发育生物学第十一章 脊椎动物胚轴形成
发育生物学8—17章课后习题答案

第八章神经系统发育1、神经胚形成答:神经胚形成:胚胎由原肠胚预定外胚层细胞形成神经管的过程。
神经胚:正在进行神经管形成的胚胎。
2、初级神经胚形成和次级神经胚形成答:初级神经胚形成:由脊索中胚层诱导上面覆盖的外胚层细胞分裂,内陷并与表皮质脱离形成中空的神经管。
次级神经胚形成:外胚层细胞下陷进入胚胎形成实心细胞索,接着在细胞索中心产生空洞形成中空的神经管。
3、什么叫神经板,神经褶,神经沟答:神经板:外胚层中线处细胞形状发生改变,细胞纵向变长加厚,形成神经板。
神经褶:神经板形成后不久,边缘加厚,并向上翘起形成神经褶。
神经沟:神经褶形成后在神经板中央出现的U型沟。
4、无脑畸形和脊髓裂与哪些基因有关,如何避免答:无脑畸形和脊髓裂均为人类胚胎的神经管闭合缺陷症。
人的后端神经管区域在27天时如不能合拢,则产生脊髓裂;若前端神经管区域不能合成,则胚儿前脑发育被停止,产生致死的无脑畸形。
它们与pax3、sonic hedghog和openbrain等基因有关。
约50%神经管缺陷可由孕妇补充叶酸加以避免。
5、斑马鱼的神经管如何形成答:斑马鱼的神经管如何形成:鸟类,哺乳类,两栖类动物胚胎的后端神经管及鱼类的全部神经管形成均采用次级神经胚形成的方式,所以斑马鱼的神经管形成也如此。
6、三个原始脑泡的发育命运答:前脑发育成为前端的端脑和后面的间脑,端脑最终形成大脑两半球,间脑形成丘脑和下丘脑区域及视觉感受区。
中脑腔最终形成大脑导水管。
菱脑再发育成前面的后脑和后面的髓脑,后脑形成小脑,髓脑形成延髓。
7、菱脑节答:菱脑节:在神经管闭合后,后脑前后轴逐渐被划分为8节,成为菱脑节,每个菱脑节是一个发育单位,节内细胞可交换而节间不能交换(其是临时性结构,到发育后期逐渐消失,但部分由后脑产生的结构如颜面神经节仍保持分节性结构)。
8、脊髓背腹区域细胞的发育命运各与哪些因子有关答:脊髓背部区域依次产生6种中间神经元(dI1-dI6),腹部则形成运动神经元和4种腹侧神经元(V0-V3)。
发育生物学——体轴与胚层
卵裂
卵裂期是指受精卵开始有丝分裂并产生由较小的细胞构成的囊胚(blastula) 的过程。主要特点包括: 分裂周期短; 分裂球的体积下降:海胆胚胎的质/核比由550降至6; 早期卵裂中合子基因大多处于休眠状态; 卵裂常经历由均等裂向不均等裂变化。
蛙的早期卵裂。 A 第一次卵裂, B 第二次卵裂, C 第四次卵裂,动物极和植 物极细胞出现差异。
小鼠胚胎上胚层细胞的程序性死亡导致囊胚腔的形成。
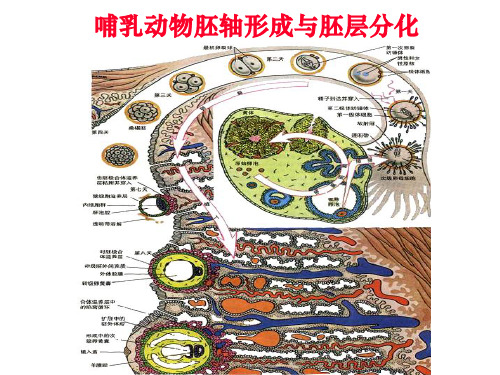
Gastrulation in the mouse embryo
6.5 d: beginning of gastrulation, the appearance of primitive streak; antero-posterior axis 7.5 d: primitive streak elongates until the bottom of the cup, forming notochord (head process)
哺乳动物胚泡的着床
胚泡在向子宫移动过程中体积增大,是由定位于滋胚层细胞 膜囊胚腔一侧的Na-K泵将外部Na+泵入腔中,最后通过渗透 作用吸水使囊胚腔增大。
随着囊胚腔内离子浓度的增加,水分就通过渗透作用进入进入囊胚腔内。囊胚 腔内液体的积累对囊胚腔壁造成一个向外的压力,这种静水压是参与形成和维持 囊胚为球形的一个重要作用力。
胚轴形成
• 胚胎细胞形成不同组织、器官,构成有序空间结构 的过程称为图式形成(pattern formation)。
• 在动物胚胎发育中,最初的图式形成主要涉及胚轴 (embryonic axes)形成及其一系列相关的细胞 分化过程。胚轴指胚胎的前-后轴(anterior posterior axis)和背 – 腹轴(dorsal -ventral axis)。胚轴的形成是在一系列基因的多层次、网 络性调控下完成的。
发育生物学试题库

发育生物学试题库(发育生物学教学组)目录:第一章章节知识点与重点........................第二章发育生物学试题总汇.......................第三章试题参考答案.............................第一章章节知识点与重点绪论1.发育和发育生物学2.发育的功能3.发育生物学的基础4.动物发育的主要特点5.胚胎发育的类型(嵌合型、调整型)6.研究发育生物学的主要方法第一章细胞命运的决定1.细胞分化2.细胞定型及其时相(特化、决定)3.细胞定型的两种方式与其特点(自主特化、有条件特化)4.胚胎发育的两种方式与其特点(镶嵌型发育依赖型发育)5.形态决定子6.胞质定域(海胆、软体动物、线虫)7.形态决定子的性质8.细胞命运渐进特化的系列实验9.双梯度模型10.诱导11.胚胎诱导第二章细胞分化的分子机制1.细胞表型分类2.差异基因表达的源由3.了解基因表达各水平的一般调控机制第三章转录后的调控1.RNA加工水平调控2.翻译和翻译后水平调控第四章发育中的信号传导1.信号传导2.