与肌肉代谢和结构有关的不同肉质表型识别基因的猪种间的肌肉转录组比较
肌肉组织学特性与肉品品质的关系

养殖技术顾问 2010. 8
誅 检疫检验
过程中一些简单的前体物质如氨基酸、糖和脂肪酸 等可以降解为许多小分子的醛、酮、胺类和其他化合 物,这些化合物既可以直接影响肉的风味,又可以相 互作用或通过某些途径与其他化合物反应生成香味 物质。
4 小结
畜禽的肌肉品质是畜禽最重要的经济性状。通 过对畜禽的肌纤维组织学特性、结缔组织和脂肪组 织与肉品质关系的分析,发现要改善畜禽肉品质,有 必要对肌纤维组织学特性及其分类的遗传机制进行 深入研究。
1 肌纤维与肉品质的关系 1.1 肌纤维直径
研究表明,肌纤维直越细,肉的嫩度越好;肌纤 维越粗,肉的嫩度越差[1]~[3]。猪的肌纤维直径与肉质之 间的关系进行了研究,发现我国地方猪种肌纤维直 径比引进猪种的肌纤维直径要细,相应的肉品的嫩 度也较好。对九龙牦牛和大通牦牛肌肉纤维直径进 行研究,证明了肌纤维直径与肉品的柔嫩度呈现曲 线相关[4]~[6]。还有研究结果也证明,肌纤维越细,肉品 嫩度越好[7]。对藏山羊的组织学研究发现藏山羊肌纤 维直径小,肉质细嫩,品质良好。对马鹿肌纤维组织 学特性的研究发现肌纤维越细,嫩度越好。在较粗的 肌纤维中,肌原纤维缔合比较牢固,较难被切断。
饲料检验判定原则为 A 类指标,铅、氟、铜、黄曲 霉毒素 B1、沙门氏菌、盐酸克仑特罗;B 类指标,水分 (微量元素预混合饲料不判定)、感官;C 类指标,标 签;D 类指标,砷。
其中 A 类指标 1 项指标不合格则为不合格;B 类指标中有 2 项以上(含 2 项)不合格则为不合格。 标签单独判定;D 类指标不判定。
陈 刚 (安徽省蒙城亳州师范高等专科学校 233500)
为更好的了解目前中国饲料产品的合格状况, 研究影响产品不合格的因素,寻找提高产品合格的 有效手段,根据农牧饲便函〔2002〕25 号《2002 年全 国饲料和饲料添加剂质量监督检测实施细则》(以 下简称《实施细则》)的要求,全国饲料检测系统对 北京等 6 个省、直辖市的饲料的生产、经营合格率进 行了大规模的监督抽查。各地基本按照《实施方案》 中的要求进行抽样、检验、判定、总结和上报。国家饲 料检测中心对全国部分省份的饲料产品连续几年进 行跟踪检测,现将检测结果报告如下。
我国近十年猪肌肉品质的研究-精品文档

报告人:经荣斌
我国养猪业和其他科技工作者, 近 10 年来,对猪肌肉品质进行 了持续不断大量的研究工作, 取得了明显的进展和丰硕的科 研成果,本文对其进行综述, 以供同行参考。
目 录
一、我国培育的瘦肉型品系(母系)猪的肉质指标
二、杂交对肌肉品质影响的研究 三、猪肉质性状侯选基因的研究 四、肌肉肌纤维的超微结构研究 五、营养对猪肌肉品质影响的研究 六、不同添加剂对猪肌肉品质的影响 七、猪肉质测定方法和测定仪器的研究
65.59
68.61 66.53 62.05 67.64
杜×大
2.5b
5.54A
33.44Aa
67.05
2.2、杂交对肌纤维直径、横截面积的影响
吴德、杨风等的研究结果:平均肌纤维直径和肌纤维横截面 积梅山猪杂种猪间则差异不显著,但杜×大>杂交>梅(p<0.05)
表3
血缘 梅山猪 1/2梅山
不同体重背最长肌肌纤维直径、横截面积
表5 背最长肌氨基酸、肌苷酸含量的比较
1/2地方猪 必需氨基酸(%) 鲜味氨基酸(%) 氨基酸总量(%) 必需氨基酸/氨基酸总量 (%) 鲜味氨基酸/氨基酸总量 (%) 肌苷酸含量(mg/g) 10.84±0.61 16.33±1.14 23.29±1.51 46.59±0.02 70.10±0.01 2.31±0.16a 1/4地方猪 10.26±1.39 17.07±1.71 23.25±1.37 44.16±0.06 73.42±0.06 2.45±0.19a 杜洛克 11.36±0.82 17.09±1.66 24.13±2.02 47.11±0.01 70.78±0.02 1.86±0.19b
1488
猪TNNC1基因和TNNC2基因在 肌肉组织中的发育性表达研究

猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达研究【摘要】本研究旨在探讨猪TNNC1基因和TNNC2基因在肌肉组织发育过程中的表达特点。
通过实验方法的使用,我们发现这两个基因在不同发育阶段表达量和模式存在差异,进一步揭示了它们在肌肉组织发育中的作用机制。
实验结果显示,TNNC1基因在早期发育阶段表达量较高,而TNNC2基因在后期发育阶段呈现上调趋势。
数据分析进一步验证了这一结论。
综合研究结论和讨论,我们认为猪TNNC1基因和TNNC2基因在肌肉组织中具有不同的发育性表达特点,这对于深入理解肌肉组织发育过程具有重要意义。
未来的研究可以进一步探究这两个基因在肌肉疾病发生发展中的作用,为相关疾病的治疗提供新的理论依据。
【关键词】猪、TNNC1基因、TNNC2基因、肌肉组织、发育性表达、研究、背景、目的、作用、表达模式、实验方法、实验结果、数据分析、特点、讨论、展望。
1. 引言1.1 研究背景肌肉组织中的TNNC1基因和TNNC2基因在发育过程中起着重要的作用,它们编码肌钙蛋白的一种亚单位,参与肌肉收缩过程。
研究表明,TNNC1基因和TNNC2基因在不同发育阶段的肌肉组织中表达量和表达模式有所不同,这与肌肉组织的生长和发育密切相关。
1.2 研究目的研究的目的是探究猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达特点,从而深入了解这两个基因在肌肉发育过程中的作用机制。
通过分析它们在不同发育阶段肌肉组织中的表达模式,可以揭示这两个基因在肌肉细胞增殖、分化和肌肉纤维形成等生物学过程中的重要功能。
通过研究猪TNNC1基因和TNNC2基因的发育性表达,可以为肌肉生长与发育的调控机制提供重要依据,为肌肉相关疾病的治疗和疾病筛查提供新的思路和方法。
通过本研究,还可以为进一步探索肌肉组织中其他相关基因的表达模式和功能提供参考,为肌肉组织发育领域的研究提供新的视角和方向。
2. 正文2.1 TNNC1基因和TNNC2基因的作用TNNC1基因和TNNC2基因是肌肉组织中的重要基因,它们都编码肌钙蛋白C(TnC),是钙离子在肌肉收缩过程中的调节因子。
猪肌肉中肌内脂肪的研究进展

猪种
梅山猪 1/2梅山猪 3/8梅山猪 1/4梅山猪 1/8梅山猪
IMF(%)2 7.2BC
5.82b
4.8b
3.27Aa
4.08Aa
注: 1.吴德等(2001).转引自.经荣斌等(2005)养猪学分会三届五次会刊(四川、绵阳) 2.肩注字母不同者差异显著(P<0.05),下同。
+1.18*
+2.01*
+2.01**
+2.24**
+2.57*
+1.45*
+2.86 **
+2.01 *
注:1.资料来源,陈润生(1989a,1989b) 2.LD=猪背最长肌;SM=猪半膜肌 3.与各自对照猪差异(+或-)是以百分单位来表示的。 4.*P<0.05, **P<0.01, 下同。
2.1 品种(系)间差异
1.前言
在我所建议的五项性状中,肌内脂肪(Intramuscular fat, 缩写为IMF,下同)对于肉的口感特性(嫩度、多汁性、滋 味……)有重要影响。国内自上世纪80年代初陈润生等 (1989a,1989b)对有代表性的中国一些地方猪的肌内脂 肪测定以来,近年来吴德等(2001)对梅山猪及其杂种猪 和曾永庆等(2004)对莱芜猪及其杂种猪的肌内脂肪含量 也做了研究。从总体看来,国内的研究还处在定量测定和 IMF与其他肉质性状的表型相关的水平上,每个组合的供测 猪样本数少则1-3头,多则不超过10头,从所取得的数据很 难分析和总结出可靠的结论,也难以在实际选择和育种改 良工作中利用。本文主要介绍国外研究者近年来对IMF的研 究方法、内容和取得的成果,以期对国内扩展IMF的研究领 域、深化研究内容、提高研究水平、促进研究成果在猪育 种改良计划中的实际应用进程有所启发和借鉴。
四川农业大学发现猪肉更好吃的秘密

四川农业大学发现猪肉更好吃的“秘密”
严循东摘录 2021年6月23日
据介绍,骨骼肌占到产肉动物体重约五分之二,其主要组成单元是肌纤维,分为慢速氧化型(型)、快速氧化型(a型)、快速酵解型(b型)等4 种,肌纤维类型的差异是影响产肉动
物肌肉品质的重要因素之一。
金龙说,过去大量研究通常将多种骨骼肌视作同一类组织,对于不同部位骨骼肌的遗传和转录
调控特性并未有精细深入地探究。
此次研究利用空间转录组学技术,揭示了3种不同肌纤维亚
型在能量代谢和脂质沉积上的差异;进一步完整构建了从头部、前肢、躯干到后肢,共47种
不同部位骨骼肌的精细转录调控图谱等,将为农业动物产肉性状提供重要指导。
此外,以往受研究方法的限制,对猪脂肪组织更多是基因二维线性的认识,一定程度上制约了
猪的肉质性状形成机制的进一步挖掘。
此次研究在现有猪参考基因组基础上,补充注释了大量
调控性转录本;采用高通量染色质空间构象捕获技术重构了猪脂肪组织的三维基因组空间结构。
“在二维的‘线性’基础上,提供了基因调控的三维‘空间’信息,相当于你要了解一个地方,拿3D
图与平面照相比,前者肯定更清楚更有参考意义。
”金龙说,此次研究还为下一步分子育种的
开展提供了重要基础数据和理论支撑;为促进猪作为人类生物学和疾病的生物学模型奠定了基础。
北京黑猪肉质性状的研究及其背最长肌转录组差异分析

北京黑猪肉质性状的研究及其背最长肌转录组差异分析随着经济和技术的快速发展,人们的生活水平不断提高,对猪肉的消费观念从量转向质,日益下降的猪肉品质与人们对高品质猪肉的追求之间的矛盾,给养猪生产者提供了新的方向。
猪肉品质主要受遗传、营养、屠宰日龄和其他环境因素的影响,本实验以北京黑猪为实验材料,探索北京黑猪的体重、背膘、部分肉质性状以及脂肪酸组分的发育性变化规律,初步确定北京黑猪的最佳屠宰日龄;通过分析不同能量水平日粮饲喂的北京黑猪肉质差异,探索能量水平对北京黑猪体重、背膘和部分肉质性状和脂肪酸组分的影响;同时,对60日龄、120日龄和180日龄的北京黑猪背最长肌进行转录组测序,筛选不同日龄之间的差异表达基因,并对差异表达基因进行功能富集分析,探索不同发育阶段北京黑猪背最长肌中转录水平的整体变化,为不同发育阶段北京黑猪相关性状的变化提供分子依据。
