【精品文档】端粒和端粒酶
端粒端粒酶和肿瘤-精选文档

端粒酶与肿瘤治疗(2)
端粒酶抑制剂治疗肿瘤的特点:
特异性、广谱性
目前这类研究已引起关注,尚有许多问题须解决 ?必须确定哪种正常细胞产生端粒酶,同时还必须评 估端粒酶对于这些细胞的重要性。 ?证明端粒酶的抑制剂是否能够如预期的那样破坏端 粒酶阳性的肿瘤。
端粒酶与肿瘤治疗(3)
? 为了研究开发抑制人体内端粒酶的药物,还必 须弄清楚这种酶究竟如何发挥功能,怎样结合到
此外,端粒的长短还和细胞分化程度相关,分化程度越高,
端粒DNA越短。如人胚细胞端粒平均长度10kb,人精细胞9kb, 而一般正常体细胞只有4kb
端粒、端粒酶与肿瘤 (1)
许多研究表明,在基因突变、肿瘤形成时,
端粒可表现出缺失、融合和序列缩短等异常 现象。 B细胞淋巴样白血病端粒融合易位型染色体
端粒与端粒酶-----端粒的概念
端粒(telomere)
是真核生物染色体线性 DNA分子末端的特殊结构,对 维持染色体稳定性和DNA完整 复制具有重要作用 。
端粒与端粒酶-----端粒的发现
20世纪30年代末:Mü ller首先提出了“端粒” 概 念
20世纪70年代: Olovnikov 提出染色体分子末端的 丢失 细胞退出细胞增殖 1978年Blackbum,et al. 发现四膜虫端粒分子末端 是重复 串联的5’ GGGGTT 3’ DNA简单序列 1985年Gueider,et al. 发现端粒酶
人的模板区序列:
5,CUAACCCUAAC-3,
模板区
与端粒的末端结合定
延伸复制的模板
端粒与端粒酶-----端粒酶的结构
发现四膜虫端粒酶是一种RNA-蛋白质复 合物(RNP结构),分子质量在200kd500kd,对RNA酶极为敏感,端粒酶内的 RNA和蛋白质对其活性似乎都是必需的
端粒与端粒酶

㈡ 端粒酶的两个主要功能
一 是端粒酶能自主地对端粒DNA富含G 的链进行延长,而富含G的链又能通过 GC配对使其终端回折形成特殊的发卡结 构,这样DNA复制时新链5′端缺失就可 以得到补齐,这就为真核生物解决了 DNA末端复制问题。
末端补齐机制:
端粒酶
GGGGTT 5‘ 5’ G-C配对回折 (GGGGTT)n
hsp90
端粒酶结合蛋白(TEP
)
TEP1、生存动力神经细胞基因(SMN) 产物、
、
PinX1、 Est1p 和Est3p
端粒酶是在染色体末端不断合成端粒序 列的酶,它可以维持端粒的长度,维持细胞 增殖潜能。端粒酶以自身RNA为模板合成端 粒酶重复序列,具有逆转录酶活性,它的活 性不依赖于DNA聚合酶,对RNA酶、蛋白酶 和高温均敏感。端粒酶活性表达能稳定端粒 的长度,抑制细胞的衰老,在生殖细胞和干 细胞中可检测到高水平的端粒酶活性。
㈠ 端粒酶的结构 端粒酶在结构上为一核糖核蛋白复合体,由RNA 和 结合的蛋白质组成,是RNA依赖的DNA 聚合酶。它是一种 特殊的能合成端粒DNA的酶,通过明显的模板依赖方式每 次添加一个核苷酸。
端粒酶实质上是一种特殊的逆转录酶
端粒酶RNA(hTR)
端粒酶逆转录酶(TERT)
端粒酶结合蛋白(TEP)
端粒酶延长端粒长度以减慢细胞衰老最早 的 证 据 来 自 Bodnar 等 的 研 究 , 1 9 9 8 年 其 在 Science 上刊文报道:将人的端粒酶基因导入 端粒酶阴性的正常人体细胞中激活其表达并培 养细胞,然后与未导入该基因的细胞比较,发 现前者端粒明显增长,细胞分裂旺盛,细胞寿 命比后者大大延长,更令人关注的是细胞并无 肿瘤样改变。
保护染色体结构和功能的完整性
端粒和端粒酶

