Tn5转座子插入p95基因不影响AcMNPV的复制
利用转座子建立细菌单基因突变株库的策略和方法[1]_图文_百.
![利用转座子建立细菌单基因突变株库的策略和方法[1]_图文_百.](https://img.taocdn.com/s3/m/9f55540955270722192ef76a.png)
平板上的转座突变株机器自动挑蓖落・-————-----—-畸到38好L板上培养转到96孔板上・-—__—_____--—’进行P僳I转另一个板I山l转另一个板l山戬.gsT分析数据整理●÷————————————一●}--—---————-—一图1建立转座子突变株库的技术路线子在铜绿假单胞菌PA01基因组中的转座插入采样数不符合泊松分布,虽然如此,但毕竟在最终还是获得了一个近饱和的突变株库。
而Garsin(71等在构建粪肠球菌的Tn917转座插入突变株库时发现,测序的8865个Tn917转座突变株突变基因只对应了610个不同的开放阅读框,远远低于预期的2400个。
这些结果说明使用随机性差的转座子会大大增加建库的成本。
因此,有的研究利用了两种类型转座子构建突变株,这是为了防止一种转座子可能有的插入热区,而造成在基因组插入位点的随机性不佳,进而造成某些基因的突变株在筛选中漏选,而另一些基因的突变株则被多次重复筛选。
两种类型转座子的使用,可以互相弥补,降低发生漏选或者重复筛选的可能性。
例如在建立铜绿假单胞菌PAl4突变株库工作中,最开始采用的是Tn5来源的细菌转座子——hphoA。
如前所述,尽管Tn5来源的转座子插入随机性非常好,研究者还是发现了至少一个热区(Hot spot,即gacA基因,同样也存在一些冷区(Cold spots。
为了避免转座位点的选择性,研究者又采用了真核生物来源的mariner转座子,该转座子能够在很多原核生物中进行转座,而且预计与TnphoA在靶位偏嗜上存在差异,故联合使用可以降低转座选择性;②对突变基因测定方法采用随机引物PCR的方式,以达到自动化的目的。
确定突变株突变基因的方法有很多,如反向PCR等,但这些方法的最大问题是需要人工参与,无法实现完全自动化测序,从而使确定突变基因的工作几乎不可能完成。
而随机引物PCR的方法可以采用机器自动化79进行,使测序工作完全不需要人工的参与,达到省时省力的目的,从而使建库所耗时间和精力大大降低,可以由一个研究组完成而不需要大规模的协作。
中国CHINET细菌耐药性监测

中国CHINET细菌耐药性监测随着抗生素的广泛使用,细菌耐药性已成为全球性的公共卫生问题。
为了应对这一挑战,中国CHINET细菌耐药性监测应运而生。
本文将介绍中国CHINET细菌耐药性监测的背景、目的和意义,并逐步引入给定的关键词。
中国CHINET细菌耐药性监测是我国重要的细菌耐药性监测平台,旨在全面监测细菌耐药性现状及其变化趋势。
通过该监测网络,我们可以获取全国范围内细菌耐药性的最新数据,从而为抗生素合理使用、疾病治疗和防控提供科学依据。
细菌耐药性是指细菌对抗生素产生抵抗力的现象。
随着抗生素的广泛使用,细菌耐药性不断增强,给疾病治疗带来极大的挑战。
因此,开展细菌耐药性监测对于了解现状、预测趋势和制定防控策略至关重要。
监测是指系统地收集和分析数据,以了解某一现象或事物的现状和发展趋势。
在细菌耐药性领域,监测可以帮助我们及时发现和解决耐药性问题,并为抗生素合理使用提供科学依据。
中国CHINET细菌耐药性监测便是通过收集和分析全国各地的细菌耐药性数据来实现这一目标。
中国CHINET细菌耐药性监测不仅可以帮助医生了解不同地区、不同医院的细菌耐药性现状及其变化趋势,还能为抗生素合理使用提供科学依据,提高疾病治疗效果,减少耐药菌的产生和传播。
该监测还可以为政府制定和调整相关政策提供数据支持,加强全球耐药性监测的合作与交流。
中国CHINET细菌耐药性监测在了解现状、预测趋势和制定防控策略方面具有重要意义,应加强对这一领域的和投入力度。
我们每个人都应该科学合理地使用抗生素,积极预防和控制细菌耐药性的产生和传播。
细菌耐药性监测是医院感染控制的重要环节,对于指导临床合理使用抗菌药物、提高治疗效果具有重要意义。
本文旨在介绍CHINET三级医院细菌耐药监测体系的应用目的、意义及监测方法,分析监测结果,提出相应对策,为抗菌药物的合理使用提供依据。
国际上先进的细菌耐药监测体系为各国细菌耐药监测中心所采用的标准化操作程序,包括收集临床菌株、细菌鉴定、药物敏感试验、数据整理分析等环节。
Tn5转座酶建库原理

Tn5引爆极速建库和长片段之战近10年间,NGS(Next Generation Sequencing)技术高速发展,测序仪器不断更新迭代,形成规模化。
在测序模式产业化的大环境下,测序样本制备成为其中很重要的一环。
样本起始量要求过高、建库流程繁琐等都会限制NGS技术的应用。
科学家们一直在不断探索,试图突破此类限制,其中转座酶(Transposase)技术引入建库中就是很大的进步,不仅解决了上述问题,转座酶也在NGS上开发了多个技术应用,拓展了NGS的适用范围。
20世纪40年代,美国遗传学家芭芭拉.麦克林托克在玉米的研究中发现了转座子(Transposon),随后,她花了整整6年的时间来研究转座子的奥秘。
在50年代,当麦克林托克向世人公布她的发现时,学术界并没有多少人认可,但是真理是经得起时间检验的,随后的几十年间科学家们在生物界各个领域证实了转座子系统的广泛存在。
1983年,瑞典皇家科学院诺贝尔奖金评定委员会终于把该年度的生理学和医学奖授予这位81岁高龄的、不屈不挠的女科学家。
麦克林托克是在遗传学研究领域第一位独立获得诺贝尔奖的女科学家,她的名字将和转座基因一起被载入科学史册。
转座基因俗称跳跃基因,它的发现改变了人们对基因组序列稳定性的认识。
理论上的突破也带来了应用上的拓展,科学家们群策群力,将转座子系统开发成基因研究的各种工具,这又反过来促进了遗传学理论的研究。
转座子标签(transposon tagging)技术是研究功能基因的有效工具之一。
比如模式植物大多都有很好的突变体库,其中玉米的两个应用最为广泛的突变体库(uniformMu和Ac/Ds突变体库)就是利用转座子创建的。
转座子标签技术克隆基因的基本原理:转座子是染色体上一段可移动的DNA片段,它可从染色体的一个位置跳到另一个位置。
当转座子跳跃而插入到某个功能基因时,就会引起该基因的失活,并诱导产生突变型。
通过遗传分析可确定某基因的突变是否由转座子引起,由转座子引起的突变便可以转座子DNA为探针,从突变株的基因组文库中钓出含该转座子的DNA片段,并获得含有部分突变株DNA序列的克隆,进而以该DNA为探针,筛选野生型的基因组文库,最终得到完整的基因。
转座因子介绍

• Mu噬菌体为一37kb的线状DNA,两端各 带一小段大肠杆菌的DNA,这与该噬菌体 插人大肠杆菌染色体上有关。 • 距末端不远处也有类似于IS的序列,但位 置不对称。靠近一端处存在与转座有关的A、 B基因,它们分别编码70000和33000两 种蛋白,在A、B与末端之间有一C区,对A、 B有负调控作用。
宿主 DNA
过量的 OUT-RNA 与其配对,抑制 IN-RNA 的转录
甲基化阻止转录酶和 DNA 结合
甲基化阻止转录酶合成 图 23- 46 几种抑制 Tn10 专座的机制,主要是通过控专座酶的合成来调节
主要类型转座子
类型
细菌插入序 列(IS)
结构特征
TIRs;转座酶和或拆 分酶编码基因 IS组份;抗药基因 TIRs;蛋白质编码 基因(有内含子)
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶位点连接。
3 转座作用的遗传效应
• • • • 引起插入突变 产生新的基因 产生染色体畸变 引起生物进化
2 转座因子的发现和检出
1951年McClintock提出转座(Transposition)和
跳跃基因(jumping gene)的新概念;
1967年Shapiro才在E.coli中发现了转座因子(transposable element)。
Barbara McClintock
(1902-1992)
转座子概述

自主的Ac 元件大部分长度都被一个含五个外显子的基因占据。其产物是转座酶。 元件末端为一个11bp的反向重复序列;在靶序列在插入位点形成8bp重复。
Spm-dSpm系统:
Spm: ❖ 自主性因子,具有转座、整合和解离活性; ❖ 两端各有一个13bp的倒置重复序列,在其靶DNA位点复制形成
3bp的正向重复。
dSpm(defective Spm): ❖ 非自主性因子,所有dSpm都是功能性Spm的缺失突变体; ❖ Spm-dSpm系统在功能上与Ac-Ds系统相似,也可以引入基因的
插入突变,影响结构基因表达,还能导致染色体断裂。
Spm/En有两个基因,tnpA 有11 个外显子,经转录拼接产生2500bp 的 mRNA,tnpB 的6000bpmRNA中包含ORF1和ORF2。
二、转座子的分类
1、插入序列(insertional sequence IS)
特征: ❖两端都有短的4-15bp的正向重复序列(direct repeats, DR) (靶序列); ❖末端有略长的15-25bp的反向重复序列(inverted repeats, IR); ❖1kb左右的编码区,它仅编码和转座有关的转座酶。
转座产生的染色体畸变
当复制性转座发生在宿主DNA原有位点附近时,往往导 致转座子两个拷贝之间的同源重组,引起DNA缺失或倒位。 若同源重组发生在两个正向重复转座区之间,就导致宿主染 色体DNA缺失;若重组发生在两个反向重复转座区之间,则 引起染色体DNA倒位。
正向重复之间的互 惠 (Reciprocal) 重 组 会将它们之间的序 列切除,中间区域 会以环状DNA 的形 式被切除(从细胞中 消失);染色体仍保 留正向重复的一个 拷贝。
