04 基因突变类型、损伤修复(精选)
遗传基因突变与DNA损伤修复.pptx

2024年9月29日星期日2时40分20秒
第8页/共112页
4、突变的可逆性
• 正突变( forward mutation):A → a(u)
• 回复突变(back mutation):a → A(v)
2 X10-6
his+
his-
4 X10-8
2024年9月29日星期日2时40分20秒
第9页/共112页
2024年9月29日星期日2 时40分20秒
③DNA链断裂 这是电离辐射引起的严重损伤事件,断链数随照射剂量而增加。
④交联 包括DNA链交联和DNA-蛋白质交联
2024年9月29日星期日2 时40分20秒
第26页/共112页
(一)电离辐射 1、诱变机理
自发突变率:10-6左右 辐射诱变率:10-3左右 2、遗传效应
(1)辐射剂量与突变频率在一定范围内成正比
第二节 基因突变的表现及检出 2、果蝇突变型的检出 (1)果蝇性连锁突变的测定 CLB法:鉴别果蝇 X - 染色体上基因的隐性突变
2024年9月29日星期日2 时40分20秒
第15页/共112页
第二节 基因突变的表现及检 出
二、突变的检出
2、果蝇突变型的检出 (1)果蝇性连锁突变的测定 Muller - 5 法
一、 基因突变的分类 1、形态突变(morphological mutation):突变体的效应在表型上
明显可见。又称可见突变(visible mutation)
2、生化突变(biochemical mutantion):突变影响生物的代谢过程,
导致一个特定的生化功能的改变或丧失。
3、致死突变(lethal mutation):影响生物体的生活力,导致个体死亡
DNA的损伤、修复

在插入酶的催化下,以正确 插入酶的催化下, 的催化下 的碱基插入空位, 的碱基插入空位,修复 DNA
聚合酶的催化下 的催化下, 在 DNA 聚合酶的催化下,以 互补链为模板, 互补链为模板,合成新的单链 片段以填补缺口 片段以填补缺口
连接酶催化连接片段 催化连接片段, 由 DNA 连接酶催化连接片段, 封闭缺口
(一)引起DNA损伤的因素: 引起DNA损伤的因素: DNA损伤的因素 自发因素: 1.自发因素: 1).脱嘌呤和脱嘧啶 DNA分子通过自发水解 经常发生脱嘧 分子通过自发水解经常 DNA 分子通过自发水解 经常 发生脱嘧 啶和脱嘌呤反应, 啶和脱嘌呤反应 , 使嘌呤碱和嘧啶碱从 DNA分子的 脱氧核糖分子的脱氧核糖 DNA 分子的 脱氧核糖 - 磷酸骨架上脱落下 来。 例如,在腺嘌呤和鸟嘌呤的N 例如,在腺嘌呤和鸟嘌呤的N- 9及脱 氧核糖C 之间的N 糖苷键常发生自发 氧核糖C-1′之间的N-糖苷键常发生自发 水解反应而断裂, 从而失去嘌呤碱基, 水解反应而断裂 , 从而失去嘌呤碱基 , 使该嘌呤碱基所编码的遗传信息丢失。 使该嘌呤碱基所编码的遗传信息丢失。
特异性的核酸内切酶识别 特异性的核酸内切酶识别 DNA 核酸内切酶 的损伤部位, 的损伤部位,并在该部位的 5' 端作一切口
糖苷酶识别受损伤的 DNA 糖苷酶识别受损伤的 碱基, 碱基,并将该碱基切除
由核酸外切酶(或 DNA 聚合 核酸外切酶( 5'→3'端逐一切除损 酶Ⅰ)从 5'→3&#碱基的脱氨基作用 碱基中的胞嘧啶(C)、腺嘌呤(A)和 鸟嘌呤(G)都含有环外氨基,氨基有时 会自发脱落,从而使胞嘧啶变为尿嘧啶 (U),腺嘌呤变为次黄嘌呤(I), 鸟嘌 呤变为黄嘌呤(X)。 这些脱氨基产物的配对性质与原来的 碱基不同,即U与A配对,I和X均与C配对。 而且DNA复制时,它们将会在子链中产生 错误而导致DNA损伤。