了解参与早期胚胎发育的细胞外信号传导途径第五章受精的机制1.受精2.受精的主要过程及相关知识3.向化性4.顶体反应5.皮质反应第六章卵裂1.卵裂特点(课堂作业)2.卵裂方式3.两栖类、哺乳类、鱼类、昆虫的卵裂过程及特点4.(果蝇)卵裂的调控机制第七章原肠作用1.了解原肠作用的方式:2.海胆、文昌鱼、鱼类、两栖类、鸟类、哺乳类的原肠作用基本过程与特点第八章神经胚和三胚层分化1.三个胚层的发育命运第九章胚胎细胞相互作用-诱导1.胚胎诱导和自动神经化、自动中胚层化2.胚胎诱导、异源诱导者3.初级诱导和次级诱导、三(多)级诱导4.邻近组织相互作用的两种类型5.间质与上皮(腺上皮)的相互作用及机制第十章胚轴形成1.体形模式2.图式形成3.果蝇形体模式建立过程中沿前后轴不同层次基因的表达4.果蝇前后轴建立的分子机制5.果蝇背腹轴形成的分子机制第十一章脊椎动物胚轴的形成1.什么是胚轴2.两栖类胚轴形成过程及分子机制3.了解鸟类、鱼类、哺乳类动物胚轴形成过程及分子机制第十二章脊椎动物中枢神经系统和体节形成机制1.脊椎动物中枢神经系统的前后轴形成2.脊椎动物中枢神经系统的背腹轴形成3.脊椎动物体节分化特征第十三章神经系统的发育1.神经系统的组织发生神经系统的组成来源(神经管、神经嵴、外胚层板)中枢神经系统的组织发生(脊髓、大脑、小脑、核团)神经系统发生过程中的组织与调控(位置、数目)2.神经系统的功能建立3.神经突起(树突和轴突)4.局部有序投射5.突触第十四章附肢的发育和再生1.附肢的起源2.附肢的早期发育附肢发育中外胚层与中胚层的相互作用附肢发育中轴性建立3.附肢再生(再生过程、再生调节)第十五章眼的发育1.视泡发育、分化2.晶状体发育、分化3.晶状体再生4.角膜发育第十六章变态1.变态2.昆虫变态的激素调控3.两栖类变态的激素调控第十七章性腺发育和性别决定1.哺乳动物的性腺发育2.哺乳动物的性别决定3.果蝇的性别决定4.雌雄同体、环境性别决定第十八章生殖细胞的发生1.精子发生:特点,过程2.卵子发生:特点,过程第十九章干细胞1.干细胞2.干细胞分类3.了解干细胞的应用第二十章动物发育的环境调控1.发育与环境关系2.环境对正常发育的调控3.环境对正常发育的干扰4.遗传与环境之间的相互作用第二章发育生物学试题总汇一、填空题(每空1分)1.发育生物学研究的主要内容是个体发育和生物种群个体发生,其主要任务是研究生命体发育的本质过程及其调节机制。
第十一章 脊椎动物胚轴形成

脊椎动物胚轴形成
第一节 两栖类胚轴形成
• 组织者和Nieuwkoop中心
• 两栖类胚轴的形成的机制
一、组织者和Nieuwkoop中心
• (一)组织者 • 在原肠作用中,由组织者能诱 导背部外胚层形成中枢神经系 统的原基——神经管,并作用 于侧中胚层共同形成胚胎的背腹轴和前-后轴。
组织者细胞在早期胚胎发育中具有重要 的功能: • ①组织者能启动原肠作用; • ②组织者细胞有能力发育成为背部中 胚层包括前脊索板、脊索中胚层等; ③组织者能诱导外胚层背部化形成神 经板并使后者发育成为神经管; • ④组织者细胞能诱导其周围的中胚层 背部化分化成为侧板中胚层而不是腹 侧中胚层。
• TGFβ家族的成员在所有脊 椎动物左-右轴形成中起着 极其重要作用。
第三节 鸟类胚轴形成
• 鸟类背—腹轴形成 • 鸟类前—后轴形成 • 鸟类左—右轴的形成
一、鸟类背-腹轴形成
• 鸟类胚胎的体轴形成在原肠胚形成过 程中完成,但背-腹极性的特化开始于 卵裂期。鸟类的背-腹极性出现的标志 是囊胚期上、下胚层的分化。 • 囊胚期胚盘(blastoderm)的形成,事 实上是在上部的碱性清蛋白(pH9.5) 与下部的酸性胚下腔液(pH6.5)之间 建立起了一个屏障。
图11.2
爪蟾组织者的形成和Nieuwkoop中心
• 在囊胚期有两个背部信号中心对于胚胎的模 式形成起着重要作用,即位于动物帽区的前 组织者和内胚层中的Nieuwkoop中心 A. 囊 胚中期 B. 囊胚晚期 C. 原肠期
二、两栖类胚轴形成的机制
• (一)Nieuwkoop中心的分子生物学 研究 • β—Catenin是Nieuwkoop中心的一个 主要细胞因子。β—Catenin基因是一 种母源性信息,其编码的蛋白质β— Catenin是一种多功能蛋白,既能锚定 细胞膜上的钙黏蛋白,又是个核内转 录因子(图11.3)。
脊椎动物胚胎发育、皮肤、运动器官、消化器官

肩带:一般不与头骨和脊柱相连,主要骨块为肩胛骨、锁骨、乌喙骨 软骨鱼类:软骨棒 硬骨鱼类:肩带与头骨愈合 四足类:肩带不与头骨和脊柱相连,两栖类肩带为固胸型和弧胸 型;乌喙骨在哺乳类退化;鸟类锁骨V型;奔跑型动物 无锁骨 腰带:四足类三块:髂骨、坐骨、耻骨 因是承重的功能,结构稳定并永远与脊柱相连;随进化出现孔洞
爬行类
眶蝶骨 额 泪 眶上骨 完整的次生腭(鳄类)将口腔与鼻腔分开,使内鼻孔后移;骨化完全; 鳞骨颧突 骨骨 鼻 鳞骨 头骨为高颅型;自接型;单枕髁;具颞窝(鳄类完整双颞窝,鸟类双颞 骨 顶骨
窝,与眼窝愈合;龟鳖类为无颞窝型)
上枕骨
鸟 类
侧 头骨高颅型,且高度愈合,骨片轻,内有蜂窝状小腔;双颞窝(与眼窝 鼓骨 面 上颌骨颧突 颧骨 鼓泡 合),次生腭不完整;单枕髁 前颌骨