转座子TnA的结构示意图
tn5转座酶随机识别dna的原理

tn5转座酶随机识别dna的原理下载温馨提示:该文档是我店铺精心编制而成,希望大家下载以后,能够帮助大家解决实际的问题。
此文下载后可定制随意修改,请根据实际需要进行相应的调整和使用。
并且,本店铺为大家提供各种各样类型的实用资料,如教育随笔、日记赏析、句子摘抄、古诗大全、经典美文、话题作文、工作总结、词语解析、文案摘录、其他资料等等,如想了解不同资料格式和写法,敬请关注!Downloaded tips: This document is carefully compiled by the editor. I hope that after you download them, they can help you solve practical problems. The documentscan be customized and modified after downloading, please adjust and use it accordingto actual needs, thank you!In addition, our shop provides you with various types of practical materials, such as educational essays, diary appreciation, sentence excerpts, ancient poems, classic articles, topic composition, work summary, word parsing, copy excerpts, other materials and so on, want to know different data formats and writing methods, please pay attention!DNA是生命的基础,它携带着生物体遗传信息的蓝图。
转座子概述

末端11bp的IR和8bp的DR,DR是由靶位点重复而成。
Ds:
各种Ds因子的长度和序列都不相同,但和Ac相关。其末端同样 有11bp的IR。
Ds比Ac短,其缺失的长度不同,一个极端的例子是Ds9因子仅 缺失194bp,另一个例子是Ds6因子长仅有2.5kb,相当于Ac两端 各1kb。
转座产生的染色体畸变
当复制性转座发生在宿主DNA原有位点附近时,往往导 致转座子两个拷贝之间的同源重组,引起DNA缺失或倒位。 若同源重组发生在两个正向重复转座区之间,就导致宿主染 色体DNA缺失;若重组发生在两个反向重复转座区之间,则 引起染色体DNA倒位。
正向重复之间的互 惠 (Reciprocal) 重 组 会将它们之间的序 列切除,中间区域 会以环状DNA 的形 式被切除(从细胞中 消失);染色体仍保 留正向重复的一个 拷贝。
转座子概述
主讲内容
一、转座子的概念 二、转座子的分类 三、转座发生的机制 四、真核生物的转座因子
一、概念ห้องสมุดไป่ตู้
转座子(transposon,简称Tn), 又称易位子,是指存在 于 染 色 体 DNA 上 可 以 自 主 复 制 和 移 位 的 一 段 DNA 序 列 。
转座子可以在不同复制子之间转移,以非正常重组方式 从一个位点插入到另外一个位点,对新位点基因的结构与表 达产生多种遗传效应。
dSpm(defective Spm): 非自主性因子,所有dSpm都是功能性Spm的缺失突变体; Spm-dSpm系统在功能上与Ac-Ds系统相似,也可以引入基因的
插入突变,影响结构基因表达,还能导致染色体断裂。
Spm/En有两个基因,tnpA 有11 个外显子,经转录拼接产生2500bp 的 mRNA,tnpB 的6000bpmRNA中包含ORF1和ORF2。
实验五转座子引起的插入突变

实验五 转座子引起的插入突变
厦门大学生命科学学院
冷落四十年的转座子理论
1983年,美国遗传学家巴巴拉.麦克林托 克(B.McClintock)由于发现了可移动 的遗传物质,被授予诺贝尔医学奖。 人们把麦氏的成就比之为一百年前另一 位伟大的遗传学家孟德尔的成就。
冷落的原因和启示
1.
2.
3.
从转座子理论和经典遗传学的关系来看,转座子理论 推翻了经典遗传学关于基因是稳定的这一传统观念, 是一种革命性的理论,因而不为经典遗传学家所接受。 从转座子理论和分子遗传学的关系来看,是由于前者 走在了时代前面,是一种超时代发现。科学界还没有 做好接受它的准备。因而遭到分子遗传学家的冷落。 从转座子理论赖以建立的实验材料看,是由于它离开 了分子生物学的主流。麦克林托克虽然身在冷泉港生 物学实验室,但她所采用的材料,与该室中极大多数 科学家不同。她没采用病毒和细菌作材料,研究基因 的拼接、剪切和重组,而是采用玉米这样的高等植物 作为研究对象。
30岁那年,麦克林托克在某些玉米籽粒 中发现了玉米色素显现着一些稀奇古怪 的模式。她观察到玉米籽粒颜色的遗传 很不稳定,有时籽粒上还出现一些斑斑 点点。 她通过耐心的记录和仔细的分析,发现 使籽粒着色的色素基因是在某一特定代 上“接上”或“拉断”的。
1951年,在冷泉港生物学专题讨论会上,麦克 林托克递交了自己的学术论文,向科学界同行 报告了她的新理论。她提出遗传基因可以转移, 能从染色体的一个位置跳到另一个位置,甚至 从一条染色体跳到另一条染色体上。她把这种 能自发转移的遗传基因称为“转座因子” 。 “转座因子”除了具有跳动的特性之外,还具 有控制其他其因开闭的作用,因此“转座因子” 又可叫做“控制因子”。
Tn5Transposas-中文
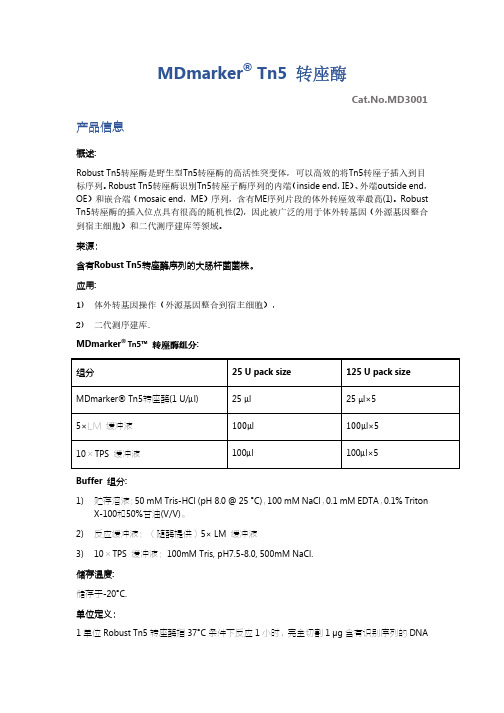
MDmarker® Tn5 转座酶Cat.No.MD3001 产品信息概述:Robust Tn5转座酶是野生型Tn5转座酶的高活性突变体,可以高效的将Tn5转座子插入到目标序列。
Robust Tn5转座酶识别Tn5转座子酶序列的内端(inside end,IE)、外端outside end,OE)和嵌合端(mosaic end,ME)序列,含有ME序列片段的体外转座效率最高(1)。
Robust Tn5转座酶的插入位点具有很高的随机性(2),因此被广泛的用于体外转基因(外源基因整合到宿主细胞)和二代测序建库等领域。
来源:含有Robust Tn5转座酶序列的大肠杆菌菌株。
应用:1)体外转基因操作(外源基因整合到宿主细胞),2)二代测序建库.MDmarker® Tn5™ 转座酶组分:组分25 U pack size 125 U pack size MDmarker® Tn5转座酶(1 U/μl) 25 μl 25 μl×55×LM缓冲液100μl 100μl×510×TPS 缓冲液100μl 100μl×5Buffer 组分:1)贮存溶液:50 mM Tris-HCl (pH 8.0 @ 25 °C),100 mM NaCl,0.1 mM EDTA,0.1% TritonX-100和50%甘油(V/V)。
2)反应缓冲液:(随酶提供)5× LM 缓冲液3)10×TPS 缓冲液:100mM Tris, pH7.5-8.0, 500mM NaCl.储存温度:储存于-20°C.单位定义:1单位Robust Tn5转座酶指37°C条件下反应1小时,完全切割1 μg含有识别序列的DNA片段所需要的酶量。
Protocols:1. 体外转基因操作MDmarker® Tn5转座酶可以将含有成对识别序列的双链DNA片段(如下图所示)随机整合到宿主细胞的基因组中。
转座因子新版

供体和受体形成旳这种构造称为共整合体(cointegrate)。
所谓共整合体,就是两个或两个以上旳复制子经过共价键连 接在一起。共整合体在原来两个分子之间结合处具有两个转 座子旳拷贝,方向为正向反复。
IS21 2132 10/11 4
2
R68-45(铜绿假单孢菌
IS50 1534 8/9
9
3
Tn5(肠杆菌)
IS51 1311 26/26
3
2 萨氏假单胞菌
(Pseudomonase savastanoi)
IS91 1800 8/9
0
1 pSU233(肠杆菌)
IS150 1443 19/24 3
(4) 交错末端旳产生和填充解释了在插入部位产生靶DNA正 向反复旳原因。
转座因子旳插入引起靶序列反复 靶序列反复旳产生过程
图10-5 转座因子旳插入机制
2. 细菌转座因子旳转座模型
(1) 剪-贴型转座(cut-and-paste transposition)
此类转座又叫简朴插入,是一种非复制型转座过程。
5 Shigella flexneri
Tn501 8200 Hgr 38
5 Pseudomonase aeruginosa
Tn1000 (Υδ) 5800 无 37
5 E. coli
起源于G+ 细菌
Tn551 5300 Ery(erythromycin) 35 5 Staphylococcus aureus
tn5转座子原理_理论说明以及概述

tn5转座子原理理论说明以及概述引言部分内容示例:1. 引言1.1 概述本文旨在探讨tn5 转座子的原理、理论说明以及其在相关领域中的应用。
对于转座子这一概念,它是指生物体内部或不同个体间基因组DNA序列移动或重排的过程。
tn5 转座子作为一种广泛存在于细菌中的转座子,具有独特的结构和功能,在生命科学领域中具有重要的研究意义和应用前景。
1.2 文章结构本文共分为五个主要部分进行阐述,分别是引言、tn5转座子原理、理论说明、实验验证与研究进展以及结论。
下面将逐一介绍各个部分内容,并给出相应章节的概览。
1.3 目的通过撰写这篇文章,旨在全面解析tn5 转座子在转座反应中所起到的关键作用和功能机制,并从理论上对其进行深入探讨。
