λ噬菌体的裂解性和溶原性的基因调控机制
最新微生物历年考研真题

微生物历年考研真题厦门大学微生物2002-2007年考研真题厦门大学2002年微生物考研试题一、填空题:(第8题 3分,其余1分/题)1.用____和____糖等成分可以制成培养真菌的半组合培养基。
2.细菌产生抗药性的3种途径分别为:染色体组上发生基因突变、____的转移和____的适应性。
3.病毒的核酸类型是及其多样的,总的来说,动物病毒以____和____居多,植物病毒以____居多,而噬菌体以____居多。
4.在实验中培养化能异养型细菌时,通常以____为碳源,____为氮源,____为生长因子,以____提供矿质元素。
5.F因子是大肠杆菌等细菌中决定____的质粒,其大小为____kb,约等于____%染色体DNA,其中1/3基因(tra区)与____有关。
6.病毒大小的单位是____,多数病毒粒子的直径在____上下。
7.电子显微镜观察表明:放线菌无性孢子的形成,只有____方式,而无____方式。
8.放线菌的形态特征,以链霉菌最典型:细胞形态呈____,营养生长期菌丝内____隔,一般呈____细胞,细胞内具有为数众多的____体:根据菌丝的结构和功能把菌丝分为____、____和____3类。
其中____能分化产生成串的分生孢子。
二、名次解释:(2分/题)1.光能自养型(photoautotroph)2.微体(microbody)3.连续培养(continuous cultivation)4.感受态因子(competence factor)5.共同抗原(common antigen)6.单细胞蛋白(single cell protein)7.紫膜(purple menbrane)8.活性污泥(activated sludge)9.类毒素(toxoid)10.Park 核苷酸( park nucleotide )11.膜边体( lomasome)12.细菌沥滤(bacterial leaching)三、问答题(没标注的3分/题,)1.龋齿的形成与某些产荚膜细菌有关吗?解释你的答案。
分子生物学名词解释

中心法则:生物体遗传信息流动途径。
现包括反转录和RNA复制等内容。
复制:是指遗传物质的传代,以母链DNA为模板合成子链DNA的过程。
转录:以DNA的一条链的一定区段为模板,按照碱基配对原则,合成一条与DNA链互补的RNA链的过程。
翻译:以mRNA为模板,氨酰-tRNA为原料直接供体,在多种蛋白质因子和酶的参与下,在核糖体上将mRNA分子上的核苷酸顺序表达为有特定氨基酸顺序的蛋白质的过程。
翻译的基本要素:tRNA、核糖体和mRNA顺式作用元件:指调控真核生物结构基因转录的DNA序列,包括启动子、上游启动子元件、增强子、加尾信号和反应元件等。
它们通过与反式作用因子相互作用来发挥转录调控作用。
反式作用因子:指真核基因的转录调节蛋白,包含DNA结合结构域和转录激活结构域。
它们与顺式作用元件、RNA聚合酶相互作用,以及转录因子之间相互协同或者拮抗,反式调控另一基因的转录。
操纵子:原核生物绝大多数基因按照功能相关性成簇串联排列,与启动子、操纵基因等调控元件共同组成一个转录单位,实现协调表达。
(原核生物中控制蛋白质合成的功能单位,包括结构基因和调控部分。
)乳糖操纵子:控制β半乳糖苷酶诱导合成的操纵子。
包括调控元件P(启动子)和O(操纵基因),以及结构基因lacZ、lacY和lacA。
在没有诱导物时,调节基因lacI编码阻遏蛋白,与操纵基因O结合后抑制结构基因转录;乳糖的存在可与阻遏蛋白结合诱导结构基因转录,以代谢乳糖。
色氨酸操纵子:控制色氨酸合成的元件之一。
大肠杆菌的色氨酸操纵子有启动子和操纵基因控制一个多顺反子mRNA的转录,控制编码色氨酸生物合成需要的各种酶,另外,还有前导区和衰减区。
当培养基中有足够的色氨酸时,操纵子关闭,,缺乏色氨酸时,操纵子开启。
诱导与阻遏:若调节蛋白和操纵基因结合后,抑制其所调控的基因转录,称阻碍物,反之诱导。
(与调节蛋白结合的效应小分子,辅诱导物)基因表达:指将来自基因的遗传信息合成功能性基因产物的过程。
λ噬菌体的基因调控
λ噬菌体的基因调控姓名学号:班级:目录λ噬菌体的发现λ噬菌体的结构组成1.基本结构2.λ噬菌体的核心λ噬菌体的生活周期I.两种发育途径简介II.调控发育途径的分子基础1.两种途径共同的早期基因表达途径2.溶源发育中基因的相互作用3.裂解途径的建立4.溶源和裂解的平衡5.溶源发育向裂解发育的转变λ噬菌体的侵染过程1.吸附2.穿入3.生物合成4.成熟与释放λ噬菌体的应用1.细菌的鉴定与分型2.耐药细菌感染的治疗3.分子生物学研究的重要工具4.遗传工程5.其他参考文献λ噬菌体的发现:1951年J. Lederberg的妻子Esther Lederberg证明了J. Lederberg和Tatum用来杂交的K12中有原噬菌体,并命名为λ,经10年的研究搞清了溶原化的实质。
在E.coli K12中是有原噬菌体的存在。
Jacob和Wollman(1956年)发现了合子诱导(zygotic induction)现象,并利用合子诱导确定了几个E.coli染色体上原噬菌体的整合位点。
他们发现Hfr(λ)×F-所得到的重组子频率要比Hfr×F-(λ)或Hfr(λ)×F-(λ)要低得多。
这是由于在Hfr(λ)×F-的杂交中,原噬菌体进入无阻遏物的受体细胞质中,进行大量复制使受体细胞裂解(图8-20b),因此不易得到重组子,此现象就称为合子诱导。
现在我们再回过头来查阅一下传递等级作图,中断杂交实验以及重组作图都是采用Hfr×F-(λ)就是不致产生合子诱导的缘故。
λ噬菌体的结构组成:1.基本结构λ噬菌体是一种温和的诱导性噬菌体,其基因组除在5'端有12个可互补的碱基外均为线性双链DNA,感染时DNA形成环状。
λ噬菌体的基因组长达50 Kb,共61个基因,其中38个较为重要。
λ-DNA的基因顺序组织如图所示,按基因组功能共分六大区域:头部编码区、尾部编码区、重组区、控制区、复制区和裂解区.它们分属四个操纵子结构:阻遏蛋白操纵子、早期左向操纵子、早期右向操纵子以及晚期右向操纵子。
噬菌体遗传分析
噬菌体的遗传分析一、噬菌体的结构:1.结构简单:蛋白质外壳、核酸、某些碳水化合物、脂肪等。
2.多样性的原因:外壳的蛋白质种类、染色体类型和结构。
3.两大类:①烈性噬菌体:T噬菌体系列(T1-T7);②温和性噬菌体: P1和λ噬菌体。
㈠、烈性噬菌体:1.结构大同小异,外貌一般呈蝌蚪状:T偶列噬菌体头部:双链DNA分子的染色体;颈部:中空的针状结构及外鞘;尾部:由基板、尾针和尾丝组成。
2.T偶列噬菌体的侵染过程(如T4噬菌体):尾丝固定于大肠杆菌,遗传物质注入破坏寄主细胞原有的遗传物质合成大量的噬菌体遗传物质和蛋白质组装许多新的子噬菌体溶菌酶裂解细菌释放出大量噬菌体。
右图为T4噬菌体侵染大肠杆菌的生活周期㈡、温和性噬菌体:例如λ和P1噬菌体,λ和P1各代表一种略有不同的溶源性类型。
1.溶源性噬菌体的生活周期:①.λ噬菌体:噬菌体侵入后,细菌不裂解附在E.coli染色体上的gal和bio位点间的attλ座位上通过交换整合到细菌染色体,并能阻止其它λ噬菌体的超数感染。
λ噬菌体特定位点的整合②P1噬菌体:不整合到细菌的染色体上,而是独立存在于细胞质内(见左下图)。
原噬菌体:整合到宿主基因组中的噬菌体。
仅少数基因活动,表达出阻碍物关闭其它基因。
原噬菌体经诱导可转变为烈性噬菌体裂解途径(见右下图)。
2.P1和λ噬菌体的特性:①P1和λ各代表不同的溶源性类型:P1噬菌体:侵入后并不整合到细菌的染色体上,独立存在于细胞质内;λ噬菌体:通过交换整合到细菌染色体上。
②溶源性细菌分裂两个子细胞:P1噬菌体复制则使每个子细胞中至少含有一个拷贝;λ噬菌体随细胞染色体复制而复制,细胞中有一个拷贝。
③共同特点:核酸既不大量复制,也不大量转录和翻译。
P1和λ噬菌体的生活周期特性二、T2噬菌体的基因重组与作图:1.噬菌体遗传性状分为两类:形成的噬菌斑形状:指噬菌斑大小、边缘清晰度、透明程度。
寄主范围:指噬菌体感染和裂解的菌株范围。
《基因工程B》复习提纲2020