基因突变——精选推荐

漫长的生物自然历史进化过程中,其中一些有利的或中性的突变,会随着生物的世代繁衍、交替而得以逐渐的稳定与累积。
这些突变的基因以及由此所引起的遗传性状变化,不仅是同种生物遗传性状多样性的根本渊源,而且也为不同物种的演化提供了丰富的原材料,并通过自然选择的作用而成为促进生物种系系统发育与不同种群产生、形成的原动力;而那些有害的突变基因,则会导致各种遗传性疾病的发生,构成和增加群体的遗传负荷。
发生在体细胞中的基因突变,即体细胞突变(somatic mutation),虽然不会传递给后代个体,但是却能够通过突变细胞的分裂增殖而在所产生的各代子细胞中进行传递,形成突变的细胞克隆(clone),成为具有体细胞遗传学特征的肿瘤病变甚或癌变的细胞组织病理学基础。
基因突变,一般具有以下几种主要特性:1.多向性任何基因座(locus)上的基因,都有可能独立地发生多次不同的突变而形成其新的等位基因,这就是基因突变的多向性。
譬如,在不同条件下,位于染色体某一基因座上的基因A可突变为其等位基因a1;也可以突变为a2或者a3、a4......a n等等其他等位基因形式,从而形成所谓的复等位基因(multiple alleles)。
遗传学上把群体中存在于同一基因座上,决定同一类相对性状,经由突变而来,且具有3种或3种以上不同形式的等位基因互称为复等位基因。
如大家所熟知的人类ABO血型系统,就是由位于9q34这一区域同一个基因座上的I A、I B和i三种等位基因形式所构成的一组复等位基因所决定的。
2.重复性基因突变的重复性是指:已经发生突变的基因,在一定的条件下,还可能再次独立地发生突变而形成其另外一种新的等位基因形式。
亦即,对于任何一个基因位点来说,其突变并非仅囿于某一次或某几次的发生,而是会以一定的频率反复发生。
例如:某一基因座上的基因A可突变为其等位基因a;基因a有可能独立地发生突变形成其新的等位基因a1;同样地,a1也可能再次地发生突变而形成其另外的等位基因a2;a2还可能突变为a3......,就其最终的群体遗传学效应而言,基因重复突变与基因多向突变的结果相似,也是群体中复等位基因存在的主要成因之一。
基因突变与损伤修复ppt课件

ppt课件.
O 28
电离辐射
• 作用机制
– 直接作用:能量直接作用于染色体和DNA – 间接作用:电离化引起细胞内化学变化导致异常
• 遗传学效应
– 辐射效应具有累加性 – 诱发基因突变和染色体畸变,突变率与剂量成正比
ppt课件.
29
电离辐射
• 过程
– 物理学阶段 – 物理化学阶段 – 生物化学阶段 – 遗传损伤
组的突变率明显上升,这些基因就是增变 基因
– DNA聚合酶的各个基因。 – dam基因,与错配修复等修复系统有关的基因。
ppt课件.
37
突变热点hot spots of mutations
• DNA的不同部分有着不同的突变率 • 突变位点在基因内的分布不随机
– 5-甲基胞嘧啶 – 短重复序列
• 与诱变剂性质有关
47复制到损伤部位时经过短暂的抑制产生一种新的dna聚合酶能催化损伤部位的dna的修复合成但识别碱基的精确度在新链中二聚体相对的位置上可以出现任何碱基其他位置上也可以出现错配
第十六章 基因突变
本章学时数:3学时 本章重点:1、突变及其特征、突变率
2、诱变 3、突变的 检出
ppt课件.