同时,详细说明了tn5转座子在实验验证和研究进展方面的最新成果,并展望未来发展方向。
通过本文,希望能够增加读者对tn5 转座子的理解,并为相关领域的研究者提供重要的参考资料和思路。
2. tn5转座子原理2.1 转座反应概述转座是指基因组中的DNA序列从一个位置移动到另一个位置的过程。
tn5转座子是一种广泛存在于细菌和真核生物中的转座子,它可以在基因组中发生自由或复制式的转座反应。
2.2 tn5转座子的结构与功能tn5转座子由两个关键部分组成:外翻酶(transposase)基因和IRs(inverted repeats)序列。
外翻酶是负责催化DNA切割和连接的酶,IRs序列则是在转座反应过程中提供结构支持和识别特定位点的重要元素。
tn5转座子通过外翻酶将自身从一个位置切割出来,并插入到新的目标位点上。
在这个过程中,外翻酶首先结合到IRs序列上,并切割出tn5转座子与目标DNA 之间的连接部分。
然后,外翻酶会介导tn5转座子在目标位点上生成一个“缺口”,并将其紧密地连接到目标DNA上。
2.3 转座机制解析tn5转座子的具体机制可以分为两步:切割与连接。
在切割步骤中,外翻酶首先结合到IRs序列上,并识别目标位点。
转座子的插入突变及转座子的应用

下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶位点连接。
(二)复制 型转座
特征:
1、转座后原 来位置上的 转座因子保 持不变;
2、转座过程 中有一共整 合体。
Tn 靶 单链交错切割
转座子被切开的 末端和靶被切开 的末端连接 交叉结构(单链转移复合
2、构建突变体库
正因为以上转座子的特征,转座子才 被广泛应用于构建插入突变体库,现 已成为研究基因功能的重要手段之一。 目前研究和应用得较多的有Tn10、 Tn5、Tn3和Mu噬菌体 。
3、鉴别菌株及群体多样性
转座子通常限制性地分布于特定的真 菌菌株或群体中,可以作为特定菌株 的诊断工具,已用于丝状真菌群体多 样性分析。在医药工业方面已用于有 益菌株的鉴别, 在植物病理学方面已 用于鉴别特定的病原 。
转座子的插入突变及转座子 的应用
主讲人:杨长友 本组成员:杨菊 孙青芝 陈骊君 陆朝军 杨长友
内容提要
一、转座子的发现 二、转座子的分类 三、转座子的转座机制 四、转座子的遗传效应 五、转座子插入位点的鉴定方法 六、转座子的应用
一、转座子的发现
转座子(transponson,简称Tn), 又称易 位子,是指存在于染色体DNA上可以自主 复制和位移的一段DNA序列。
从 3’ 末端复制 产生共合体
重组
共合体
四、转座的遗传学效应
❖ (1)插入突变失活或改变基因表达的 模式。
插入突变失活是转座的最直接效应,但 转座也可以通过干扰宿主基因与其调控元件 之间的关系或改变DNA的结构而影响基因的 表达。
四、转座的遗传学效应
细菌转座因子的插入机制

Tn5 5700bp
CamR Tn9 2638bp
IS1
IS1
( C) IS10
TetR Tn10 9300bp IS10
2.复杂转座子(TnA转座子)
• TnA转座子广泛存在于细菌中,从不同地区不同菌种分 离到的质粒都携带有TnA转座子。 • 尽管这些质粒互不相关,复制系统和转移系统都差异很 大,但每个质粒都带有TnA转座子。 • TnA能转座到大多数质粒、噬菌体及革兰阴性菌的染色 体上。
TnA转座子的基本特征见表
转座子 Tn1 Tn3 Tn21 长度/bp 5000 4957 19600 遗传标记 Amp Amp Mer Str Sul 末端反向 重复/bp 38 38 38
两侧靶点正向 重复/bp
来源 假单胞菌 沙门氏菌 志贺菌
5 5 5
Tn501
Tn1000 Tn551 Tn4430 Tn4451 Tn4456
插入序列的结构
两个IR之间仅含有编码转座酶(Transposnase 简称Tnp) 的基因,该酶是转座所必需的,并能准确识别IS两端的序 列。
转座插入的靶位点:IS插入时,在靶DNA位点产生一个39bp的正向重复序列(direct repeat sequence,DR) 。
IS插入目标DNA的靶序列(5bp)以及 在IS两端的同向重复序列(蓝色阴影)
37kb
1kb
Mu DNA包装
第二节 细菌转座因子的遗传效应和应用 一、转座因子的遗传效应
插入突变、基因重排、极性效应
1.插入突变
当 IS 、 Tn 插入到一个基因时,该基因的功能受到破坏, 其表型和一般突变体相同,如营养缺陷型、酶活性丧失等; 另外,由于Tn总是带有抗药性基因,所以转座子插入后 使该基因具有抗药性。
转座子概述
杂种不育:
当P品系雄果蝇和
M品系雌果蝇交配
时,可能由于雄果
蝇 DNA 的 进 入 而
突然生成了转座酶,
结果使很多的P因
子发生转座,造成
插入突变。这样杂
交生下的子代出现
染色体畸变和生育
力下降。
杂种不育是不对称的,它是由P品系的雄果蝇和M品系的雌果蝇杂 交产生的,但M品系的雄果蝇和P 品系的雌果蝇不会产生杂种不育。
IS1 IS2 IS4 IS5 IS10R IS50R IS903
IS序列的结构特征比较
长度(bp)
768 1327 1428 1195 1329 1531 1057
两段倒置重 靶位点正向 复区(bp) 重复区(bp)
靶位点
23
9
随机
41
5
有热点
18
11或12
AAAN20TT
16
4
有热点
22
9
NGCTNAGCN
Ac-Ds转座元件
❖ 1951年McClintock-麦克林托克提出转座(Transposition) 和跳跃基因(jumping gene)的新概念;
❖ 1967年Shapiro才在E.coli中发现了转座因子(transposable element)。
Barbara McClintock(1902-1992) Nobel Prize for Physiology or Medicine 1983
二、转座子的分类
1、插入序列(insertional sequence IS)
特征: ❖两端都有短的4-15bp的正向重复序列(direct repeats, DR) (靶序列); ❖末端有略长的15-25bp的反向重复序列(inverted repeats, IR); ❖1kb左右的编码区,它仅编码和转座有关的转座酶。
利用Tn5转座子构建杆状病毒AcMNPV随机突变体的初步研究

文章编号:042727104(2005)0420498205收稿日期:2005205209基金项目:国家自然科学基金资助项目(30070031)作者简介:李 惠(1979—),男,硕士研究生,通讯联系人钟 江教授.E 2mail :jzhong @.利用T n5转座子构建杆状病毒AcMNPV 随机突变体的初步研究李 惠,赵明磊,尹 隽,钟 江(复旦大学生命科学学院微生物学与微生物工程系,上海 200433)摘 要:以杆状病毒模式种AcMNPV 为研究对象,应用基于Tn5转座子的随机转座的方法,构建杆状病毒突变体库.将果蝇hsp70启动子后接绿色荧光蛋白基因后插入Tn5转座子,构建了可以在昆虫细胞中表达,易于跟踪的转座载体.利用体外转座系统将转座子随机插入AcMNPV 基因组,并用转座反应液转染S f21细胞,得到了表达绿色荧光蛋白的病毒突变体库.进一步纯化了两株病毒B9F 和Li6A ,进行了转座子插入位点的分析,确定两株病毒中,转座子分别插入了94K 基因和p10基因.该方法将为杆状病毒功能基因组研究提供重要的手段.关键词:AcMNPV ;Tn5转座子;转座酶;突变体中图分类号:Q 939.4 文献标识码:A 杆状病毒A utographa calif ornica multicapsid nucleopolyhedrovirus (AcMN PV )是一种应用广泛的真核表达载体1,2和具有潜力的生物杀虫剂3,4,近年来的研究显示它也有可能发展成为一种有效的哺乳动物细胞基因转导和基因治疗的载体5.AcMN PV 基因组长133894bp 6,共有154个潜在的开放阅读框.目前共有约70个基因的功能得到不同程度的研究和鉴定.同时,尚有大量基因的功能有待深入研究.高效地获得各种突变体是分析研究基因功能的重要环节.Tn5是一种细菌的转座子,两端分别带有转座元件IS50R 和IS50L ,可在Tn5转座酶催化下随机插入、整合到目标DNA 序列中,同时带入两端转座元件以内的DNA 序列.本研究构建了带有昆虫细胞表达元件和绿色荧光蛋白基因的转座载体,通过体外转座反应和细胞转染得到了带有随机插入突变的AcMN 2PV 突变库.在此基础上分离并分析了2个突变病毒株.1 材料和方法1.1 细胞、病毒和质粒草地贪夜蛾(S podoptera f rugierda )Sf21细胞系,E.coli DH5α,AcMN PV 1A 株,质粒p HZ402,pAcDZ1均为本实验室保存,转座子构建质粒pMod2购自Epicentre.Sf21细胞用TNM 2FH 培养液加10%胎牛血清培养.细菌、质粒、基因操作参照文献7.1.2 带有昆虫细胞表达框架的T n5转座子的构建以pAcDZ1为模板通过PCR 扩增果蝇hsp70启动子,引物分别为5’2ACA TGCA TGCTA 2G AA TCCCAAAACAAACTG 23’和5’2GG AA TTCTA TTCA G A GTTCTCTTCTTG 23’,反应条件为预变性95℃5min ;然后95℃1min ,56℃30s ,72℃1min 共30个循环;最后72℃延伸10min.扩增得到长度约为500bp 的产物,经电泳、胶回收试剂盒(V 2gene )纯化,并克隆到T 载体(Ta KaRa )上,得到p T 2hsp70p ,序列分析验证其正确.