通过对60d、120d、180d、210d、240d、270d和300d的北京黑猪体重、背膘、部分肉质性状以及脂肪酸组分的研究发现,随日龄的增加,北京黑猪的背膘、体重、肌内脂肪和肉色a*(红度)有增大的趋势,剪切力有下降的趋势;对脂肪酸含量(mg/g)的影响表现为:随日龄的增加,总脂肪酸、饱和脂肪酸和单不饱和脂肪酸总体上为上升的趋势,240d后变化较小,多不饱和脂肪酸基本稳定;对脂肪酸比例的影响表现为:随日龄的增加,饱和脂肪酸和单不饱和脂肪酸比例有增大的趋势,多不饱和脂肪酸比例下降。
因此,随日龄的增加,北京黑猪的肉品质整体上有一定程度的提高。
以改善肌内脂肪、肉色和脂肪酸组分为主要目标,兼顾其他因素,初步确定北京黑猪的最佳屠宰日龄为240d。
通过对不同能量水平日粮饲喂的北京黑猪体重、背膘、部分肉质性状以及脂肪酸组分的研究发现,与对照组相比,低能量组猪的pH(24h)显著降低,24h滴水损失和肉色L显著升高,其他性状如终末体重和肌内脂肪等差异不显著,其中背膘有下降的趋势(P>0.05),剪切力有增大的趋势(P>0.05);实验中能量水平对脂肪酸含量和比例的影响均为达到显著水平。
猪TNNC1基因和TNNC2基因在 肌肉组织中的发育性表达研究

猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达研究1. 引言1.1 研究背景肌肉组织中的发育性基因表达对于动物生长发育及肌肉质量的形成至关重要。
TNNC1和TNNC2基因作为肌肉细胞中的重要调控因子,在肌肉组织中发挥着重要作用。
TNNC1基因编码的蛋白质与肌肉收缩相关,而TNNC2基因编码的蛋白质则与肌肉细胞的钙离子调控密切相关。
对这两个基因在肌肉组织中的发育性表达进行研究,有助于揭示肌肉生长发育的分子调控机制。
目前,关于猪TNNC1和TNNC2基因在肌肉中的表达与调控的研究还比较有限。
针对这一问题,本研究旨在探究猪TNNC1和TNNC2基因在肌肉组织中的表达模式及其在肌肉发育过程中的调控机制,为理解动物肌肉生长发育提供更多的分子生物学依据。
通过研究这两个基因在猪肌肉组织中的表达变化及其调控因素,可以为肌肉质量的改良和遗传育种提供重要的理论依据。
【2000字】1.2 研究目的本研究旨在探究猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达特征及其调控机制。
通过对这两个基因在不同发育阶段和不同组织样本中的表达情况进行研究,揭示它们在猪肌肉发育过程中的作用机制和调控网络。
通过探讨猪TNNC1基因和TNNC2基因在肌肉组织中的相互作用及共同调控情况,为深入理解肌肉发育过程中的分子机制提供新的线索和思路。
通过本研究的开展,有望为肌肉组织发育性表达调控的研究提供重要的实验数据和理论基础,为改良猪肉品质、促进肉类生产提供科学依据,并为肌肉相关疾病的治疗和防治提供新的思路和方法。
通过对猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达研究,为深入探究肌肉发育的分子机制和调控网络提供新的研究视角和理论依据。
【字数:200】1.3 研究意义肌肉组织的发育过程是一个复杂而精密的调控过程,其受多种基因的调节和调控。
猪TNNC1基因和TNNC2基因作为肌肉收缩调节蛋白的重要成员,在肌肉组织中发挥着重要作用。
猪TNNC1基因和TNNC2基因在 肌肉组织中的发育性表达研究

猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达研究引言肌肉组织是猪体内重要的组织之一,对于肌肉组织的发育和生长过程了解不仅对于畜牧业的发展具有重要意义,同时也对于人类健康和营养具有一定的指导意义。
肌肉组织的发育受到多种因素的调控,包括基因的表达调控等。
在猪的肌肉发育过程中,TNNC1基因和TNNC2基因被认为是关键基因之一,通过调控这两个基因的表达情况可以影响猪肌肉组织的发育过程。
本研究旨在探讨猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达规律,为猪肌肉组织的发育调控提供理论依据和实验支撑。
材料与方法1. 标本采集:选择不同发育阶段的猪进行标本采集,包括出生后1周、3周、6周和成年期。
分别采集肌肉组织样本用于后续实验。
2. RNA提取与定量:采用Trizol法提取肌肉组织中的总RNA,然后通过实时荧光定量PCR法对TNNC1基因和TNNC2基因的表达量进行定量测定。
3. 蛋白提取与Western blot:提取肌肉组织中的总蛋白,然后通过Western blot法对TNNC1和TNNC2蛋白的表达水平进行检测。
4. 数据统计与分析:对实验数据进行统计分析,采用SPSS软件进行方差分析等统计方法对不同发育阶段样本的基因和蛋白表达变化进行比较分析。
结果通过实时荧光定量PCR实验发现,TNNC1基因在不同发育阶段的猪肌肉组织中的表达量呈现出逐渐增加的趋势,出生后1周时表达量最低,成年期表达量最高。
而TNNC2基因的表达量则在出生后1周到3周之间呈现出较大的增加,之后的表达量相对稳定。
通过Western blot实验结果显示,TNNC1蛋白的表达趋势与基因表达趋势一致,而TNNC2蛋白的表达水平在不同发育阶段的变化并不明显。
讨论本研究结果表明,猪TNNC1基因和TNNC2基因在肌肉组织中的发育性表达存在一定的规律性。
TNNC1基因的表达量随着发育阶段的增加呈现出逐渐增加的趋势,而TNNC2基因则在出生后1周到3周之间表达量出现明显的增加。
猪背最长肌肌纤维类型的发育性变化及其品种和性别特点

89
猪背最长肌肌纤维类型的发育性变化 及其品种和性别特点
杨晓静, 赵茹茜3 , 陈 杰, 胥清富, 韦习会, 陈 杰 ( 南京农业大学 农业部动物生理生化重点
肌纤维组成发生了急剧的变化yhc型和2a型表达下调比例下降2b型肌纤维显著上升p90日龄以后二花脸公猪背最长肌中myhc型和2a型的比例均显著高于大白猪同时大白猪背最长肌中酵解型肌纤维2b的比例增加迅速显著地高于二花脸p性别间比较myhc型纤维的比例无显著差异90二花脸母猪的myhc2b的比例显著高于二花脸公猪p0105
引物序列 2agcctctttcttctcccagggacattc23′ F: 5′ 2cacttgctaagagggacctctgag ttca23′ F: 5′ 2catctgg taacataagagg tacatctag23′ F: 5′ 2ctttcctcataaagcttcaag ttctgcc23′ F: 5′ 2atccaggctgcg taacgctctttgagg ttg ta23′ R: 5′
2 结果
211 体质量及背最长肌质量的发育性变化 见表 2。从 3 日 龄到 180 日 龄, 大 白 猪 体 质 量 均 显 著 高 于 二 花 脸 猪 ( P < 0105 ) ; 雄性二花脸猪体质量与雌性无明显差异 ( P > 0105 ) , 180 日龄时雄性有高于雌性的趋势, 即 ( 60167 ± 4125 ) kg 比 ( 52133 ±6178) kg, 但统计差异不显著 ( P > 0105 ) 。 背最长肌
基因组学和转录组学分析肌肉发育和肉质特性

基因组学和转录组学分析肌肉发育和肉质特性从前人类一直在寻找控制肌肉发展和肉质特性的基因和信号通路以期达到优化肉品生产的目的。
而近年来基因组学和转录组学技术的发展为这个目标的实现提供了新的希望。
基因组学分析基因组学是研究基因组的结构、组成、功能并应用于生物学、医学、进化学、农学等领域的科学。
基因通过转录和翻译被转译成蛋白质,而肌肉发育和肉质特性是由一系列蛋白质的互相作用所决定的。
通过基因组学分析,我们可以识别出影响肌肉发展和肉质特性的基因,为选择适合肉品生产的品种和育种计划提供基础。
一项针对猪肉的研究表明,选择性育种会导致一些重要的基因组变化,影响了肌肉生长和发生。
猪肉中神经分泌因子GPR56和alpha-actinin-3等基因对肌肉生长和发生贡献较大。
此外,一些转录因子如MyoD、Myf5和Pax7等也通过转录调控肌肉发育。
更进一步,基因组学研究还可以揭示个体之间的基因差异,从而基于这些差异预测个体的生产性能。
通过这种方式,可以更好地选择合适的家畜和育种计划以提高生产效益。
转录组学分析转录组学是研究转录组的基因表达、RNA种类和数量在不同条件下的变化及其调节机制的科学。
转录组包括在细胞特定条件下表达的所有基因。
肌肉细胞特异性基因转录调控因子如Mef2和MyoD等在肌肉细胞发育和功能dai'ta了很大作用。
Mef2是一个能够诱导肌肉细胞分化的关键转录因子。
它在早期肌肉细胞与成熟肌肉细胞中都有表达,扮演了不同的角色。
同时,MyoD可以促进肌肉细胞分化并寄共同调控成为成熟肌肉细胞所必需的蛋白质的基因。
一些WNT 通路中的基因也是特异地表达于肌肉细胞中。
通过转录组学分析,我们可以获得关于哪些基因在何种程度上表达以及在不同生长时期和条件下该表达是如何变化的信息。
这有助于揭示肌肉发育和肉质特性的分子机制并为改进家畜品质提供信息。
一项对山羊长期饲养的转录组学研究对肉品品质的研究发现,长期饲养在基因表达水平上显著影响了长饲肉羊的肉品质(如肌红蛋白含量和PH),这提示我们应该注意这些长期饲养对家畜基因调控的影响。
动物肌肉生长调控因子基因和猪脂肪沉积调控基因的研究

动物肌肉生长调控因子基因和猪脂肪沉积调控基因的研究动物肌肉生长调控因子基因和猪脂肪沉积调控基因的研究随着人们对肉类品质和营养的要求越来越高,对动物肌肉生长调控因子基因和猪脂肪沉积调控基因的研究也变得越来越重要。
这些基因对于提高肌肉质量和减少脂肪积累至关重要,因此相关研究有助于改善人类食品供应和健康状况。
动物肌肉生长调控因子基因是影响肌肉发育和质量的关键因素。
其中,MyoD家族基因是一组在肌肉分化和生长中起重要作用的基因。
研究表明,MyoD基因的表达水平与肌肉生长速度和肌纤维数量密切相关。
此外,IGF基因家族也被发现与动物肌肉生长密切相关。
IGF-1是调控肌肉生长的重要因子,它作为一种促进蛋白质合成和细胞增殖的生长因子,能够增强肌肉细胞的增殖和分化能力。
研究人员通过基因编辑技术对这些调控基因进行研究,以探索它们如何影响动物的肌肉质量和生长速度。
许多研究利用转基因技术,将MyoD和IGF基因等有关肌肉生长的基因导入到动物的基因组中,以增强它们的肌肉质量。
另一方面,猪脂肪沉积调控基因的研究对于改善猪肉质量和减少脂肪积累也具有重要意义。
研究表明,猪脂肪沉积主要受到脂肪合成相关基因的调控。
例如,猪的FASN基因编码酸敏感脂肪酸合酶,该酶参与脂肪酸生物合成的关键步骤。
同时,研究还发现,CD36和LPL等基因在猪脂肪沉积中也起着重要的作用。
除基因调控外,环境因素如饮食和运动等也会对动物肌肉生长和脂肪沉积产生影响。