1
基因突变
• 定义:指一个基因变化成它的等位基因, 是染色体上一个 座位内的遗传物质的变 化。也称点突变(point mutation)。
配对
ppt课件.
17
碱基类似物
1. 5-溴尿嘧啶(5-bromouracil,5-BU) 2. 2-氨基嘌呤(2-aminopurine 2-AP) 3. 迭氮胸苷(AZT, azidothymidine)
ppt课件.
18
5-溴尿嘧啶
基因突变及修复课件
将待分离突变株的原 始菌株以合适的稀释 度涂布到野生型菌株 和突变株均能生长的 平板(含完全培养基), 培养形成单菌落
通过一消毒“印章”将 平板菌落分别原位移 位到含完全培养基的 平板(c)和某一营养物 质缺乏的平板(d)
培养后对照两个平板上形成的单菌落,在完全培养基的平板 (c)上生长而营养物质缺乏的平板(d)上不生长的菌落即所需分 离的突变型,挑选菌落在完全培养基上分离纯化
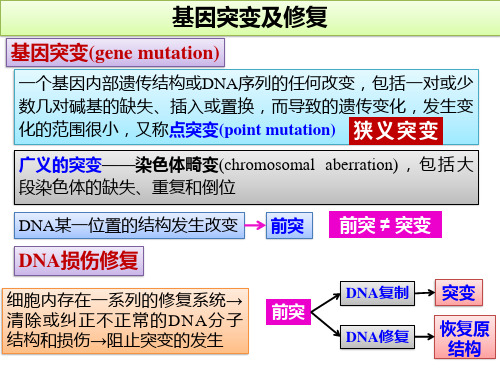
基因突变及修复
一、基因突变的类型及其分离 二、基因突变的分子基础 三、DNA损伤的修复
基因突变及修复
二、基因突变的分子基础
突变
自发突变
环境因素的影响,DNA复制过程的偶 然错误等而导致,一般频率较低,通 常为10-10~10-6
诱 变 某些物理、化学因素对生物体的DNA
进行直接作用,突变以较高的频率产生
2、诱发突变(induced mutation) 常 用 诱 变 剂
(1) 碱基类似物:5-溴尿嘧啶(胸腺嘧啶类似物)和2-氨基嘌呤(嘌 呤类似物)
(2) 插入染料:插入DNA分子的碱基对,使其分开,导致DNA 在复制过程中滑动→移码突变,如溴化乙锭、吖啶橙等染料
(3) 直接与DNA碱基起化学反应的诱变剂:如亚硝酸、羟胺和烷 化剂等
基本培养基+ 少量组氨酸 +少量诱变剂
原理:(his-)菌株在不含 组氨酸的培养基中不能 生长,或只有极少数的 自发回复突变子生长, 如果回复突变率因某种 化学诱变剂(或待测物) 的作用而增加,那么这 种化学药物可判断为具 有致癌性
基因突变及修复
二、基因突变的分子基础
3、诱变剂与致癌物质——Ames试验
基因突变及修复
一、基因突变的类型及其分离
DNA的损伤修复及突变

紫外线引起的DNA损伤 --最易形成胸腺嘧啶二聚体(TT)
19
2. 电辐射引起的DNA损伤
➢ 碱基变化
细胞中的水经辐射解离后产生大量OH-自由基,使 DNA链上的碱基氧化修饰、形成过氧化物的、导致碱 基环的破坏和脱落等。
➢ 脱氧核糖变化
脱氧核糖上的每个碳原子和羟基上的氢都能与OH-反 应,导致脱氧核糖分解,最后会引起DNA链断裂。
48
着色性干皮病(xeroderma pigmentosis,XP) 是一种切除修复有缺陷的遗传性疾病。
在研究其发病机制时,发现一些相关的基 因,称为 XPA、XPB、XPC等。这些基因的表达产物起辨认 和切除损伤DNA作用的。
XP病人是由于XP基因有缺陷,不能修复紫外 线照射引起的DNA损伤,因此易发生皮肤癌。
30
体外培养的肝癌细胞吖啶橙荧光染色
荧光显微镜下(选用蓝色激发滤片),可见含DNA的细 胞核显示黄绿色荧光,含RNA的细胞质及核仁显示橘红 色荧光。
