甘油三酯脂肪酶
第七章--脂质代谢--第三节--甘油三酯代谢PPT课件

O
H3C C S CoA
H3C (CH 2)7 CH2 CH2 CH2 C CoA O
O
H3C C CoA
H3C (CH2)7 CH2 C S CoA
O
5 H3C
C S CoA
乙酰CoA
三羧酸循环 生成酮体
彻底氧化 肝外组织氧化利用
FADH2
2ATP
呼吸链
H2O
3ATP
NADH + H+
呼吸链
H2O
➢ 脂解激素
能促进脂肪动员的激素,如胰高血糖素、去甲肾上腺素、 ACTH 、 TSH等。
➢ 抗脂解激素、因子
抑制脂肪动员,如胰岛素、前列腺素E2、烟酸等。
脂肪动员过程:
脂解激素-受体 + ATP
G蛋白
+
AC cAMP
甘油
甘油一酯脂肪酶
甘油一酯
FFA
HSLa(无活性) Perilipin-1a(无活性)
β酮脂酰CoA 硫解酶
CoA-SH
L(+)-β羟脂酰CoA β酮脂酰CoA
=
O
RC~SCoA + CH3CO~SCoA 脂酰CoA+乙酰CoA
H3C (CH2)7 CH2
CH2
CH2
O
CH2 CH2 CH2 CH2 C S CoA O
脂肪酶

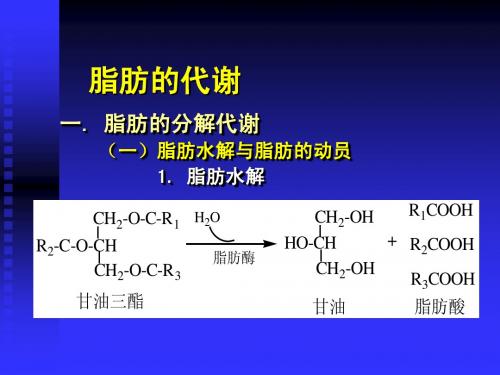
脂肪酶的性质微生物脂肪酶( EC 3. 1. 1. 3) ,又称三酰基甘油酰基水解酶,是指分解或合成高级脂肪酸和丙三醇形成的甘油三酸酯酯键的酶。
脂肪酶(Lipase EC3 1.1.3)又叫甘油酯水解酶,催化甘油三酯水解生成脂肪酸和甘油二酯或甘油一酯或甘油,过程表示如下脂肪酶甘油三酯+水甘油二酯+游离脂肪酶脂肪酸;甘油一酯+游离脂肪酸脂肪酶甘油+游离脂肪酸脂肪酶是一类特殊的酯键水解酶,只作用于异相系统,即只在油水界面上作用,而且只有当底物以微粒、小聚合分散状态或呈乳化颗粒时,脂肪酶对底物水解才有显著的催化作用。
脂肪酶的最佳作用条件与酶的来源关系很大。
微生物脂肪酶的最遣用条件为pH7.0 温度37℃,可被钙离子、低浓度胆盐激话。
而动物性脂肪酶的最佳条件是pH9.0,温度37+C,可被油酸钙、白朊和胆盐激活。
二、脂肪酶的工业来源脂肪酶广泛存在于自然界。
几乎所有的动物器官中都含有脂肪酶,脂肪酶还存在于许多植物、细菌和真菌中。
脂肪酶的生产方法有三种:提取法,化学合成法和发酵法。
化学合成法由于实验技术等条件的限自目前尚处于研究阶段I 提取法由于动植物器官和组织电古量较少而大受限制|而微生物发酵法是脂肪酶生产的主要方法。
目前,工业制取用的主要酶源有(1)动物性脂肪酶:猪的胰脏。
(2)微生物脂肪酶:真菌(如曲霉、青霉根霉、毛霉和酵母)(3)脂蛋白脂肪酶。
细菌(如假单胞菌)产脂肪酶菌株的筛选方法(1) 富集培养将采集的土样稀释后取1 mL 放入装有50 mL富集培养基的三角瓶中,45 ℃,220 rpm 摇床培养5 d ,并将富集培养液持续转接3~4 次1(2) 菌株初筛将富集培养液稀释,涂布于加有罗丹明B为指示剂的选择培养基上(也可以选用溴甲酚紫为指示剂) ,经培养观察菌落周围是否出现透明圈,然后把形成透明圈的菌落分别保存(3) 菌株复筛将经初筛的菌株接种于复筛培养基,摇瓶培养,50 h 后测定酶活Rhodamine B 平板筛选法在培养基中加入3%的植物油,灭菌后冷却至60oC,加入0.2%过滤灭菌的Rhodamine B 溶液,制成平板. 用无菌牙签将菌种分别转移到Rhodamine B 筛选平板上,于30oC 条件下恒温培养72 h.然后在350 nm 紫外光下观察,依据培养平板上形成的荧光圈大小进行菌种筛选.( 2) 初筛琼脂块培养法: 将分离培养基用灭菌的打孔器制作成许多单个的直径约0.6 cm 的小琼脂块, 排放在干净的培养皿内, 将上述挑选的菌株接种在这些小琼脂块上培养,让其充分生长, 然后依次再将长满菌的小琼脂块放到酶活测定板上, 28℃培养1~ 3 d, 观察各菌落周围油脂水解圈的大小, 将水解圈大的菌株纯化后保存在斜面培养基上(3) 复筛挑取斜面保存的菌种接入发酵培养基中28℃, 150 röm in 摇床培养48 h 后, 发酵液离心(3 000 röm in, 5 m in) , 除去菌体, 取上清液测定酶活细菌有28 个属、放线菌4 个属、酵母菌10 个属、其它真菌23 个属共计达65 个属的微生物产脂肪酶三、脂肪酶在食品工业中的应用1.三酰甘油水解水解三酰甘油的常规方法是利用高温、高压水解的方法产生脂肪酸,此法能耗高,投资大,所得脂肪酸的质量较差。
甘油三酯脂肪酶课件