其中饮食成分特别重要,营养成分的摄取量和比例的不同都会影响动物肌肉和脂肪的沉积。
因此,合理的饲养管理和营养调整可以改善肉类品质。
综上所述,动物肌肉生长调控因子基因和猪脂肪沉积调控基因的研究对于改善肉类质量、提高肌肉生长速度、减少脂肪积累至关重要。
通过对这些基因调控机制的深入研究,人们可以提高动物的肌肉质量和生产效益,进一步改善人类的食品供应和健康状况。
未来的研究将集中在基因编辑技术和环境调控策略的应用,以优化猪肉质量和减少脂肪沉积,进一步满足人们对优质食品的需求综合上述研究,肉质量和生长速度是决定动物肌肉生长和脂肪沉积的重要因素。
猪crtc3基因表达的品种差异及forskolin对其表达的影响

动物营养学报2020,32(2):870⁃880ChineseJournalofAnimalNutrition㊀doi:10.3969/j.issn.1006⁃267x.2020.02.042猪CRTC3基因表达的品种差异及forskolin对其表达的影响刘嘉琪㊀刘佳丽㊀农秋雲㊀单体中∗(浙江大学动物科学学院,动物分子营养学教育部重点实验室,杭州310058)摘㊀要:本试验旨在探究糖脂代谢通路关键基因CRTC3在不同品种猪肌肉和脂肪组织中的表达情况,并通过forskolin处理猪皮下脂肪前体细胞,研究forskolin对脂肪前体细胞分化聚酯和CRTC3基因表达的影响,阐明猪CRTC3基因表达与脂肪沉积的关系㊂试验选取杜长大猪和莱芜猪各5头,检测肌肉㊁脂肪组织中CRTC3的mRNA和蛋白表达水平以及脂肪代谢相关基因的mRNA表达水平;选取2头3日龄的杜长大仔猪,分离猪皮下脂肪前体细胞,待完全融合后用MDI诱导培养基诱导4d,然后用分化培养基继续诱导4d,完成诱导分化㊂Forskolin组在诱导分化的第1天即加入forskolin,使其终浓度为10μmol/L,对照组则加入同浓度的二甲基亚砜(DMSO)进行诱导分化㊂结果表明:在莱芜猪的背最长肌和腰大肌中,CRTC3的蛋白表达水平高于杜长大猪;在莱芜猪的皮下和内脏脂肪组织中,CRTC3及脂肪沉积相关基因过氧化物酶体增殖剂激活受体γ(PPARγ)㊁脂肪酸结合蛋白4(FABP4)㊁CCAAT/增强子结合蛋白α(C/EBPα)㊁围脂滴蛋白(PLIN)和瘦素(LEP)的mRNA表达水平显著或极显著高于杜长大猪(P<0.05或P<0.01),而脂肪棕色化相关基因NF-E2相关因子1(NRF1)㊁过氧化物酶体增殖物激活受体-γ共激活因子-1α(PGC⁃1α)㊁PRDM16㊁解偶联蛋白2(UCP2)㊁解偶联蛋白3(UCP3)的mRNA表达水平则显著或极显著低于杜长大猪(P<0.05或P<0.01)㊂进一步的研究发现,猪皮下脂肪前体细胞分化后CRTC3和脂肪沉积相关基因的mRNA表达水平极显著提高(P<0.01),脂肪棕色化相关基因的mRNA表达水平也均极显著升高(P<0.01)㊂10μmol/Lforsko⁃lin处理能抑制猪皮下脂肪前体细胞分化,极显著升高环磷腺苷效应元件结合蛋白(CREB)和脂肪棕色化相关基因的mRNA表达水平(P<0.01),促进CRTC3的进核,极显著降低CRTC3和脂肪沉积相关基因的mRNA表达水平(P<0.01)㊂上述研究结果表明,CRTC3基因与猪脂肪沉积密切相关,forskolin处理可以调控猪CRTC3及脂质代谢相关基因表达,调控猪皮下脂肪前体细胞分化聚酯㊂关键词:猪;CRTC3;脂肪沉积;分化聚酯;forskolin中图分类号:S828㊀㊀㊀㊀文献标识码:A㊀㊀㊀㊀文章编号:1006⁃267X(2020)02⁃0870⁃11收稿日期:2019-08-21基金项目:国家自然科学基金项目(31722053)作者简介:刘嘉琪(1994 ),女,广西桂林人,博士研究生,动物营养与饲料科学专业㊂E⁃mail:liu⁃jiaqi@zju.edu.cn∗通信作者:单体中,研究员,博士生导师,E⁃mail:tzshan@zju.edu.cn㊀㊀环磷酸腺苷(cAMP)-蛋白激酶A(PKA)信号通路是调控机体葡萄糖和脂质代谢的关键信号通路之一㊂研究发现,cAMP⁃PKA介导的信号通路通过cAMP反应元件结合蛋白(CREB)及其转录激活因子CRTC共同介导调控下游靶基因的转录和翻译[1]㊂CRTC3是CRTC家族中的一员,在脂肪组织中高度表达,参与了脂肪组织的发育㊁分化的过程[2]㊂但目前关于猪CRTC3的研究报道2期刘嘉琪等:猪CRTC3基因表达的品种差异及forskolin对其表达的影响较少,主要集中在其全长基因克隆和遗传序列表征上[3-4],关于其在猪脂质代谢中的功能还未见报道㊂此外,还有很多脂肪沉积相关基因参与调控脂肪代谢,如CCAAT/增强子结合蛋白α(C/EBPα)和过氧化物酶体增殖体激活受体γ(PPARγ)基因调控脂肪细胞分化聚酯[5],固醇调节元件结合蛋白-1(SREBP⁃1)基因调控胆固醇生物合成和脂肪酸合成[6-7];PPARγ基因还调控脂肪酸的合成[8],参与脂肪酸结合蛋白4(FABP4)转运,促进脂肪酸合成,进而增加机体的脂肪储存量[9]㊂过表达脂联素(ADIPOQ)可以促进游离脂肪酸的氧化和葡萄糖的吸收[10],瘦素(leptin,LEP)表达量升高可消耗脂肪,抑制脂滴生成[11]㊂脂滴是脂质储存的重要调节剂,围脂滴蛋白(peril⁃ipin,PLIN)包裹在成熟脂肪细胞的脂滴周围,维持脂质代谢的动态平衡[12]㊂㊀㊀在哺乳动物中有3种脂肪细胞:白色㊁棕色和米色,其中米色脂肪细胞常见于棕色化的白色脂肪组织中㊂棕色脂肪细胞和米色脂肪细胞含有大量的线粒体且表达较高的解偶联蛋白1(UCP1)㊂目前的研究结果表明,猪体内缺乏功能性的棕色脂肪组织和UCP1基因[13-14],但冷刺激可以调控猪体内米色脂肪细胞生成和解偶联蛋白3(UCP3)等棕色化相关基因表达[13]㊂线粒体解偶联蛋白2(UCP2)和UCP3位于线粒体内膜,能促进机体产热消耗能量,从而加快糖脂消耗,减少脂肪沉积量[15]㊂白色脂肪组织棕色化是一种重要的脂质代谢调控机制,可降低血浆甘油三酯和胆固醇水平,影响脂肪沉积[16]㊂Forskolin是一种cAMP⁃PKA信号通路激活剂,能促进白色脂肪组织棕色化[17]㊂白色脂肪组织棕色化相关基因过氧化物酶体增殖物激活受体-γ共激活因子-1α(PGC⁃1α)可与CREB和核呼吸因子(NRFs)相互作用并调节二者的活性[18];PRDM16(PRD1⁃BF1⁃RIZ1homologousdomaincontaining16)可控制棕色脂肪细胞的发育,调控棕色脂肪组织和白色脂肪组织与骨骼肌肌肉组织的形成及相互转化,进而影响脂肪和肌肉细胞的形成和发育[19]㊂但是目前关于猪白色脂肪组织棕色化相关基因表达的品种差异及forsko⁃lin对它们的影响还鲜见报道㊂㊀㊀因此,本试验以杜长大猪和我国著名的肉脂型地方品种猪莱芜猪为动物模型,利用Westernblot㊁实时荧光定量PCR和免疫荧光染色等技术,研究CRTC3及脂质代谢相关基因在不同品种猪脂肪和肌肉组织中的表达差异及其与脂肪沉积之间的关系,并进一步通过细胞试验探究forskolin处理对猪皮下脂肪前体细胞分化聚酯和CRTC3及脂质代谢相关基因表达的影响㊂1㊀材料与方法1.1㊀试验动物㊀㊀选取体重在(110.0ʃ2.5)kg的莱芜猪和杜长大猪各5头,屠宰取背最长肌㊁腰大肌㊁皮下脂肪㊁腹部脂肪组织样品,液氮速冻后保存于-80ħ冰箱㊂选取2头3日龄的杜长大仔猪,用于分离皮下脂肪组织中的前体脂肪细胞㊂1.2㊀试验方法1.2.1㊀Westernblot㊀㊀用细胞裂解液提取组织和细胞中的总蛋白质,通过二喹啉甲酸(BCA)试剂盒测定每个样品的蛋白质浓度,以确保每个蛋白质样品上样量相同㊂配制分离胶浓度为10%的十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS⁃PAGE)凝胶,进行SDS⁃PAGE分离,然后将蛋白质印迹转移至硝酸纤维素膜上㊂使用5%脱脂牛奶室温封闭1h,4ħ一抗孵育过夜;第2天用TBST洗膜3次,每次5min,然后二抗室温孵育1h后,洗膜曝光㊂1.2.2㊀实时荧光定量PCR㊀㊀使用Trizol法提取组织和细胞中的总RNA,之后用核酸分析仪检测样品RNA浓度,选取A260nm/A280nm值为1.8 2.0的RNA作为模板,按照反转录试剂盒(PrimeScriptTMRTReagentKit)的操作说明,合成cDNA㊂使用Primer6.0设计引物,引物序列见表1㊂内参基因为18SrDNA㊂然后按照实时荧光定量PCR试剂盒进行目的基因mRNA表达水平检测㊂1.2.3㊀猪皮下脂肪前体细胞分离培养㊀㊀取3日龄杜长大猪皮下脂肪组织,剪碎后用1%胶原酶Ⅰ消化液37ħ水浴消化60min,过滤,离心,弃上清,沉淀用DMEM培养基漂洗后过滤,用含10%小牛血清的DMEM完全培养基(DMEM+10%胎牛血清+青霉素-链霉素)重悬,接种于培养瓶,采用差速贴壁法纯化细胞,获得猪皮下脂肪前体细胞,将猪皮下脂肪前体细胞放置在37ħ㊁5%CO2培养箱中培养,每隔1d换1次培养基㊂178㊀动㊀物㊀营㊀养㊀学㊀报32卷表1 引物序列Table1㊀Primersequences基因Genes引物序列Primersequence(5ᶄ 