横突
带骨(肩带和腰带)和附肢骨(偶鳍和四肢骨)
肋骨
脊柱、肋骨及胸骨
脊 椎 动 物 脊 椎 骨 椎管
脉管
椎体的类型与椎间关节
双凹型 鱼、低等有尾类、低等爬行动物
后凹型
有尾类(蝾螈)、少数爬行类
前凹型
无尾类和爬行类
异凹型
即马鞍型;鸟类
双平型
哺乳类
脊 柱
脊椎骨产生顺序: 脊柱进化: 圆口类 鱼 类 两栖类 爬行类 鸟 类
哺乳类: 高度分化,结构机能无明显变化
体腔和系膜
体腔:中胚层侧板中间的空腔 鱼类--两栖类及绝大多数爬行类:围心腔1和胸腹腔1 鳄类和鸟类: 围心腔1、胸腔2、腹腔1(以结缔组织的斜隔分开) 哺乳类: 围心腔1、胸腔2、腹腔1(肌肉质的横膈 )
系膜:在体腔的背腹位置,为两层的腹膜 背系膜,腹系膜(因两侧体腔在腹部打通而大部分退化)
胚轴形成获奖课件

twist和dpp等基因旳激活解读dorsal蛋白旳浓度梯度
Dorsal蛋白旳功能
果蝇核蛋白dorsal沿背 • D沿L背蛋-腹白轴旳产浓生度区梯域度特经异过性对旳下位游腹置靶轴信基旳息因梯。旳度调控指,导控身制体 • 这种浓度梯度可活化腹侧组织分中为合子不基同因部tw分is进t (行tw发i)
和snail (sna)旳体现,用于中育胚旳层模分型化和原肠胚旳
形成,指导腹部构造旳发育;
• 同步克制腹部其他基因如dpp和zen基因旳体现,使其 只能在胚胎背侧体现,指导背部构造特化及其发育。
非洲爪蟾旳二、 两栖类胚轴旳建立
动轴1是-.植胚由物胎母极源动植物极轴旳决定-源于卵子
性•因两子栖决类定未受精卵沿动植物极存在一种极性,这 图示种母极源性性能够影响到将来卵裂旳方式。 旳•生未长受因精子卵旳极性是由沿动植物极分布旳母源性 VmgR-m1N基AR在因N爪旳A和蛋白质旳差别分布决定旳。 蟾•植爪物蟾极将旳来三个胚层旳区域划分在未受精卵就能 分布够。拟定。但胚胎旳前后轴、背腹轴和左右轴都
• 决定前后轴旳3组母体效应基因涉及: • 前端系统决定头胸部分节旳区域, • 后端系统决定分节旳腹部, • 末端系统决定胚胎两端不分节旳原头区和尾节。 • 另一组基因即背腹系统,决定胚胎旳背–腹轴。
胞质极性 (母体效应)
形态发生素
缺口基因(gap gene)
成对控制基因 (pair-rule gene)
母体效应基因产物
• 起源:在卵子发生过程中,这些母体效应基因旳mRNA由滋养 细胞合成转运至卵子,定位于卵子旳一定区域。
• 作用方式:这些mRNA编码转录因子或翻译调控蛋白因子,受 精后立即翻译,分布于整个合胞体胚盘中,激活或克制某些合 子基因旳体现,调控果蝇胚轴旳形成。
发育生物学试题库(答案参考)

发育生物学试题库(发育生物学教学组)目录:第一章章节知识点与重点 (1)第二章发育生物学试题总汇 (6)第三章试题参考答案 (18)第一章章节知识点与重点绪论1.发育和发育生物学2.发育的功能3.发育生物学的基础4.动物发育的主要特点5.胚胎发育的类型(嵌合型、调整型)6.研究发育生物学的主要方法第一章细胞命运的决定1.细胞分化2.细胞定型及其时相(特化、决定)3.细胞定型的两种方式与其特点(自主特化、有条件特化)4.胚胎发育的两种方式与其特点(镶嵌型发育依赖型发育)5.形态决定子6.胞质定域(海胆、软体动物、线虫)7.形态决定子的性质8.细胞命运渐进特化的系列实验9.双梯度模型10.诱导11.胚胎诱导第二章细胞分化的分子机制1.细胞表型分类2.差异基因表达的源由3.了解基因表达各水平的一般调控机制第三章转录后的调控1.RNA加工水平调控2.翻译和翻译后水平调控第四章发育中的信号传导1.信号传导2.了解参与早期胚胎发育的细胞外信号传导途径第五章受精的机制1.受精2.受精的主要过程及相关知识3.向化性4.顶体反应5.皮质反应第六章卵裂1.卵裂特点(课堂作业)2.卵裂方式3.两栖类、哺乳类、鱼类、昆虫的卵裂过程及特点4.(果蝇)卵裂的调控机制第七章原肠作用1.了解原肠作用的方式:2.海胆、文昌鱼、鱼类、两栖类、鸟类、哺乳类的原肠作用基本过程与特点第八章神经胚和三胚层分化1.三个胚层的发育命运第九章胚胎细胞相互作用-诱导1.胚胎诱导和自动神经化、自动中胚层化2.胚胎诱导、异源诱导者3.初级诱导和次级诱导、三(多)级诱导4.邻近组织相互作用的两种类型5.间质与上皮(腺上皮)的相互作用及机制第十章胚轴形成1.体形模式2.图式形成3.果蝇形体模式建立过程中沿前后轴不同层次基因的表达4.果蝇前后轴建立的分子机制5.果蝇背腹轴形成的分子机制第十一章脊椎动物胚轴的形成1.什么是胚轴2.两栖类胚轴形成过程及分子机制。
发育生物学名词解释(张卫红)

A.特异性蛋白质 B.DNA C.特异性 mRNA D中,错误的是
。
A.顶体反应是指受精前精子在同卵子接触时,精子顶体产生的一系列变化。
B.具有顶体结构的精子不发生顶体反应也可以进入卵子并与卵子融合
C.顶体反应释放的物质中含有大量的水解酶,因此顶体这一结构被认为具有类似溶酶体