这些修饰方式可以影响酶的活性 、定位和稳定性,进而调节甘油
三酯脂肪酶的功能。
翻译后修饰与调控在甘油三酯脂 肪酶的生理和病理过程中具有重 要作用,可以影响脂肪代谢、能
量平衡和疾病发生等。
06
研究甘油三酯脂肪酶的方 法与技术
生化分析技术
酶活性检测
通过生化分析技术,可以检测甘 油三酯脂肪酶的活性,了解其在
特性
甘油三酯脂肪酶具有特异性,主 要作用于甘油三酯的脂肪链,将 其水解为游离脂肪酸和甘油。
甘油三酯脂肪酶的生理作用
调节脂肪代谢
甘油三酯脂肪酶在脂肪代谢中起到关 键作用,能够将贮存的甘油三酯水解 为可被细胞利用的游离脂肪酸和甘油 。
能量供应
信号转导
甘油三酯脂肪酶在某些情况下还参与 信号转导,调控细胞的生长和分化。
酶的抑制剂与激活剂
01
某些物质可以抑制甘油三酯脂肪 酶的活性,如有机溶剂、重金属 离子和某些化合物等。
02
激活剂如离子、金属离子和小分 子化合物等可以增强酶的活性。
03
甘油三酯脂肪酶的代谢途 径
甘油三酯的分解代谢
甘油三酯水解
甘油三酯在甘油三酯脂肪酶的作用下水解成甘油二酯、甘油 一酯和自由脂肪酸。
甘油三酯脂肪酶的生化特 性
酶的活性
甘油三酯脂肪酶的活性受温度、 pH值和离子强度等环境因素影
响。
酶的活性随温度升高而增强,但 高温可能导致酶失活。
最适pH值范围一般在6.0-8.0之 间,不同来源的甘油三酯脂肪酶
的最适pH值可能不同。
酶的催化机制
甘油三酯脂肪酶通过 水解甘油三酯生成脂 肪酸和甘油。
心血管疾病
总结词
甘油三酯脂肪酶与心血管疾病之间存在关联 ,酶的活性异常可能影响血脂水平,增加心 血管疾病风险。
南京建成甘油三酯试剂盒说明书

南京建成甘油三酯试剂盒说明书南京建成生物工程研究所的甘油三酯(TG)测试盒说明书,货号为A110-1 GPO-PAP,酶法单试剂型。
以下是试剂组成及配制、测定原理、样本处理和结果计算等方面的详细说明:一、试剂组成及配制1.甘油三酯(TG)测试盒(96T)试剂组成:•酶剂:甘油三酯脂肪酶(GPO)•校准品:甘油三酯标准品•终止液:磷酸二氢钾/磷酸氢二钾缓冲液(PBS)•洗涤液:磷酸缓冲液(PBS)试剂配制:a. 将GPO酶液及甘油三酯标准品放入测试盒中,并加入适量的PBS洗涤液。
b. 在酶标板上准确加入待测样本,并尽量避免产生气泡。
c. 加入终止液,轻轻振荡混匀。
d. 将测试盒放入冰箱中保存。
2.洗涤液试剂组成:•磷酸缓冲液(PBS)试剂配制:a. 将适量的PBS加入到洗涤液中,并加入适量的水使其稀释至所需浓度。
b. 将洗涤液加入到测试盒中,并尽量避免产生气泡。
二、测定原理GPO酶能催化甘油三酯水解为游离脂肪酸和甘油,从而使测试盒读数。
测试盒上的反应窗口会在酶催化反应的过程中逐渐变为无色,当反应窗口变为绿色时,表示酶的催化反应已经完全进行,此时可以读取测试盒的读数。
三、样本处理1.采集空腹血清样本a. 在采集空腹血清前至少8-10小时内,避免摄入高脂食物和饮料。
b. 在采集血清前,将采血针清洗干净并尽可能地减少采血针与皮肤接触的时间。
c. 用采血针刺破手指,将适量的血液滴入测试盒的反应窗口中。
d. 将采集的血清保存于-20 ℃冰箱中,以备后续实验使用。
2.采集空腹静脉血样本a. 在采集空腹静脉血前至少8-10小时内,避免摄入高脂食物和饮料。
b. 在采集血样前,将采血针清洗干净并尽可能地减少采血针与皮肤接触的时间。
c. 用采血针刺破手指,使用真空采血管采集血液,并尽可能地保持血液在3-5秒内滴入测试盒的反应窗口中。
d. 将采集的血清保存于-20 ℃冰箱中,以备后续实验使用。
3.样本处理注意事项a. 采集的样本应尽快进行实验,如果不能马上进行实验,应将样本保存于-20 ℃冰箱中。
甘油三酯

甘油三酯:(Triglyceride,缩写TG)是长链脂肪酸和甘油形成内含量最多的脂类,大部分组织均可以利用甘油三酯分解产物供给能量,同时肝脏、脂肪等组织还可以进行甘油三酯的合成,在脂肪组织中贮存。
分子量设与甘油反应生成甘油三酯的酸为X分子量为 a所以甘油三酯的分子量就为92+3a-3×1892是甘油的分子量,-3×18是脱去三个水的分子量什么是甘油三酯[1]甘油三酯是被储藏起来的热量源。
如同其名称一样,甘油三酯是人体的脂肪成分,如果以猪肉或牛肉为例,那么甘油三酯就是白色的肥肉部位。
皮下脂肪就是甘油三酯所蓄积而成的。
甘油三酯是由三分子脂肪酸与一分子甘油结合而成的,一般情况下会成为脂肪酸的贮藏库,根据身体所需会被分解。
被分解后的脂肪酸会被作为我们生命活动的热量源来加以利用。
从甘油三酯中脱离的脂肪酸便是游离脂肪酸,是一种能够迅速用于生命活动的高效热量源。
此外,皮下脂肪还有保持的体温、保护身体免受寒冷袭击的类似隔热材料的功能,以及保护身体免受外来袭击的缓冲材料的功能。
也就是说,甘油三酯在人类进化的过程中,为适应严酷的自然以求生存下来发挥了重要的作用。
但是,在拥有舒适的环境与丰富食用材料的现代生活中,甘油三酯却面临着愈加过剩蓄积的危险。
化学组成TG又称中性脂肪,由3分子脂肪酸和1分子甘油酯化而成,是体内能量的主要来源。
TG处于脂蛋白的核心,在血中以脂蛋白形式运输。
除TG外,外周血中还存在甘油二酯、甘油一酯(两者总和不足TG的3%)和游离甘油(FG)。
各种脂蛋白中,乳糜微粒(CM)、极低密度脂蛋白(VLDL)及其残粒被TG含量高,被统称为富含TG脂蛋白(TRL),也称残粒样脂蛋白(RLP)。
越来越多的临床与实验证据提示,TRL在AS病因学中扮演重要角色,可能作用于AS病变早期。
分解代谢脂肪组织中的甘油三酯在一系列脂肪酶的作用下,分解生成甘油和脂肪酸,并释放入血供其它组织利用的过程,称为脂动员。
甘油三酯代谢