31
第二节 DNA的突变
32
如果DNA的损伤得不到有效的修复,就会造 成DNA分子上可遗传的永久性结构变化,称为 突变(mutation)。少数突变甚至有可能对细胞 是有利的。有利突变的累积可以使生物进化,使 其能更好地适合于其生存的环境。但绝大部分突 变是有害的,对于单细胞生物,不少有害突变是 致死的,对于多细胞的高等生物,有害突变会造 成病变,如代谢病和肿瘤。
通常以酮式存在,有时也以烯醇式存在。当BU先以 酮式掺入DNA,继而又变成烯醇式时,进一步复制使 DNA中 A- T对变成 G- C对。同样道理也引起 G- C向 A- T的转换,BU可以使细菌的突变率提高近万倍。26
除BU外,还有5-溴脱氧尿苷、5-氟尿嘧啶、5氯尿嘧啶及它们的脱氧核苷。
生命科学中的DNA损伤和修复

生命科学中的DNA损伤和修复DNA是生命的基础,在细胞内起着至关重要的作用。
然而,它经常受到损伤,如紫外线、化学物质、辐射等。
如果未能及时修复则可能导致疾病甚至癌症。
今天我们将探讨生命科学中的DNA损伤和修复。
一、DNA的损伤类型DNA受到的损伤形式很多,如单链断裂、双链断裂、碱基损伤等。
其中,单链断裂是指DNA中所含的两股链中的一股断裂,而另一股仍保持完整;双链断裂则是两股链都发生了断裂。
碱基损伤则是指DNA中所含的碱基受到了各种各样的损伤。
二、DNA损伤的危害DNA损伤如果未能及时修复,会导致一系列的后果。
首先是基因突变,基因突变是指DNA序列的改变,如果发生在体细胞中则可能导致癌症等疾病,而在生殖细胞中则可能传递给下一代。
此外,DNA损伤还可能导致细胞自我毁灭,造成组织和器官的功能障碍或细胞死亡,比如神经细胞坏死可导致疾病如阿尔茨海默病。
三、DNA修复机制为了避免DNA损伤的危害,细胞需要拥有多种修复机制。
DNA修复机制包括:直接反应、碱基切割修复、核苷酸切割修复和重组修复等。
(1)直接反应直接反应是一种不需要酶辅助的修复机制,它包括:碱基漂移、钯磁场旋转、邻近值作用、光诱导电子转移和物理效应等。
直接反应通常只适用于少数特定类型的损伤,比如UV损伤等。
(2)碱基切割修复碱基切割修复是指损伤的DNA区域被切割,并进行修复。
核苷酸切割打破DNA链,第一步是DNA糖-磷酸链的切割。
这种切割在大多数引起DNA损伤的情况下都是必要的,它使损伤DNA从未损伤的DNA中被分离出来,并允许损害部位被清除。
然后核酸内切酶切割DNA链,在某些情况下,切割DNA链就足够修复损伤了。
在另一些情况下,取代碱基系统需要参与。
(3)核苷酸切割修复核苷酸切割修复可以修复一些单链损伤和大多数双链断裂。
核苷酸切割修复可分为两种类型:全切割修复和片段切割修复。
全切割修复在夜间进行,就是把受损的DNA拉出来,然后把它切成很多小块,每块都用基因片断替换掉它。
DNA损伤修复与基因突变的发生

DNA损伤修复与基因突变的发生DNA是构成生命体的基础,是生命的重要组成部分。
但是在生命的过程中,DNA受到了各种外界环境因素的干扰,如放射线、紫外线、化学物质等,这些因素会导致DNA受损和突变。
然而,出现了DNA损伤,如何修复和维护DNA的稳定性成为了一个重要的问题。