用Nco Ⅰ和S ac Ⅰ双酶切p T 2hsp70p ,分离2.7kb 片段;用同样的酶双酶切p HZ402,胶回收分离800bp 的gf p 基因片段.两片段连接得到p T 2hsp70p 2gfp.以B am H Ⅰ和S ac Ⅰ双酶第44卷 第4期2005年8月复旦学报(自然科学版)Journal of Fudan University (Natural Science )Vol.44No.4Aug.2005切p T 2hsp70p 2gfp ,回收1.3kb 片段,连接到用同样的酶双酶切处理的转座子构建质粒pMod2上,得到pMod 2hsp70p 2gfp (见图1).1.3 AcMNPV D NA 的制备Sf21细胞悬浮培养至约1×106个时(参照文献8),按MO I =1pfu/cell 接种AcMN PV ,48h 后,细胞悬液以5000r/min 离心15min ,取上清液.上清液再以17000r/min 4℃离心30min ,用450μL 无菌水悬浮沉淀.加入50μL 10%的SDS ,56℃30min ;加入蛋白酶K 至100μg/mL ,再次56℃30min.用酚/氯仿法抽提病毒基因组DNA ,乙醇沉淀DNA.以50μL 无菌水溶解DNA ,4℃放置待用.图1 pMod 2hsp70p 2gfp 质粒及转座子示意图Fig.1 Structure of pMod 2hsp70p 2gfp1.4 体外转座以pMod 2hsp70p 2gfp 为模板进行PCR ,引物分别为FPP (5’2A TTCA GGCTGCGCAACTGT 23’)和RPP (5’2GTCA GTG A GCG A GG AA GCGG AA G 23’)(Epicentre ,图1),得到线性转座子DNA 片段.反应条件为:94℃预变性5min ;然后,94℃30s ,55℃30s ,72℃1min 共30个循环;最后72℃延伸10min.PCR 产物纯化后用于进行体外转座反应.转座反应参照Epicentre EZ ∶∶TN 转座酶说明书进行,10μL 反应液包括2μL AcMN PV DNA ,6μL 线性转座子序列,1μL 10×buffer 和1μL EZ ∶∶TN 转座酶.37℃反应过夜.1.5 细胞转染5×105个Sf21细胞27℃静置培养2h ,使细胞贴壁.取5μL 上述转座反应液,用Cellfectin (Invitro 2gen )转染细胞,方法参见产品说明书.转染后细胞27℃培养,Nikon HFX 倒置荧光显微镜观察.转染后72h ,收集细胞的培养上清液,作为重组病毒库的原代病毒液,4℃保存备用.1.6 突变体病毒株的分离通过空斑试验进行.六孔细胞培养板中每孔加入5×105个Sf21细胞,27℃静置2h 后,吸去上清液,加入适当稀释度的病毒液1mL ,27℃感染3h 后,吸去病毒液,同时将3%低熔点琼脂糖融解,与两倍体积的细胞培养液混合,加入到各孔中,每孔2mL.静置培养1h ,待琼脂糖凝固后,每孔再加入1mL 培养液.27℃培养3~5d ,至空斑出现.挑取空斑,悬浮于培养液中.如此重复空斑试验数轮,得到纯化的病毒株.图2 PCR 确定转座子的插入位点示意图Fig.2 Primers for the determination of transposon interstion site via PCR1.7 转座子插入位点分析沿病毒基因组每隔3kb 左右设计一个引物,共44个引物(Ac1244,图2,表1),同时根据转座子序列设计向上游和向下游的引物(Mod 2up 和Mod 2down ,图2,表1).分别以Mod 2up 、Mod 2down 与Ac1244组成引物对,共88对,进行PCR 反应.Mod 2up 或Mod 2down 可与转座子插入位点附近的一个引物产生PCR 反应产物(图2).与野生型病毒对照,并通过对PCR 产物的序列分析,确定转座子的插入位点.2 结 果2.1 携带昆虫细胞表达元件的转座子的构建PCR 扩增hsp70启动子,并接到绿色荧光蛋白基因(gf p )上游,然后再将该表达框插入转座子构994 第4期李 惠等:利用Tn5转座子构建杆状病毒AcMNPV 随机突变体的初步研究建质粒pMod2,得到带有昆虫细胞表达框架的Tn5转座子的质粒pMod 2hsp70p 2gfp (图1).将质粒转染细胞后,荧光显微镜观察可见部分细胞产生微弱绿色荧光(图未显示),表明该hsp70启动子/gfp 表达框架可以在昆虫细胞中表达.表1 用于确定转座子位点的PCR 引物的序列和在病毒基因组上的位置Tab.1 The nucleotide sequences and positions in AcMNPV genome of primers used for the determination oftransposon instertion site via PCR名称序列(5’———3’)起点位名称序列(5’———3’)起点位Ac 21tgtggaccgcagaacagata 627Ac 224cgaatatggacctaacaacc 71362Ac 22aaggctctgacgcatttcta 3545Ac 225tgtggtaatagtggcgttgg 74761Ac 23cacaacggaaggtcgtctgc 6445Ac 226ccaacaaccgagttagagta 77805Ac 24atggattgcgagtatttgcg 9338Ac 227cacggcaatacctatcatct 80922Ac 25tacgcaaggcggactacaat 12520Ac 228cgttactttccaacacccag 84065Ac 26gacgcaacacgactacactg 15473Ac 229gcaaacgacgaccgcataat 87366Ac 27cggcatcaacgagccaactt 18365Ac 230ctgaatagcgatgctgatgc 90449Ac 28ctcctccgaaggtccgtcta 21327Ac 231gcttactgtgcctgtatcaa 93342Ac 29ccagttcaacaatccctctt 24462Ac 232ttgcgagaccgtcaacataa 96487Ac 210ctccgtctggatttactgcc 27451Ac 233ctcggtgttcccgtatcgtc 99366Ac 211cgatgacctcgtggtatgga 30866Ac 234caagggcaacaaatagacgc 102229Ac 212gagaatagccgtcgccacaa 33991Ac 235gcatcaatctcccaagcaaa 105343Ac 213ccaccactaccaacaacaac 36884Ac 236cccttctttgtagatgctgt 108545Ac 214acccttcttggaacacgaca 39922Ac 237agactcgttacccgacttga 111566Ac 215aatctgccgtccagcataaa 42864Ac 238tccgagacataccacaaagc 114765Ac 216gcagaaagcgatagtgaaag 45756Ac 239tggctcataactaaactcgc 117940Ac 217agcctgctgtcgtgaatacc 48985Ac 240gcggcacataataatcgtcg 120913Ac 218aaccgctgtcgtaatcttgg 52227Ac 241cgcaagatgatggctttcct 124001Ac 219tgacgcacaacatcaactac 55542Ac 242gttcgccattagggcagtat 127099Ac 220cggctcaccgctactttctc 58834Ac 243aaaactgccgtcgtcaatac 130185Ac 221cgtttagggattctatggtg 62188Ac 244gaacggagcgtgattagtgt 133066Ac 222tcgtcgtgttgtcatagccc 65122Mod 2down acgactacgcactagccaaca NA Ac 223cgacctttccacctatcacg68114Mod 2uptcggcatggacgagctgtacNA图3 PCR 检测病毒突变株中的hsp70启动子Fig.3 PCR detection of the hs p70promoter in the mutant virus 1.DNA 分子质量标准(λDNA Hi n d Ⅲ/Eco R Ⅰ);2.B9F 突变病毒株DNA 为模板;3.Li6A 突变病毒株DNA 为模板;4.野生型AcMNPV DNA 为模板.2.2 AcMNPV 突变体库的构建PCR 扩增pMod 2hsp70p 2gfp 中转座子序列,得1.4kb 的线性化片断.将病毒基因组DNA 和该转座子片段混合,在EZ ∶∶TN 转座酶的催化下,进行体外转座.转座反应液直接用于转染Sf21细胞.一天后在荧光显微镜下观察到绿色荧光.由此得到随机插入突变病毒库.对原代病毒液进行效价测定,达2×106pfu/mL ,其中产生绿色荧光的约占20%.将该原代病毒再次感染Sf21细胞,仍然观察到细胞发出荧光,证明转座子已经成功插入病毒基因组.2.3 突变病毒株的纯化空斑试验和多孔板稀释法从突变病毒库中分离得到了2株突变病毒株,分别为B9F 和Li6A.抽提病毒基因组DNA ,并以之为模板,用扩增hsp70启动子的引物进行PCR 检测,可以从突变病毒株的DNA 中扩增得到hsp70启动子片断,以野生型病毒DNA 为模板则不能得到该片断(图3).