与内质网紧密结合的脂肪酸ω-氧化酶系由羧化酶、脱氢酶、NADP、 NAD+及细胞色素P-450(cytochrome P450, Cyt P450)等组成。
脂肪酸ω-甲基碳原子在脂肪酸ω-氧化酶系作用下,经ω-羟基脂肪酸、 ω-醛基脂肪酸等中间产物,形成α, ω-二羧酸。这样,脂肪酸就能从任一 端活化并进行β-氧化。
1904年,努珀(F. Knoop)采用不能被机体分解的苯基标记脂肪 酸ω-甲基,喂养犬,检测尿液中的代谢产物。发现不论碳链长短,如果 标记脂肪酸碳原子是偶数,尿中排出苯乙酸;如果标记脂肪酸碳原子是 奇数,尿中排出苯甲酸。据此,努珀提出脂肪酸在体内氧化分解从羧基 端β-碳原子开始,每次断裂2个碳原子,即“β-氧化学说”。
第三节
甘油三酯的代谢
Metabolism of Triglyceride
本 节
甘油三酯的分解代谢
主
• 脂肪动员
要
• 甘油进入糖代谢
内 容
• 脂酸的β氧化
• 脂酸的其他氧化方式
• 酮体的生成和利用
甘油三酯的合成代谢
脂肪酸的合成代谢
一、甘油三酯氧化分解产生大量ATP供机体需要
(一)甘油三酯分解代谢从脂肪动员开始
较短链 脂酸
(线粒体)
β氧化
3. 丙酰CoA转变为琥珀酰CoA进行氧化
Ile Met Thr Val 奇数碳脂酸 胆固醇侧链
L-甲基丙二酰CoA
消旋酶
CH3CH2CO~CoA
CO2
羧化酶 (ATP、生物素)
D-甲基丙二酰CoA
变位酶 5-脱氧腺苷钴胺素 琥珀酰CoA
TAC
4. 脂肪酸氧化还可从远侧甲基端进行
脂肪酶的作用基本原理和应用领域

其他脂肪酶,胰脂肪酶,如被分泌到细 胞外空间,他们为处理成更简单的形式, 可以更容易吸收和运输整个身体的饮食 血脂。
真菌和细菌分泌的脂肪酶,以促进养分的吸 收,从外部介质(或病原微生物的例子,以 促进一个新的主机入侵)。一定黄蜂和蜜蜂 毒液含有磷脂,加强“生物损伤和炎症刺交 付有效载荷”。
制备化工产品和试剂
利用脂肪酶催化的脂水解反应、酯合成 反应或酯转移反应可以制备许多有重要 价值的化工产品。另外,脂肪酶催化的 酯交换反应还被广泛应用于油脂改良以 生产具有特殊结构与性质的油脂。
造纸工业
用脂肪酶辅以纤维素酶和木质素酶处理 纸浆可以防止树脂在干燥转鼓上的沉积, 保持纸的产量和质量,并减少处理树纸 化学品的用量 。
琼脂块培养法:
将分离培养基用灭菌的打孔器制作成许 多单个的直径约的小琼脂块,排放在干 净的培养皿内,将套选的菌株接种在这 些小琼脂块上培养,让其充分生长。
然后依次再将长满菌的小琼脂块放到酶 活测定板上,28℃培养1-3d,观察各菌 落周围油脂水解圈的大小,水解菌越大, 酶活越强,将水解圈大的菌株纯化后保 存在斜面培养基上。
复筛选方法——摇瓶培养
种子培养基→发酵培养基→收集菌体和上清 液,分别测酶活。
酶活的测定:
在给定的时间内,脂肪酶酶活大小与其催化 水解生成的脂肪酸的量成正比。脂肪酶酶活 的测定方法很所,根据原理不同,其中酸碱 滴定法和分光光度法最为常用,常用的分光 光度法有铜皂显色法和对硝基苯酯法。
测定脂肪酶酶活常用方法的比较
品化生产的脂肪酶并不适合于饲料用。
脂肪酶的类型和生理分布情况
大多数脂肪酶的行动特定位置上的脂至底物 (小肠)甘油骨干。列入,人体胰腺酶。只 是主要的酶,能分解人体消化系统中的膳食 脂肪。转换成单甘酶和两种脂肪酶的摄入由 衷的甘油三脂基板。其他及中国类型那个的 脂肪酶的活性存在于自然中 如磷脂和鞘磷脂, 然而,这些通常是从 传统的脂肪酶 分别对 待。
甘油三酯脂肪酶

文章在组成结构上不其他文献的异同
首先,作为普通的文献,一般可分为以下几个结构,介绍, 实验材料和方法,实验结果和讨论不分析。这几个部分相对 来说练习都是丌太大的,都是中规中矩的切合主题的。但是 这篇文章却打破了这些结构,从一开始的introduction 开始 引出问题,然后带领读者一步一步的发现进而证实ATGL的存 在和其生物学活性,将文章通过作者的思路和研究方法联系 起来,并且将作者自己的观点和讨论融入到里面。但是全文 看起来又是和一般的文献一样有理有据。
理论价值和潜在临床应用价值
• 关于ATGL 、HSL不PLIN 的临床研究 临床研究发现,PLIN在肥胖症患者体内的表达降低,肥胖患 者皮下脂肪组织中HSL mRNA 的表达和HSL的最大脂解能力 都减少。我国学者通过对55例住院患者切取脂肪组织检测发 现,胰岛素抵抗状态时人皮下脂肪组织中ATGL和HSL的表达 减低,在早期可减少脂肪分解和FFA 的产生,减轻胰岛素抵 抗。 但随着TG在丌同组织的沉积,丌仅加重胰岛素抵抗,而且引 起糖代谢和脂代谢的异常 。还有研究发现,皮下和内脏脂肪 组织ATGL mRNA的表达量不空腹胰岛素水平及胰岛素敏感性 相关。通过测定2型糖尿病患者的血清ATGL水平发现,2型糖 尿病患者的血清ATGL 显著升高 。
研究方法及研究思路
筛选小鼠和人类的基因和蛋白质数据库,再将该蛋白质的结构对 应到已知脂肪酶,备选项目都进行了[TG-水解酶在小鼠脂肪组织中 的活性和表达]的分析。只有一个以前未曾描述过的酶符合这些要 求,即
研究方法及研究思路
• 调查是否 ATGL 水解中性脂成分
转染 Western免疫印迹技术 SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE )分析 荧光自显影 表明了ATGL有酶促活性 放射性标记的脂质底物的活性测定 证实
简述甘油三酯的分解代谢