第一章基因工程概述一、名词解释基因、基因操作、基因工程、重组DNA技术二、思考题1、简述基因工程操作的基本步骤。
2、简述基因工程操作的理论依据。
3、简述基因工程诞生理论上的三大发现和技术上的三大发明。
第二章基因工程的工具酶一、名词解释限制性内切酶、同裂酶、同尾酶、DNA连接酶、DNA聚合酶、逆转录酶二、思考题1、简述限制性内切酶命名的基本原则。
2、影响限制性内切酶活性的因素主要有哪些?3、影响连接反应效率的因素主要有哪些?4、大肠杆菌DNA聚合酶有哪些活性?分别简要说明其主要用途。
5、简述切口平移法标记DNA的原理。
答:首先,在Mg2+存在下利用低限量的DNase I处理双链DNA,随机产生少量切口;然后,利用大肠杆菌DNA聚合酶I的5'→3'外切酶活性在切口处向3'端平移去除核苷酸,同时,其5'→3'聚合酶活性则将切口作为引物沿5'→3'方向催化DNA合成;随着反应的进行,5'端核苷酸不断去除,3'端核苷酸不断掺入,导致切口沿着DNA合成方向移动,即切口平移。
如果在反应体系中加入标记dNTP,则这些标记dNTP将取代原有的核苷酸残基,产生带标记的DNA分子。
6、简述交换(置换)法标记DNA的原理。
答:反应体系中只有一种标记的dNTP存在时,可利用大肠杆菌DNA聚合酶I或T4/T7 DNA 聚合酶的3’→5’外切酶活性从3’端降解DNA,当露出与该标记dNTP互补的碱基时,上述酶的5'→3'聚合酶活性则催化该位置发生合成反应,用标记的dNTP置换原来的核苷酸残基,产生3’端标记的DNA。
第三章基因工程的载体一、名词解释载体、质粒、噬菌体、噬菌体溶菌周期、噬菌体溶原周期、克隆载体、表达载体、穿梭载体、整合载体、表达标签二、思考题1、简述克隆载体具备的基本条件。
2、以大肠杆菌为例,解释利用α互补筛选重组质粒的原理。
λ噬菌体溶源途径和裂解途径的基因调控

λ噬菌体溶源途径和裂解途径的基因调控摘要λ噬菌体侵染细胞后,大多数情况下进入裂解循环,λDNA复制,产生较多的噬菌体粒子。
而在以对数期以后的细菌和培养在缺乏碳源物质的培养基中的细菌作为寄主时进入溶源化途径,只有与溶源化有关的少数基因如cI才被表达。
另外溶源性细菌受到UV照射等因子诱导时,原噬菌体可以脱离细菌染色体而进行自我复制,最终导致细菌裂解,游离出大量噬菌体。
噬菌体是进入裂解循环还是整合到寄主染色体上形成溶源态,这主要取决于CI蛋白和Cro蛋白的合成及它们的调控作用。
关键词λ噬菌体,溶源化,裂解,基因调控,CⅠ蛋白,Cro蛋白一.λ噬菌体基因组和调控区λDNA分子总长度为48.5kb,编码66个基因(如下图所示),可分为三个区域:1.左臂区,自基因A到基因J,包括参与噬菌体头部蛋白质和尾部蛋白质合成所必需的全部基因。
2.中间区,介于基因J和基因N之间,这个区又称为非必需区,包含了与重组有关的基因(如基因gam)以及使噬菌体整合到大肠杆菌染色体中去的int基因和把原噬菌体从寄主染色体上切除下来的xis基因。
3.右臂区,位于N基因的右侧,包括全部主要的调控基因(cⅠ,c Ⅱ和cro),噬菌体的复制基因(O和P)以及溶菌基因(S和R)。
λ噬菌体主要的调节元件及调节基因产物的功能调节元件或调节基因产物及功能P L,O L, P R,O R左右向转录的启动子和操纵子t R(1,2,3,4,5)右向转录的终止子t L(1,2)左向转录的终止子P RE CⅠ蛋白建立启动子,受CⅡ蛋白调控P I int基因启动子,受CⅡ蛋白调控P aQ Q蛋白反义RNA启动子,受CⅡ蛋白调控P RM CⅠ蛋白基因维持启动子,受CⅠ浓度调控P R′晚期转录的启动子nut L, nut R N蛋白左右两个反终止结合位点qut Q蛋白反终止结合位点cro P L和P R的阻遏蛋白,并可阻遏P E,抑制 CI 表达c I P L和P R的主要的阻遏物,并可自主调控P RMcⅡ可以启动P RE、P I和P AQ,使λ进入溶原化途径cⅢ和CⅡ组成复合物,启动P E产生cⅠ及cro的反义RNA N t R1, t R2及t L1的反终止蛋白Q t R4的反终止蛋白.二.λ噬菌体转录调控λ噬菌体的调控有多种形式,有正调节、负调节、自主性的反馈调节、抗终止调节、反义调节及反向调节等。
λ噬菌体的裂解性和溶原性