46 容许的相互作用(primary competence):反应组织含有所有要表达的潜能,只需要环境, 但环境不能改变它的发育方向。许多组织需要含纤连蛋白和层粘连蛋白基质。 47 指令的相互作用次级感受性(secondary competence):反应组织的发育潜能不稳定,其发 育方向和过程取决于接受的诱导刺激类型。例如脊索诱导神经管底板细胞的形成。 48 感受性 competence:组织对一种特定刺激以一种特异方式产生反应的能力。它本身是一 种分化的表型,从空间和时间上区别细胞。 49 灰色新月: 精子入卵后,皮层向精子进入的方向旋转大约 30º,在动物极皮层含大量色 素而内层含有少量色素的物种中,这一胞质不同层次的相对运动形成了一个在精子进入点对 面的新月形的灰色区域,称为灰色新月。 50 神经嵴:当神经管与表面外胚层分开后,这些细胞向内迁移,侵入神经管两侧的成中胚 层细胞之间,形成一个很不规则的扁平细胞群,称神经嵴,介于神经管及其表面外胚层之间。 51 顶外胚层嵴(AER):随着鸟类和哺乳类的中胚层间质细胞进入肢区,它们分泌的因子诱
3.蛙类动物半球和植物半球汇合的背部边缘区(marginal zone)的
内陷,引发原肠作用。
A. 中胚层细胞 B.外胚层细胞
C. 动物极细胞 D.瓶状细胞
4. 晶状体来源于头部的
。
A.外胚层 B. 中胚层 C. 内胚层 D. 上胚层
发育生物学第十一章脊椎动物胚轴形成ppt课件

精子入卵后,皮层向精子进入的方向旋转大约30º
两栖类的灰色新月 精子入卵后,皮层向精子进入的方向旋转大约30º,在动 物极皮层含大量色素而内层含有少量色素的物种中,这一胞质不同层次的相 对运动形成了一个在精子进入点对面的新月形的灰色区域,称为灰色新月。
β-catenin是一种母体效应基因,其编码的蛋 白质β-CATENIN既能锚定细胞膜上的钙粘 着蛋白,又是一个核内转录因子。
β-CATENIN在受精时卵质的旋转移动过程 中在预定胚胎背部累积,在整个早期卵裂 阶段仍然主要在胚胎背部累积。到卵裂晚 期只有Nieuwkoop中心的细胞具有βCATENIN 。β-CATENIN 对于形成背部结 构是必要的。
两栖类胚胎的背- 腹轴和前-后轴是由受精时 卵质的重新分布而决定的。
a、背腹轴:受精时在精子入卵处的对面产生有色素 差异的灰色新月区,由此标志预定胚胎的背侧, 精子进入的一侧发育成为胚胎的腹侧。
b、前后轴:在动物极附近的背侧形成头部,而与其 相反的一侧形成尾,从而形成胚胎的背-腹轴和前 -后轴。
Spemann所示的蝾螈卵裂期的核等同性。
灰色新月区对 胚胎的正常发 育是必需的。
灰色新月区产 生的细胞形成 胚孔背唇(组 织者)。
Spemann等研究证实:
来源于灰色新月区细胞的胚孔背唇组 织不仅具有自我分化能力,还具有组 织和诱导邻近细胞开始原肠作用的能 力。
在原肠作用过程中,由组织者(organizer) 诱导背部外胚层形成中枢神经系统的原基— 神经管,并作用于侧中胚层共同形成背-腹 轴和前-后轴。 背唇细胞及其衍生物脊索和脊索中胚层虽然 还不足以作为整个胚胎的诱导者,在以后器 官原基形成和器官形成中都存在诱导作用, 但组织者启动了胚胎发育中的一系列诱导事 件。
脊椎动物体轴形成

脊椎动物体轴形成邹海玥 13级生物基地班 201300140153一、两栖类胚轴形成1、组织者和Nieuwkoop 中心两栖类是调整型发育的典型模型。
两栖类的背-腹轴和前-后轴是在受精时卵质的重新排列而决定。
灰色新月对胚胎的正常发育是必须的。
由灰色新月区产生的细胞形成胚孔背唇,随着背唇细胞的内陷原肠作用开始。
在原肠作用时,由于细胞重新排列,产生一系列邻近细胞之间的相互作用。
通过一系列的实验证明,组织者细胞不仅具有自我分化的能力,而且具有组织和诱导邻近细胞开始原肠作用的能力。
在原肠作用中组织者诱导背部的外胚层形成中枢神经原基——神经管,并作用于侧中胚层共同形成背-腹轴和前-后轴。
两栖类囊胚中最靠近背部的一群植物半球细胞,对组织者具有特殊的诱导能力。
Nieuwkoop 中心是由于精子入卵使卵质重排而产生的,兼具植物极和动物极细胞质的特殊区域,该区含有背部中胚层诱导信号。
如果抑制卵质旋转,胚胎将缺乏头和神经管等前端背部结构。
因此,两栖类的背-腹轴精子入卵时就决定。
Nieuwkoop在未来的发育中形成内胚层。
它具有诱导其上方的邻近细胞组织形成Spermann 组织者的能力。
使它发育成中胚层。
2、两栖类胚轴形成的机制(1)Nieuwkoop 中心的分子生物学机制爪蟾的内胚层细胞通过诱导预定中胚层细胞表达Xenopus Brachyury (Xbra)基因,进而诱导中胚层的形成。
最靠近背侧的植物半球细胞,由于表达一些特殊的细胞因子而形成Nieuwkoop中心。
β-catenin是Nieuwkoop中心的一个主要细胞因子。
GSK-3(糖原合成激酶-3 )对β-CATENIN 具有负调控作用,进而对于背侧细胞的分化起抑制作用。
如果在 1 个细胞期注射具有活性的GSK-3 ,以后胚轴的形成受到抑制。
如果在早期胚胎发育期抑制腹侧细胞内的GSK-3 ,则引起次性胚轴的形成。
β-CATENIN开始在整个卵内均匀的分布, 在腹侧细胞内因为GSK-3的存在而下降,而背侧细胞内因为DSH 的存在不会下降。
发育生物学名词解释(张卫红)