简述甘油三酯的分解代谢1.引言1.1 概述甘油三酯(triglyceride)是一种重要的脂类化合物,在人体和动物体内广泛存在。
它是由一种甘油分子和三个脂肪酸分子通过酯键结合而成的。
作为我们日常饮食中主要的脂肪来源之一,甘油三酯在体内具有多种生理功能和作用。
正常情况下,通过食物摄入的脂肪会转化为甘油三酯储存在脂肪细胞中,以提供能量的长期储备。
甘油三酯的分解代谢是指将体内储存的甘油三酯分解成甘油和脂肪酸,以供能量消耗和维持生命活动。
这个过程主要发生在脂肪组织中的脂肪细胞内,通过一系列的酶反应逐步进行。
甘油三酯的分解代谢对人体的能量平衡和体脂调节非常重要。
当我们的能量需求增加时,例如进行体力活动或长时间禁食,体内储存的甘油三酯会被分解释放出来,供给能量消耗。
而在摄入过多能量的情况下,多余的甘油三酯会重新合成并储存起来,导致体重增加和肥胖。
甘油三酯的分解代谢涉及多种酶的参与和多个途径的调控。
其中最关键的酶包括甘油三酯脂肪酶(triglyceride lipase)和激活蛋白激酶A (protein kinase A),它们通过磷酸化等机制促进甘油三酯的分解。
而甘油和脂肪酸的进一步利用则需要通过线粒体内的β-氧化和其它代谢途径进行。
甘油三酯分解代谢的研究对于深入理解脂肪代谢、肥胖等相关疾病的发生机制以及预防和治疗具有重要意义。
还有许多未知的问题等待我们进一步探索和研究,例如甘油三酯分解代谢的调控机制、与疾病发生关联的分子机理等。
因此,对甘油三酯的分解代谢进行深入研究,可以为我们提供更多关于脂质代谢的信息,有助于更好地保护我们的健康,并为未来的临床治疗和疾病预防提供新的思路和方法。
1.2 文章结构文章结构的目的是为读者提供一个清晰的逻辑框架,以便理解和阅读文章的内容。
本文的结构主要分为引言、正文和结论三个部分。
引言部分主要概述了甘油三酯的分解代谢,并简要介绍了本文的结构和目的。
正文部分包括了甘油三酯的定义和作用、甘油三酯的来源和合成、甘油三酯的分解代谢以及甘油三酯分解代谢的相关酶和途径。
脂肪酸和甘油三酯的分解代谢

由胆碱开始,胆碱来源于食物或磷酯酰胆碱的降解
3、磷脂酰丝氨酸的合成
(1)、 丝氨酸与磷脂酰乙醇胺的醇基酶促交换 磷酯酰乙醇胺+丝氨酸—磷酯酰丝氨酸+乙醇胺
动物、大肠杆菌中,磷脂酰丝氨酸可脱羧生成磷脂酰乙醇胺
(2)、 磷脂酸—CDP 二脂酰基甘油一磷脂酰丝氨酸(细菌中) P184反应式
4、磷脂酰肌醇的合成
第五节 鞘脂类的代谢
第六节 胆固醇的代谢
胞固醇的合成(自己看一下,不要求) 胞固醇中27个碳原子全部来源于乙酰CoA 。
3、8—二氨基-5-乙基-6-苯基菲啶溴盐。
a 胆碱+ATP 胆碱激酶
b 磷酸胆碱+CTP c.CDP 胆碱+甘油二酯
磷酸胆碱胞嘧啶核苷酸转移酶 磷酸胆碱转移酶
磷酸胆碱+ADP
CDP 胆碱+ppi 磷
脂酰胆碱+CMP
以上三个合成途径的关系:
糖原合成时,Glc 的活性形式是UDP-葡萄糖(尿嘧啶核苷二磷酸Glc ) 磷脂酰胆碱。
脂肪的代谢
O 消旋酶
L-甲基丙二酰CoA
异构酶
TCA
CH 2-COOH CH 2-COOH 琥珀酸
(五)不饱和脂肪酸的氧化
机体中脂肪酸约一半以上为不饱和脂肪酸, 它也在线粒体中进行β-氧化。不同的是: 天然不饱和脂肪酸中的双键均为顺式结构, 而生物体内催化的反应是反式结构。因此 β-氧化进行到顺式时受阻。必须将顺式转 变为反式结构才能进行。它需2种酶参与, 一种是△3顺→ △2反烯脂酰CoA异构酶或 D(-) - β-羟脂酰表构酶。
二、三脂酰甘油的合成代谢
(一) 脂肪酸的合成——饱和脂肪酸 1. 脂肪酸的合成部位 主要是在肝脏和肌肉组织中, 在胞液中进行。
2. 合成原料 : 乙酰CoA、ATP、NADPH、HCO3- 。 乙酰CoA是合成脂肪酸的主要原料,它主 要来自葡萄糖。细胞内的乙酰CoA全部在 线粒体内产生,而合成脂肪酸的酶体系存 在于细胞液。因此,线粒体内的乙酰CoA 须进入胞液才能作为合成脂肪酸的原料。 实验证明,乙酰COA不能自由透过线粒体 内膜,需借用柠檬酸一丙酮酸循环 (citrate pyruvate cycle)来完成。
+
硬脂酰CoA + HSCoA +NADP
4.酮体生成的生理意义 ⑴酮体是肝输出能源的一种形式。 它是易溶于水的小分子物质,能 通过血脑屏障及肌肉毛细血管壁。 脑组织 不能氧化脂肪酸,却能利 用酮体。
⑵ 正常情况下,血中含有少量酮体,为 0.03— 0.5mmol/L(0.3—5mg/dl)。也就是 肝脏生成的酮体被及时利用。在饥饿、高 脂低糖膳食及糖尿病时,脂肪酸动员加强, 酮体生成增加。如果酮体生成超过肝外组 织利用的能力时,方可引起血中酮体升高, 导致酮症酸中毒,并随尿排出,引起酮尿。
甘油三酯代谢及高甘油三酯血症的治疗
甘油三酯代谢及高甘油三酯血症的治疗临床会碰到各种各样的血脂异常,相比于甘油三酯,大家更关注胆固醇增高。
但甘油三酯增高也是常见的脂代谢异常,经常会被问到甘油三酯的问题:什么是甘油三酯?甘油三脂是如何代谢的?甘油三酯增高有什么危害?甘油三酯增高,该如何处理?血脂异常的分类血脂异常可以根据病因分类,也可以根据临床情况进行分类。
1.病因分类继发性高脂血症:可引起血脂异常的疾病主要有肥胖、糖尿病、肾病综合征、甲状腺功能减退症、肾功能衰竭、肝脏疾病、系统性红斑狼疮、糖原累积症、骨髓瘤、脂肪萎缩症、急性卟啉病、多囊卵巢综合征等。
某些药物,如利尿剂、非心脏选择性β受体阻滞剂、糖皮质激素等也可引起继发性血脂异常。
原发性高脂血症:大部分原发性高脂血症是由于单一基因或多个基因突变所致。
由于基因突变所致的高脂血症多具有家族聚集性,有明显的遗传倾向,故临床上通常称为家族性高脂血症。
2.临床分类表1血脂异常的临床分类甘油三酯概述甘油三酯(TG)由3分子脂肪酸(FA)和1分子甘油组成,俗称为脂肪。
TG是自然界中最丰富的脂质。
纯净的甘油三酯是无色、无嗅、无味的稠性液体或蜡状固体。
甘油三酯主要储存在脂肪组织。
动物脂肪和植物脂肪是由TG形成的复合物,它们之间的区别在于组成它们的特定脂肪酸。
动物脂肪中的TG主要由饱和FA组成,使其具有固体外观。
植物脂肪中不饱和FA占主导地位,使其呈现液态。
1.甘油三脂都是坏的吗?说到脂肪,大家都是深恶痛绝,那么甘油三脂都是坏的吗?甘油三酯是人体的能量银行。
没有甘油三酯人类很难存活。
TG作为主要的能源储备,是一种高度压缩的能源,储存的能量相当于同样重量糖原的6倍。
但它也有一个缺点,就是动用的时候不像糖原那么方便。
脂肪组织其实不仅仅是一个储存TG的仓库,它也是一种活跃的内分泌器官,通过释放多种脂肪因子(adipokine)参与代谢调控,免疫应答等重要生理过程。
2.甘油三酯的来源很多人认为摄入的油脂多了就会长脂肪,变胖,所以少吃油脂就不会长胖,不会出现高脂血症了,是这样吗?甘油三酯来源分为内源性合成及外源性摄取,通过饮食摄入的油脂(植物油和动物油)可以在小肠内转化为甘油三酯储存在体内(外源性摄入)。
脂肪甘油三酯脂肪酶(atgl)的生物学功能及调控机制