端粒和端粒酶一、端粒的定义和功能端粒是染色体末端的DNA序列,它们在细胞分裂过程中起到保护染色体稳定性的重要作用。
端粒由重复的DNA序列重复单元组成,其中最典型的是人类端粒的DNA序列为(TTAGGG)n(n为重复次数)。
端粒的主要功能在于防止染色体末端的损失和减缩,并保护染色体免受DNA损伤和酶解攻击。
二、端粒酶的作用2.1 端粒酶的结构端粒酶是一种酶类,负责在细胞分裂过程中补充端粒的缺失。
它是由两个基本亚单位组成的复合酶,其中一个亚单位称为TERT(telomerase reverse transcriptase),负责提供酶的催化活性,另一个亚单位称为TERC(telomerase RNA component),是一种RNA分子,提供了端粒序列的模板。
2.2 端粒酶的功能端粒酶的功能主要是通过在DNA末端添加端粒序列来延长端粒。
在染色体复制过程中,DNA的末端由于DNA聚合酶的工作机制无法完全复制,因此每次细胞分裂时,染色体的末端会逐渐缩短。
而端粒酶通过添加端粒序列,可以提供一个模板,使得DNA末端得到延长,从而保护染色体的稳定性。
三、端粒和生物老化的关系3.1 生物老化的原因生物老化是指生物体随着年龄的增长而出现的功能下降和衰老的过程。
而端粒的缩短是导致生物老化的重要原因之一。
随着细胞分裂的进行,端粒的长度会逐渐减少,当端粒长度减到一定程度时,染色体末端的保护功能将会丧失,导致染色体的不稳定性和衰老。
3.2 端粒和细胞衰老的关系细胞的衰老也与端粒的缩短密切相关。
当细胞的端粒长度减小到一定程度时,细胞进入衰老状态,无法继续分裂和增殖。
这种细胞衰老被称为克隆老化。
克隆老化意味着细胞失去了再生能力,这在许多年龄相关疾病的发生过程中起到了重要的作用。
四、端粒酶与癌症的关系4.1 端粒酶在癌症中的表达在正常细胞中,端粒酶的表达很低,因此端粒长度会逐渐减少,导致细胞衰老。
而在癌细胞中,端粒酶表达水平显著增高。
端粒和端粒酶1

一、端粒和端粒酶二、端粒端粒是真核生物染色体末端由许多简单重复序列和相关蛋白组成的复合结构,具有维持染色体结构完整性和解决其末端复制难题的作用。
端粒酶是一种逆转录酶,由RNA和蛋白质组成,是以自身RNA为模板,合成端粒重复序列,加到新合成DNA链末端。
在人体内端粒酶出现在大多数的胚胎组织、生殖细胞、炎性细胞、更新组织的增生细胞以及肿瘤细胞中。
正因如此,细胞每有丝分裂一次,就有一段端粒序列丢失,当端粒长度缩短到一定程度,会使细胞停止分裂,导致衰老与死亡。
但是许多问题用端粒学说还不能解释。
体细胞端粒长度与有丝分裂能力呈正比,这一点实验已经证实了,而不同的体细胞其有丝分裂能力是不尽相同的,胃肠黏膜细胞的分裂增殖速度就比较快,神经细胞分裂的速度就比较慢。
曾有人就不同年龄供体角膜内皮细胞的端粒长度进行研究发现角膜内皮细胞内端粒长度长期维持在一个较高的水平,而端粒酶却不表达。
另外,Kippling发现,鼠的端粒比人类长近5-10倍,寿命却比人类短的多。
这些都提示体细胞端粒长度与个体的寿命及不同组织器官的预期寿命并非一致。
生殖细胞的端粒酶活性长期维持较高的水平却不会象肿瘤那样无限制分裂繁殖;端粒长度由端粒酶控制,那何种因素控制端粒酶呢?生殖细胞内端粒酶活性较高,为什么体细胞中没有较高的端粒酶活性。
看来端粒的长度缩短是衰老的原因还是结果尚需进一步研究。
2009年,诺贝尔瑞典卡罗林斯卡医学院将诺贝尔奖生理学或医学奖授予美国加利福尼亚旧金山大学的伊丽莎白·布莱克本(ElizabethBlackburn)、美国巴尔的摩约翰·霍普金斯医学院的卡罗尔-格雷德(CarolGreider)、美国哈佛医学院的杰克·绍斯塔克(JackSzostak)以及霍华德休斯医学研究所,以表彰他们发现了端粒和端粒酶保护染色体的机理。