30 顶体反应(acrosomal reaction):是指受精前精子在同卵子接触时,精子顶体产生的一系 列变化。具有顶体结构的无脊椎动物或脊椎动物中,只有发生顶体反应的精子才能进入卵子 并与卵子融合,也只有精子与卵子接触时才发生顶体反应 31 顶体:位于精核前端,由高尔基体演化而来。顶体中含有多种水解酶,主要作用是溶解 卵子的外膜。有些动物的顶体中还有与精卵识别有关的分子。 32 精子获能( capacitation ):指射出的精子在若干生殖道获能因子的作用下,精子膜发生 一系列变化,进而产生生化和运动方式的改变的现象。
A.特异性蛋白质 B.DNA C.特异性 mRNA D.RNA;
6. 下列有关精子顶体反应的说法中,错误的是
。
A.顶体反应是指受精前精子在同卵子接触时,精子顶体产生的一系列变化。
B.具有顶体结构的精子不发生顶体反应也可以进入卵子并与卵子融合
C.顶体反应释放的物质中含有大量的水解酶,因此顶体这一结构被认为具有类似溶酶体
3.蛙类动物半球和植物半球汇合的背部边缘区(marginal zone)的
内陷,引发原肠作用。
A. 中胚层细胞 B.外胚层细胞
C. 动物极细胞 D.瓶状细胞
4. 晶状体来源于头部的
。
A.外胚层 B. 中胚层 C. 内胚层 D. 上胚层
5. 原位杂交是发育生物学研究常用的一种技术,该技术主要检测
在胚胎或组织中的分布。
46 容许的相互作用(primary competence):反应组织含有所有要表达的潜能,只需要环境, 但环境不能改变它的发育方向。许多组织需要含纤连蛋白和层粘连蛋白基质。 47 指令的相互作用次级感受性(secondary competence):反应组织的发育潜能不稳定,其发 育方向和过程取决于接受的诱导刺激类型。例如脊索诱导神经管底板细胞的形成。 48 感受性 competence:组织对一种特定刺激以一种特异方式产生反应的能力。它本身是一 种分化的表型,从空间和时间上区别细胞。 49 灰色新月: 精子入卵后,皮层向精子进入的方向旋转大约 30º,在动物极皮层含大量色 素而内层含有少量色素的物种中,这一胞质不同层次的相对运动形成了一个在精子进入点对 面的新月形的灰色区域,称为灰色新月。 50 神经嵴:当神经管与表面外胚层分开后,这些细胞向内迁移,侵入神经管两侧的成中胚 层细胞之间,形成一个很不规则的扁平细胞群,称神经嵴,介于神经管及其表面外胚层之间。 51 顶外胚层嵴(AER):随着鸟类和哺乳类的中胚层间质细胞进入肢区,它们分泌的因子诱
发育生物学复习资料重点总结

绪论1、发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
2、(填空)发育生物学模式动物:果蝇、线虫、非洲爪蟾、斑马鱼、鸡和小鼠。
第一篇发育生物学基本原理第一章细胞命运的决定1、细胞分化:从单个的全能细胞受精卵开始产生各种分化类型细胞的发育过程称细胞分化。
2、细胞定型可分为“特化”和“决定”两个阶段:当一个细胞或者组织放在中性环境如培养皿中培养可以自主分化时,可以说这个细胞或组织发育命运已经特化;当一个细胞或组织放在胚胎另一个部位培养可以自主分化时,可以说这个细胞或组织发育命运已经决定。
(特化的发育命运是可逆的,决定的发育命运是不可逆的。
把已特化细胞或组织移植到胚胎不同部位,会分化成不同组织,把已决定细胞或组织移植到胚胎不同部位,只会分化成同一种组织。
)3、(简答)胚胎细胞发育命运的定型主要有两种作用方式:第一种通过胞质隔离实现,第二种通过胚胎诱导实现。
(1)通过胞质隔离指定细胞发育命运是指卵裂时,受精卵内特定的细胞质分离到特定的裂球中,裂球中所含有的特定胞质可以决定它发育成哪一类细胞,而及邻近细胞没有关系。
细胞发育命运的这种定型方式称为“自主特化”,细胞发育命运完全由内部细胞质组分决定。
这种以细胞自主特化为特点的胚胎发育模式称为“镶嵌型发育”,因为整体胚胎好像是由能自我分化的各部分组合而成,也称自主型发育。
(2)通过胚胎诱导指定细胞发育命运是指胚胎发育过程中,相邻细胞或组织之间通过互相作用,决定其中一方或双方细胞的分化方向。
相互作用开始前,细胞可能具有不止一种分化潜能,但是和邻近细胞或组织的相互作用逐渐限制它们的发育命运,使之只能朝一定的方向分化。
细胞发育命运的这种定型方式成为“有条件特化”或“渐进特化”或“依赖型特化”,因为细胞发育命运取决于及其邻近的细胞或组织。
这种以细胞有条件特化为特点的胚胎发育模式称为“调整型发育”,也称有条件发育或依赖型发育。
胚轴的形成概论

bcd 基因编码的BCD蛋白是一种转录调节因 子。另一母体效应基因hunchback(hb)是 其靶基因之一, 控制胚胎胸部及头部部分结 构的发育。
hb在合胞体胚盘阶段开始翻译,表达区域 主要位于胚胎前部,HB蛋白从前向后也形 成一种浓度梯度。hb基因的表达受BCD蛋 白浓度梯度的控制,只有BCD蛋白的浓度 达到一定临界值才能启动hb基因的表达。
缺口基因、成对控制基因以及体节极性基 因共同调节同源异型基因(homeotic gene) 的表达,决定每个体节的发育命运。
母源性基 因系统突 变后产生 的结果 。
果蝇形体模式建成过 程中沿前后轴不同层 次基因内的表达。
不同组的基因 顺序表达沿前 后轴建立身体 的模式。
2. 前端组织中心
BICOID(BCD)蛋白浓度梯度