exo和bet是位于red区域的两个基因,常被称为red基因,red为重组相关的基因,其基因产物为核酸外切酶,参与裂解生长早期所发生的重组,并能使DNA的θ型复制转变为滚环复制。Gam为大肠杆菌外切核酸酶V的抑制物基因,gam基因产物可使宿主recB和recC基因编码的外切核酸酶失活,而外切核酸酶V则降解由滚环复制所产生的线状多联体DNA。int是整合酶基因,int基因产物能识别宿主细胞染色体DNA和噬菌体基因组上相应的att位点,并能催化二者的断裂和再连接。Att位点称为附着位点,在宿主细胞DNA上称为attB,在噬菌体DNA上称为attP, attB和attP位点分别有一段由15bp组成的同源核苷酸序列,借助同源性重组作用可使噬菌体基因组插入到宿主的染色体DNA上。Xis为切除酶基因。Int和xis基因产物共同作用,能使att位点发生重组,并将原噬菌体基因组DNA从宿主染色体中切割下来,使λ噬菌体进入裂解过程。在左区还有一个cⅢ基因,它的基因产物可行使cⅡ样的功能,即激活cI和int的转录。在b2区域内的一些基因不是噬菌体存活和感染所必需的,它们包括DNase、膜蛋白基因和int基因表达的调节位点。从λ噬菌体整个基因组来看,相邻基因之间的终止位点和起始位点常发生重叠。如在ATGA序列中,TGA是前一个基因的终止密码子,而ATG则是后一个基因的起始密码子,像这样的重叠结构,λ噬菌体基因组中就有30个,整个基因组很少有非编码区。
表1 λ噬菌体主要的调节元件及调节基因产物的功能
调节元件或
调节基因
产物及功能
PL,OL,PR,OR
植物基因工程中的λ噬菌体载体
中段:从J到N长约 20kb,是λDNA整合 和切出,溶原生长所 需的序列 ,但非溶菌 生长所必需
右臂:长约10kb,控 制溶菌和溶原生长最重 要的调控基因和序列、 以及λDNA复制起始均 在这区域内。
➢ 基因组长约50kb ,至少包括61个基因,除少数例外,大多数编码基因均是按功噬菌体结构蛋白合成、组装成熟噬菌体、溶菌生 长所需全部序列;对溶菌生长来说,中段是非必需的。
(4)重组噬菌体的体外包装,形成有感染力 的噬菌体颗粒
• 利用特殊材料,制备噬菌体包装蛋白
• 连接产物与包装蛋白混合时,就可完成包 装反应,形成有感染力的噬菌体颗粒
• 包装蛋白对所包装的DNA大小有高度选择 性, 范围:λDNA分子的75%-105%
λ噬菌体的改造
• 设计去除λDNA上的多余序列和一些限制性酶切点:因为 λDNA较大,序列中的限制性酶切点过多,妨碍其应用。
•
相信相信得力量。20.12.272020年12月 27日星 期日4时26分33秒20.12.27
谢谢大家!
•
生活中的辛苦阻挠不了我对生活的热 爱。20.12.2720.12.27Sunday, December 27, 2020
•
人生得意须尽欢,莫使金樽空对月。04:26:3304:26:3304:2612/27/2020 4:26:33 AM
植物基因工程中的 λ噬菌体载体
2013年6月29日
载体(vector)是把一个有用的目的DNA片段通过重 组DNA技术,送进受体细胞中去进行繁殖和表达的工 具。 植物基因工程技术中尤为重要的是载体,不同目的 基因需要采用不同的载体。 组成植物基因工程载体系统常见的几种载体有:
质粒 λ噬菌体 柯斯质粒(cosmid) m13单链噬菌体。
λ噬菌体的裂解性和溶原性

生命科学学院λ噬菌体的裂解性和溶原性的基因调控机制姓名:学号:班级:专业:摘要λ噬菌体(phage)有两种生存策略,一种通过感染宿主细胞,产生大量的子代噬菌体,同时宿主细胞裂解死亡,这种方式称为裂解性感染。
另一种是噬菌体的基因组以一种原噬菌体的方式潜伏于细菌中,这种增值方式称为溶原态(lysogeny)。
λ噬菌体的裂解发育、溶原发育和溶原发育到裂解发育的诱导是研究生物分子调节优异的模型。
经过四十多年的研究,在这个模型中已经发现了众多的正调节因子和负调节因子在转录水平或转录后调节基因的表达。
关键词:λ噬菌体、裂解性、溶原性目录1.摘要 (2)2.λ噬菌体的结构组成 (3)2.1壳体结构 (3)2.2λ噬菌体的核心 (3)3.λ噬菌体的生活周期 (6)3.1 λ噬菌体DNA复制 (6)3.2λ噬菌体的转录调控 (7)3.3λ噬菌体的溶原性感染 (9)3.3.1λ噬菌体溶原化状态的建立 (9)3.3.2λ噬菌体基因组的整合 (11)3.3.3原噬菌体的割离 (12)3.3.4裂解性-溶原性选择决定 (15)4.参考文献 (16)1951年Esther Lederberg 发现E.coli K12菌株经UV诱发或偶尔自发放出噬菌体。
E.coli K12中有潜伏的、无感染能力状态的噬菌体,称原噬菌体。
将这种噬菌体命名为λ。
λ侵染E.coli后可进入裂解周期(lytic cycle)或溶原周期(lysogeny)。
2.λ噬菌体的结构组成2.1壳体结构λ噬菌体是有尾噬菌体,壳体由头部和尾部组成,头部和尾部通过颈部相连。
头部通常呈二十面体对称,直径为60nm左右;尾部呈螺旋对称,无收缩性。
λ噬菌体头部蛋白主要有gpE(38kD)和gpD(12kD),他们以非二硫键进行共价连接。
2.2λ噬菌体的核心λ噬菌体核心包含线状dsDNA,分子量为30.8MD,含有48502bp,其双链DNA的两5′端叫做m端, 末端碱基为G,为左向或反时针方向转录的链。
λ噬菌体的综述

生命科学学院病毒生物学λ噬菌体综述摘要:噬菌体是一类温和噬菌体,它们感染大肠杆菌后能进行溶菌性生长(Lytic growth)和溶源性生长(Lysogenic growth)。
其溶源特性对基因重组与遗传工程研究有很大帮助。
本文就λ噬菌体的基因组结构、溶关键词:λ噬菌体溶原性溶菌性基因克隆引言:大肠杆菌噬菌体λ为长尾噬菌体科,是一类中等大小的大肠杆菌病毒,其基因组为双链线状DNA,由48502对碱基组成,分子量3.2×107 ,约50个基因,特点是相关基因成簇排列,形成若干个操纵子。
基因组两端为粘性末端,中间有相当长的DNA片段是裂解生长非必需的,这就为其作为外源基因的克隆载体提供了方便。
λ噬菌体由头和尾构成,其基因组组装在头部蛋白质外壳内部,其序列已被全部测出。
感染时吸附位点为细胞壁。
属温和性感染;感染的DNA环化并整合于宿主基因组中。
以θ环双向复制,然后通过滚环机制单向复制。
用于感染大肠杆菌的λ噬菌体改造成的载体应用最为广泛。
1、The discovery of bacteriophage lambda1951年J. Lederberg的妻子Esther Lederberg第一个证明了 J. Lederberg和Tatum用来杂交的K-12中有原噬菌体,并命名为λ,经10年的研究搞清了溶原化的实质。
从此之后,λ噬菌体被广泛用于模式物种;1962年Esther Lederberg的同事并且还是她最好的朋友Allan Campbell首次发现了λDNA整合到细菌DN A的机制;之后由λ噬菌体改造后的载体广泛的用于基因工程。
2、The characteristics of bacteriophage lambda2.1、结构特点:λ为大肠杆菌温和性噬菌体,属长尾噬菌体科,头壳为直径约50nm的二十面体,其内包裹一长线状双链DNA分子(46500bp),因分子两端各有一含12个核苷酸的黏性末端,故又可黏合成环状分子。
lambda 噬菌体复制机制