证明突变病毒株中带有插入的转座子.2.4 转座子插入位点的确定沿AcMN PV 基因组顺时针方向每隔3kb 左右设计1个寡聚核苷酸(Ac12Ac44,表1),分别以它们和根据转座子内部序列设计的寡聚核苷酸Mod 2up 和Mod 2down 作为引物,以突变病毒株的基因组DNA 为模板进行PCR ,对阳性的PCR 产物进行序列分析,从而确定了两株病毒中转座子的插入位点,结果如图4所示.这两个突变05 复旦学报(自然科学版)第44卷 株中,转座子分别插入并破坏了AcMN PV 的94K 基因和p10基因.B9F 突变株中,转座子插入位点位于94K 基因编码区,置换了基因组中114585至115089bp 之间的区域(图4A ),而Li6A 突变株中,转座子的插入位点位于AcMN PV 的p10基因编码区的119057bp 位点(图4B ).表明这两个基因都是病毒在体外细胞中复制非必需的位点.图4 B9F 和Li6A 突变株转座子插入位点示意图Fig.4 Transposon insertion sites of the mutant AcMNPV B9F and Li6A3 讨 论杆状病毒基因组学和基因功能的研究已经取得了很大的进展,对数十种病毒的基因组进行了全序列的分析,对其中的相当部分基因,特别是对于那些在病毒感染和复制中具有关键作用的基因进行了转录表达和功能的分析9.但由于杆状病毒的基因组庞大,其中一半左右的病毒基因的功能尚未有深入研究.获得大量的突变体病毒是研究病毒基因功能的重要途径之一.随机位点插入转座子在帮助人们分离大量基因功能破坏的突变方面发挥了重要的作用,已经被用在研究一些微生物乃至高等生物的基因组中10~13.杆状病毒的基因组是双链DNA ,具有感染性,DNA 转染细胞后就能开始复制,产生子代病毒.因此,杆状病毒非常适合用体外随机转座的方法获得大量的突变病毒,但目前国内外尚未有这方面研究的报道.本研究利用转座子的方法获得了一个随机突变AcMN PV 库,初步从中分离了两个突变病毒株,B9F 和Li6A ,确定了这两个病毒分离株中转座子插入位点分别为病毒的94K 基因和p10基因.AcMN PV 94K 基因是一种早期表达的基因,其基因功能还不清楚,对病毒感染细胞非必需14.p10基因则是一个在病毒感染极晚期高水平表达的基因,与病毒感染的细胞中纤维状结构有关,但对于病毒在体外培养细胞中的感染与复制也是非必需的15.这表明可以利用该方法确定病毒的非必需基因.由于破坏必需基因功能的突变会导致无法获得纯的突变株,因此尚不能以本文报道的方法进行研究,需要设计其他的研究手段,如通过改变培养条件,增加异源辅助病毒共同感染,或借助病毒全基因组的BAC (细菌人工染色体)克隆先在细菌中获得必需基因突变的病毒基因组的方法.这些研究还在进行中.利用转座子随机突变技术研究杆状病毒基因功能的工作可以帮助我们更加深入的认识病毒复制的机制,认识病毒与细胞及宿主昆虫之间的相互作用,从而为更好地利用这一病毒提供理论基础.如何更加高效地获得病毒分离株,并确定转座子的插入位点,还需要通过进一步的研究加以解决.参考文献:1 Possee R D.Baculoviruses as ex pression vectorsJ .Curr Opin Biotechnol ,1997,8:569272.2 Vialard J E ,Arif B M ,Richardson C D.Introduction to the molecular biolo gy of baculovirusesJ .Methds MolBiol ,1995,39:1224.3 Cunningham J C.Baculovirus as microbial insecticides M .In “Novel Approached to Integrated Pest Manage 2ment ”(R Reuveni Ed.).Boca Raton :CRC Press ,1995.2612292.4 Black B C ,Brennan L A ,Dierks P M ,et al .Commercialization of baculoviral insecticides M .In “The Bac 2uloviruses ”(L ler ,Ed ).New Y ork :Plenum Press ,1997.3412387.5 Ghosh S ,Parvez M K ,Banerjee K ,et al .Baculovirus as mammalian cell ex pression vector for gene therapy :an105 第4期李 惠等:利用Tn5转座子构建杆状病毒AcMNPV 随机突变体的初步研究emerging strategyJ .Mol Ther ,2002,6:5211.6 Ayers M D ,Howard S C ,Kuzio J ,et al .The complete DNA sequence of A utographa calif ornica nuclear poly 2hedrosis virusJ .V irology ,1994,202:5862605.7 Ausubel F M ,Brent R ,K ingstonR E ,et al .Short proptocols in molecular biology M .3rd ed.New Y ork :JohnWiley &S on ,Inc.,1995.(in Chinese )8 朱媛娥,徐慧斌,赵志安,等.昆虫杆状病毒表达系统表达人载脂蛋白A 2I J .生物工程学报,2003,19:6922697.9 Herniou E A ,Olszewski J A ,Cory J S ,et al .The genome sequence and evolution of baculovirusesJ .A nn RevEntomol ,2003,48:2112234.10 Yu D ,Silva M C ,Shenk T.Functional ma p of human cytomegalovirus AD169defined by global mutational anal 2ysisJ .PNA S ,2003,100:123962401.11 Carlson C M ,Dupuy A J ,Fritz S ,et al .Transposon mutagenesis of the mouse germlineJ .Genetics ,2003,165:243256.12 Otero J ,Jacobs W R J r ,G lickman M S.E fficient allelic exchan ge and transposon mutagenesis in Mycobacteriumavium by specialized transductionJ .A ppl Environ Microbiol ,2003,69:503925044.13 Castano I ,K aur R ,Pan S ,et al .Tn72based genome 2wide random insertional mutagenesis of Candida glabrataJ .Genome Res ,2003,13:905215.14 Friesen P D ,Miller L K.Divergent transcription of early 352and 942kilodalton protein genes encoded by theHin dIII K genome fragment of the baculovirus A utographa calif ornica nuclear polyhedrosis virusJ .J V irol ,1987,61:226422272.15 Vlak J M ,K linkenberg F A ,Zaal KJ ,et al .Functional studies on the p10gene of A utographa Calif ornica nu 2clear polyhedrosis virus using a recombinant expressing a p102beta 2galactosidase fusion gene J .J Gen V irol ,1988,69:765276.Preliminary Studies on the Construction of T n 5T ransposonB ased on R andom Insertional Mutants of B aculovirusLI Hui ,ZH AO Ming 2lei ,YI N J uan ,ZHONG Jiang(Depart ment of Microbiology and Microbial Engineering ,School of L if e Sciences ,Fudan U niversity ,S hanghai 200433,China )Abstract :AcMNPV ,the type species of baculovirus ,was used to construct a transposon based on mutant library using a Tn5random transposon vector.The green fluorescence protein gene led by Drosophila hsp70promoter was inserted into the Tn5transposon to construct a transposon that could be easily tracked in insect