胚轴的形成是在一系列基因的多层次、网 络性调控下完成的。
爪蟾尾芽期胚胎的前后轴、背腹轴和左右轴(中侧 轴),互成垂直角度。
第一节 果蝇胚轴的形成
现已筛选到与胚胎前后轴和背腹轴形成有 关的约50个母体效应基因(maternal effect gene)和120个合子基因(zygotic gene)。 通过对这些基因的研究,我们对果蝇胚轴 形成的调控机制已有了一个较为清晰的认 识。
决定前后轴的3组母体效应基因包括:前端 系统(anterior system)决定头胸部分节的 区域,后端系统(posterior system)决定分 节的腹部,末端系统(terminal system)决 定胚胎两端不分节的原头区和尾节。
另一组基因即背腹系统(dorsoventral system),决定胚胎的背 – 腹轴。
DL蛋白定位于细胞 核中的机制。
cactus基因与DL蛋白能 否进入细胞核这一调控 过程有关。CACTUS与 DL结合时,DL蛋白不 能进入细胞核。
发育生物学第十一章 脊椎动物胚轴形成

β-CATENIN在背腹轴特化中的作用
β-CATENIN属于Wnt信号传导途径的一个分 子,糖原合成激酶-3 (GSK-3)对βCATENIN有负调控作用,进而对背侧细胞 的分化起抑制作用。在腹侧细胞中GSK-3介 导的磷酸化作用引起β-CATENIN的降解, 而在背侧细胞中由于存在GSK-3的抑制因子 Disheveled (DSH)蛋白,所以背侧的βCATENIN不会被降解。DSH蛋白开始存在 于爪蟾未受精卵植物半球的皮层部,受精 时沿微管迁移到胚胎的背侧起作用。
β– CATENIN 是WNT信 号途径中 的成分。
DSH将β – CATENIN定 位于胚胎背部 的机制1
DSH将β – CATENIN 定位于胚胎 背部的机制 2
DSH将β –CATENIN定位于胚胎背部的机制3
β-CATENIN与一种普遍存在的转录因子 TCF3结合形成的复合物能够激活对胚轴形 成具有重要作用的其他基因,如siamois (sms)基因。SMS能激活goosecoid基因的 表达。 goosecoid基因是Nieuwkoop中心分泌蛋白因 子作用的主要靶基因之一。gcd mRNA表达 的区域与组织者的范围有关。GCD能够激 活脑形成关键基因Xotx2,使其在前端中胚 层和预定脑外胚层表达。
第一类组织者分泌性蛋白因子和BMP抑制因子
BMP4是最重要的上皮分化和腹侧化诱导因 子。对于神经发生而言,它是抑制因子或抗 神经化因子,其功能与组织者正好相反。 组织者分泌的NGN、CHD、Nodal相关蛋白 -3和FST对BMP4均有抑制作用。因此组织 者的功能是分泌具有抑制作用的可溶性蛋白 因子,而不是直接诱导中枢神经系统的发生。
动物极帽是胚胎前端的标志。
发育7-2:脊椎动物形体模式形成

第二节脊椎动物形体模式的形成及基因控制两栖类是在胚胎体轴形成方面了解最多的脊椎动物。
脊椎动物胚轴的形成不仅与定位于囊胚期分裂球中的各种决定因子相关,更重要的是与其后发育阶段中细胞的一系列相互作用相关。
一、胚胎诱导的概念脊椎动物胚胎形体模式是通过相邻细胞间一系列的相互作用渐进式决定的,而不是象果蝇中那样是完全通过形态发生决定子的细胞质定域决定的。
胚胎细胞中这种细胞间的相互作用称为胚胎诱导(embryonic induction)。
胚胎诱导现象是比利时人伟大的实验胚胎学家Spemann发现的。
如前所述,受精卵中的灰色新月区域在在胚胎发育中具有重要的作用,缺少这一部分就不能进行原肠作用(figure 10. 19);如人工损伤这一部分,则将产生畸形胚胎。
灰色新月区是产生胚孔背唇的地方。
Spemann与他的学生Mongold为了进一步研究这一区域以及所产生的胚孔背唇的作用,将蝾螈一个胚胎的胚孔背唇移植到另一个蝾螈早期原肠胚的囊胚腔中(figure 10. 21),结果在这个受体胚胎的腹面诱导产生了第二个对称排列的胚胎。
这一实验证明了背唇(脊索中胚层)具有使外胚层发育成神经系统的能力。
他们也证明了如果在外胚层下面没有脊索中胚层,则此外胚层就不能发育为神经组织。
Spemann和Mongold在1924年发表了他们的研究结果。
这一研究结果在1935年获得了Nobel Prize。
并从20世纪30年代到现在,都是发育生物学和胚胎学最关注的重大科学问题之一。
近年来一些与胚胎诱导相关的分子的发现也是现代科学中最激动人心的事件。
胚胎诱导现象最先发现与蝾螈的胚胎发育中。
以后的实验又不断地证实在无尾两栖类、鸟类、鱼类文昌鱼以及无脊椎动物等的胚胎发生过程中均有这种诱导的现象。
说明胚胎诱导作用是动物胚胎发育过程中一个普遍的现象。
在胚胎发育中,一部分细胞对其邻近细胞的形态发生产生影响,并决定其分化方向的作用称为诱导作用或胚胎诱导(induction or embryonic induction)。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
不同物种之间胚胎图式形成的机制却具惊人的相似性
斑马鱼 的囊胚
Fate map of germ-layer progenitors at late blastula stage.