Lambda噬菌体是一种可以感染大肠杆菌等细菌的噬菌体,它具有特殊的复制机制。
本文将从多个方面对Lambda噬菌体复制机制进行深入探讨。
一、Lambda噬菌体的结构Lambda噬菌体是一种双链DNA噬菌体,其直径约为55纳米。
其基因组主要由50多个基因组成,包含有富含AT的过渡核心区、左右两个对称的延伸区和一些非编码区。
Lambda噬菌体的基因组中有些基因是干扰细胞代谢的基因,有些基因编码了噬菌体颗粒结构蛋白,还有些编码了其遗传物质的复制和转录的相关酶。
二、噬菌体感染细胞的过程Lambda噬菌体感染细胞的过程大致可以分为吸附、注入、复制与释放四个阶段。
1. 吸附阶段当Lambda噬菌体接触到细菌表面时,其尾部纤毛可以选择性的吸附到细菌表面的特定受体上。
这一过程是特异性的,每一种噬菌体都有其特异性的受体。
2. 注入阶段一旦吸附到细菌表面后,Lambda噬菌体尾部会释放出一种酶,这种酶可以溶解细菌表面的细菌壁。
随后,噬菌体的DNA会由尾部注入细3. 复制阶段一旦Lambda噬菌体的DNA进入细菌细胞内,它就会利用细菌细胞内的机制开始复制。
Lambda噬菌体的复制方式有两种,一种是以自身的DNA为模板,利用细菌细胞内的酶和蛋白质进行复制;另一种是将自身的DNA整合到宿主细胞的染色体上,然后进行复制。
这两种方式取决于噬菌体的DNA是以原始圆形存在还是整合到宿主细胞染色体上。
4. 释放阶段当Lambda噬菌体的DNA在细菌内复制完成后,新的噬菌体颗粒会在细菌内自我组装,并最终将细菌破裂释放出去。
三、Lambda噬菌体的复制机制Lambda噬菌体的复制机制主要包括DNA复制和转录。
1. DNA复制Lambda噬菌体的DNA复制主要由噬菌体DNA聚合酶进行。
在细菌细胞内,Lambda噬菌体DNA形成一个线性的大环,其两端相连接,被称为原体。
当原体线性化后,就会形成一条线性的DNA。
此时,DNA聚合酶会在噬菌体的起始点(O点)和P点附近启动复制。
噬菌体裂解液的制备和菌种的保藏、噬菌体效价的测定、细菌转导测定、凝聚反应、沉淀反应

2.取0.5mL裂解液,37C预热10分钟
3.把上述S菌和裂解液混合均匀,37C预热10分钟
4.用牛液把混合液稀释至10-3及10-4
5.取10-3及10-4稀释度0.2mL涂布EMB平板,于37C培养至少48hr,观察结果
3、凝集反应
按图加抗原和抗体及对照,静置5~10min,先用肉眼观察,后用显微镜在低倍镜下观察。
2.按右图所示在皿底用记号笔画好标记。
3.接种无菌操作,取矢量培养活化的K12 S gal-菌一环,涂于S标记处;取适量活化的K12 F gal+菌液,划线接种于F标记处;取适量噬菌体裂解液,涂于λ标记处。
4.将接好的平板静置,所有菌液被培养皿吸收后再倒置,放37℃培养48~72h,观察结果。
(2)转导频率的测定
在光下观察,4个培养皿中均未发现深紫色带有金属光泽的菌落存在。
三、凝聚反应
右侧为对照组,滴加磷酸盐缓冲液(代替生理盐水)与E. coli,无明显反应。
左侧为实验组,滴加抗血清与E. coli,在室温下静置数分钟后形成肉眼可见的凝集块。
四、沉淀反应
两个载玻片菌没有观察到明显现象,没有白色沉淀线生成。
注:理想条件下,应出现如下图所示现象。
5)培养
待平板凝固后于37 ℃,培养12~16h;
根据噬菌斑数量计算噬菌体效价。
(因所测噬菌体裂解液中λdg不能形成噬菌斑,且其数量与λ数相等,故计算时乘2.)
2、细菌转导的测定
(1)点滴法
1.倒EMB平板取已灭菌的EMB培养基熔化并冷却至不烫手(约50℃),然后倒平板,冷却后翻转平板,温箱放置使表面干燥。
4、沉淀反应
将琼脂糖凝胶融化后滴加到干净的载玻片上,凝固后按下图位置打孔并在孔中滴加相应的抗原和抗体。
分子克隆简答题

简述基因克隆的两个基本特征?基因克隆的两个基本特征是一是强调外源核酸分子(一般情况下都是 DNA)在不同宿主中的繁殖,打破自然种的界限将来自于不相关物种的基因放入一个宿主中是基因操作的一个重要特征,基因操作的另一个重要特征是繁殖。
如何理解 gene 及其产物的共线性和非共线性?基因决定蛋白质的序列组成,是由密码子对应特定氨基酸所决定的。
当一个基因的核苷酸序列与其产物的氨基酸序列是一一对应时,则表明它们是共线性的。
在原核生物中,基因及其产物是共线性的。
70 年代以来,在真核生物中,发现了间断基因,后来发现这种间断基因在真核生物中普遍存在,也就是后来所说的基因间存在着内含子。
内含子指真核生物基因中不能被翻译成蛋白质的 DNA 片断,但可被转录,当两侧序列的转录 RNA 被剪接在一起时,就将内含子转录的 RNA 从整个转录物中除去。
外显子是指能够翻译成蛋白质的任一间断的基因片段,一个基因可有多个外显子。
内含子并不是一成不变的,具有相对性,对一个 DNA 片断来说,在某个基因中是内含子,但在另一个基因中却可以作为外显子。
什么是 gene cloning ?什么是亚克隆(subcloning)?基因克隆:一定程度上等同于基因的分离,即从复杂的生物体基因组中,经过酶切,消化等步骤,分离带有目的基因的 DNA 片段。
基因亚克隆:初步克隆中的外源片段往往较长,含有许多目的基因片段以外的 DNA 片段,在诸如表达、序列分析和突变等操作中不便进行,因此必须将目的基因所对应的一小段 DNA 找出来,这个过程叫“亚克隆”。
假如从一个原核生物中 cloning 某基因(1-2kb),请谈谈它的基本步骤?①对原核生物染色体 DNA 进行不完全酶切,纯化所得到的 DNA 片断,并与载体连接,转化大肠杆菌;②得到转化子后,根据目标基因两侧的已知序列设计探针,杂交,筛选转化子;③所得到的转化子中含有目标基因,进一步作亚克隆,一步步地向目标基因逼近,最后可得到目标基因什么叫基因工程(Gene Engineering),试从理论和技术两个方面谈谈 Gene Engineering 诞生的基础?基因工程(Gene Engineering )又称基因操作(Gene Manipulation)、重组 DNA 。
e42-2λ噬菌体基因表达的时序性调控
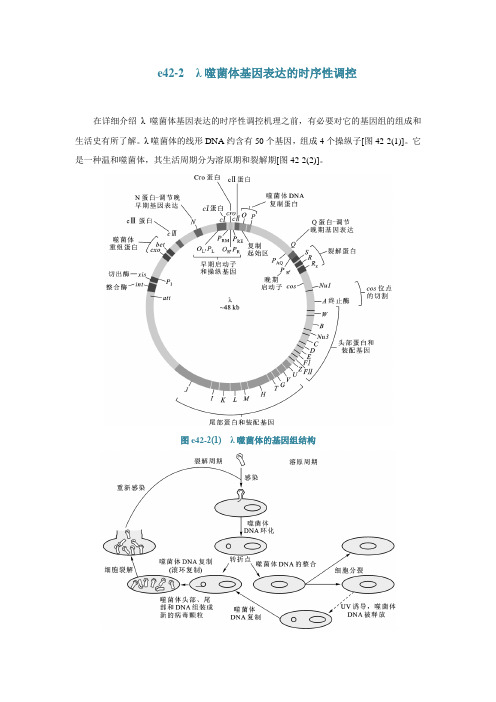
e42-2 λ噬菌体基因表达的时序性调控在详细介绍λ噬菌体基因表达的时序性调控机理之前,有必要对它的基因组的组成和生活史有所了解。
λ噬菌体的线形DNA约含有50个基因,组成4个操纵子[图42-2(1)]。
它是一种温和噬菌体,其生活周期分为溶原期和裂解期[图42-2(2)]。
图e42-2(1) λ噬菌体的基因组结构图e42-2(2) λ噬菌体的生活周期在不同的时期,λ噬菌体基因组表达的基因差别很大[图42-2(3)]。
如果进入溶原期,噬菌体DNA首先环化,前早期基因表达Cro蛋白和N蛋白。
晚早期基因表达整合酶和CI阻遏蛋白,其中CI阻止晚期基因表达,整合酶催化λ-DNA通过位点特异性重组整合到宿主DNA上(参看第三十五章DNA重组),以原噬菌体的形式存在,并随着宿主DNA的复制而复制,但不会对宿主细胞造成什么严重的损害。
在这期间,只有CI蛋白表达。
需要强调的是,整合并非是进入溶原状态必需的条件,因为某些λ噬菌体的变体虽然不能进行整合,但也能以原噬菌体的形式存在,这种情况下的λ-DNA仿佛质粒DNA一样,游离在宿主染色体DNA之外。
如果在溶原期,宿主细胞出现饥饿、中毒(如抗生素)或DNA损伤等生存压力,原噬菌体就会被激活,从溶原期转变成裂解期。
在裂解期,也是噬菌体DNA首先环化,前早期基因表达,然后是晚早期基因表达。
这时DNA发生双向θ复制。
最后是晚期基因表达,DNA通过滚环复制大量扩增,新的病毒颗粒装配,宿主细胞裂解。
图e42-2(3) λ噬菌体生活周期不同阶段的基因表达(一)前早期的基因表达在噬菌体感染宿主细胞以后,转录从左右两个主要的启动子(P L和P R)向两个方向展开。
这两个启动子可以被大肠杆菌的RNA聚合酶有效地识别,并分别终止于左右两个终止子(t L1和t R1)。
转录的终止依赖于ρ因子,转录的产物经翻译分别产生Cro蛋白和N蛋白。
Cro也是一种阻遏蛋白,它能与cI蛋白之间竞争结合操纵基因。
λ噬菌体载体大学生物学

