i n A r c t i cs h e l f w a t e r a n d i t s r e l a t i o n s h i p w i t h p h y t o p l a n k t o n b i o m a s s a n dD M S P[J].M a r i n e C h e m i s t r y,2008,109(1-2):45-59.[31] Z i n d l e r C,B r a c h e r A,M a r a n d i n o C A,e t a l.S u l p h u r c o m-p o u n d s,m e t h a n e,a n d p h y t o p l a n k t o n:I n t e r a c t i o n s a l o n g an o r t h-s o u t h t r a n s i t i n t h e w e s t e r n P a c i f i c O c e a n[J].B i o g e o-s c i e n c e s,2013,10(5):3297-3311.[32]耿澜涛.加拿大北极亚北极海水中溶解甲烷的分布及其生物地球化学研究[D].武汉:中国地质大学,2017.G e n g L T.S t u d i e s o n t h e D i s t r i b u t i o n o f D i s s o l v e d M e t h a n e a n dI t s B i o g e o c h e m i s t r y i n C a n a d i a n A r c t i c a n d S u b-A r c t i c S e a s[D].W u h a n:C h i n a U n i v e r s i t y o f G e o s c i e n c e s,2017.[33] W a n n i n k h o f R.R e l a t i o n s h i p b e t w e e n w i n d s p e e d a n d g a s e x-c h a n g e o v e r t h e o c e a n r e v i s i t e d[J].L i m n o l o g y a nd O ce a n o g r a-p h y:M e t h o d s,2014,12(6):351-362.3949中国海洋大学学报2024年T e m p o-S p a t i a l D i s t r i b u t i o n s a n d I n f l u e n c i n g F a c t o r s o f C H4a n dD M S D u r i n g S c a l l o p F a r m i n g i n Y a n t a i O f f s h o r eZ h a n g S h e n g h u i1,S u n J i n g2,Z h a o M i a o3,4,H u X i a o k e1(1.K e y L a b o r a t o r y o f C o a s t a l B i o l o g y a n d B i o r e s o u r c e U t i l i z a t i o n,Y a n t a i I n s t i t u t e o f C o a s t a l Z o n e R e s e a r c h,C h i n e s e A c a d-e m y of S c i e n c e s,Y a t a i264006,C h i n a;2.S h a n d o ng M a r i n e R e s o u r c e a n d E n v i r o n m e n t R e s e a r ch I n s ti t u t e,S h a n d o n g K e y L a b o r a t o r y o f M a r i n e E c o l o g i c a l R e s t o r a t i o n,Y a n t a i264006,C h i n a;3.J i a n g s u K e y L a b o r a t o r y o f M a r i n e B i o r e s o u r c e s a n d E n v i r o n m e n t,J i a n g s u O c e a n U n i v e r s i t y,L i a n y u n g a n g222005,C h i n a;4.J i a n g s u I n s t i t u t e o f M a r i n e R e s o u r c e s D e v e l o p-m e n t,L i a n y u n g a n g222005,C h i n a)A b s t r a c t: M e t h a n e(C H4)a n d d i m e t h y l s u l f i d e(D M S)a r e i m p o r t a n t v o l a t i l e b i o g e n i c a c t i v e g a s e s, w h i c h p l a y i m p o r t a n t r o l e s i n r e g u l a t i n g c l i m a t e c h a n g e.I n o r d e r t o e x p l o r e t h e i m p a c t o f r a f t c u l t u r e o f s c a l l o p o n C H4a n d D M S,f i e l d i n v e s t i g a t i o n s w e r e c o n d u c t e d i n Y a n t a i o f f s h o r e d u r i n g M a y,J u n e, J u l y,A u g u s t,S e p t e m b e r a n d N o v e m b e r i n2019,r e s p e c t i v e l y.T h e r e s u l t s s h o w e d t h a t s c a l l o p f i l t e r f e e d i n g c o u l d i n d i r e c t l y i n p u t o r g a n i c m a t t e r i n t o s e d i m e n t,a n d r e s u l t i n g i n a n25.64%h i g h e r a v e r a g e C H4c o n c e n t r a t i o n i n t h e b o t t o m l a y e r o f s c a l l o p f a r m i n g a r e a c o m p a r e d w i t h t h a t i n n o n-c u l t u r e a r e a. M e a n w h i l e,s c a l l o p f i l t e r f e e d i n g c o u l d a l s o r e p a c k a g e t h e p h y t o p l a n k t o n i c D M S P i n t o f e c a l D M S P,a n d t h e n d e g r a d e D M S P i n t o D M S b y a t t r a c t i n g a t t a c h e d b a c t e r i a.C o m p a r e d w i t h t h e n o n-f a r m i n g a r e a,t h e D M S c o n c e n t r a t i o n i n t h e s u r f a c e l a y e r o f t h e f a r m i n g a r e a i n c r e a s e d b y24.13%.I n a d d i t i o n,t h e C H4 c o n c e n t r a t i o n s i n s u r f a c e s e a w a t e r w e r e s u p e r s a t u r a t e d,a n d D M S P m a y b e a n i m p o r t a n t p o t e n t i a l s o u r c e o f C H4i n t h e s u r f a c e s e a w a t e r.Y a n t a i o f f s h o r e i s a n i m p o r t a n t s o u r c e o f a t m o s p h e r i c C H4a n d D M S.D u r i n g t h e w h o l e s c a l l o p b r e e d i n g c y c l e,t h e s e a-a i r f l u x e s o f C H4a n d i n t h e i n v e s t i g a t e d s e a a r e a w a s a b o u t9.69ˑ106a n d3.33ˑ107g,r e s p e c t i v e l y.K e y w o r d s:C H4;D M S;D M S P;s h e l l f i s h f a r m i n g责任编辑徐环。
皮质醇对鱼类性别分化过程的影响及调控机制研究进展

第38卷第4期大连海洋大学学报Vol.38No.4 2023年8月JOURNAL OF DALIAN OCEAN UNIVERSITY Aug.2023DOI:10.16535/ki.dlhyxb.2022-293文章编号:2095-1388(2023)04-0726-11皮质醇对鱼类性别分化过程的影响及调控机制研究进展高蕊1,2,3,闫红伟1,2,3∗,刘鹰2,4,刘奇1,2(1.大连海洋大学海洋科技与环境学院,辽宁大连116023;2.设施渔业教育部重点实验室(大连海洋大学),辽宁大连116023;3.辽宁省河鲀良种繁育及健康养殖重点实验室,辽宁大连116023;4.浙江大学生物系统工程与食品科学学院,浙江杭州310058)摘要:鱼类的性别决定与分化一直是发育生物学㊁繁殖生理学及鱼类遗传育种学领域的研究热点,与高等脊椎动物不同,鱼类的性别决定与分化具有较强的可塑性,容易受到许多环境因素如温度㊁种群密度和pH等的影响㊂探究鱼类的性别决定与分化的分子机制对鱼类性别控制育种至关重要,也有利于提高鱼类养殖效益㊂皮质醇(cortisol)作为鱼类最主要的糖皮质激素,在鱼类性别分化过程中具有重要作用,本文综述了鱼类性别分化与性转变过程中,外源皮质醇影响鱼类的性别分化㊁环境因素影响内源皮质醇水平的变动规律,以及皮质醇调控性别分化与性转变的分子机制,并提出未来应在皮质醇诱导鱼类雄性化的作用机制㊁皮质醇与表观遗传因子的互作关系等方面重点开展研究,以期为深入探究皮质醇的功能特性及调控机理提供科学参考㊂关键词:皮质醇;鱼类性别分化;环境因子;分子机制中图分类号:S917.4㊀㊀㊀㊀文献标志码:A㊀㊀性别决定(sex determination)是性别形成的生物学过程,是指未分化的具有双向潜能的性腺决定其向精巢或卵巢方向发育的过程[1]㊂性别分化(sex differentiation)以性别决定为前提,是未分化的性腺在性别确定后,发育为精巢或卵巢的过程[2]㊂鱼类是脊椎动物中种类最大的类群,在脊椎动物系统演化过程中具有承前启后的地位㊂与鸟类㊁哺乳动物等脊椎动物相比,鱼类的性别决定与分化过程更为复杂,其具有原始性㊁多样性和易变性的特点,既受到遗传因素的作用,又受某些外部环境因素如温度[3]㊁光照[4]㊁pH[5]和种群密度[6]等的影响㊂因此,揭示鱼类的性别决定与分化机制对理解脊椎动物性别形成过程具有重要的理论意义㊂更为重要的是,许多鱼类在生长㊁繁殖和形态上存在雌雄差异㊂在养殖生产上,如果对某些鱼类开展性别控制和单性养殖,可大幅提高经济效益,故阐明鱼类性别决定及分化机制在育种方面具有重要的应用价值㊂性类固醇激素在鸟类㊁爬行动物㊁两栖类动物及鱼类等的性别分化中扮演着重要角色㊂1953年,Yamamoto等[7]首次用雌激素处理青鳉(Oryzias