cell easily.In vit ro transposition was carried out using the transposon and AcMNPV genomic DNA to allow the random insertion of transposon into the virus genome.The transposed genome was then used to transfect insect cells S f21,and a library of mutant viruses capaple of ex 2pressing green fluorescence protein was obtained.Two mutant viruses ,B9F and Li6A ,was isolated ,and the transposon in 2sertion sites was determined within the coding region of 94K gene and p10gene ,respectively.This technology will be very useful in the functional genomics study of baculoviruses.K eyw ords :AcMNPV ;Tn5transposon ;transposase ;mutant205 复旦学报(自然科学版)第44卷 。
转座子插入引起的基因突变

转座子插入引起的基因突变一、原理转座子(Tn)是能在不同复制子之间转移位置的核苷酸顺序。
它一般来自抗药性质粒,由一个或几个抗药性基因加上两端两个顺序相同(但是方向不一定相同)的核苷酸片段(称为插入顺序IS)构成。
当一个转座子转移位置而插入某一基因时,能使这一基因失活,即发生突变。
各个转座子的抗药性基因、转移频率、插入位置、插入顺序都可以不同。
有些转座子对于染色体的各个位置插入没有偏向,而另一些则偏向于插入某些特定的位置。
转座子的插入突变有两上表型效应,即基因突变的表型效应和由转座子带来的抗药性。
因此利用转座子可以有效地取得任何一种突变型。
一个很有效的方法是通过转座子诱发F’因子上的染色体基因的插入突变。
大肠杆菌2-2菌株的染色体上携带有Tn5转座子(含kan抗性基因)。
要使Tn5插入到F’Lac+Pro+/Str s的乳糖发酵基因中去以引起Lac+突变,可先将这一F’因子转移到Δ(Lac Pr o)Str r受体细胞中去,再观察是否发生了插入突变。
在能排除供体的含链霉素的培养基上选择Pro+受体菌落,若是抗卡那霉素的,那就是说受体细菌不但接受了F’因子,而且这一F’因子带有Tn5,如果它又由Lac++变为Lac-,不能在以乳糖为唯一碳源的培养基上生长,那么,这一插入位置就是在乳糖发酵基因中。
为进一步提高效率,还可以采取一种措施,使在特定条件下Lac+细菌不能生存而Lac-细菌能够生存。
已经知道galE突变型在有乳糖和半乳糖存在的情况下,由于细胞中积累有毒的半乳糖代谢中间产物UDP-gal和gal-1-P而死亡,所以如果采用galE突变型作为受体细菌,在含有0.02%乳糖和0.2%甘油的基本基上,受体细菌如果接受了一个F’Lac+Pro+,就不能生存。
如果lac+基因由于插入了Tn5而成为lac+,那么受体细菌便能生存下来。
二、材料、试剂、器具1、受体菌:F-/Δ(lac proB) gaE str r2、供体菌:F’lac+pro+/Δ(lac pro)nal r带Tn5于染色体某处。
Tn5转座酶建库原理
Tn5引爆极速建库和长片段之战近10年间,NGS(Next Generation Sequencing)技术高速发展,测序仪器不断更新迭代,形成规模化。
在测序模式产业化的大环境下,测序样本制备成为其中很重要的一环。
样本起始量要求过高、建库流程繁琐等都会限制NGS技术的应用。
科学家们一直在不断探索,试图突破此类限制,其中转座酶(Transposase)技术引入建库中就是很大的进步,不仅解决了上述问题,转座酶也在NGS上开发了多个技术应用,拓展了NGS的适用范围。
20世纪40年代,美国遗传学家芭芭拉.麦克林托克在玉米的研究中发现了转座子(Transposon),随后,她花了整整6年的时间来研究转座子的奥秘。
在50年代,当麦克林托克向世人公布她的发现时,学术界并没有多少人认可,但是真理是经得起时间检验的,随后的几十年间科学家们在生物界各个领域证实了转座子系统的广泛存在。
1983年,瑞典皇家科学院诺贝尔奖金评定委员会终于把该年度的生理学和医学奖授予这位81岁高龄的、不屈不挠的女科学家。
麦克林托克是在遗传学研究领域第一位独立获得诺贝尔奖的女科学家,她的名字将和转座基因一起被载入科学史册。
转座基因俗称跳跃基因,它的发现改变了人们对基因组序列稳定性的认识。
理论上的突破也带来了应用上的拓展,科学家们群策群力,将转座子系统开发成基因研究的各种工具,这又反过来促进了遗传学理论的研究。
转座子标签(transposon tagging)技术是研究功能基因的有效工具之一。
比如模式植物大多都有很好的突变体库,其中玉米的两个应用最为广泛的突变体库(uniformMu和Ac/Ds突变体库)就是利用转座子创建的。
转座子标签技术克隆基因的基本原理:转座子是染色体上一段可移动的DNA片段,它可从染色体的一个位置跳到另一个位置。
当转座子跳跃而插入到某个功能基因时,就会引起该基因的失活,并诱导产生突变型。
通过遗传分析可确定某基因的突变是否由转座子引起,由转座子引起的突变便可以转座子DNA为探针,从突变株的基因组文库中钓出含该转座子的DNA片段,并获得含有部分突变株DNA序列的克隆,进而以该DNA为探针,筛选野生型的基因组文库,最终得到完整的基因。
转座子类型资料

二组件的关系 相同 推测相同
组件的功能 二者皆有功能 预计有功能 有功能
IS 10L
反向 有2.5%的差异 无功能
Tn5
5700
KanR
IS 50R
有功能
IS 50L
反向 1 bp的改变 无功能
复合转座子的结构特征
• (1)中间区域含编码转座酶以外的标记基因; • (2)两端具有插入序列; • (3)两末端是反向重复序列; • (4)靶位点存在短正向重复序列。
II类转座因子家族 P Hobo Mariner FB Tc1 Ac/Ds Spm/dsom Mu/mn Tam1 Tam3 TU/puppy
ITR
TSD
31
8
12
8
28
2
大
9
54
2
10/11
8
13
3
200
9
13/14
3
12
8/5
大
8
反转录病毒ห้องสมุดไป่ตู้ 含有LTR的反转录转座子 反转录转座子
非LTR的反转录转座子
BACK
=
Tn3
转座子
靶质粒
第一步 切口DNA
的
复
第四步 完成复制
制
式
转
座 第五步
Res位点两侧重组
第二步 连接自由末端 第三步 转座子复制
第六步
Conservative transposition
不供 形体 成保 共持 合裂 体缺 。。
Nonreplicative transposition
5、转座引起的遗传效应
转座子
1、转座子的基本概念 2、转座子的转座特征 3、转座子的类型 4、转座子的转座机制 5、转座引起的遗传效应
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
文章编号:042727104(2008)0320301205收稿日期:2007207213作者简介:尹 隽(1969—),女,讲师;通讯联系人钟 江,男,教授,E 2mail:jzhong@.Tn5转座子插入p95基因不影响AcMNPV 的复制尹 隽,胡志鹏,宋大新,钟 江(复旦大学生命科学学院微生物学与微生物工程系,上海200433)摘 要:从Tn5转座子介导的AcMNPV 随机插入突变体库中,分离到一株复制正常的突变体AcApra41.突变定位发现Tn5转座子插入了病毒p95基因中.为了排除AcApra41中还有其他突变,利用同源重组法构建了p95基因定点插入突变的重组病毒AcGFP 2P95in.PCR 确认p95基因中插入了Tn5转座子;Westernblot 也证实A 2cApra41和AcGFP 2P95in 感染的细胞中,P95蛋白的分子量都因为插入突变而变小,由野生型的95ku 变为55ku.病毒复制动态曲线和荧光显微镜观察证实带有该插入突变的病毒能够在Sf9细胞中正常复制,并表达极晚期基因.这一结果表明完整的P95蛋白对病毒复制是非必须的.关键词:杆状病毒;突变体;复制;基因表达中图分类号:Q812 文献标识码:A 杆状病毒(Baculoviridae )是一类主要感染节肢动物的病毒,基因组为双链环状DNA,大小约为80~180kb [1].一方面它们作为基因表达载体已被广泛地用于生产各种蛋白质,另一方面它们也可以作为生物杀虫剂使用.在所有的杆状病毒中,苜蓿银纹夜蛾核型多角体病毒(Auto gra pha cali fornica multiplenucle 2opolyhedrovirus,AcMNPV )是被研究得最为深入的一种病毒,该种病毒的基因组大小为133894b p,包括156个潜在的阅读框[2].尽管有一些基因已经被研究得很清楚,比如病毒的结构蛋白,必需的顺式作用因子等[327],但还是有许多基因的功能尚不清楚.为了能高效地研究这些未知基因,我们用Tn5转座子介导的随机插入突变法构建了AcMNPV 突变库(李惠等[8],尹隽等,待发表).本文研究了从突变体库中筛选到的一株p95基因带有插入突变的突变体.研究结果提示完整的P95蛋白对病毒在细胞中的复制是非必须的.1 材料和方法1.1 细胞、病毒和菌株草地贪夜蛾细胞系Sf9细胞用TNM 2FH 培养基(Sigma 2Aldrich,MO,USA)补充10%的小牛血清、100U/mL 青霉素和100U/mL 链霉素培养.