EcoR I
cos
cI
cos
λgt10(4.3kb),可插入外源 DNA大小为0-6kb,插入使cI基因 失活,可通过免疫学方法筛选
λ噬菌体载体的主要类型
λgt11载体为大肠杆菌β半乳糖苷酶插入失活载体的代表
EcoR I Stop code
cos
lacZ
cos
λgt11(4.37kb),可插入外源DNA 大小为0-7.2kb,插入使lacZ基因失 活,可通过组织化学的方法筛选
EcoRⅠ BamHⅠ SalⅠ
中间可替代区(14kb)右臂(9kb)
EcoRⅠ BamHⅠ SalⅠ
SalⅠ BamHⅠ EcoRⅠ
λ噬菌体载体的优点
lλ-噬菌体载体在体外包装成噬菌体颗粒后,可以高 效转染大肠杆菌,感染宿主细胞的效率几乎可达 100%,而质粒DNA的转化率只有千分之一
lλ-DNA 载体的装载上限为23kb,远远大于质粒的装 载量
λ噬菌体载体的主要类型
l取代型载体具有成对的克隆位点,在这两个位点 之间的λDNA区段可以被外源插入的DNA片段所置 换。
λ噬菌体载体的主要类型
以λEMBL3和 λEMBL4为代表。
EMBL3载体 左臂(20kb)
中间可替代区(14kb)右臂(9kb)
EMBL4载体 左臂(20kb)
SalⅠ BamHⅠ EcoRⅠ
λ
λ噬菌体的生物学特性
l 噬菌噬菌体,在感染 宿主后可进入溶原状态,也可进入裂解循 环。
l λ 噬菌体结构简单,仅由外壳包装蛋白和 λ-DNA组成
l λ噬菌体基因组DNA全长48502个核苷酸, 可以编码至少30个基因
λ-DNA的结构特点
l λ噬菌体基因组DNA全长48502个核苷酸,
分子生物学_南京大学中国大学mooc课后章节答案期末考试题库2023年