latipes),获得了性反转的功能性雌性个体㊂自此,国内外研究人员逐步开展了激素诱导鱼类性别转变的研究㊂17β-雌二醇(17β-estradiol,E2)是硬骨鱼类卵巢发育和雌性性别维持所必需的雌激素,在性别决定期间,经E2处理能造成斑马鱼(Danio rerio)[8]㊁尼罗罗非鱼(Oreochromis niloticus)[9]等许多鱼类雌性化㊂11-酮基睾酮(11-ketotestoster-one,11-KT)是鱼类最主要的雄激素,经11-KT处理会造成蜂巢石斑鱼(Epinephelus merra)[10]㊁条纹锯鮨(Centropristis striata)[11]等许多鱼类雄性化㊂皮质醇(cortisol)既是鱼类主要的糖皮质激素(glucocorticoid,GC),也是一种类固醇激素,其被认为是连接外部环境刺激与内部生理反应的关键因㊀收稿日期:2022-09-28㊀基金项目:国家自然科学基金青年基金(31902347);国家海水鱼产业技术体系资助(CARS-47);辽宁省科技厅面上项目(2022-MS-351);辽宁省教育厅科研项目(LJKMZ20221092,LJKZ0712);大连市重点领域创新团队项目(2021RT07);辽宁省科技特派团项目(2022JH5/10400070)㊀作者简介:高蕊(1998 ),女,硕士研究生㊂E-mail:gaorui199809@㊀通信作者:闫红伟(1985 ),女,博士,副教授㊂E-mail:yanhongwei@子[12]㊂当鱼体受到外界应激因子刺激后,下丘脑-垂体-肾间组织(hypothalamus-pituitary-interrenal, HPI)轴会迅速做出反应,由下丘脑释放促肾上腺皮质激素释放激素(corticotropin-release hormone, CRH)作用于垂体,刺激垂体前叶促肾上腺皮质激素细胞分泌促肾上腺皮质激素(adrenocortico-tropic hormone,ACTH),ACTH刺激肾间细胞分泌皮质醇激素,并作用于全身各个靶器官[13]㊂近年来研究发现,皮质醇可能介导了环境因素影响鱼类性别分化的过程㊂在一些鱼类中,外源皮质醇处理会造成基因型为雌性的个体雄性化,而在皮质醇处理时,使用雌激素或皮质醇合成抑制剂在一定程度上能回救上述皮质醇所造成的雄性化[14-15]㊂研究还发现,高温㊁高密度养殖等环境因素会造成一些鱼类皮质醇水平增加,进而导致鱼类个体雄性化[16-17]㊂在雌雄同体的鱼类中,由于种群社会结构变化所造成的性转变往往伴随着皮质醇水平的改变㊂目前,皮质醇作用于鱼类性别分化与性转变的具体机制尚不明确,故探讨皮质醇对鱼类性别分化的影响并厘清其调控机制,有助于认知外部环境因素影响鱼类性别的作用途径及性别分化的内分泌机制㊂本研究中,综述了外源皮质醇处理对鱼类性别分化的作用规律,鱼类性别分化或性转变期环境因素对鱼类内源皮质醇水平的影响规律,以及皮质醇调控鱼类性别分化与性转变的分子机制,以期为研究鱼类性别形成的调控机制提供科学参考㊂1㊀外源皮质醇处理对鱼类性别分化的影响1985年,Van den Hurk等[18]分别用皮质醇㊁皮质醇代谢物可的松(cortisone)处理性别未分化的虹鳟(Oncorhynchus mykiss)均能获得性反转的雄性个体,表明糖皮质激素会影响鱼类的性别分化,这意味着鱼类的性别分化受HPI轴调控㊂上述发现引起了国内外学者的广泛关注㊂随后,研究人员陆续在雌雄异体鱼类如博纳里牙汉鱼(Odon-testhes bonariensis)[19]㊁漠斑牙鲆(Paralichthys lethostigma)[20],以及雌雄同体鱼类如条纹锯鮨[21]㊁斜带石斑鱼(Epinephelus coioides)[22]中发现了皮质醇类似的作用效果㊂1.1㊀皮质醇处理对雌雄异体鱼类性别分化的影响在雌雄异体鱼类中,皮质醇处理会造成斑马鱼[14]㊁尼罗罗非鱼[15]等模式鱼类,以及黄颡鱼(Tachysurus fulvidraco)[23]㊁褐牙鲆(Paralichthys olivaceus)[24]等经济鱼类的雄性化,且皮质醇诱导的雄性化具有剂量依赖性(表1)㊂如对孵化后7d 的博纳里牙汉鱼分别投喂皮质醇含量为400㊁800mg/kg的饲料63d,产生的雄性比例分别为95%和100%[19]㊂对孵化后60d的漠斑牙鲆分别投喂皮质醇含量为100㊁300mg/kg的饲料28d,产生的伪雄鱼比例分别为71%和87%[20]㊂皮质醇处理的起始时期不同,影响的效果也不同㊂如在尼罗罗非鱼中,对孵化后5d且基因型为雌性的幼鱼投喂皮质醇含量为1000mg/kg的饲料25d,会造成幼鱼性腺中卵母细胞的缺失,而在孵化后40d开始用皮质醇处理,处理50d后幼鱼性腺中仍具卵母细胞[15]㊂此外,经皮质醇处理所产生的伪雄鱼可能具有生殖功能㊂如采用皮质醇含量为300mg/kg的饲料投喂孵化后12d的黄颡鱼24d,所产生的伪雄鱼具有精小叶结构和生理性雄鱼特有的生殖突[23]㊂上述经外源皮质醇处理造成鱼类的雄性化,在一定程度上可被雌激素或皮质醇合成抑制剂回救㊂如对孵化后30d且基因型为雌性的褐牙鲆研究发现,对照组(正常饲料)㊁皮质醇组(100mg/kg饲料)㊁联合使用皮质醇(100mg/kg饲料)与E2 (1mg/kg饲料)饲喂组所产生的雄性比例分别为3.3%㊁50%和0%[24]㊂而分别用皮质醇(50mg/kg 饲料)㊁皮质醇合成抑制剂美替拉酮(500mg/kg 饲料)㊁联合使用皮质醇(50mg/kg饲料)与美替拉酮(metyrapone)(500mg/kg)饲料对15日龄的斑马鱼进行为期1个月的饲喂,所产生的雄性比例分别为100%㊁61.9%和48.6%[14]㊂1.2㊀皮质醇处理对雌雄同体鱼类性别分化的影响皮质醇处理会诱导一些雌雄同体鱼类的雄性化(表1)㊂分别用皮质醇(300mg/kg饲料)㊁皮质醇受体拮抗剂米非司酮(mifepristone)(6.25mg/kg 饲料)饲喂性别未分化的雌雄同体雌性先熟的条纹锯鮨幼鱼,处理84d后所产生具有精巢的个体比例分别为31.6%和50.9%,具有兼性性腺的个体比例分别为68.4%和32.0%[21],推测皮质醇通过与皮质醇受体结合的方式对鱼类性别进行调控㊂而在某些雌雄同体鱼类中,对成鱼进行皮质醇处理也会造成雌性转变为雄性㊂如对雌性三斑海猪鱼(Halichoeres trimaculatus)成鱼进行为期42d的皮质醇饲喂(1000mg/kg饲料),可造成血浆E2水平下降,性腺中出现生精细胞[16]㊂通过腹腔注射让雌性斜带石斑鱼成鱼摄入皮质醇(50mg/kg体质727第4期高蕊,等:皮质醇对鱼类性别分化过程的影响及调控机制研究进展表1㊀外源皮质醇处理诱导鱼类雄性化过程中性别比例及相关基因表达的变化Tab.1㊀Changes in sex ratio and sex-related genes expression in the processes of exogenous cortisol treatment induced mascu-linization in fish生殖策略reproductive strategy物种species皮质醇处理cortisol treatment方式method浓度concentration起始时期start date时长duration雄性比例percentageof male性别相关基因sex relatedgene参考文献reference雌雄异体gonochorism虹鳟(Oncorhynchusmykiss)浸泡处理30mg/100L受精后41d30d92%(受精后150d)80%(受精后300d)Van denHurk等[18]漠斑牙鲆(Paralichthyslethostigma)饲喂处理100mg/kg饲料孵化后60d(XX基因型幼鱼)28d(第14天和第21天间存在12d的暂缓期)71%(孵化后138d)Mankiewicz等[20]饲喂处理300mg/kg饲料孵化后60d(XX基因型幼鱼)28d(第14天和第21天间存在12d的暂缓期)87%(孵化后138d)黄颡鱼(Pelteobagrusfulvidraco)饲喂处理300mg/kg饲料孵化后12d24d97%(孵化后62d)84%(孵化后122d)齐飘飘[23]博纳里牙汉鱼(Odontesthesbonariensis)饲喂处理400mg/kg饲料孵化后7d63d95%(孵化后126d)amhʏcyp19a1aˌHattori等[19]饲喂处理800mg/kg饲料孵化后7d63d100%(孵化后126d)amhʏcyp19a1aˌ饲喂处理1000mg/kg饲料孵化后5d(XX基因型幼鱼)25dcyp19a1aˌgsdfʏ尼罗罗非鱼(Oreochromisniloticus)饲喂处理(1000mg皮质醇+800mg17β-雌二醇)/kg饲料孵化后5d(XX基因型幼鱼)175d彭锟[15]饲喂处理1000mg/kg饲料孵化后5d(XX基因型幼鱼)55dcyp19a1aˌgsdfʏ饲喂处理1000mg/kg饲料孵化后40d(XX基因型幼鱼)50d cyp19a1aˌ雌雄同体hermaphroditism条纹锯鮨(Centropristisstriata)饲喂处理300mg/kg饲料性别未分化的幼鱼84d31.6%,但68.4%具有兼性性腺(停止处理时)Miller等[21]三斑海猪鱼(Halichoerestrimaculatus)饲喂处理200mg/kg饲料雌性成鱼42d0,但42.9%具有兼性性腺Nozu等[16]饲喂处理1000mg/kg饲料雌性成鱼42d11.1%,但88.9%具有兼性性腺(停止处理时)皮下注射2mg/kg体质量雌性成鱼60d0dmrt1ʏamhʏsox9ʏcyp19a1aʏ(停止处理时)斜带石斑鱼(Epinepheluscoioides)皮下注射10mg/kg体质量雌性成鱼60d0,但100%具有兼性性腺dmrt1ʏamhʏsox9ʏcyp19a1aʏ(停止处理时)Chen等[22]皮下注射50mg/kg体质量雌性成鱼60d50%,但50%具有兼性性腺(停止处理时)dmrt1ʏamhʏsox9ʏcyp19a1aʏ(停止处理时)量),可使其性别转变为雄性[22]㊂然而在停止处理后,已经发生性转变的斜带石斑鱼精子停止性转变,发育中的精子也消失了,这表明皮质醇引起的性转变具有暂时性㊂以上研究表明,皮质醇对鱼类的性别分化具有重要作用,可在一定程度上造成鱼类向雄性方向分化,并且进行外源皮质醇处理时,不同浓度㊁处理时间和处理方式对鱼类性别分化的影响效果不同㊂827大连海洋大学学报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第38卷皮质醇可以通过皮质醇受体发挥调控作用,皮质醇合成抑制剂或雌激素在一定程度上可回救皮质醇诱导的雄性化㊂值得注意的是,在一些鱼类中,皮质醇诱导所产生的伪雄鱼具有生殖突,推测具有生殖功能㊂而在一些鱼类中,皮质醇处理造成的雄性化却是暂时的,停止处理后性别的改变将不可持续㊂目前,大多数研究未将皮质醇处理产生的伪雄鱼饲养至性成熟,尚不能确定皮质醇处理产生的伪雄鱼是否真正具有生殖功能,未来尚需深入研究㊂2㊀环境因素对鱼类内源皮质醇水平的影响鱼类的性别分化与性转变受到外部环境因素的影响,在多种环境因素介导的鱼类性别分化与性转变过程中,内源皮质醇水平往往发生了相应的变化(表2),故推断皮质醇可能是响应外部环境信号与鱼类性别分化及性转变的重要因子[20,25]㊂2.1㊀温度在许多鱼类㊁爬行动物和两栖类生命早期阶段,温度对性别分化过程具有决定作用㊂1981年, Conover等[26]首次证明,温度会影响大西洋银汉鱼(Menidia menidia)的性别㊂此后,研究人员开展了一系列温度对鱼类性别分化的研究㊂如对孵化后9d的奥利亚罗非鱼(Oreochromis aureus)进行为期25d的温度试验,在27㊁37ħ条件下产生的雄性后代比例分别为63.0%和97.8%,这表明高温会使奥利亚罗非鱼偏雄性化[27]㊂对孵化后15~25d 且基因型为雌性的斑马鱼进行高温(37ħ)处理,可得到100%伪雄鱼[28]㊂此外,高温处理还会诱导金鱼(Carassius auratus)[29]㊁半滑舌鳎(Cynoglos-sus semilaevis)[30]和褐牙鲆[3]产生雄性化㊂以上温度诱导产生的雄性化可能是一种由皮质醇介导的热应激结果㊂孵化后几周的环境温度决定了博纳里牙汉鱼幼鱼的性别[31],在17ħ时雄性比例为0%,在24ħ时为69.2%,在29ħ时可达100%,且在29ħ时饲育的幼鱼体内皮质醇水平始终高于17ħ[19]㊂24ħ条件下对博纳里牙汉鱼幼鱼投喂皮质醇含量为800mg/kg的饲料,可使雄性比例达到100%[32]㊂在褐牙鲆[24]和青鳉[25]中也存在类似现象,高温条件下饲喂的幼鱼皮质醇水平往往较高,而雄性比例也相对较高㊂同时研究还发现,在一些鱼类中,饲喂E2或美替拉酮可回救由高温诱导的幼鱼个体雄性化[24-25,33]㊂2.2㊀种群密度种群密度也会对鱼类性别分化产生一定影响㊂高密度会造成拥挤胁迫,使幼鱼向雄性化转变[34]㊂早期研究发现,欧洲鳗鲡(Anguilla