端粒酶英文名称:telomerase一种反转录酶,由蛋白质和RNA两部分组成核糖蛋白复合体,其中RNA是一段模板序列,指导合成端粒DNA的重复序列片段。
端粒与端粒酶PPT精选文档

细胞内正常复制。这一方面证实了端粒对染色体的保
护作用,也使 DNA的大片段克隆成为可能,为后来
的人类基因组测序奠定了基础。
6
一、端粒与端粒酶的发现
1984年,伊丽莎白在试验中发现了一 个有趣的现象:不论是四膜虫还是酵 母自身的端粒序列都可以在酵母中被 保护和延伸。而带着四膜虫端粒DNA 的人工染色体进入到酵母后,复制后 被加上的是酵母端粒序列而非四膜虫 的端粒序列。
17
三、端粒酶的结构与功能
目前认为端粒酶主要由3个部分构成,即端粒 酶 RNA(telomerase RNA,TR)、端粒酶相关蛋 白质(telomerase — associated protein, TP1/ TP2) 和端粒酶逆转录酶(telomerase reverse transcriptase,TERT)。
长寿梦想的“天梯”
——端粒与端粒酶
主人:杨长友
重庆师范大学生命科学学院 生物化学与分子生物学专业
1
2009年诺贝尔生理学或医学奖
贡献:揭示了 “how chromosomes are protected by telomeres and the enzyme telomerase” (染色体是如 何被端粒和端粒酶保护的)。
9
一、端粒与端粒酶的发现
实验过程大致如下: 1、将底物寡聚核苷酸(端粒DNA、随机序列 DNA)进行放射性标记; 2、将高浓度的寡聚核苷酸底物与高浓度的四 膜虫细胞裂解液一起孵育; 3、通过放射性标记的核苷酸来检测体外端粒 序列的合成。
10
一、端粒与端粒酶的发现
结果显示,当四膜虫细胞裂解液加入四膜虫或 酵母端粒序列DNA时,其明显被重新加上了 DNA碱基,而且以 6个碱基递增的方式延长, 与四膜虫端粒重复基本单位为 6个碱基正好吻 合,而对于随机序列的DNA底物并不发生延伸。
端粒与端粒酶

• 1989年,一位Yale大学的研究人员公布了一项 科学发现,海拉癌细胞含有一种叫做端粒酶的物 质,能使细胞不死。这让控制生物衰老的神秘物 质——端粒酶走进了人们的视线。
参考文献
1.周宏远,肖林,严军,等.恶性肿瘤染色体端粒行为异常与 癌变发生关系研究【J】.肿瘤防治研究所,2001,28 2.Maruyama Y,Hanai H,Kaneko E.Telomere length and telomerase activity in intestinal metaplasia adenoena and welldifferentiated adenocarcinema of the stomach【J】.Nippon Rinsho,1998,56,(5):1186-1189 3.何剑,陈琼,易红,等.端粒酶的反义核苷酸抑制肺癌细胞端粒酶 活性的研究[J]. 中国现代医学杂志,2004,14(11):14-17.
端粒酶(Telomerase)
一种RNA依赖性的DNA聚合酶,能维持端粒酶的长度; 除了人体生殖细胞和部分体细胞外,端粒酶几乎对其他所有细胞不起作用,但它却能 维持癌细胞端粒的长度,使其无限增殖。
恶性肿瘤
在本质上是基因病; 研究发现,几乎所有的人体恶性肿瘤细胞中可检测到端粒酶的活性和端粒缩短。
端粒、端粒酶与恶性肿瘤发生的相关关系
的肿瘤特异性,是更加理想的肿瘤端粒酶抑制靶点
端粒酶:灵丹妙药Or学术骗局?