分子生物学_南京大学中国大学mooc课后章节答案期末考试题库2023年1.下列蛋白质合成抑制剂中,既能抑制原核生物蛋白质合成、又能抑制真核生物蛋白质合成的是:参考答案:嘌呤霉素2.如果大肠杆菌突变后,DNA连接酶的表达量提高5倍,那么快速标记新合成的DNA后分离冈琦片段,会发现参考答案:冈琦片段比未突变的菌株长3.下列关于原核生物转座子的说法,正确的是参考答案:都可以编码转座酶4.下列有关内蛋白子的说法,错误的是:参考答案:迄今发现的内蛋白子仅存在于原核生物中5.下列关于原核生物RNA转录起始的叙述,错误的是参考答案:全酶和核心酶在DNA上的结合位点是相同的6.Holliday模型中主要步骤的正确顺序是参考答案:单链断裂-形成Holliday中间体-分支迁移-Holliday中间体的拆分7.下列有关真核生物mRNA前体加帽修饰的叙述,错误的是参考答案:加帽过程发生在转录完成之后8.下列有关I型内含子和II型内含子的叙述,错误的是参考答案:拼接过程都需要鸟苷或鸟苷酸9.下列有关跨越合成的说法,错误的是参考答案:跨越合成总是导致DNA突变10.在下列哪种培养基条件下,大肠杆菌乳糖操纵子的表达水平最高参考答案:低葡萄糖、高乳糖11.下列哪一种生物中没有光复活机制参考答案:胎盘类哺乳动物12.下列有关真核生物mRNA前体3’加尾过程的叙述,正确的是参考答案:细胞核与细胞质中都有poly(A)聚合酶13.下列哪一种蛋白质与λ噬菌体溶原状态的建立与维持密切相关?参考答案:CI蛋白14.为大肠杆菌DNA连接酶的作用提供能量的是参考答案:NAD+15.真核生物的翻译起始因子eIF4F实际上是由三种亚基组成的复合物,其中具有帽子结合活性的亚基是参考答案:eIF4E16.下列有关原核生物和真核生物肽链延伸机制的叙述,错误的是参考答案:都需要延伸因子EF-Tu和EF-G17.原核生物基因表达和调控的单位称为参考答案:操纵子18.下列有关人类基因组计划的说法,正确的是参考答案:根据基因组计划的测序结果,人与小鼠约有99%的基因是相同的19.克隆羊多莉的基因组与其它同龄的羊相比参考答案:端粒比与其它同龄羊的短20.所谓核开关是指下列哪种物质形成的调节基因表达的结构?参考答案:mRNA21.λ噬菌体是一种以大肠杆菌为宿主的参考答案:温和噬菌体22.下列关于真核生物转座子的说法,正确的是参考答案:玉米中的Ac-Ds系统属于保留型DNA转座子23.下列因素中,影响翻译正确起始的是参考答案:SD序列与反SD序列的相互作用24.λ噬菌体晚早期基因的表达主要依赖于下列哪一种蛋白质的调控?参考答案:N蛋白25.下列有关乳糖操纵子的说法正确的是参考答案:乳糖操纵子中既有正调控又有负调控26.下列有关选择性拼接的叙述,错误的是参考答案:大多数真核生物mRNA前体存在多种拼接方式27.下列有关EF-Tu和EF-G的叙述,错误的是参考答案:都可以与氨酰tRNA结合28.端粒酶是一种蛋白质-RNA复合物,其中RNA的作用是参考答案:作为端粒合成的模板29.下列有关氨酰tRNA合成酶的叙述,错误的是参考答案:大多数生物体中有20种结构功能相近、高度同源的氨酰tRNA合成酶30.原核生物利用tmRNA进行反式翻译应对参考答案:non-stop mRNA31.下列有关真核生物mRNA前体拼接机制的叙述,正确的是参考答案:真核生物mRNA前体可能存在多种拼接方式32.染色质折叠的正确顺序是参考答案:形成核小体,形成30 nm纤维,放射环结构33.下列各项中对DNaseI最敏感的是参考答案:“裸露”的DNA,即未与组蛋白结合的DNA34.由核基因编码的线粒体蛋白的合成部位是:参考答案:细胞质的游离核糖体35.水解酶由高尔基体进入溶酶体的信号是:参考答案:N寡糖链上具有甘露糖-6-磷酸36.蛋白质可以通过下列哪种修饰锚定在膜上?参考答案:脂基化37.蛋白质糖基化发生的主要部位是参考答案:内质网和高尔基体38.对于大多数基因来说,CpG序列的甲基化参考答案:抑制基因转录39.错配修复机制的特征是参考答案:低效率、高耗能;40.下列有关真核生物RNA聚合酶抑制剂的说法,错误的是参考答案:RNA聚合酶III对放线菌素D最敏感41.下列有关原核生物RNA转录后加工的说法,正确的是参考答案:mRNA一般不进行转录后加工42.RecA是一种多功能蛋白,除了具有ATPase活性外,还具有下列哪种酶活性?参考答案:辅蛋白酶43.下列突变类型中,不可能由转座作用引起的是参考答案:单个碱基的突变44.下列有关锌指结构的叙述,错误的是参考答案:具有相同的氨基酸组成45.转录激活因子可以与下列哪一种顺式作用元件结合参考答案:增强子46.下列有关增强子的叙述,错误的是参考答案:有严格的组织、细胞和基因特异性47.选择性拼接是真核生物中常见的基因表达调控方式,下列选项中不可能发生的是参考答案:改变外显子的连接顺序48.RNA干扰属于下列哪一种水平上的基因表达调控参考答案:翻译水平49.糖皮质激素受体可以通过下列哪一步骤被活化参考答案:与激素配体结合并转移到细胞核内50.下列有关组蛋白乙酰化的叙述,错误的是参考答案:组蛋白的乙酰化只能在细胞质中进行51.下列有关玉米非自主型转座子的叙述,正确的是参考答案:只有当基因组同时含有属于同一家族的自主型转座子时,转座才可以发生52.大肠杆菌依靠甲基化程度的不同区分亲本链和新合成的子链,下列说法中正确的是参考答案:如果两条链都被甲基化,则错配修复效率极低,即使修复也很容易造成突变53.下列关于Mu噬菌体的说法,错误的是参考答案:两端有重复序列54.下列有关断裂基因的叙述,错误的是参考答案:内含子序列都比外显子序列短55.直接修复机制的特征是参考答案:高效率、高耗能56.同源重组和位点特异性重组的共性是参考答案:都有利于遗传物质多样性的积累57.Tn10转座到一个新的DNA靶点时,在靶点两侧会参考答案:形成正向重复序列58.下列转座子中不属于逆转座子的是参考答案:果蝇的P元件59.大多数真核生物中的同源重组的机制可以用下列哪一种模型解释?参考答案:双链断裂模型60.转录起始过程的正确顺序是:参考答案:闭合启动子复合物、开放启动子复合物、结合最初几个核苷酸、启动子清除61.下列有关线粒体翻译体系的叙述,正确的是参考答案:存在更宽松的摆动规则62.摆动学说认为密码子与反密码子相互配对时,配对有一定自由度的碱基对是参考答案:密码子的第3位与反密码子的第1位63.在三种终止密码子中,使用频率最高的是参考答案:UAA64.不依赖序列的特异性,只依赖于序列的同源性的DNA重组方式是参考答案:同源重组65.下列有关重组修复的说法,错误的是参考答案:属于倾向差错的复制机制66.下列关于逆转录病毒的说法,正确的是参考答案:逆转录酶又称RDDPase,一般性质与DDDPase相似67.下列关于真核生物转录起始的叙述,错误的是参考答案:三种RNA聚合酶使用不同的σ因子识别启动子68.下列哪一项不是DNA复制起点的特征?参考答案:具有回文结构69.下列有关端粒酶的说法,错误的是参考答案:体细胞中端粒酶活性较高70.下列关于原核生物转录终止的叙述,错误的是参考答案:原核生物中有两种不同的终止子,但作用机制相同71.真核生物特有的tRNA转录后加工方式是参考答案:添加CCA72.下列原核生物核酸内切酶中,可以被RNase处理失活的是参考答案:RNase P73.真核生物mRNA的帽子结构中的磷酸二酯键是参考答案:5’-5’磷酸二酯键74.大肠杆菌中最主要的同源重组途径是参考答案:RecBCD途径75.如果溶酶体水解酶蛋白缺乏甘露糖残基,那么该水解酶将:参考答案:分泌到胞外76.下列有关tRNA内含子拼接的叙述,错误的是参考答案:拼接过程属于自我催化77.λ噬菌体DNA的复制方式为参考答案:滚环复制78.宿主细胞的状态决定λ噬菌体的命运。
分子生物学课件λ噬菌体基因表达的调控

OR3
CI
O R2
PR
CRO
O R1
P RM
G/C as axis IR seq of 17 bp
OL3
O L2
O L1
CI
OR1, OR2, OR3 序列的差异, 决定了CI-p, CRO-p 与其的结合力
For CI-p (repressor) For CRO-p (repressor)
OR1 > OR2 = OR3 OR1 = OR2 < OR3
P RE
Cis- factor
Trans-factor
F的基因组成
C gene (与溶原直接相关)
λ ? host ? lysogenic ? tuibid colony C gene mut. ? lytic ? Clear plaque
P RE
Early
N-P Anti-termination
Delayed early stage
Anti-termination in Rho-dependent terminator
TR1; Rho-dependent terminator 8s Cro RNA
Np
NuTR
Cro
CII
16s RNA
Immediate early stage; N & Cro genes transcription from P L & P R
Delayed early stage; N-p required for anti-termination at T L & T R
Late stage; Q-p required
λ噬菌体基因表达的调控
2006
烈性噬菌体( lytic phages ) 温和噬菌体( temperate phages )
λ噬菌体的裂解性和溶原性