Fate map of dorsal–ventral tissue progenitors at late blastula stage.
爪蟾卵经紫外线照射 后引起背部结构的缺 失,可以由注射NGN 蛋白而挽救。NGN对 背部结构的诱导呈剂 量依赖性。
NGN在组织 者区的定位
Chordin mRNA在组织者区的定位
第一类组织者分泌性蛋白质因子:BMP抑制因子。
组织中心产生 的信号分子 (如Chordin、 Noggin、 Follistatin---神经诱导分子) 可拮抗腹部化 信号(如 BMP4)。
Spemann等研究证实: 来源于灰色新月区细胞的胚孔背唇组 织不仅具有自我分化能力,还具有组 织和诱导邻近细胞开始原肠作用的能 力。
在原肠作用过程中,由组织者(organizer) 诱导背部外胚层形成中枢神经系统的原基— 神经管,并作用于侧中胚层共同形成背-腹 轴和前-后轴。 背唇细胞及其衍生物脊索和脊索中胚层虽然 还不足以作为整个胚胎的诱导者,在以后器 官原基形成和器官形成中都存在诱导作用, 但组织者启动了胚胎发育中的一系列诱导事 件。
第十一章 脊椎动物胚轴形成
第一节 两栖类胚轴形成
两栖类是调整型胚胎发育的经典模型,对其 胚轴的形成机制有较多的了解。 脊椎动物胚轴形成的机制
A、定位于囊胚期大量分裂球中的各种决定因子 B、发育过程中发生在邻近细胞之间的一系列相互作 用。
Life cycle of the African claw-toed frog Xenopus laevis
爪蟾组织者和Nieuwkoop中心及中胚层诱导模型
二、两栖类胚轴形成的机制
1. Nieuwkoop中心的分子生物学研究
爪蟾的内胚层细胞通过诱导预定中胚层细 胞表达Xenopus Brachyury (Xbra)基因, 进而诱导中胚层的形成。 最靠近背侧的植物半球细胞,由于表达一 些特殊的细胞因子而形成Nieuwkoop中心。 β-CATENIN是Nieuwkoop中心的一个主要细 胞因子。
下包完成30%时的胚盘
下包胚盘的边缘区域的放大,示边缘下包的内卷和细胞内移
通过下包胚盘的边缘细胞内卷或上胚层细胞分层形成下胚层
斑马鱼原肠作用时细胞的集中和延伸把下胚层和上胚层细胞带 到背部形成胚盾,胚盾内脊索中胚层向动物极延伸。
移植的胚盾诱导受体 胚胎产生第二胚轴
Maternal organizers. The YSL is a source of a signal that induces marginal cell fates.