• 端粒酶(Telomerase),在细胞中负责端粒的延长的一 种酶,是基本的核蛋白逆转录酶。
成人体内的少数细胞可制作出这种巨型蛋白复合物,但制 作量非常之少,因此只有部分成分已被确定。 端粒酶仅仅解决了复制长度的问题,并不能解决DNA复 制时的变异问题。端粒能否决定生命长度,还在研究中, 尚未成定论。
端粒和端粒酶的研究进展

• 端粒和端粒酶简介 • 端粒和端粒酶的研究历史与现状 • 端粒和端粒酶与人类健康 • 端粒和端粒酶的实验研究方法 • 总结与展望
01
端粒和端粒酶简介
端粒的结构和功能
端粒的结构
端粒是由DNA和蛋白质组成的结 构,位于染色体末端,保护染色 体免受损伤和降解。
端粒的功能
端粒的主要功能是维持染色体的 稳定性和完整性,防止染色体融 合和降解,同时参与细胞分裂和 衰老过程。
相关疾病。
端粒和端粒酶的调控机制
03
目前,研究者们正在深入研究端粒和端粒酶的调控机制,以期
更好地理解其在细胞生命活动中的作用。
未来研究方向和展望
01
深入探究端粒和端粒酶的作用机制
未来研究需要进一步深入探究端粒和端粒酶的作用机制,以更好地理解
其在细胞生命活动中的作用。
02
开发基于端粒和端粒酶的治疗方法
未来可以开发基于端粒和端粒酶的治疗方法,用于治疗相关疾病。
03
加强跨学科合作与交流
未来需要加强跨学科合作与交流,促进端粒和端粒酶研究的深入发展。
03
端粒和端粒酶与人类健康
端粒和端粒酶与衰老
端粒与衰老
端粒是染色体末端的保护结构,随着细胞分裂次数的增加,端粒长度逐渐缩短, 导致基因组不稳定和细胞功能异常,最终引发衰老。
端粒酶与衰老
端粒酶是一种维持端粒长度的酶,通过激活端粒酶可以延长端粒长度,从而延缓 衰老过程。
端粒和端粒酶与疾病的关系
端粒与心血管疾病
心血管疾病患者中,端粒长度缩短与动脉粥样硬化、心肌梗死等 疾病的发病风险增加相关。
端粒与癌症
端粒酶的异常激活可以导致细胞无限增殖,形成肿瘤,因此与癌症 的发生和发展密切相关。
端粒和端粒酶

端粒和端粒酶09级食品科学与工程郭晓敏学号:170112009012一、端粒(一)端粒的发现1938年两位卓越的遗传学家Mc.Clintock和Muller分别以玉米、果蝇为材料发现经过各种途径损伤后所断裂的染色体很不稳定,容易降解,断口具有“粘性”,彼此之间很容易相互粘连而造成染色体易位、倒位、缺失、双着丝点等类型的染色体畸变。
而染色体的天然末段似乎从不与其它断口连接,也不彼此连接,这是因为染色体天然末端有类似“帽子”的结构,虽不含功能基因,但具有维持染色体稳定性的功能。
据此,Muller提出了“端粒”(telomere)这个术语。
(二)端粒的结构端粒( telomere) 是保护真核细胞染色体末端并维持其完整的特殊的DNA/蛋白质复合物,它像“帽子”一样扣在染色体的两端。
1.端粒DM的结构端粒DNA由两条长短不同的DNA链构成,一条富含G,另一条富含C。
富含G的那条链5’一3’指向染色体末端,此链比富含c的链在其3’末端尾处可多出12~16个核苷酸的长度,即3’悬挂链,一定条件下能形成一个大的具有规律性很高的鸟嘌呤四联体结构,此结构是通过单链之间或单链内对应的G残基之间形成H00gsteen碱基配对,从而使4段富含G的链旋聚成一段的四链体DNA。
也有人认为,端粒G链序列可以形成稳定的发卡结构,它和四联体结构都被认为与端粒DNA的保护功能有关.2. 端粒结合蛋白的结构端粒结合蛋白包括端粒酶、保卫蛋白复合体( sheherin) 和非保卫蛋白。
保卫蛋白复合体由端粒重复序列结合因子l(TRFl),端粒重复序列结合因子(TRF2)。
端粒保卫蛋白1( P0rrl),TRFl相互作用核蛋白(TIN2),TIN2相互作用蛋白l(TINTl )及阻抑和活化蛋白1趣印1)组成,各分布在染色体端粒上,能保持端粒结构的相对稳定。
非保卫蛋白有DNA修复蛋白RADSO,NBSl ,胍E11,Ku86和DNAPKcs等,各分布不局限在端粒上。
端粒和端粒酶

(一种有纤毛的单细胞池糖微生物)进一步揭示了这一端粒的 初步结构,发现它是由几个核苷酸(富含G)组成的DNA重复 片断,重复的次数由几十到数千不等。