生命科学学院λ噬菌体的裂解性和溶原性的基因调控机制XX:学号:班级:专业:摘要λ噬菌体(phage)有两种生存策略,一种通过感染宿主细胞,产生大量的子代噬菌体,同时宿主细胞裂解死亡,这种方式称为裂解性感染。
另一种是噬菌体的基因组以一种原噬菌体的方式潜伏于细菌中,这种增值方式称为溶原态(lysogeny)。
λ噬菌体的裂解发育、溶原发育和溶原发育到裂解发育的诱导是研究生物分子调节优异的模型。
经过四十多年的研究,在这个模型中已经发现了众多的正调节因子和负调节因子在转录水平或转录后调节基因的表达。
关键词:λ噬菌体、裂解性、溶原性目录1.摘要 (2)2.λ噬菌体的结构组成 (3)2.1壳体结构 (3)2.2λ噬菌体的核心 (3)3.λ噬菌体的生活周期 (6)3.1 λ噬菌体DNA复制 (6)3.2λ噬菌体的转录调控 (7)3.3λ噬菌体的溶原性感染 (9)3.3.1λ噬菌体溶原化状态的建立 (9)3.3.2λ噬菌体基因组的整合 (11)3.3.3原噬菌体的割离 (12)3.3.4裂解性-溶原性选择决定 (15)4.参考文献 (16)1951年Esther Lederberg 发现E.coli K12菌株经UV诱发或偶尔自发放出噬菌体。
E.coli K12中有潜伏的、无感染能力状态的噬菌体,称原噬菌体。
将这种噬菌体命名为λ。
λ侵染E.coli后可进入裂解周期(lytic cycle)或溶原周期(lysogeny)。
2.λ噬菌体的结构组成2.1壳体结构λ噬菌体是有尾噬菌体,壳体由头部和尾部组成,头部和尾部通过颈部相连。
头部通常呈二十面体对称,直径为60nm左右;尾部呈螺旋对称,无收缩性。
λ噬菌体头部蛋白主要有gpE(38kD)和gpD(12kD),他们以非二硫键进行共价连接。
2.2λ噬菌体的核心λ噬菌体核心包含线状dsDNA,分子量为30.8MD,含有48502bp,其双链DNA的两5′端叫做m端,末端碱基为G,为左向或反时针方向转录的链。
3.4 温和噬菌体与溶源性