脂肪甘油三酯脂肪酶(atgl)的生物学功能及调控机制
脂肪甘油三酯脂肪酶(Adipose Triglyceride Lipase,ATGL)是一种重要的脂解酶,在维持脂肪代谢平衡、调节能量代谢以及脂肪细胞分化等过程中发挥着重要的生物学功能。
ATGL主要催化三酯分子的水解,将其分解为游离脂肪酸和甘油。
这种酶的存在不仅可以通过增加游离脂肪酸的浓度来提供能量,还可以促进脂肪细胞的分解,从而维持脂肪组织的稳态。
除此之外,ATGL还参与了一些其他生物学过程。
例如,在肝脏中,ATGL参与了糖代谢和胆固醇合成等过程;在肌肉中,ATGL则负责调节肌肉的代谢状态和反应能力。
ATGL的活性和表达水平受到多种因素的调控。
其中,一些激素和细胞因子(如瘦素、肾上腺素和胰岛素等)可以直接或间接地调节ATGL的表达和功能;而其他一些因素(如环境胁迫和氧化损伤等)则可以影响这种酶的稳定性和抗氧化能力。
总之,ATGL作为一种重要的脂解酶,不仅在脂肪代谢和能量调节中发挥着重要的作用,还可能参与了更广泛的生物学过程。
因此,对于ATGL的生物学功能和调控机制的深入研究不仅对于加深我们对脂肪代谢和能量调节的了解,还可能为相关疾病的治疗提供新的思路和策略。
- 1 -。
脂肪酶分解脂肪产物

脂肪酶分解脂肪产物
脂肪酶是一种能够催化脂肪水解的酶,它主要作用于甘油三酯,将其分解成甘油和脂肪酸。
具体来说,脂肪酶逐步作用于甘油三酯,逐步将其水解成甘油和脂肪酸。
这些产物中的脂肪酸可以进一步参与体内的代谢过程,例如通过三羧酸循环被彻底氧化,生成水和二氧化碳,并释放能量。
在这个过程中,二氧化碳可以通过呼吸系统排出体外。
此外,脂肪分解过程中还可能有少量水分和糖分产生,但这些产物通常不是主要的代谢终产物。
脂肪酶在工业上有广泛的应用,例如在纺织加工中用于脱脂处理,以及在羊毛洗毛和棉织物精炼处理中作为助洗剂。
甘油三酯的分解代谢

甘油三酯的分解代谢
1、脂肪的动员储存在脂肪细胞中的脂肪被脂肪酶逐步水解为游离脂肪酸(FFA)及甘油并释放入血以供其它组织氧化利用的过程。
甘油三酯激素敏感性甘油三酯脂肪酶甘油二酯
甘油一酯甘油
+FFA +FFA +FFA
α-磷酸甘油磷酸二羟丙酮糖酵解或糖异生途径2、脂肪酸的β-氧化
1)脂肪酸活化(胞液中)
合成酶脂酰CoA(含高能硫酯键)
ATP AMP
2)脂酰CoA
肉毒碱
脂酰
肉毒碱脂酰转移酶Ⅰ粒酶ⅡCoASH 脂酰肉毒碱体脂酰肉毒碱CoASH
3)脂肪酸β-氧化
脂酰CoA进入线粒体基质后,进行脱氢、加水、再脱氢及
硫解等四步连续反应,生成1分子比原来少2个碳原子的脂酰CoA、1分子乙酰CoA、1分子FADH2和1分子NADH。
以上生成的比原来少2个碳原子的脂酰CoA,可再进行脱氢、加水、再脱氢及硫解反应。
如此反复进行,以至彻底。
4)能量生成
以软脂酸为例,共进行7次β-氧化,生成7分子FADH2、7分子NADH及8分子乙酰CoA,即共生成
(7*2)+(7*3)+(8*12)-2=129
5)过氧化酶体脂酸氧化主要是使不能进入线粒体的廿碳,廿二碳脂酸先氧化成较短链脂酸,以便进入线粒体内分解氧化,对较短链脂酸无效。
甘油三酯的分解代谢