3、1972年,Watson发现了一个问题:他发现模板(template) 和引物(primer)的存在,而且只能从5`到3`端方面合成。 这样,在引物被去除之后,细胞每分裂一次,在新合成的
因为端粒长度的缩短而导致细胞分裂的停止的机制尚不清楚, 目前有三种理论: 1. 第一种认为不断缩短的端粒最终将会影响到邻近的基因。 2. 第二种认为端粒DNA的完全丢失将会产生损伤的信号,这将会 激活P53。 3. 第三种认为不是损伤的信号,而是缩短了的端粒本身激活了 P53。
二、端粒酶
1、概念: 端粒酶是一种RNA与蛋白的复合体,
它以自身RNA上的一个片段为模板通过逆 转录合成端粒重复序列,并通过一种RNA 依赖性聚合酶(如逆转录酶)机制加到染 色体3’末端以延伸端粒。 2、组成:RNA(作为模板)
蛋白质(反转录酶)
3、作用机制: 在端粒DNA的复制时,端粒酶既有
模板,又有逆转录酶这两方面的作用。其 与端粒3´端结合后,以其RNA为模板, 经
➢ 端粒酶可以延长正常细胞的生命周期并且加速人类干 细胞复制的能力: 正常的细胞在其生命周期中只能分裂60-100次,而端 粒酶可以促进这个能力,显著的延缓衰老的进程。
➢ 端粒酶活动受细胞类型的特异性调节 端粒酶在体细胞中很少表达,但在骨髓、肿瘤细胞、 外周血白细胞如部分粒细胞、T细胞、单核细胞/B细胞 和精子细胞这些高增殖的细胞中却相当活跃。
DNA互补链的5`端必将留下一个小空缺(一段端粒)。(见 图1)这就不能解释DNA从头(denovo)合成的问题。这样, 细胞染色体长度会越来越短,稳定性越来越差,直至最终细
端粒及端粒酶

端粒及端粒酶:它们的作用机制和它们的功能转变效应摘要:端粒及端粒酶的分子特性在大多数真核生物中是保守的。
那么,端粒和端粒酶是怎样作用的,它们怎样相互作用来提高软色体稳定性,我们将在此讨论。
关键字:端粒、端粒酶1,简介端粒,真核生物软色体末端的DNA—蛋白质复合物,保护着基因组免受不稳定因子的侵扰。
这些因子包括软色体终端区域的降解,端粒的溶解,要么随着其它端粒,或者一段断裂的DNA末端,或者不恰当的重组。
这些过程都是潜在的灾难;例如,端粒溶解能导致爽双着丝粒染色体形成,这是不稳定遗传的,并且导致分裂细胞子代遗传容量的不稳定或者遗传信息的丢失。
端粒DNA由串联重复序列,简单的,经常富含—G—,该序列特异性的被活化的端粒酶识别。
这些串联重复形成了一个包含许多端粒蛋白结合位点分子“脚手架”,这就使高度次序结合的复合物依次成核,尽管还难以定义,这些保护性的端粒蛋白包括端粒DNA序列特异性结合蛋白。
端粒处生成的DNA—蛋白复合物是有活力的;在分裂间期,端粒结合蛋白间歇性的,以大约数分钟一个的速率相互交换,取决于哪一个蛋白组件被检查。
2,端粒酶:聚合酶和保护器端粒DNA的完全复制需要端粒酶,一种专门的胞内核糖核蛋白RNP反转录酶(RT)。
它的核心酶包含TERT蛋白,这种蛋白不但含有一个RT同源域,还有其它必要的保守域和一段RNA组件TER。
通过复制一小段内部RNA模板序列,端粒酶从5’—3’向软色体末端合成端粒DNA链,从而得到延长。
被调节的软色体DNA终端延长弥补着由于核酸酶作用和不完全的终端DNA复制造成的缩短。
一套多组分的“端粒内稳态”系统,一方面阻止着端粒的过分延长。
联合的端粒蛋白间的相互作用对这种稳态作用非常重要,这也在顺式端粒中起作用。
相反的,这种端粒稳态系统起着在端粒缩短时端粒的延长作用,从而在含有端粒酶的细胞中保持端粒重复序列处在在一个合适的范围内。
端粒酶还作为一种胞内RT来合成端粒中短的,重复的DNA序列。
端粒与端粒酶

染色体是真核生物遗传物质的载体, 染色体是真核生物遗传物质的载体,维持其稳定性对于 高等生物至关重要.然而,作为生命蓝图的承载者, 高等生物至关重要.然而,作为生命蓝图的承载者,染 色体却处在一个危机四伏的环境中, 色体却处在一个危机四伏的环境中,经常遭到细胞内多 种不利因素的干扰和破坏.可以想象, 种不利因素的干扰和破坏.可以想象,如果得不到有效 保护, 稳定性难以维持, 保护,DNA稳定性难以维持,后果将不堪设想.染色体 稳定性难以维持 后果将不堪设想.