温和噬菌体与溶源性温和噬菌体和溶源性温和噬菌体(lysogenic phage):噬菌体感染宿主细胞后,它们的核酸和寄主细胞的染色体同步复制,并随着寄主细胞的生长繁殖而传递下去,寄主细胞不裂解,这种噬菌体被称为温和噬菌体或溶源性噬菌体。
温和噬菌体进入的这种状态被称为溶源性(lysogeny)。
含有温和噬菌体的宿主细胞叫溶源细菌或溶源细胞。
原噬菌体:附着或融合在溶源细胞染色体上的温和噬菌体的核酸被称为原噬菌体或前噬菌体(Prophage)。
λ噬菌体的溶源性反应:进入宿主后线状基因组依靠粘性末端环化λ噬菌体的DNAλ噬菌体的裂解循环和溶源循环Phage DNA circularizesorPhage溶源细菌正常复制,原噬菌体被传递给子代细菌裂 解循 环细菌染色体噬菌体吸附、侵入噬菌体DNA 环化、进入循环 细胞裂解,噬菌体释放新的噬菌体复制、组装噬菌体DNA 整合到细菌染色体中成为原噬菌体原噬菌体噬菌体DNA偶尔,原噬菌体从细菌染色体上脱落下来从新进入循环溶源循环原噬菌体随着细菌的分裂传递下去Integration of lambdaDNA into the host Lambda DNA integrates at specific attachment (att) sites on both the host and phage genomes. Host genes near att include gal, galactose utilizat -ion; bio, biotin synthesis. Lambda integrase is required, and specific pairing of the complementary ends results in integration of lambda DNARegulation of lytic and lysogenic events in phage lambdacI ,Cro ,溶源循环cI ,Cro ,裂解循环1、可自发裂解溶原细菌绝大多数不发生裂解,只有极少数溶原细菌中的原噬菌体从寄主细胞的DNA上脱离,进行大量复制,并接着成熟为噬菌体粒子,导致宿主细胞裂解,释放成熟的噬菌体粒子,这种现象称为溶原细菌的自发裂解。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
λ噬菌体的裂解性和溶原性的基因调控机制摘要:λ噬菌体(phage)有两种生存策略,一种通过感染宿主细胞,产生大量的子代噬菌体,同时宿主细胞裂解死亡,这种方式称为裂解性感染。
另一种是噬菌体的基因组以一种原噬菌体的方式潜伏于细菌中,这种增值方式称为溶原态(lysogeny)。
λ噬菌体的裂解发育、溶原发育和溶原发育到裂解发育的诱导是研究生物分子调节优异的模型。
经过四十多年的研究,在这个模型中已经发现了众多的正调节因子和负调节因子在转录水平或转录后调节基因的表达。
关键词:λ噬菌体、裂解性、溶原性1951年J. Lederberg的妻子Esther Lederberg证明了J. Lederberg和Tatum用来杂交的K12中有原噬菌体,并命名为λ,经10年的研究搞清了溶原化的实质。
λ噬菌体的基因组长达50 Kb,共61个基因,其中38个较为重要。
其生活史如图8-15所示,可分为裂解周期和溶原周期。
细菌处于溶原化状态时,细胞质中有一些λ CⅠ基因的产物CⅠ蛋白,这是一种阻遏蛋白,可以阻止λ左、右两个早期起动子的转录,使之不能产生一些复制及细胞裂解的蛋白。
λ的DNA随着宿主的染色体复制而复制。
但在UV诱导下Rec蛋白可降解CⅠ蛋白(见第17章),诱导90%的细胞裂解。
有时λ也可自发地(10-5)从宿主的染色体上游离出来,进行复制,最终导致宿主细胞的裂解,此称为治愈(curing)。
游离在细胞质中的λ可以进行滚环复制,产生多个拷贝,并合成头部和尾部蛋白,包装成完整的λ噬菌体,使细胞裂解,释放出λ噬菌体再感染新的细胞。
(图8-19)。
因为λ噬菌体的DNA也有整合在染色体上和游离于细胞质中两种状态,所以也称做附加体。
但和F因子不同,λ噬菌体有细胞外形式,而F因子无细胞外形式。
在E.coli K12中是有原噬菌体的存在。
Jacob和Wollman(1956年)发现了合子诱导(zygotic induction)现象,并利用合子诱导确定了几个E.coli染色体上原噬菌体的整合位点。
他们发现Hfr(λ)×F-所得到的重组子频率要比Hfr×F-(λ)或Hfr(λ)×F-(λ)要低得多。
这是由于在Hfr(λ)×F-的杂交中,原噬菌体进入无阻遏物的受体细胞质中,进行大量复制使受体细胞裂解(图8-20b),因此不易得到重组子,此现象就称为合子诱导。
现在我们再回过头来查阅一下传递等级作图,中断杂交实验以及重组作图都是采用Hfr×F-(λ)就是不致产生合子诱导的缘故。
裂解发育噬菌体的感染周期可以分为早期(复制前)和晚期(复制后)两个阶段。
噬菌体基因组复制和产生蛋白质颗粒,并组装进子代噬菌体,开始裂解进程。
噬菌体的DNA注入宿主细胞中后经早期发育和晚期发育最终可以裂解从而释放出子代噬菌体,完成整个裂解发育。
早期感染是指从噬菌体到进入复制开始的时期,晚期感染是指从复制开始到最后细胞裂解释放出子代噬菌体颗粒这一时期。
噬菌体内的必需基因组不大,当噬菌体的DNA注入细菌体内时,若发生裂解发育,在早期发育过程中噬菌体的DNA会被优先复制,并转录成mRNA,mRNA可以代替宿主细胞的mRNA重新引导噬菌体的活性,从而使噬菌体大量增殖,所以一个噬菌体感染将产生大量经过复制和重组的子代噬菌体。
λ噬菌体内个基因的功能裂解周期在噬菌体的裂解周期调控中,调控子编码相关功能蛋白质的基因成簇排列,少数调控开关可以控制裂解进程的通路。
在整个调控过程中调控基因以级联反映发生作用,一个基因的表达是下一个基因的表达所必须的。
噬菌体基因表达最初阶段必须依赖宿主进行转录,最初只有少数基因进行表达,在大多数情况下称为早期基因(early gene),在λ噬菌体中成为早早期基因(immediate early gene)。
它们是噬菌体裂解周期的早期基因调控部分,可以进行级联反应从而引发下一阶段的转换。
λ噬菌体有两个早早期基因,N和cro,它们由宿主RNA聚合酶转录。
其中N基因可以编码一个抗终止因子,该因子作用于nut位点,允许转录继续进入迟早期基因;cro基因有上冲功能:阻止阻抑物的合成,关闭早早期基因的表达。
当收到早期基因的诱导之后,迟早期基因(delayed early gene)或中期基因(middle gene)开始表达。
这类基因和早期基因包括了噬菌体装配自身的颗粒外壳和裂解细胞以外的所有噬菌体功能。
迟早期基因包括2个用于复制的基因和7个用于重组的基因,还有3个编码调控因子的基因,其中调控因子具有相反的功能。
λ噬菌体裂解级联途径与溶原途径连锁当噬菌体的DNA开始复制时,可以引起晚期基因(late gene)的表达。
每套基因都含有一种为下一套基因表达所需的调控因子,利用这些连续控制形成一个级联反应,从而使同基因在特定时期内被开启或关闭。
pN是抗终止因子,可以使RNA聚合酶通过来那个调早早期基因的终点而继续转录。
pQ是迟早期基因的产物,也是抗终止因子,能是RNA聚合酶转录晚期基因。
通过λ噬菌体的裂解级联反应,我们可以发现整个程序是预先安排就绪的,都是从两个启动子P L和P R开始转录早早期基因N和cro。
由于λ噬菌体是利用抗终止作用进行下一个阶段的表达。
启动子P L、P R位于cⅠ的左右两测。
与每一个启动子相连的为操纵基因(O L、O R)。
裂解级联反应具有连续性,因而调控区可以为它提供能控制整个裂解周期的作用位点。
阻抑物通过组织RNA聚合酶靠近这些启动子来阻止噬菌体基因组进入裂解周期。
阻抑物c Ⅰ的突变可以使噬菌体无法维持溶原态进入裂解周期。
在噬菌体处于溶原状态时,阻止cro基因的表达式非常重要的。
阻抑物结合在OL位点上时关闭从PL开始的N基因转录,结合到OR位点上时可以关闭cro基因的转录,同时也是cⅠ基因转录所需要的。
在OL处时,阻抑物可以阻止RNA聚合酶在PL处起始转录,从而停止N基因的表达。
在OR处,阻抑物的结合一直乐PR的使用,因此cro和其他右向早期基因不能正确表达。
当阻抑物结合在OR处时,RNA聚合酶可以有效启动转录,从而促进cⅠ基因的表达,形成一个自主调控的正回路,在这个回路中需要有阻抑物不断合成期自身。
从而控制回路保持了溶原态的稳定存在,只要阻抑物含量充分,cⅠ基因就可以继续表达,结果是OLOR被继续无限期地占用。
通过阻遏整个裂解级联反应,从而使原噬菌体保持溶原状态。
溶原通过一个自主调控回路维持,如果一个步骤被干扰,裂解周期开始阻抑物的存在可以为噬菌体侵入细菌提供免疫性,当第二个噬菌体DNA侵入溶原细胞时,已存在噬菌体所产生的阻抑物同样可以抑制新噬菌体的级联反应,从而使新的噬菌体进入溶原状态。
当λ噬菌体DNA侵入细胞时,在整个控制回路中没有阻抑物的存在,所以细菌RNA 聚合酶不能通过此回路而转录cⅠ。
因而在λ噬菌体刚侵入细胞时,需要另外两个基因cⅡ和cⅢ进行转录。
这套系统可以产生起始的阻抑物,从而诱导cⅠ途径的表达。
因而,溶原反应建立和维持的整个反应过程可以用下面如来表示。
溶原化需要级联反应,接着级联反应停止,被阻抑物所维持的自主调控系统所取代cⅡ蛋白以另一种间接方式促进了溶原态的产生,从而引起了Q基因启动子的转录,这个转录物事Q区域的反义转录得到的。
通过cⅠ阻抑物基因的转录来直接启动溶原态的机制,同时也通过抑制cro和Q基因的裂解途径所需的控制基因,间接促进了溶原态的产生。
λ噬菌体可以选择进入溶原状态或开始裂解状态。
我们已经看到,溶原是通过建立自控的维持回路起始的,这个回路通过在两个部位上施加影响抑制整个裂解级联反应。
建立溶原的过程实际上是通过控制裂解级联反应的某些步骤来实现的。
cro蛋白可以负责阻抑物的合成和抑制早期基因的表达,从而使噬菌体进入溶原状态。
它可以形成一个小的二聚体作用于免疫区,并具有两种作用:维持回路阻止阻抑物的合成,抑制早期基因从P R、P L进行表达。
溶原与裂解途径有非常密切的关系,溶原与裂解都遵循同一条代谢途径,两者都经历早早期基因的表达,并延伸进入迟早期基因。
进入裂解周期的其实时间是cro蛋白在O R3处的结合,这就停止了P RM处开始的溶原维持回路。
Cro蛋白必须与O R1、O R2或O L1、O L2结合,调节早期基因的表达。
通过停止合成cⅡ和cⅢ蛋白,这一行为即导致停止从PRE合成阻抑物。
当不稳定的cⅡ和cⅢ蛋白降解时,阻抑物已建回路就关闭。
在某种特定条件下,cⅡ蛋白的水平决定了感染的结果。
增强cⅡ蛋白稳定性的突变会增加溶原化的频率,这种突变发生在cⅡ自身或其他基因中。
cⅡ蛋白不稳定的原因是它对于宿主蛋白酶降解的敏感性,它在细胞中的浓度受到cⅢ和宿主功能的影响。
其次是λ噬菌体cⅢ蛋白的作用:它保护cⅡ蛋白不受降解。
虽然cⅢ蛋白不能保证cⅡ蛋白继续存在,但缺少cⅢ蛋白时,cⅡ蛋白实际上总是没有活性。
宿主细胞对于cⅡ蛋白水平的影响,为细菌干扰λ噬菌体选择溶原化或裂解提供了一种方法。
当λ噬菌体倾向于是生长很好的细胞裂解,但而当细胞饥饿时更倾向于进入溶原化状态。
看法:λ噬菌体在整个基因的表达中采取了一种非常聪明的策略,这种策略可以使侵染进入细菌的噬菌体充分利用其周围的环境。
当其进入的宿主细胞处于良好的状态和环境中时,噬菌体便进入裂解状态;而当宿主细胞没有处于最佳状态时,噬菌体便倾向于进入溶原化状态,从而使自身得到最大程度上的复制。
参考文献:1.Lewin B.著, 余龙等译基因Ⅷ. 科学出版社,2005,北京n B. D. et. al. Revisited gene regulation in bacteriophage λ. Curr. Opin. Genet. Dev.15:145-1523.Max G. Bacteriophage λ: The Untold Story. J. Mol. Biol. 1999, 293: 177-1804.Donald L. C. et. al. A new look at Bacteriophage λgenetic networks. J. Bacteriol 2007, 189(2): 298 –3045.Harrison E. et.al. Establishment and Maintenance of Repression by Bacteriophage Lambda: the role of the cI, cII and cIII proteins. Proc. Nat. Acad. Sci. USA, 1971, 68: 21 90-21946.Ronald A. A. et. al. How Cro and lambda-repressor distinguish between operators : thestructural basis underlying a genetic switch. Proc. Nat. Acad. Sci. USA.1998, 95: 34 31-34367.Brooks, K. and Clark, A.J. (1967) J. Virol. 1, 283^293.。