anguilla)在800㊁1600㊁3200g/m3养殖密度下,雄性比例分别为69%㊁78%和96%[35]㊂此外,种群密度还会影响欧洲舌齿鲈(Dicentrarchus labrax)[36]㊁斑马鱼[14]和博纳里牙汉鱼[17]等鱼类性别分化㊂研究发现,在鱼类早期发育阶段,高密度养殖造成的个体雄性化可能也与皮质醇水平有关㊂在日本鳗鲡(Anguilla japonica)性别分化期间,成群饲养个体的血清皮质醇水平显著高于单独饲养个体(P<0.05),推断这是造成鳗鲡高密度养殖个体雄性偏多的原因[37]㊂在斑马鱼的性别分化期也发现,养殖密度越高,体内皮质醇水平越高,雄性比例也就越高[14]㊂在对XX基因型博纳里牙汉鱼的研究中也得出类似结论,用四周均为镜面的水槽饲养博纳里牙汉鱼比无反射光水槽饲养时雄性比率更高[17]㊂推断由环境拥挤引起的鱼类个体雄性化过程中,负责处理视觉信息的大脑具有重要作用㊂综上,在鱼类性别分化期,高密度养殖造成的拥挤胁迫能促使鱼类体内皮质醇水平升高,而高皮质醇水平是造成鱼类雄性化的一个重要原因㊂2.3㊀种群社会关系种群中的社会关系变化会造成雌雄同体鱼类,如黑双锯鱼(Amphiprion melanopus)[38]㊁双带锦鱼(Thalassoma bifasciatum)[39]和蓝带血虾虎鱼(Ly-thyrpnus dalli)[12]等的性转变,期间也往往伴随着皮质醇水平的变化㊂目前,在皮质醇作用于雌雄同体鱼类性别转变的假设中,Perry等[39]认为,在雌雄同体雌性先熟的双带锦鱼中,功能性雄性通过攻击雌性来提高雌性体内的皮质醇水平,从而抑制雌性11-KT合成,进而阻止双带锦鱼雌鱼的雄性化㊂如果将功能性雄性从社会群体中移除,雌性的压力降低,体内皮质醇水平会下降,雌鱼便会出现雄性化㊂然而与Perry等[39]假设所矛盾的是,当去除同样为雌雄同体雌性先熟的圆拟鲈(Parapercis cylindrica)功能性雄性后,完成皮质醇植入的功能性雌性仍可性转变为雄性,这表明高皮质醇水平并不能阻止圆拟鲈的雄性化[40]㊂蓝带血虾虎鱼是一种具有一夫多妻制种群社会关系且雌雄同体雌雄同步成熟的鱼类,研究发现,通常蓝带血虾虎鱼雄性体内皮质醇927第4期高蕊,等:皮质醇对鱼类性别分化过程的影响及调控机制研究进展表2㊀环境因素诱导雄性化过程中鱼类皮质醇水平和性别比例的变化Tab.2㊀Changes in cortisol level and sex ratio in the processes of environmental factors induced masculinization in fish处理treatment物种species生殖策略reproductivestrategy试验experiment处理条件treatment condition起始时期start date时长duration皮质醇水平cortisol level雄性比例percentageof male参考文献reference温度temperature 博纳里牙汉鱼(Odontesthesbonariensis)雌雄异体17ħ24ħ29ħ孵化后0d孵化后0d孵化后0d126d126d126d29ħ组>24ħ组>17ħ组(孵化后21d),24ħ组>29ħ组>17ħ组(孵化后49d)69.2%100%Hattori等[19]褐牙鲆(Paralichthysolivaceus)雌雄异体18ħ27ħ孵化后30d(XX基因型幼鱼)孵化后30d(XX基因型幼鱼)70d70d27ħ组>18ħ组(孵化后60㊁100d)3.3%100%Yamaguchi等[24]密度density鳗鲡(Anguillajaponica)雌雄异体一个玻璃容器中养殖1尾鱼一个玻璃容器中养殖2尾鱼一个玻璃容器中养殖4尾鱼一个玻璃容器中养殖8尾鱼性别分化早期性别分化早期性别分化早期性别分化早期14d14d14d14d8尾/容器组>4尾/容器组=2尾/容器组>1尾/容器组Chiba等[37]斑马鱼(Danio rerio)雌雄异体养殖密度9尾/L养殖密度19尾/L养殖密度37尾/L养殖密度74尾/L受精后6d受精后6d受精后6d受精后6d84d84d84d84d74尾/L组>9尾/L组54.4%61.4%71.6%80.1%Ribas等[14]社会关系social relations蓝带血虾虎鱼(Lythyrpnusdalli)雌雄同体雌雄同步成熟移除种群中的功能性雄性功能性雌性个体皮质醇水平先升高后下降,并在功能性雄性移除后的3d达到峰值Solomon-Lane等[12]水箱颜色color of tanks漠斑牙鲆(Paralichthyslethostigma)雌雄异体22.9ħ下,将幼鱼分别养殖在灰色㊁黑色和蓝色水箱中将幼鱼养殖在灰色㊁黑色和蓝色水箱中,处理的前50d温度控制在19ħ,后维持在23ħ孵化后60d孵化后60d93d93d处理34d时,蓝色组>灰色组>黑色组,在处理65㊁93d时,3个处理组皮质醇水平无显著性差异灰色㊁黑色组为50%左右,蓝色组为95%灰色㊁黑色组为55%左右,蓝色组为74%Mankiewicz等[20]水平较低,反而体型较大的雌性体内皮质醇水平较高,但在移除种群中功能性雄性后,体型较大的雌鱼皮质醇水平开始上升,并在移除雄性后的1~3d 时达到峰值,后逐渐降低[12]㊂这表明,在性别转变初期,高浓度的皮质醇对鱼类性转变为雄性起着促进作用,这与Perry等[39]的假设不同㊂而对于雌雄同体雄性先熟的黑双锯鱼,雄性和雌性皮质醇水平并无差异,但当功能性雌性去除后,种群内各个体的皮质醇水平不断升高,并逐渐有个体性转变为新的功能性雌性[38]㊂由此可见,皮质醇的升高不仅能促进雌雄同体鱼类性转变为雄性,还可能在雌雄同体鱼类性转变为雌性的过程中发挥作用㊂综上所述,皮质醇水平的高低一定程度上反映了一些雌雄同体鱼类在种群中的社会地位㊂皮质醇参与了雌雄同体鱼类性转变的过程,而高水平皮质醇水平可能促进了一些雌雄同体鱼类的性转变㊂但目前相关研究还较少,尚需进一步探究皮质醇在雌雄同体鱼类性转变中的作用机制㊂2.4㊀其他环境因素光照㊁养殖水槽颜色也会对鱼类的性别分化造成影响㊂研究发现,这些环境因素可通过影响鱼体皮质醇水平对鱼类性别分化进行调控,如将刚孵化的基因型为雌性的青鳉在绿光环境下饲养60d,可诱导产生伪雄鱼且伪雄鱼具有生殖功能,所产生的精子可与正常卵子结合孵育全雌子代,推测绿光造037大连海洋大学学报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第38卷成青鳉雄性化是由皮质醇水平变化引起的[4]㊂在探究养殖水箱对漠斑牙鲆性别分化影响时发现,蓝色水箱中饲养的鱼皮质醇水平较高,相应的雄性比例也较高,推测背景颜色影响了漠斑牙鲆的性别,表明环境因素在性别决定期间充当压力源,并最终造成雄性偏向性[20]㊂总之,外部环境因素影响着鱼类的性别分化与性转变,而皮质醇可能是连接外部环境因素与鱼类性别调控的重要纽带,在鱼类的性别分化与性转变中起到了重要作用㊂3㊀皮质醇调控性别分化与性转变的分子机制研究认为,皮质醇可能通过3种方式介导硬骨鱼类性别分化与性转变[41-42]:一是通过HPI轴与下丘脑-垂体-性腺(hypothalamus-pituitary-gonadal, HPG)轴相互作用,调控鱼类的性别变化;二是通过皮质醇和雄激素合成通路的交互作用,共同调控鱼类的性别变化;三是通过控制鱼类性别相关基因的转录来调控鱼类性别变化㊂3.1㊀HPG轴与HPI轴的相互作用HPG和HPI轴的相互作用共同调控鱼类的性别分化㊁发育和繁殖等重要生理过程[41](图1)㊂脑内一些神经递质通过影响体内皮质醇水平,间接调控着性别分化与性转变过程[43]㊂参与HPG和HPI轴相互作用的神经递质主要包括去甲肾上腺素(norepinephrine,NE)㊁精氨酸催产素(arginine vasotocin,AVT)㊁多巴胺(dopa-mine,DA)㊁血清素(serotonin,5-HT)㊁亲吻素(kisspeptin)和褪黑素(melatonin,MEL)等㊂其中,NE能影响促性腺激素释放激素(gonadotropin-releasing hormone,GnRH)的释放和促性腺激素(gonadotropins,GtHs)的产生[44],且对促肾上腺皮质激素释放因子(corticotropin-releasing factor, CRF)具有调控作用[45]㊂AVT与哺乳动物精氨酸加压素(arginine vasopressin,AVP)同源,是研究鱼类行为及性转变所关注的主要激素[46]㊂对于群居性雌雄同体的鱼类,一个社会群体中功能性雄性死亡后,可能会使体型较大的雌鱼下丘脑中的AVT和NE水平上升,造成雌鱼性转变为雄性[47]㊂这些快速的神经化学变化反过来也影响了GnRH和促黄体生成素(luteinizing hormones,LH)的释放,促进鱼类卵巢细胞凋亡并提高皮质醇水平[43]㊂一般认为,脑中NE活动增加会导致血清皮质醇水ACTH 促肾上腺皮质激素;AVT 精氨酸催产素;CRF 促肾上腺皮质激素的释放因子;DA 多巴胺;E2 17β-雌二醇; FSH 促卵泡激素;FSHR 促卵泡激素受体;GnIH 促性腺激素抑制激素;GnRH 促性腺激素释放激素;GR 皮质醇激素受体;Kisspeptin 亲吻素;LH 促黄体生成素;LHR 促黄体生成素受体;MEL 褪黑素;NE 去甲肾上腺素;T 睾酮;5-HT 血清素;11-KT 11-酮基睾酮㊂ACTH adrenocorticotropic hormone;AVT arginine vasotocin; CRF corticotropin-releasing factor;DA dopamine;E2 17β-es-tradiol;FSH follicle-stimulating-hormone;FSHR follicle-stimu-lating hormone receptor;GnIH gonadotropin-inhibitory hormone; GnRH gonadotropin-releasing hormone;GR glucocorticoid re-ceptor;LH luteinizing hormones;LHR luteinizing hormones re-ceptor;MEL melatonin;NE norepinephrine;T testosterone; 5-HT serotonin;11-KT 11-ketotestosterone.图1㊀下丘脑-垂体-性腺轴与下丘脑-垂体-肾间组织轴的关系[41]Fig.1㊀Diagram of the relationship between hypothala-mus-pituitary-gonadal(HPG)axis and hypo-thalamus-pituitary-interrenal(HPI)axis[41]平快速上升,而5-HT的减少会消除NE对AVT信号的抑制,使鱼类在性腺改变的第一阶段维持高皮质醇水平[43]㊂Kisspeptin是一种下丘脑神经肽,研究发现,在雌性大鼠中,位于下丘脑弓状核区的Kisspetin细胞和糖皮质激素受体(glucocorticoid re-ceptor,GR)共表达[48],推测GR是HPI和HPG 轴间联系的纽带,环境信号因子能通过HPI轴作用于GR来影响HPG轴[49]㊂MEL对鱼类的昼夜节律㊁血压和季节性繁殖具有调控作用㊂研究发现, MEL和NE能瞬时调节GnRH产生,以促进LH的生成,启动性转变[50]㊂随后皮质醇和促性腺激素137第4期高蕊,等:皮质醇对鱼类性别分化过程的影响及调控机制研究进展抑制激素(gonadotropin-inhibitory hormone,GnIH)增加,进而抑制GnRH和GtHs信号传导㊂在这些因素的共同作用下,鱼体内皮质醇水平增加㊂皮质醇通过抑制E2合成所必需的cyp19a1a(cytochrome P450,family19,subfamily A,polypeptide1a)基因转录,来调控E2的合成和雌性相关基因的表达,导致卵巢退变为精巢[42-43]㊂3.2㊀皮质醇和雄激素合成通路间的交互作用与哺乳动物不同的是,硬骨鱼类最主要的雄激素不是睾酮(testosterone),而是11-KT㊂11-KT的合成与皮质醇的合成㊁代谢过程间存在交互关系,两过程均有11β-羟化酶(11β-hydroxylase, Cyp11b)和11β-羟基类固醇脱氢酶(11β-hydroxys-teroid dehydrogenase,Hsd11b)参与催化(图2)㊂睾酮在Cyp11b催化下转化为11β-羟基睾酮(11β-OH-testosterone),11β-羟基睾酮在Hsd11b催化下转化为11-KT[42,51-52]㊂同样,11-脱氧皮质醇(11-deoxycortisol)在Cyp11b催化下转化为皮质醇,后皮质醇在Hsd11b催化下代谢为可的松[32,42]㊂皮质醇也可通过调节编码Hsd11b的hsd11b2 (hydroxysteroid11-beta dehydrogenase2)基因表达,调控鱼类11-KT的生成,进而影响鱼类性别分化与性转变㊂有研究发现,在皮质醇诱导博纳里牙汉鱼雄性化的过程中,hsd11b2表达上调㊂有趣的是,睾酮和11-KT的升高要先于cyp19a1a基因表达量的下降[32]㊂对斜带石斑鱼的研究中也发现了类似的现象,腹腔注射皮质醇(50mg/kg体质量)能够使hsd11b2表达迅速上调,11-KT水平随之升高,编码Cyp11b的cyp11b2(cytochrome P450family11 subfamily B member2)基因表达上调和cyp19a1a基因表达下调均发生在11-KT升高之后[22]㊂由此推测,在一些鱼类性别分化与性别转变过程中,皮质醇可通过直接调控雄激素合成相关基因表达的方式来提高11-KT水平,进而激活雄性化通路㊂在雄性通路被激活后,雌激素合成相关的通路则被抑制㊂㊀DHEA dehydroepiandrosteron;OH hydoxy.