探索一步步深入
1984年,Carol与Liz精心讨论设计实验,用四膜虫的核抽提液与体外的 1984 端粒DNA进行温育,试图在体外检测到这个"酶"活性,看到端粒的延伸 。经过不断优化条件,尤其是把底物换成体外合成的高浓度的端粒DNA 后,同年的圣诞节终于清楚地看到了"酶"活性。在测序胶的同位素曝光 片上,端粒底物明显被从新加上了DNA碱基,而且每六个碱基形成一条 很深的带,与四膜虫端粒重复基本单位为六个碱基正好吻合。这种酶活 性不依赖于DNA模板,只对四膜虫和酵母的端粒DNA进行延伸,而对随机 序列的DNA底物不延伸;并且该活性不依赖于DNA聚合酶。由于同源重组 对序列没有特异性的要求并且依赖于DNA聚合酶的活性,至此,她们澄 清了这两种假说,证明了有一种"酶"来延伸端粒DNA。这种酶后来被命 名为"端粒酶"(telomerase)。
探索一步步深入
20世纪五六十年代 20世纪五六十年代,当科学家们尝试解析真核生物如何实现染色体DNA的精 世纪五六十年代 确复制时,又一个端粒相关的难题产生了。DNA聚合酶进行每一轮线性DNA的 复制都会导致少量末端核苷酸的丢失,其结果是,真核生物线性染色体,作 为基因的载体,会在细胞分裂过程中逐渐缩短。1972年,James Watson提出 了"末端复制问题",他同时推测,真核生物需要一种特殊机制来确保线性染 色体末端的完整复制。同时,Alexey Olovnikov也推测,染色体末端的逐渐 缩短将导致细胞的衰老。 20世纪七十年代 20世纪七十年代,Hayflick首次提出将体外培养的正常人成纤维细胞的“ 世纪七十年代 有限复制力”作为细胞衰老的表征。在此过程中,细胞群中的大部分细胞 经历了一定次数的分裂后便停止了,但它们并没有死亡,仍保持着代谢活 性,只是在基因表达方式上有一定的改变。于是Hayflick猜测细胞内有一 个限制细胞分裂次数的“钟”,后来通过细胞核移植实验发现,这种“钟 ”在细胞核的染色体末端——端粒。
端粒与端粒酶
端粒酶结合蛋白(TEP
)
TEP1、生存动力神经细胞基因(SMN) 产物、
、
PinX1、 Est1p 和Est3p
端粒酶是在染色体末端不断合成端粒序 列的酶,它可以维持端粒的长度,维持细胞 增殖潜能。端粒酶以自身RNA为模板合成端 粒酶重复序列,具有逆转录酶活性,它的活 性不依赖于DNA聚合酶,对RNA酶、蛋白酶 和高温均敏感。端粒酶活性表达能稳定端粒 的长度,抑制细胞的衰老,在生殖细胞和干 细胞中可检测到高水平的端粒酶活性。
㈠ 端粒酶的结构 端粒酶在结构上为一核糖核蛋白复合体,由RNA 和 结合的蛋白质组成,是RNA依赖的DNA 聚合酶。它是一种 特殊的能合成端粒DNA的酶,通过明显的模板依赖方式每 次添加一个核苷酸。
端粒酶实质上是一种特殊的逆转录酶
端粒酶RNA(hTR)
端粒酶逆转录酶(TERT)
端粒酶结合蛋白(TEP)
端粒酶延长端粒长度以减慢细胞衰老最早 的 证 据 来 自 Bodnar 等 的 研 究 , 1 9 9 8 年 其 在 Science上刊文报道:将人的端粒酶基因导入 端粒酶阴性的正常人体细胞中激活其表达并培 养细胞,然后与未导入该基因的细胞比较,发 现前者端粒明显增长,细胞分裂旺盛,细胞寿 命比后者大大延长,更令人关注的是细胞并无 肿瘤样改变。
端粒
染色体
细胞分裂
端粒
细胞分裂
细胞将停止分裂而趋于老化
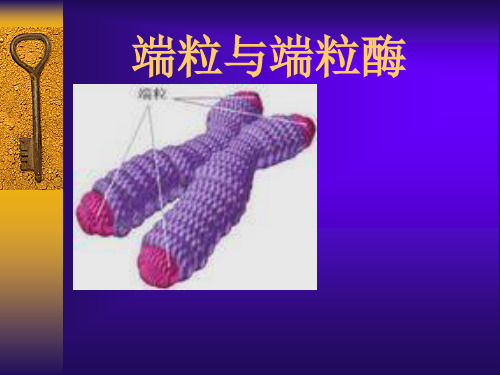
端粒:是真核细胞线性染色体末端特殊结构。
由端粒DNA和端粒相关蛋白组成。 端粒DNA:为不含功能基因的简单、高度重复序列, 在生物进化过程中具有高度保守性。
不同物种的端粒DNA 序列存在差异。
人类及其它脊椎动物染色体端粒的结构是 5′TTAGGG3′的重复序列, 长约15kb。体细胞的端粒有 限长度(telomere restriction fragments TRFS)大多数 明显短于生殖细胞,青年人的TRFs又显著长于年长 者,提示TRFs随着细胞分裂或衰老,在不断变短, 主要是由于DNA聚合酶不能完成复制成线性DNA末 端所致。
【精品文档】端粒和端粒酶

Assumptions
染色体的自然末端不同于非正常的 DNA断裂末 端,它有一个特殊的结构来避免染色体之间的 相互融合 .