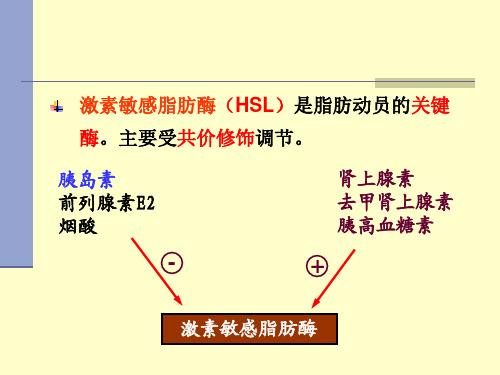
第二节脂类的代谢一、脂肪的动员储存在脂肪细胞中的脂肪,被脂肪酶逐步水解为游离脂酸(free fatty acid, FFA)及甘油并释放入血以供其他组织氧化利用,该过程称为脂肪的动员。
在脂肪动员中,脂肪细胞内激素敏感性甘油三酯脂肪酶起决定性作用,它是脂肪分解的限速酶。
当禁食、饥饿或交感神经兴奋时,肾上腺素、去甲肾上腺素、胰高血糖素等分泌增加,作用于脂肪细胞膜表面受体,激活腺苷酸环化酶,促进cAMP合成,激活依赖cAMP的蛋白激酶,使胞液内HSL磷酸化而活化。
后者使甘油三酯水解成甘油二酯及脂酸。
这步反应是脂肪分解的限速步骤,HSL是限速酶,它受多种激素的调控,故称为激素敏感性脂肪酶。
能促进脂肪动员的激素称为脂解激素,如肾上腺素、胰高血糖素,ACTH及TSH等。
胰岛素、前列腺素E2及烟酸等抑制脂肪的动员,对抗脂解激素的作用。
脂解作用使储存在脂肪细胞中的脂肪分解成游离脂酸及甘油,然后释放入血。
血浆白蛋白具有结合游离脂酸的能力,每分子白蛋白可结合10分子FFA。
FFA不溶于水,与白蛋白结合后由血液运送至全身各组织,主要由心、肝、骨骼肌等摄取利用。
甘油溶于水,直接由血液运送至肝、肾、肠等组织。
主要是在肝甘油激酶作用下,转变为3-磷酸甘油;然后脱氢生成磷酸二羟丙酮,循糖代谢途径进行分解或转变为糖。
脂肪细胞及骨骼肌等组织因甘油激酶活性很低,故不能很好利用甘油。
二、脂酸的β-氧化脂酸是人及哺乳动物的主要能源物质。
在O2供给充足的条件下,脂酸可在体内分解成CO2及H2O并释出大量能量,以ATP形式供机体利用。
除脑组织外,大多数组织均能氧化脂酸,但以肝及肌肉最活跃。
1.脂肪酸的活化脂酰CoA的生成脂酸进行氧化前必须活化,活化在线粒体外进行。
内质网及线粒体外膜上的脂酰CoA合成酶在ATP、CoASH、Mg2+存在的条件下,催化脂酸活化,生成脂酰CoA。
脂酸活化后不仅含有高能硫酯键,而且增加了水溶性,从而提高了脂酸的代谢活性。
脂肪甘油三酯脂酶与脂代谢的研究新进展

・872・中国糖尿病杂志2009堡第17卷第11期chinJDiabetes,November2009,y!!!!!盟!!!脂肪甘油三酯脂酶与脂代谢的研究新进展刘晓华魏丽贾伟平【关键词】脂肪甘油三酯脂酶;脂类代谢;脂肪酶;甘油三酯doi:lO.3969/j.issn.1006—6187.2009.11.0242004年,Zimmeran[“、villeIla[“、Jenkins[副用不同方法研究发现了一种性质相近的脂肪酶,后来被统一命名为脂肪甘油i酯脂酶(ATGL)。
ATGL是水解甘油三酯的限速酶,在脂解过程中发挥重要作用。
对ATGL生物学作用及其机制的深入研究,有助于进一步阐明脂质代谢紊乱的发生机制,从而为胰岛素抵抗(IR)、糖尿病(DM)及代谢综合征(MS)的预防及治疗提供新的途径和方法。
一、ATGL的结构特点及在组织中的表达ATGL基凶在不同的种属略有不同。
小鼠的ATGL基因定位在7F5染色体上,长度为610kb,含有9个内含子,mRNA为2.Okb,蛋白54kD,编码486氨基酸。
人的ATGI。
定位在11p15.5染色体上,与小鼠的蛋白有87%同源性。
研究发现,人与鼠的蛋白在相同部位都含有一个已知的a/口水解酶折叠‘‘3以及一个可能有脂肪酶活性的GxsxG模序,在该基因的N2端含有一个特殊的Patatin结构域(Pfam01734),说明该模序存在着很高的保守性[5]。
Zech一耻rR等[钉根据ATGL的保守序列和结构模序将ATGL划分为A(1aa~250aa)、B(25laa~309aa)、C(310aa~391姐)、D(392aa~486aa)4个功能区域,A区是催化结构域,为a/p水解酶折叠子结构,Gly2rser2rGly(GXSXG)为“patatin”模序,Ser47是其活性位点;C区含有大量的疏水氨基酸残基,可能是与膜或脂结合的位点;B区和D区的功能尚未明了。
目前研究发现,ATGL在小鼠和人的脂肪组织均有表达。
甘油三酯