3ᶄ)产物大小Productlength/bp18SrDNAF:CCCACGGAATCGAGAAAGAGR:TTGACGGAAGGGCACCA122CRTC3F:GACAAGCCAGGACGACAGTTR:GGTGCTTCTCTTCCTTCCAA112PPARγF:TGTGGACCTGTCGGTGATGR:TGGAGTGGAAATGCTGGAGA88FABP4F:GGAAGGTGGCTGGCATGGCR:CTCCATCTAAGGTTATGGTGCTCTTG177C/EBPαF:GGTGGACAAGAACAGCAACGR:TCACTGGTCAGCTCCAGCAC129PLINF:CCCTGGTGGCGTCTGTATGR:GCGGCATATTCAGCAGTGTC349LEPF:AGATCCTCACCAGTCTGCCTTCCR:CCAGGCTCTCCAAGGTCTCCAG141NRF1F:GTGGCCACCTACACTGAACAR:CCAGATGGGCTGTTACCTCA130PGC⁃1αF:GAGATTCCGTATCACCACCR:CTTTCAGACTCCCGCTTC340PRDM16F:TACACGTGCAGGTACTGTGGR:GAGGTGTCTGTCCAGGTTGG231SREBP⁃1F:AAGCGGACGGCTCACAATGR:CGCAAGACGGCGGATTTAT122ADIPOQF:ACGGTCTACTTGAAGGATGTGAR:TCCAGATAGAGGAGCACAGAG125UCP2F:ATGGTTGGATTCAAGGCCACR:CCTTTCTCCCTGGATCTGC141UCP3F:CAACAGGAAGTACAGCGGGAR:CACCATCTCGGCACAGTTCA130㊀㊀PPARγ:过氧化物酶体增殖体激活受体γperoxisomeproliferatorsactivatedreceptorγ;FABP4:脂肪酸结合蛋白4fattyacidbindingprotein4;C/EBPα:CCAAT/增强子结合蛋白αCCAAT/enhancerbindingproteinα;PLIN:围脂滴蛋白perilipin;LEP:瘦素leptin;NRF1:NF⁃E2相关因子1NF⁃E2relatedfactor1;PGC⁃1α:过氧化物酶体增殖物激活受体-γ共激活因子-1αPPARγcoactivator1α;SREBP⁃1:固醇调节元件结合蛋白-1sterolregulatoryelement⁃bindingprotein⁃1;ADIPOQ:脂联素adiponectin;UCP2:解偶联蛋白2uncouplingprotein2;UCP3:解偶联蛋白3uncouplingprotein3㊂下图同Thesameasbelow㊂1.2.4㊀猪皮下脂肪前体细胞诱导分化和forskolin处理㊀㊀将培养的猪皮下脂肪前体细胞接种于12孔细胞培养板,待完全融合后用MDI成脂诱导培养基[DMEM完全培养基+0.25mmol/L磷酸二酯酶抑制剂(IBMX)+100nmol/L地塞米松(DEX)+500nmol/L胰岛素(insulin)+100nmol/L罗格列酮(ROS)]诱导4d,然后用分化培养基(DMEM完全培养基+500nmol/Linsulin)继续诱导4d,完成诱导分化㊂Forskolin组在诱导分化的第1天即加入forskolin,使其终浓度为10μmol/L,对照组则加入同浓度的二甲基亚砜(DMSO)进行诱导分化㊂1.2.5㊀免疫荧光染色㊀㊀细胞用磷酸盐缓冲溶液(PBS)洗涤3次后进行免疫荧光染色,具体步骤参见文献[4]㊂1.2.6㊀油红O染色㊀㊀称取0.5g油红O溶于100mL异丙醇中配好保存,使用时将其同蒸馏水按3ʒ2稀释为工作液,通过定性滤纸过滤㊂将细胞培养板的培养液吸出,加入固定液15min,弃掉固定液,加入油红O2782期刘嘉琪等:猪CRTC3基因表达的品种差异及forskolin对其表达的影响工作液染色15min,然后用PBS洗涤后拍照观察㊂1.3㊀统计分析㊀㊀实时荧光定量PCR数据结果用 平均值ʃ标准误(meanʃSE) 表示㊂采用Graphpad软件进行显著性分析和作图,P<0.01表示差异极显著,P<0.05表示差异显著,P>0.05表示差异不显著㊂2㊀结㊀果2.1㊀CRTC3在不同品种猪肌肉组织中的表达差异㊀㊀Westernblot结果表明,莱芜猪背最长肌和腰大肌中CRTC3的蛋白表达水平均高于杜长大猪(图1-A㊁图1-B)㊂同时,莱芜猪背最长肌中CRTC3的mRNA表达水平显著高于杜长大猪(P<0.05,图1-C),而在腰大肌中2个品种猪之间无显著差异(P>0.05,图1-D)㊂这表明在肌肉组织中,猪CRTC3的表达在不同品种猪间存在差异㊂2.2㊀CRTC3及脂质代谢相关基因在不同品种猪皮下脂肪组织中的表达差异㊀㊀Westernblot结果表明,莱芜猪皮下脂肪组织中CRTC3的蛋白表达水平高于杜长大猪(图2-A),同时,莱芜猪皮下脂肪组织中CRTC3的mRNA表达水平显著高于杜长大猪(P<0.05,图2-B)㊂此外,莱芜猪皮下脂肪组织中脂质沉积相关基因如PPARγ㊁FABP4㊁C/EBPα和PLIN的mRNA表达水平显著高于杜长大猪(P<0.05,图2-C),LEP的mRNA表达水平极显著高于杜长大猪(P<0.01,图2-C);莱芜猪皮下脂肪组织中脂肪棕色化相关基因如PRDM16㊁UCP2和UCP3的mRNA表达水平显著低于杜长大猪(P<0.05,图2-D),NRF1和PGC⁃1α的mRNA表达水平极显著低于杜长大猪(P<0.01,图2-D)㊂㊀㊀A:杜长大猪和莱芜猪背最长肌中CRTC3的蛋白表达水平(1㊁3为杜长大猪,2㊁4为莱芜猪);B:杜长大猪和莱芜猪腰大肌中CRTC3的蛋白表达水平(1㊁3为杜长大猪,2㊁4为莱芜猪);C:杜长大猪和莱芜猪背最长肌中CRTC3的mRNA表达水平(n=5);D:杜长大猪和莱芜猪腰大肌中CRTC3的mRNA表达水平(n=5)㊂∗:差异显著(P<0.05)㊂㊀㊀A:theproteinexpressionlevelofCRTC3inlongissimusdorsimuscle(LDM)ofDLYpigs(1,3)andLaiwupigs(2,4);B:theproteinexpressionlevelofCRTC3inpsoasmajor(PMM)muscleofDLYpigs(1,3)andLaiwupigs(2,4);C:themRNAexpressionlevelofCRTC3inLDMofDLYpigsandLaiwupigs(n=5);D:themRNAexpressionlevelofCRTC3inPMMofDLYpigsandLaiwupigs(n=5).∗:significantdifference(P<0.05).图1㊀猪肌肉组织中CRTC3表达的品种差异Fig.1㊀BreeddifferenceofCRTC3expressioninmuscletissueofpigs2.3㊀CRTC3及脂质代谢相关基因在不同品种猪内脏脂肪组织中的表达差异㊀㊀与杜长大猪相比,莱芜猪内脏脂肪组织中CRTC3的蛋白表达水平更高(图3-A),CRTC3的mRNA表达水平显著高于杜长大猪(P<0.05,图3-B)㊂莱芜猪内脏脂肪组织中PPARγ㊁LEP的mRNA表达水平极显著高于杜长大猪(P<0.01,图3-C),FABP4㊁C/EBPα和PLIN的mRNA表达水378㊀动㊀物㊀营㊀养㊀学㊀报32卷平均显著高于杜长大猪(P<0.05,图3-C);而莱芜猪内脏脂肪组织中NRF1㊁PGC⁃1α㊁PRDM16和UCP3的mRNA表达水平均显著低于杜长大猪(P<0.0,图3-D),UCP2的mRNA表达水平极显著低于杜长大猪(P<0.01,图3-D)㊂㊀㊀A:杜长大猪和莱芜猪皮下脂肪组织中CRTC3的蛋白表达水平(1㊁3为杜长大猪,2㊁4为莱芜猪);B:杜长大猪和莱芜猪皮下脂肪组织中CRTC3的mRNA表达水平(n=5);C:杜长大猪和莱芜猪皮下脂肪组织中脂肪沉积相关基因的mRNA表达水平(n=5);D:杜长大猪和莱芜猪皮下脂肪组织中脂肪棕色化相关基因的mRNA表达水平(n=5)㊂∗:差异显著(P<0.05);∗∗:差异极显著(P<0.01)㊂㊀㊀A:theproteinexpressionlevelofCRTC3insubcutaneousadiposetissue(SAT)ofDLYpigs(1,3)andLaiwupigs(2,4);B:themRNAexpressionlevelofCRTC3inSATofDLYpigsandLaiwupigs(n=5);C:themRNAexpressionlevelsofadiposedepositionrelatedgenesinSATofDLYpigsandLaiwupigs(n=5);D:themRNAexpressionlevelsofadiposebrown⁃ingrelatedgenesinSATofDLYpigsandLaiwupigs(n=5).∗:significantdifference(P<0.05);∗∗:extremelysignificantdifference(P<0.01).图2㊀猪皮下脂肪组织中CRTC3及脂质代谢相关基因表达的品种差异Fig.2㊀BreeddifferenceofexpressionofCRTC3andlipidmetabolismrelatedgenesinsubcutaneousadiposetissue2.4㊀猪皮下脂肪前体细胞分化前后CRTC3及脂质代谢相关基因的表达变化㊀㊀由分化前后的照片发现,猪皮下脂肪前体细胞分化后聚集了大量的脂滴(图4-A),分化后CRTC3和磷酸化CREB(p⁃CREB)的蛋白表达水平有明显提高(图4-B)㊂与分化前相比,猪皮下脂肪前体细胞分化后CRTC3和CREB的mRNA表达水平极显著升高(P<0.01,图4-C㊁图4-D)㊂此外,分化后的脂肪前体细胞中PPARγ㊁FABP4㊁C/EBPα㊁PLIN㊁SREBP⁃1㊁ADIPOQ和LEP的mRNA表达水平极显著高于分化前的脂肪前体细胞(P<0.01,图4-E),同时NRF1㊁PGC⁃1α㊁PRDM16㊁UCP2和UCP3的mRNA表达水平在分化后也极显著升高(P<0.01,图4-F)㊂2.5Forskolin处理对猪CRTC3定位、表达和皮下脂肪前体细胞分化的影响㊀㊀通过免疫荧光染色发现,在一般情况下,CRTC3定位在293T细胞的细胞质和细胞核内,forskolin处理促进了猪皮下脂肪前体细胞中CRTC3蛋白由细胞质进入细胞核内(图5-A)㊂由图5-B的油红O染色可见,forskolin处理后脂滴较对照组明显减少,且猪皮下脂肪前体细胞中CREB的mRNA表达水平极显著高于对照组(P<0.01,图5-C),同时PGC⁃1α㊁PRDM16㊁UCP2和UCP3的mRNA表达水平极显著高于对照组(P<0.01),UCP3的mRNA表达水平显著高于对照组4782期刘嘉琪等:猪CRTC3基因表达的品种差异及forskolin对其表达的影响(P<0.05),NRF1的mRNA表达水平与对照组相比有升高的趋势但差异不显著(P>0.05,图5-D)㊂而Forskolin组猪皮下脂肪前体细胞中CRTC3在蛋白和mRNA表达水平上均较对照组极显著降低(P<0.01,图5-E㊁图5-F),同时PPARγ㊁FABP4㊁C/EBPα㊁PLIN和LEP的mRNA表达水平也极显著低于对照组(P<0.01,图5-G)㊂㊀㊀A:杜长大猪和莱芜猪内脏脂肪组织中CRTC3的蛋白表达水平(1㊁3为杜长大猪,2㊁4为莱芜猪);B:杜长大猪和莱芜猪内脏脂肪组织中CRTC3的mRNA表达水平(n=5);C:杜长大猪和莱芜猪内脏脂肪组织中脂肪沉积相关基因的mRNA表达水平(n=5);D:杜长大猪和莱芜猪内脏脂肪组织中脂肪棕色化相关基因的mRNA表达水平(n=5)㊂∗:差异显著(P<0.05);∗∗:差异极显著(P<0.01)㊂㊀㊀A:theproteinexpressionlevelofCRTC3invisceraladiposetissue(VAT)ofDLYpigs(1,3)andLaiwupigs(2,4);B:themRNAexpressionlevelofCRTC3inVATofDLYpigsandLaiwupigs(n=5);C:themRNAexpressionlevelsofadiposedepositionrelatedgenesinVATofDLYpigsandLaiwupigs(n=5);D:themRNAexpressionlevelsofadiposebrowningrelatedgenesinVATofDLYpigsandLaiwupigs(n=5).