科学家Hermann Muller 将这种特殊结构命名为 端粒(Telomere, 在希腊语中 ,telos表示末 端,meros 表示片段 ).
Experiments
Jack Szostak
人工染色体
1980年,Elizabeth Blackburn在会议上对这一重 大发现的报告,引起了Jack Szostak的极大兴趣。 他那时候正试图在酿酒酵母 中构建人工线性染色体,让 它能够在细胞中像自然染色 体一样复制。但当环状质粒 线性化并转入细胞后,很快 就被降解掉。
人工染色体
端粒序列的发现使Jack Szostak有机会把 线性质粒末端连接到四膜虫的端粒DNA , 然后再倒入酵母细胞中。奇迹发生了:线 性质粒不再被降解,而是在细胞内稳定存 在并复制。 这是人工染色体的最早雏形,它使得DNA 的大片段克隆成为可能,后来为人类基因 组测序工作立下了汗马功劳。
Elizabeth Blackburn 和 Carol Greider 的实验
四膜虫在接合细胞的大核发育过程中,大核产生 了非常丰富的小染色体,每一个小染色体都在末 端加上了端粒,可以推测:如果“酶”的假说成立, 此时细胞内的酶活性应该是非常高的.
Elizabeth Blackburn 和 Carol Greider 的实验
用四膜虫的核抽提液与体外的端粒 DNA进行温 育,试图在体外检测到这个酶的活性 ,看到端粒 的延伸 .
Elizabeth Blackburn 和 Carol Greider 的实验
经过不断优化条件,尤其是把底物换成体外合成的高浓 度的端粒DNA后,Carol通过曝光x光片,清楚地看到了 “酶”的活性:在测序胶的同位素曝光片上,端粒底物明 显被重新加上了DNA碱基,而且每六个碱基形成一条很 深的带,与四膜虫端粒重复基本单位为六个碱基正好吻 合.
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
端粒酶RNA亚基的发现
Daniel Gottschling利用TPE效应设计出实验,筛选酵母RNA亚基 基因: 把URA和ADE连个基因通过遗传重组的方法置入端粒区域,由于 TPE效应,URA和ADE不表达. 酵母只能在含有uracil的培养基中生长,且克隆是红色的.
再转入cDNA表达库,某些调控端粒长度的基因通过过表达可以改 变端粒的长度.如果端粒长度变得足够短,TPE效应就会消失,URA 和ADE两个基因就会启动表达. 这种短端粒的酵母能够在不含有uracil的培养基中生长,并且酵母克 隆显示出白色
端粒酶催化亚基
端粒酶既然能够利用RNA模板亚 基来复制DNA,那么很容易推测 这个蛋白亚基可能具有RNA dependent DNA polymerase 活性(依赖于RNA的DNA聚合酶 活性),即逆转录酶活性.更进一步 说,它的蛋白质序列里应该包含逆 转录酶特有的结构区域.
用四膜虫的核抽提液与体外的端粒DNA进行温 育,试图在体外检测到这个酶的活性,看到端粒 的延伸.
Elizabeth Blackburn 和 Carol Greider 的实验
经过不断优化条件,尤其是把底物换成体外合成的高浓度 的端粒DNA后,Carol通过曝光x光片,清楚地看到了“酶” 的活性:在测序胶的同位素曝光片上,端粒底物明显被重 新加上了DNA碱基,而且每六个碱基形成一条很深的带, 与四膜虫端粒重复基本单位为六个碱基正好吻合.