图2㊀硬骨鱼类类固醇激素合成[42,52]Fig.2㊀Schematic representation of steroidogenesis in teleost fishes[42,52]3.3㊀皮质醇调控鱼类性别相关基因的转录皮质醇可通过先与糖皮质激素受体结合,再与应答基因启动子区域内的糖皮质激素反应元件(glucocorticoid response elements,GRE)结合,直接控制性别相关靶基因转录进而调控鱼类性别转变㊂一些硬骨鱼类的cyp19a1a㊁fshr(follicle stimu-lating hormone receptor)和dmrt1(double-sex and mab-3-related transcription factor1)基因启动子区域存在GRE[25,53-54],皮质醇-糖皮质激素受体复合物能通过与这些基因上的GRE作用,对鱼类性别开展调控㊂除此之外,皮质醇还能通过其他未知方式调控性别相关基因的转录㊂尽管在鱼类的抗苗勒氏管激素amh(anti-Müllerian hormone)基因上未发现GRE,但皮质醇却可以通过调控amh表达影响鱼类性别㊂3.3.1㊀皮质醇抑制cyp19a1a的转录㊀在鱼类中,睾酮是E2合成的原料[55]㊂睾酮可以在性腺芳香化237大连海洋大学学报㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第38卷。
2019年南京师范大学海洋科学与工程学院考研招生目录及参考书目

不接受同等学 10 力考生报考
复试参考书目
651 动物生物学: 《普通动物学》(第 4 版),刘凌云等主编,高等教育出版社,2009 年 880 水生生物学: 《高级水生生物学》,刘建康,科学出版社,2000 年。 F146 动物学: 《普通动物学》(第 4 版),刘凌云等主编,高等教育出版社,2009 年。
考研咨询、更多南京师范大学考研信息请关注新浪微博:南京师范大学考研百事通
专业及方向 095134
初试科目
渔业发展 ①101 思想政治理论
同等学力 人数 加试科目 ⑤F146 动物学 ⑥池塘养殖学 15 复试科目
考研咨询、更多南京师范大学考研信息请关注新浪微博:南京师范大学考研百事通
(专业学位) ②204 英语二 ⑦鱼类学 01 (全日制)水生动 ③340 农业知识综合二 物繁殖与遗传育种 ④880 水生生物学 02 (全日制)水生动 物疾病防治 03 (全日制)渔业资 源可持续利用 340 农业知识综合二: 《水产经济动物增养殖学》,李明云主编,海洋出版社,2011。 初试参考书目 880 水生生物学: 《高级水生生物学》,刘建康,科学出版社,2000 年。 F146 动物学: 复试参考书目 《普通动物学》(第 4 版),刘凌云等主编,高等教育出版社,2009 年。
⑤F145 大学数 不接受同等学 17 学 力考生报考
650 海洋专业综合: 《海洋资源》,辛仁臣、刘豪,中国石化出版社,2008 年。 初试参考书目 879 海洋科学概论: 《海洋科学导论》,冯士筰等编,高等教育出版社。 F145 大学数学: 《高等数学》(第七版,上下册),同济大学编,高等教育出版社,2014 年。 初试科目 复试科目 同等学力 加试科目 人数
2019 南京师范大学海洋科学与工程学院考研招生目录及参考书目
波罗的海治理实践对跨界海洋空间规划的启示

学报社会科学版2022年第3期波罗的海治理实践对跨界海洋空间规划的启示∗郭雨晨1,2㊀练梓菁3(1.广东省海洋发展规划研究中心,广东广州510275;2.南方海洋科学与工程广东省实验室(珠海),广东珠海519082;3.中山大学海洋科学学院,广东珠海519082)摘㊀要:随着海洋空间规划在世界范围内的兴起与发展,跨界海洋空间规划逐渐成为了国与国之间探寻海上合作的新议题.欧洲为目前跨界海洋空间规划项目开展的集中区域,波罗的海区域又在整个欧洲跨界海洋空间规划发展中扮演着 排头兵 的角色.以波罗的海跨界海洋空间规划的实现路径为研究内容,旨在为我国今后与其他东南亚沿海国的跨界海洋空间规划合作提供参考.通过分析得出,欧盟法律政策㊁区域政府间合作机制㊁沿海国海洋空间规划制度的普遍建立㊁区域数据共享平台建设与欧盟资助,以及该区域网络化治理特征等因素是促成波罗的海区域跨界海洋空间规划发展与活跃的关键原因所在.关键词:波罗的海;跨界海洋空间规划;欧盟;网络化治理中图分类号:P 74㊀㊀文献标识码:A㊀㊀文章编号:1672G335X (2022)03G0058G10D O I :10.16497/j .c n k i .1672G335X.202203006一㊁引言海洋空间规划是以生态系统方法为基础的,分析㊁分配人类海洋活动时空分布,从而实现可持续发展目标的过程.[1]跨界海洋空间规划则为区域㊁国家或地区层面在海洋空间规划事务方面进行的跨界合作.海洋自然环境的流动性,海洋资源与海洋活动分布㊁影响的跨区域性,以及基于陆海统筹的海洋管理思路是进行跨界海洋空间规划的必要性所在.[2]广义的跨界海洋空间规划包括跨行政边界海洋空间规划㊁跨地理边界海洋空间规划和跨治理边界海洋空间规划三层内涵.[3]狭义的跨界海洋空间规划,也是目前学术和实务界讨论最多的,一般指跨行政边界的海洋空间规划.欧洲是目前在海洋空间规划方面开展最多跨界合作的区域.根据欧盟政策的定义, 跨界合作 可大致分为两种.一种是指具有一条共享边界的邻国之间的合作.边界可能是已确定的,也可能是还未划分㊁仍有争议的.[4]另一种是指多个具有管辖权的实体(例如国家㊁州㊁省㊁地方机构)参与的合作.这些实体可能并不共享同一边界,而在更广阔的地域范围内合作,例如波罗的海区域㊁地中海区域等.[5]上述两种跨界合作都是基于大生态系统管理方法,旨在最大程度降低行政边界划分对海洋生态系统的割裂性管理所造成的影响.在欧洲各区域较为活跃的跨界海洋空间规划合作中,波罗的海区域扮演了 排头兵 的角色.因此,本文选取波罗的海区域为研究对象,拟从多维度对海洋空间规划跨界合作所需的现实条件与实现途径进行分析.跨界海洋空间规划的经验具备一定的普遍性,在目前亚洲众多沿海国开展海洋空间规划立法与制度建设的当下,波罗的海区域的实践可供我国和世界其他区域沿海国就海85∗收稿日期:2022G02G20基金项目:国家社会科学基金重大研究专项 新时代海洋强国建设 (20V H Q 005)的阶段性成果作者简介:郭雨晨(1989G㊀),女,江苏徐州人,广东省海洋发展规划研究中心助理研究员,南方海洋科学与工程广东省实验室(珠海)成员,法学博士,主要从事海洋空间规划㊁海洋管理与海洋法研究.中国海洋大学学报(社会科学版)2022年5月洋空间规划的跨界合作进行参考.二、波罗的海跨界海洋空间规划发展现状波罗的海位于斯堪的纳维亚半岛与欧洲大陆之间,为典型的半闭海,通过斯卡格拉克海峡㊁卡特加特海峡㊁大贝尔特海峡㊁小贝尔特海峡,以及厄勒海峡等狭窄的海峡与大西洋相连,深水交换较少.波罗的海水面面积约42万平方公里,三分之一以上的水域深度不足30米.因此,与其表面积相比,波罗的海总体水量很小.[6]波罗的海流域约居住8500万人,沿岸国包括丹麦㊁德国㊁波兰㊁立陶宛㊁拉脱维亚㊁爱沙尼亚㊁俄罗斯㊁芬兰和瑞典等国家.除俄罗斯外,其余八国均为欧盟成员国.因此,波罗的海区域也是欧盟与俄罗斯之间重要的区域联系纽带.波罗的海的浅海㊁半封闭特征,加之约有200条河流注入波罗的海,带入河流沿岸的城市㊁工农业污染物,使得波罗的海的海洋环境问题一直比较突出和棘手;并且,波罗的海海域为逆时针环流,且少有深水交换;这种水循环模式使得一些沿岸国家的污染物(例如俄罗斯和波兰)被转移到其他国家的管辖水域.[4]因此,波罗的海的自然地理特征在很大程度上决定了沿岸国在海洋开发与保护方面合作的紧迫性与必要性.即便是在冷战时期,波罗的海区域也成功通过了世界上首个综合全面的海洋环保协议 1974年的«保护波罗的海区域海洋环境公约»(又称«赫尔辛基公约»).在海洋空间规划的跨界合作方面,波罗的海区域目前已完成了B a l t S e a P l a n㊁P l a nB o t h n i a㊁B a l t i c S C O P E㊁P a nB a l t i cS c o p e等数个项目(见表1),并开展了多项与跨界海洋空间规划相关的辅助性项目.例如,B a l t i cL I N e s(2016 2019年)项目关注与 航运路线 和 能源走廊 相关的海洋空间规划数据基础设施的建设,以提高波罗的海区域海洋空间规划中航运路线和能源走廊的跨国一致性;B a l t i c R I M (2017 2020年)项目主要针对波罗的海水下文化遗产保护,通过文化遗产管理人员和空间规划人员的合作识别㊁指定海洋文化遗产区,将文化遗产管理整合到海洋空间规划中;C a p a c i t y4M S P(2019 2021年)项目则基于波罗的海区域最新完成的海洋空间规划项目及成果,旨在促进利益相关者㊁政策制定者和决策者的对话交流,拓展海洋空间规划相关知识等.表1㊀波罗的海区域主要跨界海洋空间规划项目目标与参与机构项目名称/时间项目目标参与机构B a l t S e a P l a n (2009 2012年)(1)改善海洋空间规划的信息库;(2)促进各国将海洋空间规划纳入国家海事战略;(3)制定波罗的海区域共同的空间规划战略 «波罗的海愿景2030»;(4)在八个试点区域实施海洋空间规划;(5)促进海洋空间规划的能力培养或研究德国联邦海洋与水文局,梅克伦堡G前波美拉尼亚州交通㊁建设和区域发展部,世界自然基金会(德国);波兰格丁尼亚海事处,什切青海事处,格但斯克海事研究所;丹麦奥尔胡斯大学国立环境研究所;爱沙尼亚塔尔图大学海洋研究所,波罗的海环境论坛(爱沙尼亚);立陶宛克莱佩达大学海岸研究与规划所,波罗的海环境论坛(立陶宛):波罗的海环境论坛(拉脱维亚);瑞典皇家理工学院㊁瑞典环境保护局P l a nB o t h n i a (2010 2012年)(1)实施芬兰海洋水下环境清单计划,研究海洋栖息地情况;(2)制定波的尼亚海海洋空间规划草案芬兰海事研究中心,芬兰环境研究所;瑞典国家住房㊁建筑及规划委员会,原瑞典渔业委员会;波罗的海远景战略委员会(V A S A B);赫尔辛基公约委员会(H E L C OM);N o r d r e g i oB a l t i c SC O P E (2015 2017年)(1)加强各国海洋空间规划主管部门与利益相关者合作;(2)针对跨界问题制定解决方案,提高各国实施国家海洋空间规划的连贯一致性;(3)就试验区域航运㊁能源㊁渔业和自然保护区四个议题开展跨界问题讨论瑞典海洋与水资源管理局;德国联邦海洋与水文局;波兰什切青海事处;丹麦海事局;爱沙尼亚财政部;拉脱维亚环境保护与区域发展部;芬兰环境研究所;V A S A B;H E L C OM;N o r d r e g i oP a nB a l t i c S c o p e (2018 2019年)(1)促进各国国家海洋空间规划流程的跨界合作与咨询;(2)加快生态系统方法和数据共享的落实;(3)研究海洋空间规划中的陆海相互作用,探索陆海统筹中的海陆一体化规划方式瑞典海洋与水资源管理局;德国联邦海洋与水文局;波兰什切青海事处;丹麦海事局;拉脱维亚环境保护与区域发展部;爱沙尼亚财政部;芬兰环境研究所,芬兰奥兰自治省政府;V A S A B;H E L C OM;N o r d r e g i o㊀㊀注:资料来源于作者收集整理.952022年第3期郭雨晨㊀练梓菁:波罗的海治理实践对跨界海洋空间规划的启示三、波罗的海跨界海洋空间规划法律政策保障与政府间合作机制波罗的海跨界海洋空间规划的法律与政策依据主要来自全球层面㊁欧盟层面和波罗的海区域层面.在全球性公约层面,«联合国海洋法公约»和«生物多样性保护公约»为各沿海国国内海洋空间规划及跨界合作提供了基本的法律框架.