∗:significantdifference(P<0.05);∗∗:extremelysignificantdifference(P<0.01).图3㊀猪内脏脂肪组织中CRTC3及脂质代谢相关基因表达的品种差异Fig.3㊀BreeddifferenceofexpressionofCRTC3andlipidmetabolismrelatedgenesinvisceraladiposetissueofpigs3㊀讨㊀论3.1㊀CRTC3基因表达的品种差异㊀㊀脂肪沉积的关键信号网络需要机体糖脂代谢相关基因的参与,CRTC3是一个重要的糖脂代谢基因㊂大量研究表明CRTC3可能通过与糖脂代谢相关基因的互作,调节激素的分泌㊁能量代谢和脂肪酸代谢反应,进而影响机体肌肉和脂肪组织的脂肪沉积[2]㊂全身敲除CRTC3基因后,小鼠体内棕色脂肪组织量增加,能量消耗量更大,说明CRTC3基因的缺失促进了白色脂肪组织的脂解作用;在高脂饮食条件下,全身敲除CRTC3基因小鼠较野生型小鼠明显瘦小,研究者发现这是由于CRTC3受到脂肪组织中的β-肾上腺素能受体信号的调控,从而促进脂肪沉积[1]㊂而关于CRTC3在脂肪和肌肉中的表达模式及功能的研究甚少㊂本试验比较了杜长大猪和地方品种猪莱芜猪间背最长肌和腰大肌CRTC3基因的表达差异,发现CRTC3基因在莱芜猪中的表达水平更高;通过比较皮下脂肪和内脏脂肪组织中CRTC3的mRNA表达水平发现,莱芜猪的CRTC3基因表达水平均高于杜长大猪,同时发现,莱芜猪的PPARγ㊁FABP4㊁C/EBPα㊁PLIN㊁LEP等脂肪沉积相关基因的mRNA表达水平显著或极显著高于杜长大猪,而莱芜猪中NRF1㊁PGC⁃1α㊁PRDM16㊁UCP2㊁UCP3等白色脂肪棕色化相关基因的mRNA表达水平显著或极显著低于杜长大猪,表明品种间脂质代谢和米色脂肪细胞数目可能存在差别㊂有研578㊀动㊀物㊀营㊀养㊀学㊀报32卷究表明,猪体内虽然没有功能性的UCP1基因和棕色脂肪[13-14],但存在米色脂肪细胞[13],当猪受到冷刺激时可以通过调控米色脂肪细胞生成和UCP3等相关基因表达调控脂质代谢和产热[13]㊂也有研究表明,CRTC家族中的CRTC2能够促进PGC⁃1α和线粒体基因在肌肉细胞中的表达[20],进而推测CRTC家族可能会调控白色脂肪棕色化及相关基因表达㊂而关于CRTC3是否可以调控猪皮下脂肪组织中棕色化相关基因表达还有待于进一步研究㊂细胞试验研究还发现,分化后的皮下脂肪前体细胞生成大量的脂滴,检测到CRTC3和聚酯相关基因PPARγ㊁FABP4㊁C/EBPα㊁PLIN和LEP的mRNA表达水平在分化后得到极显著升高,同时脂肪棕色化特异性基因NRF1㊁PGC⁃1α㊁PRDM16㊁UCP2㊁UCP3的mRNA表达水平也极显著升高㊂这些结果表明,猪CRTC3基因表达与猪脂肪沉积及相关基因表达可能存在密切的关系,其表达水平可能影响猪的脂质代谢和脂肪沉积,但其具体功能还有待于进一步研究㊂㊀㊀A:猪皮下脂肪前体细胞分化前后的照片;B:猪皮下脂肪前体细胞分化前后CRTC3㊁pCREB㊁CREB的蛋白表达水平(1㊁3:分化前;2㊁4:分化后);C:猪皮下脂肪前体细胞分化前后CRTC3的mRNA表达水平(n=4);D:猪皮下脂肪前体细胞分化前后CREB的mRNA表达水平(n=4);E:猪皮下脂肪前体细胞分化前后脂肪沉积相关基因的mRNA表达水平(n=4);F:猪皮下脂肪前体细胞分化前后脂肪棕色化相关基因的mRNA表达水平(n=4)㊂∗∗:差异极显著(P<0.01)㊂㊀㊀A:theimageofporcinesubcutaneouspreadipocytesbeforeandafterdifferentiation;B:theproteinexpressionlevelof⁃CRTC3,pCREBandCREBinporcinesubcutaneouspreadipocytesbefore(1,3)andafterdifferentiation(2,4)(n=4);C:themRNAexpressionlevelofCRTC3inporcinesubcutaneouspreadipocytesbeforeandafterdifferentiation(n=4);D:themRNAexpressionlevelofCREBinporcinesubcutaneouspreadipocytesbeforeandafterdifferentiation(n=4);E:themRNAexpressionlevelsofadiposedepositionrelatedgenesinporcinesubcutaneouspreadipocytesbeforeandafterdifferentiation(n=4);F:theex⁃pressionmRNAlevelsofadiposebrowningrelatedgenesinporcinesubcutaneouspreadipocytesbeforeandafterdifferentiation(n=4).∗∗:extremelysignificantdifference(P<0.01).图4㊀猪皮下脂肪前体细胞分化前后CRTC3及脂质代谢相关基因的表达变化Fig.4㊀ExpressionchangesofCRTC3andlipidmetabolismrelatedgenesinporcinesubcutaneouspreadipocytesbeforeorafterdifferentiation6782期刘嘉琪等:猪CRTC3基因表达的品种差异及forskolin对其表达的影响㊀㊀A:经(Forskolin组)和不经forskolin处理(对照组)时293T细胞中CRTC3定位的免疫荧光图像;B:经和不经forskolin处理时猪皮下脂肪前体细胞分化后的油红O染色图片;C:经和不经forskolin处理时猪皮下脂肪前体细胞中CREB的mRNA表达水平(n=4);D:经和不经forskolin处理时猪皮下脂肪前体细胞中脂肪棕色化相关基因的mRNA表达水平(n=4);E:经和不经forskolin处理时猪皮下脂肪前体细胞中CRTC3的蛋白表达水平(1㊁3:对照组;2㊁4:Forskolin组);F:经和不经forskolin处理时猪皮下脂肪前体细胞中CRTC3的mRNA表达水平(n=4);G:经和不经forskolin处理时猪皮下脂肪前体细胞中脂肪沉积相关基因的mRNA表达水平(n=4)㊂∗:差异显著(P<0.05);∗∗:差异极显著(P<0.01)㊂㊀㊀A:theimmunofluorescenceimagesoflocalizationofCRTC3in293Tcellswith(Forskolingroup)orwithoutforskolintreat⁃ment(controlgroup);B:theoil⁃redOstainingmapsofporcinesubcutaneouspreadipocyteswithorwithoutforskolintreatment;C:themRNAexpressionlevelofCREBinporcinesubcutaneouspreadipocyteswithorwithoutforskolintreatment(n=4);D:themRNAexpressionlevelsofadiposebrowningrelatedgenesinporcinesubcutaneouspreadipocyteswithorwithoutforskolintreatment(n=4);E:themRNAexpressionlevelofCRTC3inporcinesubcutaneouspreadipocyteswith(2,4)orwithout(1,3)forskolintreatment(n=4);F:theproteinexpressionlevelofCRTC3inporcinesubcutaneouspreadipocyteswithorwithoutforskolintreatment(n=4);G:themRNAexpressionlevelsofadiposedepositionrelatedgenesinporcinesubcutaneouspreadipo⁃cyteswithorwithoutforskolintreatment.∗:significantdifference(P<0.05);∗∗:extremelysignificantdifference(P<0.01).图5㊀Forskolin处理对猪CRTC3定位、表达和皮下脂肪前体细胞分化的影响Fig.5㊀EffectsofforskolintreatmentonlocalizationandexpressionofporcineCRTC3anddifferentiationofsubcutaneouspreadipocytes3.2㊀Forskolin调控猪皮下脂肪前体细胞分化聚酯和CRTC3等基因表达㊀㊀Forskolin是一种腺苷酸环化酶的激活剂,可以提高cAMP水平,进而调控cAMP⁃PKA信号通路㊂通过PKA和细胞外信号调节激酶(ERK)信号通路协同增加胆囊收缩素(CCK)基因转录,激活CREB促使其磷酸化,促进CRTC3的进核[21]㊂我们前期的研究结果表明,CRTC3的亚细胞定位与糖脂代谢和脂肪沉积密切相关[2,13,22]㊂CREB可通过不同的识别位点激活启动子,并响应cAMP778㊀动㊀物㊀营㊀养㊀学㊀报32卷和血清介导细胞色素c转录,从而影响细胞线粒体的呼吸作用,进而促进脂质代谢[18]㊂本试验中,在猪皮下脂肪前体细胞诱导分化的过程中加入forskolin,油红O染色发现forskolin处理显著抑制了脂滴的形成,CREB的mRNA表达水平显著升高㊂有研究发现,CREB通过抑制PPARγ和诱导PGC⁃1α协调空腹时肝脏脂质和葡萄糖代谢的能力,促进葡萄糖异生和脂肪酸氧化,从而促进糖脂的代谢,降低脂肪的沉积[23]㊂与该报道一致,本试验中,forskolin处理后猪皮下脂肪前体细胞中UCP2等线粒体相关基因的mRNA表达水平显著或极显著升高,而CRTC3和脂肪沉积相关基因的mRNA和蛋白表达水平极显著下降㊂这些结果表明,CRTC3在猪脂质代谢和脂肪沉积中发挥重要的调控作用㊂但是,关于CRTC3是如何调控猪脂质代谢和脂肪沉积的还有待于进一步研究㊂4㊀结㊀论㊀㊀CRTC3的mRNA和蛋白表达水平在地方品种猪莱芜猪的肌肉和脂肪组织中显著高于外来杂交品种猪杜长大猪;诱导猪皮下脂肪前体细胞分化可以促进CRTC3基因的表达,而forskolin处理能抑制猪皮下脂肪前体细胞分化聚酯和降低CRTC3等基因表达㊂这些结果表明,猪CRTC3可能在参与了脂质代谢的调控过程,并且在猪的脂质代谢和脂肪沉积过程中发挥重要的作用㊂参考文献:[1]㊀SONGY,ALTAREJOSJ,GOODARZIMO,etal.CRTC3linkscatecholaminesignallingtoenergybal⁃ance[J].Nature,2010,468(7326):933-939.