带有AcMNPV 基因组的质粒(bacmid )来源于Bac 2to 2Bac 系统(Invitro gen,CA,USA ),保存于大肠杆菌中,它转染Sf9细胞后可以复制产生感染性的AcMNPV [9].以在病毒多角体启动子下游插入绿色荧光蛋白基因(gfp )的bacmid (bacGFP )作为起始病毒基因组,利用体外转座系统构建了AcMNPV 随机插入突变体库(尹隽,发表中). E.coli BJ5183(RecA +)作为重组用菌株,E.coli EC100(Epicentre,WI,USA)用于保存bacmid.用于培养含bacmid 菌株的抗生素浓度分别为:卡那霉素(Kan )50m g/mL,庆大霉素(Gen )7m g/mL,阿普拉霉素(Apra )20m g/mL.1.2 Tn5转座子插入定位参见李惠等[8],简单介绍如下.根据AcMNPV 基因组序列,全基因组每隔1.5kb 沿同一方向设计了88个引物(分别命名为v1~v88,具体序列略).在转座子上设计两个引物Tn 2u p (5′2GTCTCCGACCT 2第47卷 第3期2008年6月复旦学报(自然科学版)JournalofFudanUniversit y (NaturalScience )Vol.47No.3Jun.2008GATG 2CAGCTC 23′)和Tn 2down (5′2ACGACTACGCACTAGCCAACA 23′)(图1).用v1~v88分别和Tn 2up,Tn 2down 组对作为引物,以突变体bacmidDNA 作为模板进行PCR 扩增,结果v45和Tn 2down 得到阳性PCR 产物.产物割胶回收后进行DNA 序列分析(上海英俊生物技术公司),得到转座子插入位置的上游一侧的序列.在距离上游插入位点约500b p 处再设计一个引物p95down (5′2CGGCGTCGGTCGTTTGAA 23′),以p95down 和Tn 2up 作为引物再进行PCR,得到的产物同样测序,得到转座子插入位置下游一侧的序列(图1).图1 转座子插入位置及引物示意图Fig.1 Diagramofthelocusoftrans posoninsertionandsitesof primersOrf 81,Ac 2TLP ,p 95:AcMNPV 编码基因;hr3:AcMNPV 基因组同源序列3,Tn5:Tn5转座子;A pra :阿普拉霉素基因;v45,p95up,p95down,Tn 2up,Tn 2down:实验中所用引物的位置和方向.数字代表在AcMNPV 上的相对位置1.3 定向重组构建bacGFP 2P95in 重组病毒在距离p95down 上游1kb 处设计引物p95up (5′2CTGATAACGTGCATCAACCG 23′)(图1),以bacmidDNA 为模板,p95up 和p95down 为引物进行PCR 扩增,得到2.6kb 的片段,包括1.6kb 转座子序列和位于转座子序列两侧各0.5kb 的基因组序列.将该片段转化带有bacGFP 的大肠杆菌BJ5183菌株(BJ51832bacGFP ),让片段与bacGFP 发生同源重组.由于片段中的转座子序列含有Apra 抗性基因及启动子,用Kan/Gen/A pra 抗性平板筛选得到的就是同源重组后的bacmid 重组子.抽提重组bacmidDNA,以之为模板,p95up 和p95down 为引物进行PCR,并对扩增产物进行电泳检测.如果能得到2.6kb 产物,即为转座子插入p95基因的正确克隆.取同源重组bacmidDNA 电转化 E.coli EC100感受态细胞,得到的同源重组bacmid 命名为bacGFP 2P95in.抽提bacGFP 2P95inDNA,用Cellfectin (Invitro gen )转染Sf9细胞,得到重组病毒AcGFP 2P95in.同样以bacGFP 转染Sf9细胞,得到对照病毒AcGFP.1.4 重组病毒AcGFP 2P95in 的鉴定为了确定重组病毒的正确性,分别用AcGFP 和AcGFP 2P95in 病毒感染Sf9细胞(感染复数MOI=5pfu/cell ),3d 后收取上清,分别抽取病毒基因组DNA.500μL 抽提液(20%PEG80001.6mol/LNaCl )与500μL 病毒上清混合,室温放置30min 后,15000r/min 离心15min.沉淀用20μLddH 2O 溶解后,加入80μL 裂解液(10mmol/LTrispH8.0,10mmol/LEDTA,20m g/mLRNaseA,0.25%SDS,80μg 蛋白酶K ).50℃反应1h 后,用等体积酚抽提.水相加入1/10体积3mol/LNaAc 及2倍体积的无水乙醇沉淀,即得到子代病毒的基因组DNA.以此为模板,p95up 和p95down 为引物进行PCR 扩增:94℃30s,58℃30s,72℃150s,共30个循环.1.5 Westernblot 检测P95蛋白在Sf9细胞中的表达24孔板中每孔接种大约2×105个细胞,同时加入适量病毒液使MOI 约为5pfu/cell.27℃培养72h 后,将贴壁细胞吹打下来,5000r/min 离心5min,沉淀用1×PBS 漂洗后,加入40μL2×SDS 样品缓冲液,进行SDS 2PAGE (10%)电泳分析和Westernblot 检测.一抗为用原核pET 系统表达的P95蛋白自制的鼠抗P95抗体(稀释倍数为1∶500),二抗为碱性磷酸酯酶标记的羊抗兔IgG 抗体(Sigma 公司,稀释比例为1∶30000).以BCIP/NBT 为底物,进行显色反应.203 复旦学报(自然科学版)第47卷 1.6 重组病毒AcGFP 2P95in 生长曲线测定以MOI=5分别用AcGFP 和AcGFP 2P95in 感染Sf9细胞,在不同的时间取上清,按标准方法测TCID50[10].2 结 果2.1 Apra41筛选和转座子在基因组中的定位从一个Tn5介导的杆状病毒随机插入突变体库中,抽提bacmidDNA 转染Sf9细胞,从中挑选出一株复制完全正常的突变体病毒AcApra41.该突变体能在细胞中产生明显的细胞病理效应,能形成子代病毒,且其极晚期启动子能启动报道基因gfp 的表达.通过对全基因组进行PCR 扩增,确定转座子插入位置(参见李惠等[8]),发现引物v45和Tn 2down 配对进行PCR 扩增可以得到1.3kb 的阳性条带,表明转座子反向插入病毒p95基因的编码区.测序结果证实转座子上游插在p95基因编码区的69304位点.在距离69304下游约500b p 处设计引物p95down,用p95down 和Tn 2up 进行PCR,得到0.5kb 产物,测序结果表明转座子下游与病毒基因组的交界在p95基因编码区的69294位点(图1).转座子插入造成11b p 的正向重复,破坏了p95基因的编码序列.2.2 定向重组构建转座子插入p95的重组病毒为了排除AcApra41中除了p95位点转座子插入外还有其他突变的可能性,用同源重组法构建了p95位点定向突变的bacmidbacGFP 2P95in,转染Sf9细胞得到重组病毒AcGFP 2P95in.同样用bacGFP 转染Sf9细胞得到对照病毒AcGFP.抽提AcGFP 2P95in 和AcGFP 感染细胞后产生的子代病毒DNA,通过PCR 验证定向重组病毒的正确性,结果如图2所示.以p95up 和p95down 为引物进行PCR 扩增时,AcGFP 产生1.0kb 片段,而AcGFP 2P95in 产生一个2.6kb 的片段(图2),表明有1.6kb 的转座子插入.这些与预期完全一致.以p95up 和Tn 2down,及p95down 和Tn 2up 引物对进行PCR 扩增,仅AcGFP 2P95in 可以得到约0.7kb 的产物(图2),而AcGFP 没有产物,这些也与预期结果完全一致.为了进一步检验突变体病毒p95基因被破坏.用抗P95蛋白N 端的抗体检测AcGFP,AcGFP 2P95in 和AcApra41感染的Sf9细胞中P95蛋白的表达,发现AcGFP 感染的细胞中有一条95ku的条带(图3).AcGFP 2P95in 和AcApra41感染的细胞中缺少该条带,但均有一条约55ku 的条带,表明P95蛋白由于转座子的插入而截断了(图3).图2 PCR 检验定向重组病毒AcGFP 2P95inFig.2 ConfirmationofrecombinantvirusAcGFP2P95inwithtar getedinsertionofTn5图3 病毒感染Sf9细胞中P95蛋白的Westernblot 检测Fig.3 Westernblotanal ysisofP95proteininSf9cells infectedb yrecombinantviruses 2.3 AcGFP 2P95in 在Sf9细胞中的复制分别用AcGFP 和AcGFP 2P95in 感染Sf9细胞,MOI=5.测定感染5,24,48和72h 后培养上清中的病毒效价,并用荧光显微镜(Nikon,FX35A )观察感染72h 后细胞表达GFP 的情况.结果表明,AcGFP 2303 第3期尹 隽等:Tn5转座子插入p95基因不影响AcMNPV 的复制 P95in 与AcGFP 有类似的病毒生长曲线(图4),且两株病毒由极晚期多角体启动子表达外源基因的水平也相当(图5).表明转座子插入p95基因并未造成AcGFP 2P95in 病毒的复制和基因表达发生明显的变化.