人工染色体
端粒序列的发现使Jack Szostak有机会把 线性质粒末端连接到四膜虫的端粒DNA , 然后再倒入酵母细胞中。奇迹发生了:线 性质粒不再被降解,而是在细胞内稳定存 在并复制。 这是人工染色体的最早雏形,它使得DNA 的大片段克隆成为可能,后来为人类基因 组测序工作立下了汗马功劳。
端 粒----稳定线性染色体的末端结构
Def. 存在于染色体末端的一段 特殊的DNA重复序列
端粒和端粒结合蛋白组成核蛋 白复合物,广泛存在于真核生 物细胞中;
不同种类细胞的端粒重复单位 不同,大多长约5—8bp;
端粒的结构与功能具有保守性.
端粒的功能
保护染色体 末端
A
B
防止染色体 复制时丢失
决定细胞寿 命
Cபைடு நூலகம்
E
TPE效应
固定染色体 位置
为什么染色体的自然末端不容易相互融合?
Assumptions
染色体的自然末端不同于非正常的DNA断裂末 端,它有一个特殊的结构来避免染色体之间的相 互融合.
科学家Hermann Muller将这种特殊结构命名 为端粒(Telomere,在希腊语中,telos表示末 端,meros表示片段).
Experiments
D
端粒酶活性的发现
Liz女士的实验室在研究四膜虫端粒序列的过程中还发现 了一个有趣的现象:带着四膜虫端粒DNA的人工染色体 导入到酵母后,被加上了酵母的端粒而不是四膜虫的端 粒序列.
由于端粒是由重复序列组成的,当时人们普遍猜想同源重 组是延伸端粒补偿染色体末端隐缩的机制.但是同源重 组只能复制出更多本身的序列,为什么在四膜虫端粒上 加的是酵母的端粒序列而不是四膜虫本身的序列呢?这 个现象是同源重组无力解释的.
Nobel Prize in Medicine Awarded for Creaking DNA Puzzle
端粒酶亚基
Liz实验室与Tom Cech合作,对端粒酶活性进一步定性. 用RNA酶处理样品,降解样品的RNA→酶活性消失 用蛋白酶消化→酶活性消失
端粒酶RNA亚基的发现
RNA亚基有一段RNA序列正好和四膜虫的端粒DNA序 列互补,端粒酶正式利用RNA亚基的这段序列作为模板 重复复制出端粒DNA
Jack Szostak
Elizabeth Blackburn
四膜虫
人工染色体
四膜虫
四膜虫是一种特殊的模式生物, 有两个细胞核。小核很稳定,含 5对染色体,用于生殖传代;而 大核在接合细胞的发育过程中, 染色体可断裂成200—300个小 染色体,端粒非常丰富。
Elizabeth Blackburn
Elizabeth Blackburn 和 Carol Greider 的实验
这种酶活性不依赖于DNA模板,只 对四膜虫和酵母的端粒DNA进行 延伸,而对随即序列的DNA底物不 延伸;并且该活性不依赖于DNA聚 合酶.
而同源重组对序列没有特异性要求, 并且依赖于DNA聚合酶的活性.
至此,她们澄清了这两种假说,证明 了有一种酶来延伸端粒DNA,即端 粒酶(telomerase)
Elizabeth Blackburn 和 Carol Greider 的实验
四膜虫在接合细胞的大核发育过程中,大核产生了 非常丰富的小染色体,每一个小染色体都在末端 加上了端粒,可以推测:如果“酶”的假说成立,此 时细胞内的酶活性应该是非常高的.
Elizabeth Blackburn 和 Carol Greider 的实验
【精品文档】端粒和端粒酶
Assumptions
1.复制前联体、环化(linear→circle) 2.发卡结构 3.可变末端(alterable terminal)多个短重复序列
(short repetitive sequences) 4.蛋白质干预
Puzzles 2
1939年,潜心玉米遗传性状研究的 Barbara McClintock (因发现玉米的转座子获诺贝尔学奖)注意 到:在减数分裂后期偶然产生的染色体断裂很容易重新 融合起来形成“桥”,在紧接着的有丝分裂中,这种染色 体“断裂---融合---桥---断裂”的循环不断继续。
1978年,Elizabeth Blackburn利用四膜虫纯化 了rDNA,以rDNA为模板通 过体外合成掺入dNTP的实 验,推断出四膜虫的染色体 上有由许多重复的5’CCCCAA-3’六碱基序列组 成的特殊结构区域,此区域 即端粒。
人工染色体
1980年,Elizabeth Blackburn在会议上对这一 重大发现的报告,引起了Jack Szostak的极大兴趣。 他那时候正试图在酿酒酵母 中构建人工线性染色体,让 它能够在细胞中像自然染色 体一样复制。但当环状质粒 线性化并转入细胞后,很快 就被降解掉。