[2]㊀LIUJQ,XUZY,WUWC,etal.RegulationroleofCRTC3inskeletalmuscleandadiposetissue[J].Jour⁃nalofCellularPhysiology,2018,233(2):818-821.[3]㊀LEESH,HURMH,LEEEA,etal.Genomiccharac⁃terizationoftheporcineCRTC3andtheeffectsofanon⁃synonymousmutationp.V515Fonleanmeatpro⁃ductionandbellyfat[J].MeatScience,2018,137:211-215.[4]㊀LIUJQ,YOUWJ,XUZY,etal.Rapidcommunica⁃tion:porcineCRTC3geneclone,expressionpattern,anditsregulatoryroleinintestinalepithelialcells[J].JournalofAnimalScience,2018,96(7):2622-2628.[5]㊀LEEJE,SCHMIDTH,LAIB,etal.Transcriptionalandepigenomicregulationofadipogenesis[J].Molec⁃ularandCellularBiology,2019,39(11):e00601-18.[6]㊀PARRAGAA,BELLSOLELLL,FERRÉ⁃D AMARÉAR,etal.Co⁃crystalstructureofsterolregulatoryele⁃mentbindingprotein1aat2.3åresolution[J].Struc⁃ture,1998,6(5):661-672.[7]㊀MENÉNDEZ⁃HURTADOA,VEGA⁃NUᶄÑEZE,SAN⁃TOSA,etal.Regulationbythyroidhormoneandretin⁃oicacidoftheCCAAT/enhancerbindingproteinαandβgenesduringliverdevelopment[J].BiochemicalandBiophysicalResearchCommunications,1997,234(3):605-610.[8]㊀BERGERJ,MOLLERDE.ThemechanismsofactionofPPARs[J].AnnualReviewofMedicine,2002,53:409-435.[9]㊀徐秋良,张庆莉,陈玉林.绵羊脂肪细胞型脂肪酸结合蛋白基因(FABP4)cDNA的克隆㊁表达及其结构模拟分析[J].农业生物技术学报,2011,19(3):483-489.[10]㊀HUE,LIANGP,SPIEGELMANBM.AdipoQisanoveladipose⁃specificgenedysregulatedinobesity[J].JournalofBiologicalChemistry,1996,271(18):10697-10703.[11]㊀李宏睿,孙文夏,潘杰.瘦素功能研究进展[J].中国动脉硬化杂志,2004,12(1):108-112.[12]㊀LONDOSC,SZTALRYDC,TANSEYJT,etal.RoleofPATproteinsinlipidmetabolism[J].Biochimie,2005,87(1):45-49.[13]㊀LINJ,CAOCW,TAOC,etal.ColdadaptationinpigsdependsonUCP3inbeigeadipocytes[J].JournalofMolecularCellBiology,2017,9(5):364-375.[14]㊀ZHENGQT,LINJ,HUANGJJ,etal.ReconstitutionofUCP1usingCRISPR/Cas9inthewhiteadiposetis⁃sueofpigsdecreasesfatdepositionandimprovesther⁃mogeniccapacity[J].ProceedingsoftheNationalA⁃cademyofSciencesoftheUnitedStatesofAmerica,2017,114(45):E9474-E9482.[15]㊀RICQUIERD,BOUILLAUDF.Theuncouplingpro⁃teinhomologues:UCP1,UCP2,UCP3,StUCPandAt⁃UCP[J].BiochemicalJournal,2000,345(2):161-179.[16]㊀姚旋,张颖,单仕芳,等.褐色脂肪组织研究的最新进展和科学意义[J].中国细胞生物学学报,2011,33(3):227-236.[17]㊀SEAMONKB,DALYJW.Forskolin:auniquediter⁃peneactivatorofcyclicAMP⁃generatingsystems[J].JournalofCyclicNucleotideResearch,1981,7(4):8782期刘嘉琪等:猪CRTC3基因表达的品种差异及forskolin对其表达的影响201-224.[18]㊀VERCAUTERENK,PASKORA,GLEYZERN,etal.PGC⁃1⁃relatedcoactivator:immediateearlyexpres⁃sionandcharacterizationofaCREB/NRF⁃1bindingdomainassociatedwithcytochromecpromoteroccu⁃pancyandrespiratorygrowth[J].MolecularandCel⁃lularBiology,2006,26(20):7409-7419.[19]㊀郭晓强,王晓红,边巍.PRDM16和棕色脂肪细胞分化[J].生命的化学,2009,29(1):29-32.[20]㊀WUZD,HUANGXM,FENGYJ,etal.TransducerofregulatedCREB⁃bindingproteins(TORCs)inducePGC⁃1αtranscriptionandmitochondrialbiogenesisinmusclecells[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2006,103(39):14379-14384.[21]㊀罗献梅,陈代文.棕色脂肪组织的生理功能及影响因素[J].饲料工业,2007,28(19):22-26.[22]㊀SHANTZ,XIONGY,ZHANGPP,etal.Lkb1con⁃trolsbrownadiposetissuegrowthandthermogenesisbyregulatingtheintracellularlocalizationofCRTC3[J].NatureCommunications,2016,7:12205.[23]㊀HERZIGS,HEDRICKS,MORANTTEI,etal.CREBcontrolshepaticlipidmetabolismthroughnuclearhor⁃monereceptorPPAR⁃γ[J].Nature,2003,426(6963):190-193.978㊀动㊀物㊀营㊀养㊀学㊀报32卷∗Correspondingauthor,professor,E⁃mail:tzshan@zju.edu.cn(责任编辑㊀菅景颖)BreedDifferenceofPorcineCRTC3ExpressionandItsRegulationbyForskolinLIUJiaqi㊀LIUJiali㊀NONGQiuyun㊀SHANTizhong∗(KeyLaboratoryofAnimalMolecularNutrition,MinistryofEducation,CollegeofAnimalScience,ZhejiangUniversity,Hangzhou310058,China)Abstract:CRTC3isakeygeneinglucoseandlipidmetabolismpathway.Inthisstudy,weaimedtostudytheexpressionpatternofporcineCRTC3inmuscleandadiposetissuesofdifferentbreeds,andtheeffectsoffors⁃kolintreatmentonexpressionofCRTC3geneanddifferentiationinporcinesubcutaneouspreadipocytes.FiveLaiwupigsandfiveDurocˑLandraceˑYorkshire(DLY)pigswereselectedtodetectthemRNAandproteinexpressionlevelsofCRTC3andlipidmetabolismrelatedgenesinmuscleandadiposetissues.Beforedifferenti⁃ation,theporcinesubcutaneouspreadipocyteswereseparatedfrom3⁃day⁃oldDLYpiglets.TheabovemediumwasreplacedbyDMEMcontainingMDIinduction4days.ThenthemediumwasreplacedbyDMEMcontai⁃ninginsulinuntilday8.10μmol/Lforskolinwasaddedatthe1stdayofdifferentiationandinductioninforsko⁃lingroup,andthedimethylsulfoxide(DMSO)withthesameconcentrationwasaddedincontrolgroup.Theresultsshowedasfollows:comparedwithDLYpigs,LaiwupighadahigherCRTC3proteinexpressionlevelinlongissimusdorsimuscleandpsoasmajormuscle.ThemRNAexpressionlevelsofCRTC3andfatdepositionrelatedgenesperoxisomeproliferatorsactivatedreceptorγ(PPARγ),fattyacidbindingprotein4(FABP4),CCAAT/enhancerbindingproteinα(C/EBPα),perilipin(PLIN)andleptin(LEP)insubcutaneousandvis⁃ceraladiposetissuesofLaiwupigsweresignificantlyhigherthanthoseofDLYpigs(P<0.05orP<0.01).However,themRNAexpressionlevelsofadiposebrowningrelatedgenesNF⁃E2relatedfactor1(NRF1),PPARγcoactivator1α(PGC⁃1α),PRD1⁃BF1⁃RIZ1homologousdomaincontaining16(PRDM16),uncoup⁃lingprotein2(UCP2)anduncouplingprotein3(UCP3)insubcutaneousandvisceraladiposetissuesofLaiwupigsweresignificantlyorextremelysignificantlylowerthanthoseofDLYpigs(P<0.05orP<0.01).Further⁃more,themRNAexpressionlevelsofCRTC3,adiposedepositionrelatedgenesandadiposebrowningrelatedgenesinporcinesubcutaneouspreadipocyteswereextremelysignificantlyincreasedafterdifferentiation(P<0.01).Treatmentwith10μmol/Lforskolininhibitedthedifferentiationofporcinesubcutaneouspreadipo⁃cytes,upregulatedthemRNAexpressionlevelsofcAMP⁃responseelementbindingprotein(CREB)andadi⁃posebrowningrelatedgenes,promotedtheentryofCRTC3intothenucleus.Moreover,itreducedthemRNAexpressionlevelsofCRTC3(P<0.01),aswellasadiposedepositionrelatedgenes(P<0.01).Therefore,CRTC3isaadiposedepositionrelatedgeneforselectingbioactiveornutrientfactors,whichshouldbeapplica⁃bletoregulationoffatdeposition.[ChineseJournalofAnimalNutrition,2020,32(2):870⁃880]Keywords:pigs;CRTC3;fatdeposition;differentiation;forskolin088。
贵州地方猪骨骼肌中3种 microRNAs 的表达差异

贵州地方猪骨骼肌中3种 microRNAs 的表达差异刘畅;罗志宇;冉雪琴;王嘉福【期刊名称】《贵州农业科学》【年(卷),期】2014(000)003【摘要】为探明 miR-1、miR-133、miR-206基因在不同品种猪肌肉组织中的表达及其与屠宰性能间的关系,以6月龄大白猪、糯谷猪、关岭猪及黔南黑猪的背最长肌为材料,采用实时荧光定量 PCR 方法,研究了3种 microRNAs (miRNAs)在贵州地方猪种和大白猪背最长肌中的表达差异。
结果表明:成年期miR-133的表达量高于另外2种 miRNA 分子。
miR-133在糯谷猪中的表达量是大白猪的1.58倍(P <0.05),与其眼肌面积、花油重高度负相关,与屠宰率高度正相关。
miR-206在糯谷猪中的表达量是大白猪的2.23倍(P <0.05),与其瘦肉率高度负相关,与板油重中度正相关。
在关岭猪背最长肌中 miR-133的表达量最高,是大白猪的2.67倍(P <0.05),miR-133和 miR-206的表达量与眼肌面积、胴体重高度正相关,与花油重、板油重和脂肪比负相关;但 miR-206的表达量明显低于其他猪种。
不同品种猪中 miR-133和 miR-206存在差异表达,对地方猪品种的骨骼肌生长和脂肪沉积有一定的调节作用。
【总页数】5页(P112-116)【作者】刘畅;罗志宇;冉雪琴;王嘉福【作者单位】贵州大学农业生物工程重点实验室,贵州贵阳 550025; 贵州大学动物科学学院,贵州贵阳 550025;贵州大学动物科学学院,贵州贵阳 550025;贵州大学动物科学学院,贵州贵阳 550025;贵州大学农业生物工程重点实验室,贵州贵阳 550025【正文语种】中文【中图分类】S813.3【相关文献】1.贵州地方山羊品种肌肉生长抑制素基因的表达差异 [J], 杨家大;彭舒2.贵州地方山羊CPT-1A基因在不同品种和组织中的表达差异 [J], 杨家大3.广西巴马小型猪2型糖尿病模型骨骼肌糖代谢及能量代谢相关基因的表达差异[J], 严雪瑜;蒋钦杨;吴延军;梁家充;郭亚芬;兰干球4.microRNA-155在长白猪和通城猪骨骼肌发育过程中的表达分析 [J], 赵拴平;昝林森;唐中林;李奎5.外泌体microRNA在猪成熟和闭锁卵泡中的表达差异及功能分析 [J], 陈慧芳;黄绮亮;胡智超;潘晓婷;吴志胜;白银山因版权原因,仅展示原文概要,查看原文内容请购买。
猪肌纤维发育变化及其与肉质相关性研究的开题报告

猪肌纤维发育变化及其与肉质相关性研究的开题报告
一、研究目的和意义
猪肉是我国主要的肉类食品之一,具有广泛的消费群体和市场需求。
猪肉的肉质是影响消费者购买和食用的主要因素之一,而猪肌纤维对肉质的影响极其重要。
因此,本研究旨在探究猪肌纤维发育变化及其与肉质相关性,为猪肉肉质评价和养殖提供科
学依据。
二、研究内容和方法
1. 研究目标:采用猪肌肉样本,分析猪肌纤维发育变化规律,探究其与肉质相关性。
2. 研究方法:选取养殖期间、品种不同的猪为研究对象,采用光学显微镜、电子显微镜等技术手段,对猪肌纤维进行形态学和生长发育方面的观察和分析。
同时,通
过肉质指标测定和实验数据分析,探究猪肌纤维发育变化与肉质品质之间的关系。
3. 研究内容:本研究主要围绕以下几个方面开展深入研究:(1)猪肌纤维发育
变化规律;(2)肉质指标的测定及与猪肌纤维发育变化之间的相关性;(3)探讨影
响猪肌纤维发育的内外因素和调控机制。
三、研究预期结果和应用价值
1. 研究预期结果:通过对猪肌纤维发育变化及其与肉质相关性的深入研究,期望获得以下预期结果:(1)揭示猪肌纤维生长发育的规律及其与肉质品质的关系;(2)研究内外因素对猪肌纤维发育的调节作用,为提高猪肉肉质品质提供技术支撑;(3)为猪肉肉质评价、猪肉生产等提供科学依据,具有一定的理论和应用价值。
2. 应用价值:本研究的研究成果,可以为猪肉生产实践提供参考和指导,同时也能够推动猪肉肉质研究的深入发展,为提高我国猪肉产业的竞争力和市场占有率做出
贡献。
基于转录组测序的巴马香猪和杜洛克猪骨骼肌差异IncRNA分析
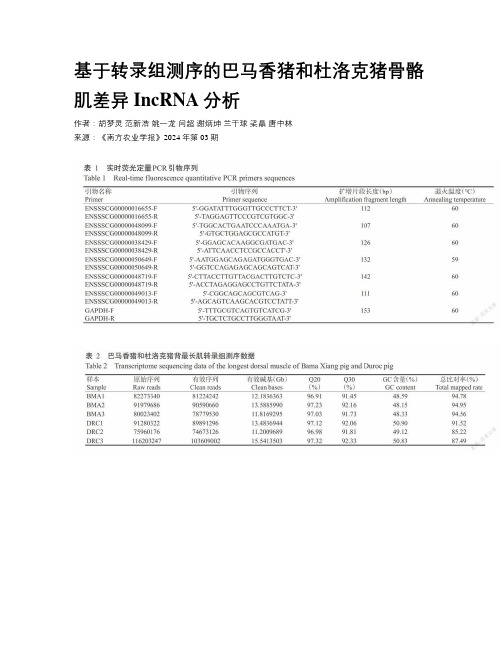
基于转录组测序的巴马香猪和杜洛克猪骨骼肌差异IncRNA分析作者:胡梦灵范新浩姚一龙闫超谢炳坤兰干球梁晶唐中林来源:《南方农业学报》2024年第03期摘要:【目的】通过对巴马香猪和杜洛克猪背最长肌组织进行转录组测序鉴定差异表达lncRNA,筛选与骨骼肌发育形成相关的候选基因,為明确lncRNA对骨骼肌生长发育的调控机制提供理论基础。
【方法】采集12月龄的巴马香猪和杜洛克猪背最长肌组织进行转录组测序,以错误发现率(FDR)<0.05且|log2Fold Change|>1为标准筛选差异表达基因(DEGs)和差异表达lncRNA,并进行实时荧光定量PCR验证,预测差异表达lncRNA靶基因并进行GO 功能注释和 KEGG信号通路富集分析,选取表达差异最大的IncRNA构建其与靶基因互作网络。
【结果】在巴马香猪和杜洛克猪背肌间共鉴定出6316个DEGs和675个差异表达lncRNA;GO功能注释分析结果表明,差异表达IncRNA靶基因主要涉及代谢过程、肌肉组织发育及骨骼肌细胞增殖和分化等生物过程;KEGG信号通路富集分析结果表明,差异表达lncRNA靶基因在Hippo和Wnt信号通路显著富集(P<0.05),说明差异表达lncRNA与骨骼肌细胞的增殖、自噬和分化相关;对表达差异最大的lncRNA-MSTRG.16703进行实时荧光定量PCR验证,发现其在巴马香猪中的相对表达量极显著高于杜洛克猪(P<0.01),与转录组测序结果一致。
lncRNA-MSTRG.16703与靶基因互作网络分析结果表明,上调靶基因包括QKI、MBNLI和YBX2,QKI和MBNLI在均与可变剪接相关。
【结论】在巴马香猪和杜洛克猪间发现的差异表达lncRNA是调控骨骼肌发育的候选基因,其靶基因主要富集在Hippo和Wnt等骨骼肌发育相关信号通路。
lncRNA-MSTRG.16703的表达差异最大且在巴马香猪中上调,其靶基因在骨骼肌细胞增殖分化和肌纤维形成中有重要调控作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