[7]1991年的«跨界背景下的环境影响评价公约»和该公约下的«战略环境影响评估协定»,以及1998年的«在环境问题上获得信息㊁公众参与决策和诉诸法律的公约»分别对跨界环境影响评估㊁战略影响评估,以及公众与利益相关者参与事务提供部分法律支持.另外,国际海事组织的公约和相关制度也与跨界海洋空间规划相关.欧盟与波罗的海区域层面的法律政策与政府间合作机制对波罗的海跨界海洋空间规划的兴起和发展起到了直接推动作用.(一)欧盟法律政策2007年,欧盟委员会发布«综合海洋政策»(I M P),意图打破过去欧盟海洋事务部门化㊁碎片化的管理方式,在政策层面对欧盟海洋管理的各层级㊁部门予以协调和统筹.作为统筹整合海洋事务㊁部门和政策途径之一的海洋空间规划,也被列入了«综合海洋政策»待制定和实施的工作计划.2008年,欧盟发布了海洋空间规划指导性文件 «海洋空间规划路线图»(R o a d m a p f o rM a r i t i m e S p a t i a l P l a n n i n g)以落实«综合海洋政策»中关于在欧盟建立海洋空间规划制度的要求.该文件制定出台的目的不仅在于推进成员国各自海洋空间规划的实施,还鼓励㊁倡导成员国的海洋空间规划合作,以实现整个欧洲海域空间规划的协调和管理.[8]除了«海洋空间规划路线图»之外,2008年发布的与海洋空间规划密切相关的文件,还有欧盟的«海洋战略框架指令»(M S F D),该文件为成员国建立以生态系统方法为基础的海洋空间规划制度提供了法律框架.[9]2009年,欧盟波罗的海战略(E U S B S R)出台,该战略旨在加强波罗的海区域的合作,以应对其面临的共同挑战,并在对生态环境负责的前提下促进经济发展.战略提出了包括保护波罗的海海洋环境,通过能源和运输系统加强波罗的海各国的紧密联系,以及消除波罗的海区域各国市场壁垒,发展共同的经济㊁文化和旅游基础等主要目标,并指出跨领域行动的实施对于整个战略的执行至关重要,其中就包括发展综合海洋治理体系以及海域与陆域空间规划.[10]2014年,欧盟出台了更为详细的«海洋空间规划指令»(M S P D),要求成员国在2021年3月前制定本国的海洋空间规划.[11]«海洋空间规划指令»明确了海洋空间规划的总体目标和编制要求,并罗列了成员国制定海洋空间规划时需要重点关注的几个问题.除了海洋空间规划文件中常提及的考虑陆海相互作用㊁运用生态系统方法㊁重视利益相关者参与外,还提出了成员国之间的海洋空间规划合作㊁成员国与非成员国之间的海洋空间规划合作的要求.除了以上与海洋空间规划直接相关的欧盟指令和政策,欧盟的«鸟类指令»和«栖息地指令»中关于海洋自然保护区网建设的规定,以及«共同渔业政策»«水框架指令»等相关规定都是有力支持和推动欧盟各国在海洋空间规划建设以及合作的法律政策依据.(二)波罗的海区域政府间合作机制与数据共享平台欧盟法律政策为欧洲跨界海洋空间规划提供了宏观目标与框架,波罗的海地区空间规划和发展方面的政府间多边合作机制 波罗的海远景战略委员会(以下简称V A S A B)和«保护波罗的海区域海洋环境公约»的执行机构 赫尔辛基公约委员会(以下简称H E L C OM)则为波罗的海沿岸国在跨界海洋空间规划中提供了更具体且契合区域特征的合作平台,以及海洋管理的相关标准与技术性支持.促进波罗的海区域海洋空间规划是V A S A B目前的工作重点之一.V A S A B早在2001年第五届部长级会议就已达成将空间规划向海洋推进的共识.第五届部长级会议发布的«维斯马宣言»(W i s m a r D e c l a r a t i o n)是波罗的海区域首个表明实施海洋空间规划的政治意愿文件.[12]维斯马部长会议还通过了 V A S A B2010+空间发展行动计划 ,该计划将海岸带区域和岛屿开发确定为空间规划的跨国合作主题之一,并将空间规划的范围拓展至领海.之后,V A S A B陆续发布了与海洋空间规划相关的行动计06中国海洋大学学报(社会科学版)2022年5月划㊁宣言及战略文件.2010年10月,V A S A B与H E L C OM建立了海洋空间规划联合工作组(H E L C OMGV A S A B M S P WG,以下简称工作组).该工作组由政府机构和欧盟高级官员组成,为波罗的海跨区域海洋空间规划提供长期㊁稳定的合作机制,确保沿海国在波罗的海建立协调一致的区域海洋空间规划进程.该工作组是波罗的海地区国家在«海洋空间规划指令»协商谈判过程中进行区域性意见交换的平台,并被指定为欧盟波罗的海战略中海洋空间规划事务的 负责人 .V A S A B与H E L C OM还建立了数据专家组(B S R M S PD a t aE S G),作为工作组的子组织.2010年,H E L C OM和V A S A B通过了«波罗的海海洋空间规划原则»,将海洋空间规划的基本原则与欧盟指令㊁«保护波罗的海区域海洋环境公约»,以及公约体系下的现有数据平台(例如地理信息数据库 H E L C OM G I Sd a t a s e t s)相联系,目的在于为波罗的海区域 量身打造 海洋空间规划的发展战略.2013年,H E L C OM和V A S A B部长级会议通过了«波罗的海海洋空间规划路线图(2013 2020年)».这是推动波罗的海沿海国国内和跨界海洋空间规划发展的纲领性文件,以保证2020年在波罗的海区域完成制定和实施海洋空间规划的目标.这部文件设定了波罗的海海洋空间规划需达到的目标,以及实现目标的关键步骤;并且还拟定了相关指南的制定计划以配合这些步骤的实现.这些步骤不仅包括适用生态系统方法㊁加强数据信息交换共享㊁保障公众参与这些海洋空间规划制定与实施的基本要求,还包括促进政府间海洋空间规划合作㊁加强对海洋空间规划专业人员的培养㊁总结次区域海洋空间规划实践经验㊁建立国内与区域海洋空间规划框架,以及对海洋空间规划进行评估的要求.根据«波罗的海海洋空间规划路线图(2013 2020年)»的要求,H E L C OM和V A S A B于2016年通过了«波罗的海区域实施生态系统方法的海洋空间规划指南»和«跨界咨询㊁公众参与与合作指南»两部文件,为生态系统方法在海洋空间规划的适用以及其成员国开展跨界海洋空间规划的相关问题提供了具体说明.最新的指南是2019年初通过的«跨界海洋空间规划输出数据结构指南».该指南规定了跨界海洋空间规划的输出数据规范,以促进跨界海洋空间规划数据的兼容性和互用性.除了各指南,支持波罗的海国内以及跨界海洋空间规划的两大数据平台 H E L C OM MA D S和B A S E MA P S也已建立,旨在为波罗的海各国的国内和跨界海洋空间规划提供可视化地理空间数据. MA D S提供与H E L C OM工作相关的地理空间数据.数据类型包括状况评估㊁监测㊁人类活动压力㊁生物多样性㊁航运㊁陆地和海洋基本情况等.[13]B A S E MA P S则提供了一个交互式地图用户界面,用户可以直接访问波罗的海空间规划原始数据,并了解波罗的海各国海洋空间规划的情况.[14]除了与H E L C OM合作,V A S A B还指导㊁参与多项跨界海洋空间规划项目,并为项目成员和利益相关者搭建交流㊁学习的对话平台,也提供海洋空间规划事务的专业培训.例如,2009年10月,V A SGA B专家组和利益相关者在维尔纽斯举行了讨论波罗的海海洋空间规划挑战的研讨会,以强化V A S A B 在海洋空间规划事务中所起到的先锋作用,以及所发挥的 知识信息库 的功能;2013年,V A S A B与波罗的海地区多所大学合作开展的海洋空间规划教育课程,为参与波罗的海海洋空间规划的专业人士㊁非政府机构㊁咨询公司等提供培训课程.四、波罗的海跨界海洋空间规划的现实基础根据欧盟2014年«海洋空间规划指令»的规定,成员国须在2021年3月前制定本国的海洋空间规划.因此,波罗的海沿岸国海洋空间规划的国内立法和制度建立,在该指令的出台后得以加速推进.海洋空间规划制度在绝大多数波罗的海沿岸国(除俄罗斯外)的建立与实践是跨界海洋空间规划在波罗的海开展的现实基础.在波罗的海沿海九国中,丹麦于2016年通过了«海洋空间规划法»(A c t o n m a r i t i m e s p a t i a l p l a nG162022年第3期郭雨晨㊀练梓菁:波罗的海治理实践对跨界海洋空间规划的启示n i n g),建立了丹麦海洋空间规划制度的框架.丹麦的国家级海洋空间规划项目于2017年1月启动,并于2021年9月完成了公众咨询与战略环境影响评价,进入最后的修改审核阶段.规划将覆盖丹麦在北海与波罗的海的内水㊁领海与专属经济区.[15]爱沙尼亚海洋空间规划的主要国内法依据是2015年通过的新«爱沙尼亚规划法»(E s t o n i a nP l a nGn i n g A c t).规划分为国家级和区域级,两个区域级海洋空间规划 H i i u I s l a n d和Pär n uB a y已分别于2016年和2017年通过.国家级海洋空间规划已进行公众咨询并将报请政府批准.[16]拉脱维亚海洋空间规划的主要法律依据是2011年颁布的«空间发展规划法»(S p a t i a lD e v e l o p m e n t P l a n n i n g L a w).2019年5月,拉脱维亚政府通过了«海洋规划2030»(M a r i t i m eP l a n2030).规划覆盖拉脱维亚内水㊁领海与专属经济区.[17]立陶宛议会在2015年通过了«立陶宛共和国领土全面规划»(C o m p r e h e n s i v eP l a no f t h eT e r r i t o r y o f t h eR e p u b l i c o f L i t h u a n i a),其中包含海洋空间规划的部分.该规划文件于2020年到期,新规划文件编制正在筹备之中.[18]波兰从2007年就开始了试验性规划的尝试,旨在为新的立法活动提供海洋空间规划的方法和经验.«波兰共和国海域与海事管理法»(A c t o n s e a a r e a s o f t h eR e p u b l i c o f P o l a n d a n d t h em a r i t i m e a dGm i n i s t r a t i o n)是波兰海洋空间规划的主要法律依据.2021年5月,«波兰内水㊁领海与专属经济区发展规划(1:200000)»正式实施.[19]芬兰的海洋空间规划是由地区议会制定与批准的,是不具有法律约束力的规划.规划范围覆盖领海与专属经济区.芬兰将其管辖海域分为了四个区域,由八个地区议会和奥兰岛自治政府负责进行规划,其中三个区域的海洋空间规划已于2020年底完成.[20]德国是欧洲最早开展海洋空间规划的国家之一,目前已经完成了包括波罗的海专属经济区海洋空间规划㊁北海专属经济区海洋空间规划㊁石荷州波罗的海和北海领海海洋空间规划㊁梅前州波罗的海领海海洋空间规划㊁下萨克森州北海领海空间规划等五个区域级规划.[21]第二轮北海和波罗的海专属经济区规划已于2021年底正式发布实施.[22]«瑞典环境法规»(S w e d i s hE n v i r o n m e n t a l C o d e)与«规划与建筑法»(P l a na n dB u i l d i n g A c t)是瑞典海洋空间规划的主要法律依据.瑞典位于斯卡格拉克海峡与卡特加特海峡㊁波的尼亚湾,以及其他波罗的海管辖范围内的三部海洋空间规划编制正在进行中,预计2021年底完成,规划将范围覆盖领海绝大部分区域与专属经济区.[23]俄罗斯并非欧盟成员,鉴于海洋空间规划的重要性日渐显著,虽未启动正式的海洋空间规划项目,但俄罗斯已在涅瓦湾㊁芬兰湾等区域进行了多项试验性规划的尝试.[24]并就海洋空间规划的立法㊁程序等相关事宜与其他波罗的海国家积极开展交流.例如,2014年,俄罗斯和波兰在维斯图拉泻湖的合作项目 V I L A 旨在探寻两国在该跨界区域社会经济发展和环境保护方面的合作,项目也涉及到空间规划的内容.2014年,俄罗斯与德国合作启动的为期三年的 俄罗斯联邦对波罗的海沿海地区空间利用中的 环境友好理念 咨询援助项目,目的在于为涅瓦河河口和芬兰湾的空间管理中树立 环境友好 的概念,以在满足社会经济发展需求和 环保优先 的要求中实现平衡.虽然大部分波罗的海沿岸国已建立起的国内海洋空间规划制度,为其跨界合作奠定了一定的基础,但沿岸国层次不齐的海洋空间规划发展状况在其跨界合作中也带来了一定的阻碍:国内海洋空间规划发展进程更先进的国家更愿意处理具体的规划问题和实际矛盾,并且利益相关者对于规划事务也更熟悉并且具有更高参与度.[25]但也正是由于跨界海洋空间规划的开展,为海洋空间规划发展较好的国家和正在起步的国家提供了交流平台,也为后者提供了难得的交流学习机会.在项目资金来源方面,目前已在波罗的海开展的跨界海洋空间规划项目和相关的辅助项目主要由欧洲区域发展基金(E R D F)㊁欧洲海洋与渔业基金(E M F F),以及欧盟委员会海洋与渔业事务总司(E U 26中国海洋大学学报(社会科学版)2022年5月D GGM a r e)进行资助(见表2).有些项目的参与国也会进行部分资金支持.这些资助是欧洲跨界海洋空间规划能够从理论层面向实际执行层面转换的最有力支撑.表2㊀波罗的海跨界海洋空间规划项目资助情况项目名称/期间项目金额主要资助部门㊁基金B a l t S e a P l a n(2009 2012年)370万欧元欧盟区域发展基金(I n t e r r e g项目)B a l t i cSC O P E(2015 2017年)260万欧元欧洲海洋与渔业基金P a nB a l t i c S C O P E(2018 2019年)330万欧元欧洲海洋与渔业基金P l a nB o t h n i a(2010 2012年)50万欧元欧盟委员会海洋与渔业事务总司B a l t i cL I N e s(2016 2019年)240万欧元欧盟区域发展基金(I n t e r r e g项目)B a l t i c R I M(2017 2020年)299万欧元欧盟区域发展基金(I n t e r r e g项目)C a p a c i t y4M S P109万欧元欧盟区域发展基金(I n t e r r e g项目)(俄罗斯也为该项目提供了部分资金)㊀㊀㊀㊀㊀注:资料来源于作者收集整理.五、波罗的海区域的网络化治理特征除上述讨论的法律政策基础㊁政治合作机制等因素外,波罗的海区域的网络化治理特征也是波罗的海区域跨界海洋空间规划发展的重要助力.网络化治理简而言之就是政府部门与非政府部门(包括私营部门㊁第三部门)共同参与治理的多级治理模式.[26](P290)网络化治理被认为是对信息化时代日趋分权和多样化社会的回应.在传统的政府治理模式框架下,政策的制定㊁执行和实施通过自上而下的命令来实现.随着权力的分散㊁社会的日益多元及组织界限的模糊,单纯依靠政府治理社会的难度大大增加,面临的社会问题也日益复杂.在此背景下,网络化治理这种治理模式有利于减少公共政策实施的阻力,强化其助力,促进更有效的资源整合和利用.[27]二十世纪九十年代,网络化治理在波罗的海区域开始兴起,原因主要包括以下三方面:[28]其一,冷战结束后,诸多新的国际或跨政府组织(如波罗的海国家理事会(C B S S))成立,已存在的政府间组织(如H E L C OM)进一步发展;其二,1992年,里约热内卢会议签署了«21世纪议程»,引入并提倡综合性和参与性治理的理念,给予了非政府实体参与社会治理的机会与地位;其三,欧盟1995年和2004年的两次东扩,拓宽了波罗的海国家的合作领域.波罗的海地区网络化治理大致可分为三种类型:国际制度或政府间合作㊁国际政策网络㊁跨国网络.[29]第一种类型即传统的政府间合作形式,在这种模式下,各国的政府部门为决策制定者和执行者,其他参与者(如非政府组织或地方政府)没有决策权.尽管近些年非政府组织在这种合作形式下获得了观察员身份,但仅可发表观点,仍不能参与决策.具有代表性的合作平台和组织有H E L C OM㊁V A SGA B㊁波罗的海议会会议(B S P C)等.第二种类型为国际政策网络,是政府部门(包括中央以及地方政府机构)与非政府部门共同治理的模式,所有成员参与决策的制定与执行.因此,除了国家法律规定,地方实体和非政府组织的倡议也会对决策制定产生影响.代表性合作平台与战略有波罗的海发展论坛(B D F)㊁«波罗的海二十一世纪议程»(B a l t i c21)等.最后一种类型是跨国网络模式,是指除中央政府外的多层次主体参与(例如地方政府机构㊁非政府组织㊁科研机构㊁商业协会等)的 自治 伙伴关系.代表性组织有波罗的海城市联盟(U B C)㊁潜艇员网络(S u b m a r i n e r N e t w o r k)㊁波罗的海次区域合作(B S S S C)等.这类伙伴关系,在波罗的海跨界海洋空间规划发展的过程中,不仅为其成员搭建了稳定的36。
南黄海布格重力异常的小波多尺度分析-海洋科学

根据上述原理 , 应用小波变换对南黄海布 格重 力数据 ( 图 1) 进行了多尺度分解。本研究区范围为 32 ~ 37. 5 N, 122 ~ 126 E, 布格 重力 异常 资料 为 5 km ! 5 km 网格化数据。 研究表明[ 4] , 随着阶数的增大 , 小波所反映的波 长范围是逐渐增大的。根据重力场理论, 不同波长 的重力变化反映不同深度的密度变化 , 密度变化体
3
南黄海布格重力小波多尺度分解结果
1
南黄海地质构造特征
南黄海地区兼跨中朝、 扬子 和华南三大构造单 元。北部靠近山东半岛, 与北黄海相连, 位于中朝块 体; 南部与东海相连, 位于浙闽隆褶带上 ; 中部为扬 子块体。海区从北向南, 依次有千里岩隆起 , 北部盆 地, 中部隆起, 南部盆地, 勿南沙隆起, 另外还有东部 隆起 , 这些构造被 一系列多 期次、 多层 次的断 阶逼近场 ( 图 6) 表现 平缓, 其场值为 12 ~ 31 mGal, 主要反映莫霍面的起伏和上地幔岩石密 度的变化 , 总体上 看, 本 区莫霍面 由西 向东逐 渐抬 升, 局部发育隆起与坳陷。
F ig . 6
图 6 南黄海布格重力异常四 阶小波变换逼近图 T he for th w avelet transfor m approx imat ion o f the Bouguer g rav ity ano malies in the South Yellow Sea
M arine Sciences/ Vo l. 32, N o. 11/ 2008
研究论文
A RT ICL E
Fig . 4
图 4 南黄海布格重力异常三阶小波变换细节图 T he third o rder wav elet transfor m detail o f Bouguer gr avit y anomalies in t he South Y ello w Sea
我们从大海那里看到了生机和勃发——2008年海洋知识夏令营营员回访

我们从大海那里看到了生机和勃发——2008年海洋知识夏令
营营员回访
佚名
【期刊名称】《海洋世界》
【年(卷),期】2009(000)007
【摘要】2008年7月26日上午,由国家海洋局主办、海洋出版社承办的2008全国海洋知识夏令营活动正式开营。
在5.12汶川地震发生后,为了抚慰四川地震灾区青少年的心灵,用大海的开放、包容和博大的胸怀帮助他们早日走出阴霾,同时向受灾的学生们献出海洋人的一份爱心,此次夏令营邀请了20名来自四川地震重灾区的学生,
【总页数】3页(P34-36)
【正文语种】中文
【中图分类】P7-4
【相关文献】
1.与大海的约会——2008全国海洋知识夏令营纪行 [J], 屠强
2.明年还要参加海洋知识夏令营!——第六届全国海洋知识夏令营随笔 [J], 屠梦晨
3.感受浮潜——青岛39中美国海洋夏令营营员手记 [J], 张亦嘉;王紫剑;张惠
4.尚勇勉励航天夏令营营员:热爱科学成就科技梦想 [J], 无
5.始自2008年——首届全国海洋知识夏令营回顾 [J], <海洋世界>综合;姚海科因版权原因,仅展示原文概要,查看原文内容请购买。
东海近岸大气中的^(210)Po、^(210)Bi和^(210)Pb及其沉降入海通量评估

东海近岸大气中的^(210)Po、^(210)Bi和^(210)Pb及其沉降入海通量评估马瑞阳;钟强强;王浩;杜娟;刘文靖;张洁然;黄德坤;于涛【期刊名称】《海洋学报》【年(卷),期】2024(46)3【摘要】大气^(210)Po、^(210)Bi和^(210)Pb的沉降通量是海洋中核素示踪颗粒物动力学过程(颗粒有机碳输出、颗粒物输运)的基础参数,为揭示我国近海地区^(210)Po、^(210)Bi和^(210)Pb活度浓度的时空变化规律并估算其沉降入海通量,本文于2016年9月至翌年2月和2021年9-11月分别对上海及厦门地区近地表大气气溶胶中^(210)Po、^(210)Pb和^(210)Bi的活度浓度进行了连续观测;基于^(210)Po-^(210)Pb活度比(^(210)Po/^(210)Pb)和^(210)Bi-^(210)Pb活度比(^(210)Bi/^(210)Pb)两种示踪法计算了气溶胶颗粒物的滞留时间,并利用一维简单气溶胶沉降速率模型估算了3种核素以大气沉降方式输入东海的通量。
结果显示,2016年上海秋、冬两季^(210)Po、^(210)Bi、^(210)Pb 3种核素活度浓度的变化范围分别为0.11~1.27 m Bq/m^(3)、0.45~1.83 m Bq/m^(3)和1.12~6.10 m Bq/m^(3);2021年秋季厦门^(210)Po、^(210)Bi、^(210)Pb 3种核素活度浓度的变化范围分别为0.05~0.85 m Bq/m^(3)、0.83~2.52 mBq/m^(3)和0.17~1.32 m Bq/m^(3),上海近地表气溶胶中3种核素的活度浓度秋季平均值比厦门地区高。
利用^(210)Po/^(210)Pb和^(210)Bi/^(210)Pb计算得到上海和厦门近地面大气的气溶胶滞留时间存在显著差异,基于^(210)Po/^(210)Pb计算上海气溶胶滞留时间均值为(94±54)d,基于^(210)Bi/^(210)Pb计算上海气溶胶滞留时间均值为(6.4±4.8)d,造成这种差异的原因很可能是两种示踪法本身具有的系统性差异。
中华虎头蟹幼体发育的初步观察

第27卷第3期大连海洋大学学报Vol.27No.3 2012年6月JOURNAL OF DALIAN OCEAN UNIVERSITY Jun.2012文章编号:2095-1388(2012)03-00 -05中华虎头蟹幼体发育的初步观察王红伟1,原振政1,祁峰2,刘谞2,李晓东2,姜玉声1,刘海映3,郑岩2,赵静1,刘涵1 (1.大连海洋大学水产与生命学院,辽宁大连116023;2.盘锦光合蟹业有限公司,辽宁盘锦124200;3.大连海洋大学海洋科技与环境学院,辽宁大连116023)摘要:对中华虎头蟹幼体发育的过程进行观察㊂结果表明:幼体发育分为4个主要时期,包括3期溞状幼体和1期大眼幼体㊂海水盐度为28,水温为(25.1±0.8)℃时,Ⅰ期溞状幼体历时4~5d发育至Ⅱ期溞状幼体,Ⅱ期溞状幼体历时4~5d发育至Ⅲ期溞状幼体,Ⅲ期溞状幼体历时5~6d发育至大眼幼体,大眼幼体经过5~6d发育至Ⅰ期仔蟹,幼体自孵化完成全部发育过程共需要18~22d㊂幼体大小㊁第1颚足和第2颚足外肢羽状刚毛数目㊁尾叉棘刺数目㊁腹部肢芽大小是区分各时期溞状幼体的主要形态特征㊂关键词:中华虎头蟹;幼体发育;形态特征中图分类号:S966.1 文献标志码:A 中华虎头蟹Orithyia sinica又称乳斑虎头蟹,俗称虎头蟹㊁鬼头蟹,属甲壳纲㊁十足目㊁馒头蟹科㊁虎头蟹属,是中国与朝鲜半岛沿海特有的大型经济蟹类[1],在中国南北沿海均有分布,地处辽东湾的辽宁盘锦㊁营口及丹东鸭绿江口附近海域资源量相对较多㊂中华虎头蟹肉质细嫩,味道鲜美,具有很高的食用价值㊂因每年可捕获的时间短暂,自然资源稀少,市场供不应求㊂此外,中华虎头蟹色泽鲜艳,背面观与中国民间传统的布艺老虎相似,也有学者提出饲养此蟹作为观赏宠物[2]㊂但是随着近些年捕捞强度的增大及栖息环境的恶化,虎头蟹资源日渐衰退,对其生物学与人工繁殖技术的研究已引起人们的重视㊂目前,国内有关中华虎头蟹生物学方面的报道仅见于温度㊁盐度对成蟹存活与摄食的影响[2],以及其视网膜电图特性的昼夜节律性的研究[3]㊂国外,Kim等[4-5]研究了中华虎头蟹溞状幼体和大眼幼体的生长与蜕壳;Koo 等[6-7]研究了温度对中华虎头蟹幼体存活㊁生长㊁蜕壳的影响;Hong[8-9]对中华虎头蟹前溞状幼体和溞状幼体的形态进行了描述㊂蟹类生活史中通常具有多个幼体阶段,幼体经过多次蜕壳,最终发育为仔蟹㊂幼体发育是其生物学研究的重要内容,也是人工繁育的理论依据㊂目前,已有对中华绒螯蟹Eriocheir sinensis[10]㊁三疣梭子蟹Portunus trituberculatus[11]㊁锯缘青蟹Scylla serrata[12]㊁远海梭子蟹Portunus pelagicus[13]和日本蟳Charybdis japonica[14]等主要经济蟹类幼体发育的研究,促进了其增养殖业的发展㊂然而,国内外尚未见有关中华虎头蟹幼体发育完整过程的报道㊂本试验中,作者对中华虎头蟹各期幼体的形态特征及发育时间进行观察与记录,以充实此稀有蟹种的生物学研究,旨在为其人工繁育以及资源的合理利用与保护提供基础资料㊂1 材料与方法1.1 材料试验用亲蟹于2010年5月末由渔船采捕,捕获地点如图1所示㊂将亲蟹运输至辽宁省盘锦市光合水产有限公司码头,共计145只㊂1.2 方法1.2.1 亲蟹培育 将亲蟹饲养于5m×5m×1.5m 的水泥池中,池底铺有10cm细砂,中间用砖块隔出10m2的空地,便于换水与投饵㊂海水盐度为 收稿日期:2011-06-25 