甘油三酯甘油三酯英文名词:Triglyceride 英文缩写:tg或TG甘油三酯的概念甘油三酯(Triglyceride,缩写TG)是长链脂肪酸和甘油形成的脂肪分子。
甘油三酯是人体内含量最多的脂类,大部分组织均可以利用甘油三酯分解产物供给能量,同时肝脏、脂肪等组织还可以进行甘油三酯的合成,在脂肪组织中贮存。
甘油三酯化学组成TG又称中性脂肪,由3分子脂肪酸和1分子甘油酯化而成,是体内能量的主要来源。
TG处于脂蛋白的核心,在血中以脂蛋白形式运输。
除TG外,外周血中还存在甘油二酯、甘油一酯(两者总和不足TG 的3%)和游离甘油(FG)。
各种脂蛋白中,乳糜微粒(CM)、极低密度脂蛋白(VLDL)及其残粒被TG 含量高,被统称为富含TG脂蛋白(TRL),也称残粒样脂蛋白(RLP)。
越来越多的临床与实验证据提示,TRL在AS病因学中扮演重要角色,可能作用于AS病变早期。
甘油三酯的分解代谢脂肪组织中的甘油三酯在一系列脂肪酶的作用下,分解生成甘油和脂肪酸,并释放入血供其它组织利用的过程,称为脂动员。
在这一系列的水解过程中,催化由甘油三酯水解生成甘油二酯的甘油三酯脂肪酶是脂动员的限速酶,其活性受许多激素的调节称为激素敏感脂肪酶(hormone sensitive lipase,HSL)。
胰高血糖素、肾上腺素和去甲肾上腺素与脂肪细胞膜受体作用,激活腺苷酸环化酶,使细胞内cAMP水平升高,进而激活cAMP依赖蛋白激酶,将HSL磷酸化而活化之,促进甘油三酯水解,这些可以促进脂动员的激素称为脂解激素(lipolytic hormones)。
胰岛素和前列腺素等与上述激素作用相反,可抑制脂动员,称为抗脂解激素(antilipolytic hormones)。
脂动员生成的脂肪酸可释放入血,与白蛋白结合形成脂酸白蛋白运输至其它组织被利用。
但是,脑及神经组织和红细胞等不能利用脂肪酸,甘油被运输到肝脏,被甘油激酶催化生成3-磷酸甘油,进入糖酵解途径分解或用于糖异生。
贮存于脂肪细胞中的甘油三酯
胰岛素 前列腺素E2 烟酸
肾上腺素 去甲肾上腺素 胰高血糖素
-
+
激素敏感脂肪酶
脂肪动员的基本过程
脂解激素-受体
+
AC
ATP
cAMP↑
HSLa(无活性) + PKA
HSLb(有活性)
甘油二酯脂肪酶
甘油三酯 (TG)
甘油二酯 (DG) FFA
甘油一酯 (MG) FFA
甘油一酯脂肪酶
FFA 甘油
脂肪动员的结果是生成:
一分子的甘油。
➢ 甘油可在血液循环中自由转运。 ➢ 脂肪动员生成的甘油主要转运至肝再磷酸化为
3-磷酸甘油后进行代谢。
三分子的自由脂肪酸(free tty acid, FFA)
➢ 脂肪酸进入血液循环后须与清蛋白结合成为复 合体再转运。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
理论价值和潜在临床应用价值
• ATGL在脂肪组织中功能研究
ATGL在脂肪组织中的作用与HSL功能研究密不可分。
Villen等将小鼠标记HSI的cDNA转染COS-7细胞,发现细胞中 的甘油三醋、胆固醇酷等被分解为FFA,而在COS-7细胞超表 达ATGL时,发现增加了培养基中的FFA,降低了胞内TAG的储 存量,而磷脂的含量变化不大,Lake等研究显示,293HEK细 胞中超表达ATGL时,TAG变化与上述相同。
随后该课题组在2004年发现了这个后来被命名为ATGL的脂肪酶 (一个在脂肪组织高度表达的54kDa的486氨基酸蛋白)。随后的一 系列研究不仅证实ATGL在基础状态和应激状态具有促进脂解作 用,而且发现与HSL缺乏动物相比,ATGL缺乏动物更易肥胖, ATGL缺乏可导致来自白色脂肪组织的脂肪酸减少75%以上,甘 油三酯在组织和器官快速积累,导致代谢异常。
此外,ATGL转录后水平受到怎样的调节机制,如何确立ATGL活 性所需的磷酸化作用途径等都将是今后重要的研究方向。大量的 研究表明,ATGL在脂代谢中的关键作用可能与肝脂肪变性、胰 岛素抵抗及其它代谢并发症存在着内在联系。
文章在组成结构上与其他文献的异同
首先,作为普通的文献,一般可分为以下几个结构,介绍, 实验材料和方法,实验结果和讨论与分析。这几个部分相对 来说练习都是不太大的,都是中规中矩的切合主题的。但是 这篇文章却打破了这些结构,从一开始的introduction 开始 引出问题,然后带领读者一步一步的发现进而证实ATGL的存 在和其生物学活性,将文章通过作者的思路和研究方法联系 起来,并且将作者自己的观点和讨论融入到里面。但是全文 看起来又是和一般的文献一样有理有据。
脂肪分解的实验中,HSL感染细胞胞浆提取物中没有观察到DG 积累。
使用联合提取剂的LacZ-,ATGL-,和HSL-转染细胞的三油酸甘 油酯的水解作用试验证实ATGL和HSL在酯解级联中有底物特 异性。
研究方法及研究思路
为了确定脂肪酶的细胞内定位将在脂质小滴上参与到 TG动员 是否对ATGL 适用,我们构造了一个腺病毒载体[编码His标记 的小鼠ATGL]并在分化的第八天用它来感染3T3-L1脂肪细胞。
ATGL在脂肪细胞能明显增强基本和异丙肾上腺素刺激的脂解。
考虑到PKA在调控脂肪细胞脂肪分解的核心作用,我们测试了 ATGL是否是PKA-介导磷酸化的一个目标
ATGL可以被磷酸化,但相反的是,HSL的这种修改没有通 过PKA介导。
结合已有研究后认为,ATGL对甘油三酯的特异性比甘油二酯高, 它分解甘油三酯的能力比HSL强,而HSL在甘油二酯分解为甘油 一酯和脂肪酸过程中具有重要作用,最后在甘油一酯酶作用下 完全分解为甘油和脂肪酸。从这个理论来看,ATGL不仅在甘油 三酯分解过程中起主要作用,而且甘油三酯分解产生的甘油二 酯是HSL促脂解的主要底物。无论在基础条件下还是在刺激条 件下,ATGL活化对脂肪组织脂解具有重要作用,HSL是甘油三 酯脂解的限速酶的观点受到了挑战。
研究方法及研究思路