与肌肉代谢和结构有关的不同肉质表型识别基因的猪种间的肌肉转录组比较摘要背景:肉的品质取决于发生在肌肉组织中的生理过程,这可能会涉及到一系列与肌肉结构和新陈代谢特性有关的基因。
了解在屠宰过程中肉类表型之下的生物现象对揭示肉品质的变化是必要的。
因而,要对肌肉转录组进行分析,比较两个高对比度猪品种(长白(LW)和巴斯克(B))的基因表达图谱,可以知道不同的饲养环境会影响到肉的品质。
LW是养猪业中占有优势的品种,这不利于肉类质量属性的标准的设定。
B 是以低瘦肉、高脂肪含量、高肉质为特性的本地品种,该品种从遗传学的角度与欧洲其他的猪品种相距甚远。
方法论/主要发现:进行转录组分析使用一个自定义的15k微阵列,突出了不同品种之间表达有差异的1233个基因(使用α=0.05多重检测),其中的635个在B品种中高度表达,598个在LW品种中高度表达。
在不同的饲养环境中基因的表达没有不同。
另外,12个差异表达基因的表达水平被实时RT-PCR验证微阵列数据量化。
集群功能注释强调了与转录品种差异有关的四个主要集群:代谢过程、骨骼肌结构和组织、细胞外基质、溶酶体,蛋白质水解。
从而,突显出与肌肉生理和肉类品质变化有关的许多基因。
结论/意义:总之,这些结果将有助于更好地了解肌肉生理和肉品质变化潜在的分子和生物过程。
另外,本研究是识别猪肉品质的分子标记和调控手段后续发展的第一步。
引言因市场对瘦肉的需求日益增长,故引导猪的繁殖计划向获得现代肉用型猪发展。
有目的通过增加日增重和胴体瘦肉率来提高猪生产性能的同时也提高了猪的生长速率、饲料转化率、瘦肉含量和腰眼面积的增长,减少了背膘厚和胴体脂肪含量。
然而,消费者对猪肉的评价中扮演重要角色的一些肉品质性状,比如系水力、颜色、PH、肌内脂肪含量和嫩度也受到影响。
影响肉品质的因素是复杂的,它依赖于猪的基因型、环境条件、宰前处理、屠宰过程。
此外,肉品质的变化取决于发生在肌肉中的生理过程,这个过程涉及到许多与肌肉结构和代谢特点有关的基因。
弄清楚肌肉品质性状有不同的选择和非选择品种间的转录组表达谱差异,可能有助于理解肉品质变化的生理过程。
为了这个目的,设计该实验研究在胴体脂肪含量和肉品质方面有鲜明对照的两品种(LW和B)猪的背最长肌的基因表达谱。
LW在现代养猪业中是最有优势的品种,它以高瘦肉率、低脂肪含量和高日增重为特点,但有标准的肉品质。
相比之下,B是本地的品种,以低瘦肉率和高脂肪含量、高肉品质为特点,它从遗传学角度来说与其他欧洲的猪品种相去甚远。
此外,尽管越来越多的出版物关注基因表达与猪肉质量的关系,目前的转录组分析是第一个从事对高肉质量的B品种的研究。
我们研究的目的是探讨LW和B两种猪与肌肉性状和肉品质方面有关系的LM转录组谱,从而阐明文献报道中导致这两种猪产生巨大表现差异的生物现象和提高我们对肉品质的综合判断。
这两种猪要么在可选择的饲养条件下饲养(A,室内圈养和室外散养;n=10),要么在传统饲养条件下(C,全漏缝地板,n=10),研究表明不同的饲养方式已经影响了一些肌肉和肉品质的性状。
为了获得关于基因表达图谱的精确信息,转录组分析时使用了一种新的、特定的猪肌肉微阵列,即15K Genmascqchip,其中85%的探针与一个特定的注释序列和9169个特定基因相关。
对基因本体论生化过程和功能注释集群的功能分析正在进行,来突出生物网络和肌肉生理、肉类品质相关基因的密切关系。
结果增重、机体组成和背最长肌特征如表1所示,B和LW两个猪品种在增重、机体成分、最长肌组成、生物物理特征和肉的感官性状上表现出很大的不同。
由于B的日增重低,要达到相同的体重,B比LW需要更长的时间(85天),同时表现出更高的背膘厚(75%)和更低的腰肌比例(230%)。
在LM 组成上,B的含量水比LW稍高,而总的蛋白质含量基本相近。
LM的胶原蛋白含量和糖酵解潜力比B高,但肌内脂肪含量(251%)比B低。
LW也表现出高的滴水损失和剪切力,低嫩度、香味和多汁性。
饲养环境对增重、机体成分、LM组成和生物物理特征没有显著影响。
A 饲养环境中肉的嫩度比C饲养环境要低(4.0 vs.4.4, P= 0.018),但多汁性和香味并没有什么不同。
转录分析用定制的15K骨骼肌猪微阵列(Genmascqchip)进行B与LW肌肉转录组的比较,该微阵列可以通过基因表达综合平台公开利用,编号为GPL11016。
简单说,这种新的猪的骨骼肌微阵列能被很好的注释(超过70%),从而可以研究与8622个人类Entrez Gene ID相对应的9169个独特的基因。
基于网络的基因集分析工具包主要用于针对生化过程的基因本体论分类。
GO-slim(代表高水平基因本体论)用于聚焦最重要的过程。
如图1所示,13个生化过程十分突出。
其中代谢过程是最重要的一类(涵盖50%的基因),而增重过程仅涵盖不到5%的基因,仍有20%左右的基因没有归类。
通过对饲养在两个不同的饲养环境中(C, n= 10每种和A, n= 10每种)的两个品种(B, n = 20和LW,n =20)进行转录分析来比较它们肌肉表达谱的差异。
LW的基因表达并没有因为饲养环境的变化而发生改变,因为在A与C猪中没有发现差异表达探针。
相比之下,我们观察到一个对基因表达有影响的优良品种,有12%的探针在B与LW品种间表达是有差异的(P≤0.05)。
基因表达在品种间显示出显著的不同,这些品种可以根据叠化值分为两列。
叠化值可以通过当基因在B品种高度表达时B与LW样品表达比率来表示或是通过当基因在LW 品种高度表达时LW与B样品表达比率来表示。
差异表达基因与1223个独特的注释基因有关,其中635个基因在B品种高度表达(表S1),而598个基因在LW品种高度表达(表S2)。
详细的基因名称、描述、识别、叠化值和适当显著水平的BH值,在表S1和表S2中有具体说明。
如果有冗余(每个基因有超过一个探针),则叠化值总是与和微阵列数据高度一致的探针集相似。
显著差异表达(FC>2)和丰富的信息性(至少有一个相关的GO BP term)都在表2中说明,表2中的基因是在B品种中高度表达的(2<FC<=2.6),表3中的基因是在LW品种中高度表达的(2<FC<=5)。
B品种中在最长肌上高度表达的15个基因中,有四个参与了脂质代谢:磷脂酶A成员A1 (PLA1A)、蛋白质脂肪酸转移酵素亚族β(FNTB)、鞘磷脂磷酸二酯酶酸类似物3A (SMPDL3A)、激素敏感脂肪酶(LIPE)。
这四个基因参与转录或翻译:锌指蛋白410、锌指蛋白24、细胞质聚腺苷酸化元素相关蛋白2、角蛋白和Ⅱ型细胞骨架7。
还有三个基因参与离子转运或离子体内平衡:线粒体的钠氢交换、钙离子激活通道钾离子交换、液泡融合蛋白MON1同系物A。
LW品种中在最长肌上高度表达的26个基因中,有6个基因参与了转录或RNA的生成:锌指蛋白7、RNA聚合酶相关蛋白RTF1、干扰素调节因子8、LSM3同系物、去乙酰化酶3和锚蛋白重复域1。
5个基因参与防御、免疫系统或应激过程:白介素10受体亚族β、谷胱甘肽过氧化物酶8、天冬酰胺合成酶、干扰素调节因子8和锚蛋白重复域1。
有4个基因参与了氧化还原过程:aldoketo还原酶1族成员B1、谷胱甘肽过氧化物酶8、metaxin 3、乙醛酸盐还原酶/羟基丙酮酸还原酶。
最后,有3 个基因参与了葡萄糖的代谢过程:葡萄糖磷酸变位酶1、溶质载体5族成员4、乙醛酸盐还原酶/羟基丙酮酸还原酶。
通过定量RT-PCR验证基因芯片分析在差异表达基因中,选出12个通过实时定量PCR来验证微阵列差异表达结果。
其中6个基因在LW中高度表达(ADAMTS8, SPARC,GLOD4, ANKRD1, HHATL 和IGF1),6 个基因在B中高度表达(LIPE, ZNF24, FOS, FABP3,PPARD 和FHL3),在微阵列分析中FC 的扩展值在1.2—4.3之间,RT-PCR分析也是这样。
结果见图2。
用RT-PCR的方法,所有12个基因在两品种间表达有差异,使用两种方法,如微阵列和RT-PCR,每一个基因的FC值是相似的。
品种间差异表达的功能分析这两列基因,在B中高度表达的635个基因和在LW中高度表达的598个基因,使用了注释数据库进行基因本体BP富集分析,该数据库包括可视化和集成发现生物信息资源。
重要结果(显著性水平P<=0.05)见于表4。
与脂质代谢和运输、碳水化合物代谢和运输有关的基因本体BP,富集在B中高度表达的基因列表。
而生物吸附、蛋白质聚合趋化和细胞骨架组织有关的基因本体BP,富集在LW中高度表达的基因列表。
为了减少冗余并研究一个网络格式中的功能相关基因,使用DAVID工具完成了功能注释集群。
为在B(表5)中和LW(表6)中高度表达的基因列表,我们使用三个基因本体论术语、BP、细胞元件(CC)、分子函数(MF)、BIOCARTA和关于基因、基因组的KEGG [20]等方法建立了一个功能相关集群的生物模块。
根据黄等人的研究,一个相当于P=0.05的非对数标尺的富集数据1.3,已经用于集群意义的阈值来使用。
在B中高度表达的基因列表中,发现有6个改进群有超过1.3的改进数据。
根据功能把基因分为3个群:细胞骨架基因(集群1-B)、液泡和溶酶体(集群2-B)、葡萄糖代谢过程(集群4-B)。
其他三个集群(集群3-B、5-B、6-B)与转录过程有关。
在LW中高度表达的基因列表中,确定了8个集群,它们与细胞外胶原蛋白(集群1-LW, 2-LW和5-LW)、多糖绑定(集群3-LW)、细胞运动(集群4-LW)、收缩纤维和肌动蛋白聚合(集群6-LW and 7-LW)和趋药性(集群8-LW)有关。
讨论在遗传背景上,B品种是欧洲七个国家的11个品种中以“独特”为特点的本地品种。
尽管有越来越多的出版物关注基因表达与猪肉品质的关系,本研究是第一个对非选和具有高肉质猪品种的转录组分析。
我们的目标是阐明可以揭示文献中报道的B与LW品种肌肉表型差异的生物现象。
整个骨骼肌组织的转录图谱显示了一个挑战,即因为基因表达的变化可能会反映出存在于该组织中的各种细胞的mRNA组成的变化。
然而,即使我们不能把表达的改变归因于一个特定的细胞类型,我们可以假设肌纤维是主要的一种细胞类型,假设在组织上品种的比较是作为一个整体进行的。
在转录组分析之后,LW与B之间的12个差异表达的基因已经通过定量PCR分析完全确定下来了,因此也能证明新的GenmascqChip是一个研究猪的基因表达的功能强大的工具,从而可以更好地理解肌肉生理。
从遗传背景方面比较骨骼肌基因表达的差异,仅有这些研究数据是远远不够的。
本研究中两品种间差异表达的基因数目与文献记载的数量相同。