基金项目:辽宁省教育厅实验室专项(2008S062);国家海洋局近岸海域生态环境重点实验室资助项目(201019);辽宁省海洋与渔业厅科技计划项目(2011025);辽宁省高等学校优秀人才支持计划项目(LJQ2011074);大连市青年科技人才基金资助项目(2007J23JH023) 作者简介:王红伟(1985-),男,硕士研究生㊂E-mail:woshi415deren@ 通信作者:姜玉声(1977-),男,副教授㊂E-mail:jys@28,温度为25℃,连续充气㊂早晚按体质量的5%投喂光滑河蓝蛤㊁冰鲜鱼各1次,并根据实际摄食情况进行调整㊂每天早㊁晚半量换水㊂1.2.2 幼体培育 各发育时期的幼体均在容积为300L的圆形玻璃缸中培育㊂幼体孵化前加砂滤海水200L㊂培育至Ⅰ期溞状幼体时不换水,每日加注新水;培育至Ⅱ期溞状幼体时开始换水,每日早晚各换水1/5;培育至Ⅲ期溞状幼体时换水量增加至每日1/2;大眼幼体每日全量换水㊂对于Ⅰ期与Ⅱ期幼体,每4h投喂轮虫1次,并补充少量单胞藻及微粒配合饲料㊁虾片等代用饵料;Ⅲ期溞状幼体以卤虫无节幼体为主要饵料;对于大眼幼体投喂卤虫无节幼体和活糠虾㊂幼体培育期间,海水盐度为28,温度为(25.1±0.8)℃,随着幼体的变态逐渐增加充气量㊂1.2.3 幼体发育观察 采用光诱方法,用烧杯捞取不同发育时期的幼体及幼体蜕下的壳置于特制的玻璃凹槽中,于Olympus C-BD230解剖镜下观察并拍照,记录幼体各发育时期与特征,并随机取15只幼体进行形态参数的测量㊂图1 亲蟹的采捕地点Fig.1 The location where where tiger crab brood stocki used in this experiment were collected2 结果2.1 幼体发育时期中华虎头蟹溞状幼体经3次蜕皮变态为大眼幼体,每次蜕皮后,其形态均发生变化,因此将溞状幼体阶段分为3个时期,各发育时期幼体的形态见图2㊂海水盐度为28,水温为(25.1±0.8)℃时,Ⅰ期溞状幼体历时4~5d发育至Ⅱ期溞状幼体;Ⅱ期溞状幼体历时4~5d发育至Ⅲ期溞状幼体;Ⅲ期溞状幼体历时5~6d发育至大眼幼体;大眼幼体经过5~6d发育至Ⅰ期仔蟹㊂本试验条件下,幼体自孵化共需要18~22d发育为仔蟹㊂2.2 幼体的形态中华虎头蟹各发育时期幼体的形态描述见表1和图3㊂溞状幼体分头胸部和腹部,头胸甲上下与左右各具一刺,位于上方的背刺与下方的额刺尤其长,其尖端间距约为体长的3倍㊂前侧各生有一对复眼㊂腹部分六节,最后一节呈叉状,其上具有棘刺,Ⅰ期溞状幼体尾叉棘刺5对,随着蜕壳每期幼体棘刺数目增加1对㊂第二腹节左右生有一对侧齿㊂Ⅰ期溞状幼体腹部2~5节上肢芽不明显,Ⅱ期溞状幼体腹部肢芽明显,至Ⅲ期时肢芽明显长大,呈棒状㊂3 讨论3.1 幼体发育的分期蟹类是甲壳动物中的一个大家族,其幼体发育模式多样,不同种类刚孵化的幼体所处的发育时期有所不同㊂中华绒螯蟹㊁三疣梭子蟹㊁锯缘青蟹等种类先要在卵膜内度过无节幼体和原溞状幼体时期,之后破膜孵化,初孵的幼体称为Ⅰ期溞状幼体[15-17]㊂河流溪蟹Sinopotamon henanens胚胎除了经历上述两个发育时期外,还存在卵内大眼幼体期,经过此期后便直接孵化出仔蟹[18]㊂中华虎头蟹破膜时为溞状幼体,但其溞状幼体只有3期,之后变态为大眼幼体,这与其它已报道的蟹类不同(表2)㊂中华绒螯蟹㊁锯缘青蟹㊁远海梭子蟹腹部肢芽出现在溞状幼体Ⅲ期[13],而中华虎头蟹腹部肢芽出现在溞状幼体Ⅰ期,Ⅱ期时已较明显,后者是否在卵膜内多度过一个或更多溞状幼体时期,还需要对其卵内幼体的发育过程做进一步研究㊂3.2 幼体的形态特征中华虎头蟹溞状幼体和大眼幼体的形态特征与其它几种常见经济蟹类有较多不同(表2),前者2大连海洋大学学报 第27卷注:A Ⅰ期溞状幼体;B Ⅱ期溞状幼体;C Ⅲ期溞状幼体;D 大眼幼体;E 大眼幼体蜕的壳;FⅠ期仔蟹㊂Note:A ZoeaⅠ;B ZoeaⅡ;C ZoeaⅢ;D Megalopa;E Shell of megalopa;F Crab stageⅠ.图2 中华虎头蟹幼体发育过程及形态Fig.2 Different larval stages of tiger crab O .sinica图3 中华虎头蟹各发育时期溞状幼体的特征Fig.3 Morphological characters of different stage zoeae in tiger crab O .sinica较明显的特征是:溞状幼体较大,具有长的背刺与额刺,Ⅰ期和Ⅲ期幼体两刺顶部的间距分别为7.15㊁11.81mm,是体长的3倍左右,与身体大小比例很不相称㊂研究表明,水生动物的幼体能够通过增大或增厚身体表面的结构,如棘刺㊁刚毛㊁壳等,来加强防御能力,进而降低被捕食的风险[19-20]㊂中华虎头蟹多在河口栖息和繁殖,这一区域饵料丰富,往往也是其它鱼㊁虾㊁蟹的索饵与繁殖场,虎头蟹浮游的溞状幼体被捕食的风险性很高㊂长期的自然选择可能导致其溞状幼体保留了如此长的棘刺以应对众多的捕食者㊂然而,捕食者是否对海洋蟹类幼体身体构造具有表型可塑性,表型3第3期 王红伟,等:中华虎头蟹幼体发育的初步观察表1 中华虎头蟹幼体各发育时期的形态Tab.1 Morphological characters of different stage larvale of O.sinica幼体发育时期Stage 形态特征Mor⁃phological characters 个体大小/mm SizeⅠ期溞状幼体第一颚足外肢羽状刚毛数4根;尾叉棘刺5对;腹部肢芽不明显体长2.63~2.65背刺长度3.16~3.55额刺长度4.18~4.71背刺-额刺间距7.15~8.68Ⅱ期溞状幼体第一颚足外肢羽状刚毛数为6根;尾叉棘刺6对;腹部出现4对肢芽体长2.66~2.77背刺长度3.41~3.80额刺长度5.45~6.22背刺-额刺间距9.39~10.10Ⅲ期溞状幼体第一颚足外肢羽状刚毛数为8根;尾叉棘刺7对;腹部4对肢芽明显,呈棒状㊂体长3.61~3.68背刺长度4.43~4.94额刺长度5.56~7.08背刺-额刺间距11.01~11.81大眼幼体期形态与仔蟹相似,腹部能够卷曲于头胸甲下,游泳时伸展,腹部游泳足发达,生有刚毛;具有螯足与其它4对步足体长3.96~4.19头胸甲长2.68~3.04头胸甲宽2.51~2.68结构又是怎样演化为遗传特征的,如此一系列问题将有待进一步研究㊂中华虎头蟹大眼幼体的腹部能够自由弯折,游泳时伸展开来,或卷折于头胸甲下,此时幼体则沉入水底爬行或静止,看上去与仔蟹十分相似㊂这一特征与其它蟹类差异明显,多数蟹类大眼幼体腹部发达但不能自由折叠,凭借发达的腹部,幼体具有较强的游泳能力,利于捕食及躲避敌害功击㊂中华虎头蟹大眼幼体的运动能力则相对较弱,常仰卧或俯卧于养殖容器底部,实验室养殖时,需要较大的饵料密度才能保证幼体的摄食量㊂如此特殊的大眼幼体在自然环境中如何捕食及躲避敌害的追捕,将有待于进一步研究㊂3.3 幼体发育时期的判断虾蟹类人工繁育中需要准确区分各发育时期的幼体,以便及时调整投喂㊁换水等技术措施㊂本试验结果表明,幼体大小㊁第1和第2颚足外肢羽状刚毛数目㊁尾叉棘刺数目㊁腹部肢芽大小是中华虎头蟹溞状幼体分期的主要依据㊂中华虎头蟹溞状Ⅰ期与Ⅱ期幼体大小差异并不明显,加之不同母本的同一期幼体间大小有时也存在差异,难以从大小上表2 常见经济蟹类溞状幼体与大眼幼体的形态特征[13]Tab.2 Morphological comparison of zoeal and megalop larvae in several commercially important crabs[13]种类species 溞状幼体分期与形态特征stages and characters时期第1颚足外肢末端羽状刚毛数/根腹部肢芽出现时期大眼幼体形态特征stages and characters of megalopa体长/mm头胸甲第1触角外肢参考文献reference中华虎头蟹ⅠⅡⅢ468Ⅰ3.96~4.19两侧呈双角状突起5节,第4~5节具感觉毛,数目为8㊁2本文中华绒螯蟹ⅠⅡⅢⅣⅤ4681012Ⅲ4.90~5.36两侧呈双角状突起4节,第2~4节各具一排感觉毛,数目为4㊁4㊁4㊂第3节具2~4根刚毛梁象秋等[10]三疣梭子蟹ⅠⅡⅢⅣ461013~14Ⅲ3.70~4.16较圆钝,两侧不呈双角状突起5节,第2~4节各具一排感觉毛,数目为4㊁6㊁8㊂第5节具2根刚毛孙颖民等[11]锯缘青蟹ⅠⅡⅢⅣⅤ46810~1112~13Ⅲ3.55较圆钝,两侧不呈双角状突起5节,第1~4节各具一排感觉毛黄胜南等[12]远海梭子蟹ⅠⅡⅢⅣⅤ48101212Ⅲ3.20~3.67较圆钝,两侧不呈双角状突起5节,第2~4节各具一排感觉毛,数目为8㊁6㊁5㊂第3节具1根刚毛,第5节具2根刚毛廖永岩等[13]日本蟳ⅠⅡⅢⅣⅤVI4689~1011~1213-14Ⅳ3.58~3.84较圆钝,两侧不呈双角状突起5节,第2~5节各具一排感觉毛㊂第3节具1根刚毛(光滑)㊂第5节具2根刚毛(1根光滑,1根刺状)阎愚等[14]4大连海洋大学学报 第27卷判断幼体时期,给生产操作造成了困难㊂这一时期可以借助仪器观察幼体腹部肢芽是否明显㊁第1和第2颚足外肢羽状刚毛的数目㊁尾叉棘刺数目等特征进行判断,而对于有经验的技术人员则可直接观察水体中幼体蜕下的壳,判断幼体是否进入下一时期㊂另外,笔者发现,从Ⅱ期溞状幼体开始其背刺与额刺中上部呈现红色,此特征无需借助仪器设备即可分辨,便于在实际生产中应用㊂参考文献:[1] 沈嘉瑞,刘瑞玉.我国的虾蟹[M].北京:科学出版社,1976.[2] 廖永岩,吴蕾,蔡凯,等.盐度和温度对中华虎头蟹存活和摄饵的影响[J].生态学报,2007,27(2):627-629.[3] 柴敏娟,郑微云.几种甲壳动物视网膜电图特性的昼夜节律性比较[J].海洋与湖沼,1990,21(3):212-217.[4] Kim Y H,Studies on Orithyia sinica (Linnaeus)in the WestemSea 2Weight of ovary,number of eggs spawned and egg diameter [J].Bulletin of the Korean Fisheries Society.Pusan,1983,16(1):14-16.[5] Kim Y H,Chung S C.Studies on the growth and molting of the ti⁃ger crab,Orithyia sinica (Linnaeus)[J].Bulletin of the KoreanFisheries Society Pusan,1990,23(2):93-108.[6] Koo J G,Jee J H,Kim J M,et al.Effect of water temperature onsurvival,growth and intermolt period of tiger crab,Orithyia sinica (Linnaeus)larvae[J].Journal of Fish Pathology,2004,17(2):139-144.[7] Koo J G,Kim S G,Jee J H,et al.Effects of ammonia and nitrite onsurvival,growth and moulting in juvenile tiger crab,Orithyia sinica (Linnaeus)[J].Aquaculture Research,2005,36(1):79-85.[8] Hong S Y.Zoeal stages of Orithyia sinica (Linnaeus)(Decapoda,Calappidae)reared in the laboratary[J].Publ Inst Mar Sci Nntl Fish Univ Busan,1976,9:17-23.[9] Hong S Y.The prezoeain various decapod crustaceans[J].Joumal of Natural Histary,1988,22(4):1041-1075.[10] 梁象秋,严生良,郑德崇,等.中华绒螯蟹Eriocheir sinsis H.Milne-Edwards 的幼体发育[J].动物学报,1974,20(1):61-75.[11] 孙颖民,阎愚,孙进杰.三疣梭子蟹的幼体发育[J].水产学报,1984,8(3):219-226.[12] 黄胜南,李婉丽.锯缘青蟹Scylla serrata (Forskal)幼体发育的研究[J].水产学报,1965,2(4):24-34.[13] 廖永岩,余波,董学兴.远洋梭子蟹幼体发育的研究[J].台湾海峡,2001,20(4):533-546.[14] 阎愚,孙颖民,宋志乐.日本蟳幼体发育的研究[J].水产学报,1989,13(1):74-79.