Pitx2决定心脏弯曲和肠盘绕的方向
第二节 鱼类胚轴形成
不同物种之间胚体图式形成的机制却具惊 人的相似性:在此发育过程中,都涉及母 体效应基因的产物-母体效应决定子的定 位、外源信号的影响和细胞间的相互作用 ,涉及一系列信号传导分子的作用。 有关这些信号传导机制的研究是目前最活 跃的领域之一。
β-CATENIN与TGF-β家族蛋白协同作用诱 导组织者形成的机制
总之,为维持正常的发育,在胚胎背部细 胞中必须含有β-CATENIN,并使sms基因表 达,SMS与TGF-β基因家族的蛋白质协同作 用使gcd基因激活,在SMS和GCD等共同作 用下才能形成组织者。 组织者的形成涉及多种基因的激活,存在 于Nieuwkoop中心的分泌蛋白激活位于其上 方中胚层细胞中一系列转录因子,后者再 激活编码组织者分泌产物的一些基因。
Hensen氏节由胚胎后端的Koller’s sickle形成
后端化因子 RA的作用
中侧轴特化因子
脊椎动物中侧轴形成的关键事件都是原肠 胚左侧侧板中胚层表达的nodal-相关基因调 控。 爪蟾的nodal-相关基因-1(Xnr-1)的表达区 域仅限于原肠胚左侧,Xnr-1激活pitx-2在胚 胎左侧专一性表达。VG1蛋白可以增加胚胎 左右对称的特化,异位表达VG1可导致胚胎 左右逆转。
背部中胚层诱 导形成组织者 的可能机制
gcd基因的激活也同时受到定位于植物半球和 Nieuwkoop中心的TGF-β家族蛋白产物 (Vg1、VegT和Nodal相关蛋白)的协同作用。 β-CATENIN与VG1、VEGT信号相互作用的 结果形成Nodal-相关蛋白从背侧到腹侧的浓 度梯度。Nodal-相关蛋白的浓度梯度使中胚 层细胞分化。含有大量Nodal相关蛋白的分化 为组织者,较少的分化为侧板中胚层,不含 Nodal相关蛋白的分化成腹侧中胚层。
β-CATENIN在背腹轴特化中的作用
β-CATENIN属于Wnt信号传导途径的一个分 子,糖原合成激酶-3 (GSK-3)对βCATENIN有负调控作用,进而对背侧细胞 的分化起抑制作用。在腹侧细胞中GSK-3介 导的磷酸化作用引起β-CATENIN的降解, 而在背侧细胞中由于存在GSK-3的抑制因子 Disheveled (DSH)蛋白,所以背侧的βCATENIN不会被降解。DSH蛋白开始存在 于爪蟾未受精卵植物半球的皮层部,受精 时沿微管迁移到胚胎的背侧起作用。
Brachyury基因在非洲爪蟾原肠胚时期预定中胚层细胞的表达
组织者的精细结构。组织者特异基因的表达,可以把 早期组织者细分成含有不同信息组合的亚区。
β-catenin是一种母体效应基因,其编码的蛋 白质β-CATENIN既能锚定细胞膜上的钙粘 着蛋白,又是一个核内转录因子。 β-CATENIN在受精时卵质的旋转移动过程 中在预定胚胎背部累积,在整个早期卵裂 阶段仍然主要在胚胎背部累积。到卵裂晚 期只有Nieuwkoop中心的细胞具有βCATENIN 。β-CATENIN 对于形成背部结 构是必要的。
动物极帽是胚胎前端的标志。
Axis formation in the zebrafish embryo. -3
Dorsal–ventral and mesendodermal patterning of the blastula embryo. Antagonists such as Boz, Chordin, and Dkk1 inhibit ventralizing BMP signals.
β– CATENIN 是WNT信 号途径中 的成分。
DSH将β – CATENIN定 位于胚胎背部 的机制1
DSH将β – CATENIN 定位于胚胎 背部的机制 2
DSH将β –CATENIN定位于胚胎背部的机制3
β-CATENIN与一种普遍存在的转录因子 TCF3结合形成的复合物能够激活对胚轴形 成具有重要作用的其他基因,如siamois (sms)基因。SMS能激活goosecoid基因的 表达。 goosecoid基因是Nieuwkoop中心分泌蛋白因 子作用的主要靶基因之一。gcd mRNA表达 的区域与组织者的范围有关。GCD能够激 活脑形成关键基因Xotx2,使其在前端中胚 层和预定脑外胚层表达。
Xwnt8引起中胚 层的腹部化并抑 制头部结构的形 成
Xwnt8在缘区的表达
注射CERBERUS(CBR)可诱导出两个头部结构
FRZB的表达与 功能。注射 FRZB引起躯干 部形成的抑制
7
外胚层分化为 上皮和神经组 织的不同机制
后端化因子 随着前端神经系统的区域性特化神经管, 后端开始进行分化。后端的分化由胚胎后 端产生后端化因子进行调控。 成纤维细胞生长因子eFGF、WNT3a和视黄 酸(Retinoic acid,RA)都是后端化因子。 RA主要作用于后脑的图式形成;eFGF对于 脊髓的分区最为重要;而WNT3a可以抑制 前端控制基因的表达,还可以协调RA和 eFGF的作用。
第一类组织者分泌性蛋白因子和BMP抑制因子
BMP4是最重要的上皮分化和腹侧化诱导因 子。对于神经发生而言,它是抑制因子或抗 神经化因子,其功能与组织者正好相反。 组织者分泌的NGN、CHD、Nodal相关蛋白 -3和FST对BMP4均有抑制作用。因此组织 者的功能是分泌具有抑制作用的可溶性蛋白 因子,而不是直接诱导中枢神经系统的发生。
爪蟾和其他脊椎动物胚胎前-后轴的形成在 背-腹轴形成之后,胚胎的背部一旦建立随 即开始中胚层细胞的内卷运动,并建立前后轴。 最先经过胚孔背唇迁入的中胚层细胞产生 前端结构,从侧唇和腹唇迁入的中胚层细 胞形成后端结构。
原肠作用过程中细胞内卷的动力来自深层内卷缘区细胞
Nieuwkoop中心
Nieukoop中心对正常发育是必需的
背方部分含有 Nieukoop中心, 形成一个背部 化的胚胎,但 缺少肠;腹方 部分没有 Nieukoop中心, 形成一腹部化 的胚胎,缺少 背部和头部结 构。
一、组织者和Nieuwkoop 中心
Spemann所示的蝾螈卵裂期的核等同性。
灰色新月区对 胚胎的正常发 育是必需的。 灰色新月区产 生的细胞形成 胚孔背唇(组 织者)。
Axis formation in the zebrafish embryo. -1
Axis formation in the zebrafish embryo. -2
鱼类胚胎前后轴的形成是由两个信号中心 调控的: A、第一个信号中心是原肠胚期神经外胚层和 表皮外胚层之间的一小团神经细胞分泌的 前端发育诱导因子。 B、第二个信号中心是原肠胚期胚盘边缘侧板 脊索中胚层的前体细胞产生的后端发育诱 导因子。
精子入卵后,皮层向精子进入的方向旋转大约30º
两栖类的灰色新月 精子入卵后,皮层向精子进入的方向旋转大约30º ,在动 物极皮层含大量色素而内层含有少量色素的物种中,这一胞质不同层次的相 对运动形成了一个在精子进入点对面的新月形的灰色区域,称为灰色新月。