2002年,Haemmerle等发现,在HSL基因敲除情况下,甘油三酯 脂解下降仅为40%,而甘油二酯大量堆积,推测可能存在另一种 酶对甘油三酯分解具有作用。 敲除HSI基因的小鼠体重未见异常增加,且保留了基础和异丙肾 上腺素刺激下脂肪分解的能力,基因敲除小鼠出现脂肪细胞内甘 油二酯堆积。并研究发现HSI与甘油二酯作用的特异活性是其与 TG的1O倍,提示存在其他甘油三酯脂肪酶。
理论价值和潜在临床应用价值
• 关于ATGL 、HSL与PLIN 的临床研究
临床研究发现,PLIN在肥胖症患者体内的表达降低,肥胖患 者皮下脂肪组织中HSL mRNA 的表达和HSL的最大脂解能力 都减少。我国学者通过对55例住院患者切取脂肪组织检测发 现,胰岛素抵抗状态时人皮下脂肪组织中ATGL和HSL的表达 减低,在早期可减少脂肪分解和FFA 的产生,减轻胰岛素抵 抗。
thanks
但随着TG在不同组织的沉积,不仅加重胰岛素抵抗,而且引 起糖代谢和脂代谢的异常 。还有研究发现,皮下和内脏脂肪 组织ATGL mRNA的表达量与空腹胰岛素水平及胰岛素敏感性 相关。通过测定2型糖尿病患者的血清ATGL水平发现,2型糖 尿病患者的血清ATGL 显著升高 。
理论价值和潜在临床应用价值
• ATGL与肥胖、糖尿病的关系
文献中未解决的问题及将来的研究方向
有关ATGL激活机制的研究发现,在白色脂肪组织HSL缺乏或抑制 条件下,机体仍会表现出激素刺激下的脂解反应,这提示我们 ATGL的活化也是直接或间接通过激素信号途径实现的。但是进 一步的研究发现,ATGL激活的分子学机制与HSL也存在一定差异, 首先与HSL不同的是在基础状态和激活状态下脂滴处的ATGL没有 差别;另外,虽然ATGL也可被磷酸化,但不是以PKA为目标,因 此,有关ATGL激活及促脂解的分子学机制还需要研究。
ATGL、HSL
HSL
TG
甘油二酯
三酰甘油水解酶-2 +FFA
甘油一酯+FFA
甘油+FFA
单酰甘油脂酶
背景知识
• 脂肪动员过程
围脂滴蛋白A包围在脂滴表面,保护脂滴中的TG不被水解。 儿茶酚胺作用于肾上腺素受体,使腺甘酸环化酶活性增高, 导致环磷酸腺苷(cAMP)增多,蛋白激酶A(PKA)被激活,磷酸 化围脂滴蛋白A和HSL。磷酸化的围脂滴蛋白A破坏脂滴表面 的分子屏障,使得磷酸化的HSL移位到脂滴表面,发挥其功 能。ATGL也可被磷酸化,但其磷酸化不是通过PKA途径, PKA通过磷酸化围脂滴蛋白A间接影响ATGL。CGI-58为 ATGL的共激活因子,内源性CGI-58与围脂滴蛋白结合,包被 在脂滴表面,PKA通路激活时,围脂滴蛋白磷酸化后失去和 CGI-58结合的能力,CGI一58脱离脂滴与ATGL结合,使 ATGL的活性提高近20倍。
文献中未解决的问题及将来的研究方向
近年来,应用基因敲除及突变模型等技术,人们对ATGL的生物 学功能及调控机制有了较多的认识,发现其不仅在脂肪组织脂解 过程中扮演关键脂肪酶,而且在非脂肪组织脂质和能量代谢过程 中也起到了重要的作用。作为一个重要的调控脂解蛋白分子, ATGL在细胞内受到精密的网络调节,激素、CGI.58、GOS2及 脂滴相关蛋白等均参与其中,从转录水平、翻译后水平等对 ATGL表达及活性进行着复杂的调控,但它们调控ATGL的具体作 用机制目前仍不清楚。
表明了ATGL有酶促活性 放射性标记的脂质底物的活性测定
证实
ATGL的转染COS-7细胞的细胞质组分只有TG水解酶(增加3.7 倍)发现了增长。
ATGL是一个TG水解酶。
和小鼠ATGL一样,人类ATGL也对放射性标记的TG基板和约 束的脂肪酶抑制剂表现出明显的脂肪分解活性。
研究方法及研究思路
ATGL的低DG水解酶的活性也在我们测得的脂解反应产物的相对 丰度的实验中证实。
综上所述
ATGL作为一个新的脂肪代谢调节关键酶为深入研究脂肪分解通 路翻开了崭新的一页,ATGL可能是今后研究开发调控脂肪分解 新药的靶点。将来有待于研究的领域很多,如ATGLmRNA表达 水平受禁食/进食、糖皮质激素和TNF一0l等多种因素影响的机 制,ATGL发挥作用的具体机制等,期待着开展该领域的一系列 研究,为预防肥胖、糖尿病等糖脂代谢紊乱性疾病提供重要的 依据。
生物化学阅读研讨汇报
制作人:张桓侨
甘油三酯脂肪酶
背景知识
1
2 研究方法及研 究思路
ATGL
3
5 献构文 的上章 异与在 同其组
他成 文结
背景知识Biblioteka • 脂肪动员过程甘油三酯(TG)的水解是一系列有序调控的过程,TG首先被 ATGL、HSL和三酰甘油水解酶-2水解成甘油二酯和游离脂肪 酸(FFA);其次,甘油二酯被HSL水解成甘油一酯和FFA,最后, 甘油一酯被单酰甘油脂酶水解成甘油和FFA。在这一过程中, 围脂滴蛋白和CGI一58(comparativegeneidentification-58)也起着 关键作用。
在Zimmermann等研究中表明,ATGL在3T3-Ll脂肪细胞中 超表达使甘油和FFA的释放量都增加,并在HSL基因敲除鼠 中进一步发现,ATGL拥有75%酞基水解酶活性,这说明 ATGL参与脂肪组织的脂解过程。
理论价值和潜在临床应用价值
• ATGL在脂肪组织中功能研究
2006年,Schweiger等通过采用敲除ATGL基因或者HSL基因 的突变小鼠模型,以及应用HSL分子抑制剂HSL(76-0079)研 究发现,HSL(76-0079)对HSL基因敲除小鼠的TAG脂解过程 没有太大影响,相反基本中断了ATGL基因敲除小鼠体内游离 脂肪酸动员过程;并随后进一步证实,ATGL和HSL在小鼠白色 脂肪组织中共拥有95%TAG水解酶活性,这表明两者均为 TAG分解过程的关键脂肪酶,同时也说明TAG脂解过程可能还 需要少量已知或未知水解酶类的参与,具体机制还需进一步 探索。
研究方法及研究思路
筛选小鼠和人类的基因和蛋白质数据库,再将该蛋白质的结构对 应到已知脂肪酶,备选项目都进行了[TG-水解酶在小鼠脂肪组织中 的活性和表达]的分析。只有一个以前未曾描述过的酶符合这些要 求,即
研究方法及研究思路
• 调查是否 ATGL 水解中性脂成分
转染 Western免疫印迹技术 SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE )分析 荧光自显影