图4 病毒感染细胞后胞外病毒增长的动态Fig.4 ThekineticsofextracellularvirustitreincellsinfectedbyAcGFP 2P95inandAcGFP 图5 AcGFP 2P95in 和AcGFP 表达绿色荧光蛋白基因的比较(200×)Fig.5 ComparisonofGFPex pressioninvirusinfectedcells byfluorescentmicrosco py左图:亮场显微镜;右图:荧光显微镜(200×)3 讨 论本研究从Tn5转座子随机插入杆状病毒突变库中分离到一个复制正常的突变体病毒AcApra41,确定该突变体中转座子位于病毒基因组69294~69304b p 处,破坏了p95基因的完整性.为了确保重组病毒的表型是由转座子的插入引起的,应用同源重组的方法构建了定向重组病毒AcGFP 2P95in.生长曲线测定表明重组病毒的复制状况与对照病毒类似,报告基因GFP 的表达也一致,说明p95基因的完整性对病毒复制并不是必须的.AcMNPV p95基因编码一个由846个氨基酸组成的蛋白质,理论分子量为95ku.该基因的同源基因在BmNPV,O pMNPV 等多种杆状病毒中都有编码[11212].实验表明该蛋白是病毒核衣壳的结构蛋白[2,12].同源性分析表明该蛋白还带有几丁质结合域,但它们与蛋白功能的关系还不清楚.本研究得到的p95基因插入突变的突变体,插入位置位于基因中间偏后位置,插入后破坏该基因,可以编码产生一个截短型P95蛋白.用抗P95蛋白N 端序列的抗体进行Westernblot 证实P95蛋白确实由野生型的95ku 变为一个约55ku 的蛋白.试验结果表明,转座子的插入并不影响病毒的正常复制和基因表达,这有三种可能:1)P95虽然是组成病毒粒子的结构蛋白,但对病毒的结构和在细胞中的复制并不是必须的;2)P95蛋白对病毒复制是必须的,但其N 端的序列已经能够发挥作用,不需要C 端的序列;3)P95的C 端序列对病毒复制也是必须的,在感染的细胞中它也能由某未知启动子驱动表达,但由于West 2ernblot 所用的抗体是针对N 端序列的,故无法检测到.虽然本研究的结果提示前两个可能性更大,但由于C 端尚有一段1.1kb 的长序列,目前尚不能完全排除第三种可能性.有研究表明BmNPV p95基因能转录出大小不同的两个mRNA [11],预示AcMNPVP95基因也可能存在这种情况.参考文献:[1] Theilmann DA,BlissardGW,BonningB,et al .Baculoviridae[C]//Fau quetCM,Ma yoMA,ManiloffJ,et al .VirusTaxonom y 2EighthRe portoftheInternationalCommitteeonTaxonomyofViruses.NewYork:Sprin ger,2005:112921185.[2] Ayres MD,HowardSC,KuzioJ,et al .Thecom pleteDNAse quenceof Auto gra pha cali fornica nuclear poly 2hedrosisvirus[J].Virolo gy ,1994,202:5862605.403 复旦学报(自然科学版)第47卷 [3] Braunagel SC,RussellWK,Rosas2AcostaG,et al.Determinationofthe proteincom positionoftheocclusion2derived virusof Auto gra pha cali fornica nucleo polyhedrovirus[J].Proc Natl Acad Sci USA,2003,100:97972 9802.[4] Hefferon KL.BaculovirusLateEx pressionFactors[J].J Mol Microbiol Biotechnol,2004,7:892101.[5] Herniou EA,OlszewskiJA,Cor yJS,et al.The genomese quenceandevolutionofbaculoviruses[J].Ann RevEntomol,2003,48:2112234.[6] Jehle JA,BlissardGW,Bonnin gBC,et al.Ontheclassificationandnomenclatureofbaculoviruses:A proposalforrevision[J].Arch Virol,2006,151:125721266.[7] Okano K,VanarsdallAL,MikhailovVS,et al.Conservedmoleculars ystemsoftheBaculoviridae[J].Virolo2gy,2006,344:77287.[8] Li H,ZhaoML,YinJ,et al.Theconstructionand preliminar yanal ysisofaTn5trans posonbasedrandommu2tantlibrar yofbaculovirus[J].Front Biol China,2006,1:1152119.[9] Luckow VA,LeeSC,Barr yGF,et al.Efficient generationofinfectiousrecombinantbaculovirusesb ysite2spe2cifictrans poson2mediatedinsertionofforei gn genesintoabaculovirus genome propagatedin Escherichia coli[J].J Virol,1993,67:456624579.[10] O’Reilly DR,MillerLK,LuckowVA.Baculovirusex pressionvectors,alaborator ymanual[M].NewYork:W.H.FreemanandCom pany,1992.[11] Lu ML,SweversL,IatrouK.The p95GeneofBomb yxmorinuclear polyhedrosisvirus:tem poralex pressionandfunctional properties[J].J Virol,1998,72:478924797.[12] Russell RLQ,RohrmannGF.CharacterizationofP91,a proteinassociatedwithvirionsofanOr gyia pseudot2sugatabaculovirus[J].Virolo gy,1997,233:2102223.InsertionalDisru ptionofAcMNPV p95GenedidnotAffectVirusRe plicationinSf9CellsYINJuan,HUZhi2pen g,SONGDa2xin,ZHONGJian g(Department of Microbiolo gy and Microbial Engineerin g,School of Li fe Sciences,Fudan Universit y,Shan ghai200433,China)Abstract:Are plication2competentmutantvirus,AcA pra41,wasisolatedfromalibrar yofAcMNPVrandominsertional mutantsconstructedwithTn5trans posonmediatedmuta genesis.TheTn5trans posonwasdeterminedtobelocatedinthep95geneofAcMNPV genome.Toexcludethe possibilitythatAcA pra41containedothermutations,arecombinantvirus, AcGFP2P95in,whichhadtar getedinsertionofTn5in p95geneinexactl ythesamewa yasinAcA pra41,wasconstructed byhomolo gousrecombination.TheinsertionofTn5trans posonwasconfirmedwithPCR.Theinsertionoftrans posonre2 sultedinthea ppearanceofatruncatedformofP95proteinof55kuincellsinfectedwithAcGFP2P95inasshownb y Westernblotusin gantibod ya gainsttheN2terminal partofP95protein,incontrastwiththefulllen gthP95proteinof95 kuincellsinfectedwithwildt ypevirus.Virusre plicationcurveandfluorescentmicrosco picobservationindicatedthat AcGFP2P95inre plicatedandex pressedviral genesnormall yaswildt ypevirusinSf9cells.Theresultsindicatedthatafull2 lengthP95proteinwasnotessentialforvirusre plication.Ke ywords:Baculovirus;mutants;re plication;geneex pression 503 第3期尹 隽等:Tn5转座子插入p95基因不影响AcMNPV的复制 。