[15] 堵南山,赵云龙,赖伟.中华绒整螯蟹胚胎发育的研究[C]//甲壳动物学论文集:第2辑.青岛:青岛海洋大学出版社,1992:128-135.[16] 韦受庆,罗远裕.青蟹胚眙发生的研究[J].热带海洋,1986,5(3):57-62.[17] 薛俊增,堵南山,赖伟.三疣梭子蟹胚胎发育早期的组织学研究[J].动物学研究,2001,22(1):69-73.[18] Pace F,Harris R,Jaccarini V.The embryonic development of theMeditemanean freshwater crab[J].Journal of Zoology,1976,180(1):93-106.[19] Agrawal A A.Phenotypic plasticity in the interactions and evolu⁃tion of species[J].Science,2001,294:321-326.[20] Bronmark C,Miner J G.Predator induced phenotypical change inbody morphology in crucian carp [J].Science,1992,256:1348-1350.Larval development of tiger crab Orithyia sinicaWANG Hong⁃wei 1,YUAN Zhen⁃zheng 1,QI Feng 2,LIU Xu 2,LI Xiao⁃dong 2,JIANG Yu⁃sheng 1,LIU Hai⁃ying 3,ZHENG Yan 2,ZHAO Jing 1,LIU Han 1 (1.College of Fisheries and Life Science,Dalian Ocean University,Dalian 116023,China;2.Panjin Guanghe Crab Industrial Co.Ltd.,Panjin124200,China;3.College of Marine Science and Environment,Dalian Ocean University,Dalian 116023,China)Abstract :Larval development of tiger crab Orithyia sinica was observed under a light microscope at a water temper⁃ature of (25.1±0.8)℃.It was found that 4larval stages including 3zoeal stages and 1megalop stage were distin⁃guished by the first stage of juvenile crab.The larvae hatched as a zoea (Z)which was metamorphosed into the first stage juvenile crab by 4times of moult,during which ZⅠlarvae developed into ZⅡlarvae in 4-5days,ZⅡlarvae developed into ZⅢlarvae in 4-5days,and ZⅢlarvae lasted for 5-6days,then became megalop whichmolted into juvenile crab 5-6days later.The whole process of larval development took place during 18-22d.The developmental stages of the larvae were primarily distinguished by body size,number of hair and spine on the max⁃illiped and telson,and size of pleopodal buds.Key words :tiger crab Orithyia sinic ;larval development;morphological character5第3期 王红伟,等:中华虎头蟹幼体发育的初步观察。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
---------------------------------------------------------------最新资料推荐------------------------------------------------------
海洋学(二)--耿晨曦
一、不会。
根据地壳均衡说,地壳漂浮于液体底层之上,地壳高的地方地壳较厚,即高山会有深的山根作为补偿,如果山峰被侵蚀,其会上浮,实际降低小于不会。
根据地壳均衡说,地壳漂浮于液体底层之上,地壳高的地方地壳较厚,即高山会有深的山根作为补偿,如果山峰被侵蚀,其会上浮,实际降低小于 500m。
当有大量物质沉积在某一特定区域,新沉积物的巨大质量将会使地壳下沉。
同样,当大量的物质在特定地区被侵蚀,地表会上升以弥补。
因此,当山脉被侵蚀而降低高地,地壳将会向上回跳(回跳量较侵蚀量小),(在一定程度上)造成进一步侵蚀。
现在某些可在地表见到的地层可能曾经在地壳深处被其他地层覆盖,但因为上方地层被侵蚀消失,使下方地层向上回跳而出现在地表。
当有大量物质沉积在某一特定区域,新沉积物的巨大质量将会使地壳下沉。
同样,当大量的物质在特定地区被侵蚀,地表会上升以弥补。
因此,当山脉被侵蚀而降低高地,地壳将会向上回跳(回跳量较侵蚀量小),(在一定程度上)造成进一步侵蚀。
1 / 7
现在某些可在地表见到的地层可能曾经在地壳深处被其他地层覆盖,但因为上方地层被侵蚀消失,使下方地层向上回跳而出现在地表。
二、因为海水是不断流动的。
时间太久会导致地磁场改变。
三、我国海岸带在历史上经历了多次变迁,海岸线有的像陆地推进,有的向海洋推进。
其原因有自然原因,也有不可忽视的人为因素。
我国海岸带在历史上经历了多次变迁,海岸线有的像陆地推进,有的向海洋推进。
其原因有自然原因,也有不可忽视的人为因素。
海岸主要受海水动力因素侵蚀所产生的各种形态,又称海蚀地貌。
它是海岸地貌的一大类别。
怎么会被侵蚀?塑造海岸侵蚀地貌的主要动力因素是波浪和潮流,但高纬度地带的海岸还受到冰冻的侵蚀,热带和亚热带的海岸则受到丰富的地表水和强烈的化学风化作用的侵蚀。
海岸侵蚀地貌的发育过程,除与沿岸海水动力的强弱和海岸的纬度地带性有关以外,还受组成海岸的岩性的抗蚀能力所制约。
结构致密、坚硬岩石海岸,抗蚀能力较强,但因裂隙和节理发育,多海蚀洞、海蚀拱、海蚀柱、海蚀崖。
松软岩石海岸,抗蚀能力较海岸主要受海水动力因素侵蚀所产生的各种形态,又称海蚀地貌。
---------------------------------------------------------------最新资料推荐------------------------------------------------------ 它是海岸地貌的一大类别。
怎么会被侵蚀?塑造海岸侵蚀地貌的主要动力因素是波浪和潮流,但高纬度地带的海岸还受到冰冻的侵蚀,热带和亚热带的海岸则受到丰富的地表水和强烈的化学风化作用的侵蚀。
海岸侵蚀地貌的发育过程,除与沿岸海水动力的强弱和海岸的纬度地带性有关以外,还受组成海岸的岩性的抗蚀能力所制约。
结构致密、坚硬岩石海岸,抗蚀能力较强,但因裂隙和节理发育,多海蚀洞、海蚀拱、海蚀柱、海蚀崖。
松软岩石海岸,抗蚀能力较差,海蚀崖后退较快,易形成海蚀平台。
石灰岩海岸,在海水溶蚀下具有独特的蜂窝状海蚀地貌形态。
海蚀地貌通常被作为判别地区构造运动和海平面变化的标志之一。
被侵蚀是属于怎样一种情况?比如中国。
中国海岸侵蚀具有普遍性、多样性和加剧发展第三个特点,侵蚀岸线长度占全国大陆岸线的差,海蚀崖后退较快,易形成海蚀平台。
石灰岩海岸,在海水溶蚀下具有独特的蜂窝状海蚀地貌形态。
海蚀地貌通常被作为判别地区构造运动和海平面变化的标志之一。
被侵蚀是属于怎样一种情况?比如中国。
中国海岸侵蚀具有普遍性、多样性和加剧发展第三个特点,侵蚀
3 / 7
岸线长度占全国大陆岸线的 1 /3 以上,尤以渤海和黄海沿岸较重。
但因沿海泥沙来源丰富、堆积条件较好,使总体侵蚀强度通常稍轻于国外许多海岸。
中国海岸的侵蚀因素有海平面上升、风暴潮加剧、入海泥沙减少和多种不合理的人为开发活动等以上,尤以渤海和黄海沿岸较重。
但因沿海泥沙来源丰富、堆积条件较好,使总体侵蚀强度通常稍轻于国外许多海岸。
中国海岸的侵蚀因素有海平面上升、风暴潮加剧、入海泥沙减少和多种不合理的人为开发活动等 9 种,其中种,其中 7 种与全球变化有直接或间接关系。
它们主要通过增强海洋动力作用、减少沿岸泥沙和降低海岸稳定性三个原因,加剧海岸侵蚀。
岛屿会越变越小?海岸侵蚀导致一些地区的地下水质被破坏、盐碱地增加、海滨浴场和渔港遭到破坏,沿岸农田、公路和居民区也受到威胁。
导致海岸侵蚀的原因很多,除风暴潮等引起的自然过程外,生态恶化、径流减少、不合理采砂、护岸植被破坏等是重要的人为原因。
海岸侵蚀会使海岸线减短,比如海水倒灌。
所以导致海岛越变越小。
种与全球变化有直接或间接关系。
它们主要通过增强海洋动力作用、减少沿岸泥沙和降低海岸稳定性三个原因,加剧海岸侵蚀。
---------------------------------------------------------------最新资料推荐------------------------------------------------------ 岛屿会越变越小?海岸侵蚀导致一些地区的地下水质被破坏、盐碱地增加、海滨浴场和渔港遭到破坏,沿岸农田、公路和居民区也受到威胁。
导致海岸侵蚀的原因很多,除风暴潮等引起的自然过程外,生态恶化、径流减少、不合理采砂、护岸植被破坏等是重要的人为原因。
海岸侵蚀会使海岸线减短,比如海水倒灌。
所以导致海岛越变越小。
其实,对于近些年来海岸带的变迁主要是人为因素所导致的,人类活动对自然造成了巨大的影响。
主要的人为因素有:
其实,对于近些年来海岸带的变迁主要是人为因素所导致的,人类活动对自然造成了巨大的影响。
主要的人为因素有:
1 、乱砍滥伐人类活动对植被的破坏使得大量的泥沙进入河水中,而河水携带大量泥沙流入海洋时,沙在海岸附近堆积起来,长年累月,沉积为陆地,这时海岸线就会向海洋推移。
我国黄河就是含沙量巨大,每年人类活动对植被的破坏使得大量的泥沙进入河水中,而河水携带大量泥沙流入海洋时,沙在海岸附近堆积起来,长年累月,沉积为陆地,这时海岸线就会向海洋推移。
我国黄河就是含沙量巨大,每年携带大量泥沙沉积在河口,使得海岸带向外延伸。
5 / 7
2 、港口建设据统计,我国的港口建设是近年来我国海岸线变化的最主要因素。
港口建设使用了大量先进的大型器材,我国拥有不少的港口,而这些港口的建设占据了很大的沿海面积,大量的工程建设使得海岸带明显的向外扩张。
据统计,我国的港口建设是近年来我国海岸线变化的最主要因素。
港口建设使用了大量先进的大型器材,我国拥有不少的港口,而这些港口的建设占据了很大的沿海面积,大量的工程建设使得海岸带明显的向外扩张。
3 、填海造陆围海造田等人为的填海活动都直接地使得海岸带向海洋延伸。
说到填海造地,人们可能自然而然的想到荷兰,确实荷兰用其先进的技术,填海造地,大大拓展了他们的土地面积,而现在我国也进行了大量的填海造地工程,使得海岸带向外扩展。
围海造田等人为的填海活动都直接地使得海岸带向海洋延伸。
说到填海造地,人们可能自然而然的想到荷兰,确实荷兰用其先进的技术,填海造地,大大拓展了他们的土地面积,而现在我国也进行了大量的填海造地工程,使得海岸带向外扩展。
4 、径流变化河流的流向不是一成不变的,或因为自然原因或人为原因,我国几大河流都经历过改道,而河流改道会形成新的河口,造成海岸带的变化。
如黄河改道可能使得新形成的河口出现大量淤积,使得海岸线快
---------------------------------------------------------------最新资料推荐------------------------------------------------------ 速的扩张。
河流的流向不是一成不变的,或因为自然原因或人为原因,我国几大河流都经历过改道,而河流改道会形成新的河口,造成海岸带的变化。
如黄河改道可能使得新形成的河口出现大量淤积,使得海岸线快速的扩张。
5 、其他因素盐场建设、渔业设施建设等沿海的工程项目,其大都会占领海洋,使得海岸线向海洋扩展。
盐场建设、渔业设施建设等沿海的工程项目,其大都会占领海洋,使得海岸线向海洋扩展。
7 / 7。