本文将从DNA损伤与修复的关系、基因突变的发生机制以及DNA修复和突变应用方面对这一问题进行讨论。
DNA损伤与修复的关系DNA在生物体内为避免损伤的发生,具有详细的修复系统和机制,以保持DNA的完整性和稳定性。
DNA损伤主要分为两类:基因突变和染色体畸变。
前者是DNA的单个碱基或小片段发生突变,如:DNA碱基损伤(如硝基化、脱氨基、酸化等)和DNA单链断裂;后者是染色体片段的变异、插入、缺失等。
多种机制负责DNA修复,包括基础修复、核苷酸切换修复、异源结束修复、同源重组修复以及DNA电子传输修复等多个层面和路径。
每个机制都具有独特的异同,各自适用于不同类型、不同程度的DNA损伤。
基础修复机制又称为直接修复,适用于对DNA双链断裂具有良好修复能力,重要的是它可以修复简单而单一的损伤。
对于单个碱基损伤,真核生物细胞通常通过两个酶进行修复:O6-甲基-鸟嘌呤-DNA甲基转移酶和构成II的酶。
O6-甲基-鸟嘌呤-DNA甲基转移酶负责从DNA中将甲基基团转移到基团中,而构成II的酶,则负责恢复鸟嘌呤的结构。
因此,基础修复机制对DNA的双链断裂修复很有帮助。
核苷酸切换修复机制是针对DNA中插入的碱基和缺失的碱基进行修复。
这种机制通过分解未成对碱基,在一个互补核苷酸插入到未成对的位点上,在DNA聚合酶的作用下修复了某一段DNA序列的完整性。
此机制对大量、复杂碱基损伤有良好作用。
异源结束修复机制主要利用两条不同的染色体,通过交换DNA中的部分,来维持信号序列的完整性。
正确地,它主要用于DNA双链断裂、复制错误等比较复杂情况下DNA的修复。
同源重组修复机制则是一种在DNA双链断裂情况下的修复机制,主要通过病变染色体找到另一个同源的染色体,以其作为模板,在染色体的可替代区域引进修复甚至重组事件。
第4章 DNA损伤、修复及重组

• TT 、 CC 、 CT 之间都可 形成二聚体。
• 紫外线引起的DNA损伤最易形成胸腺嘧啶二聚 体(TT)。
(2)电离辐射造成的DNA损伤 • 碱基的变化:细胞中的H2O经辐射分解后产生•OH, 使碱基氧化修饰,形成过氧化物,导致碱基的破 坏和脱落。 • 脱氧核糖的变化:•OH可以使脱氧核糖分解,引 起DNA单链或双链断裂。 • DNA的交联,包括DNA链间交联和DNA与蛋白质 的交联。
• 以胸腺嘧啶二聚体为例,含有二聚体的DNA仍可 进行复制,但复制到二聚体时要暂停一下,然后 越过此处障碍,在二聚体的后面以未知的机制开 始复制,这种起始复制可能不需引发。 • 这样在合成的子链上留下一个大缺口,而其互补 链则复制成完整的双链。然后由完整双链中的母 链与带缺口的子链发生重组。
• DNA合成时,复制叉遇到嘧 啶二聚体,会跳跃过损伤部 位,在下游约1000个核苷酸 处重新开始。复制的子链形 成一个缺口。 • recA蛋白可以识别并结合于 此,并同时识别同源双链区。 • 在recA蛋白的作用下,同源 双链发生重组交换,无损伤 的母链断裂修补缺损的空隙。 • 母链的缺损部位以互补链为 模板,进行合成修复。
(四)烷基转移修复
• 烷化剂所引起的最常见的DNA损伤时使鸟嘌呤O6 甲基化。在大肠杆菌中存在 O6-甲基鸟嘌呤转移酶, 可修复甲基化的碱基。将O6甲基鸟嘌呤的甲基转 移至该酶的一个半胱氨酸上,酶自身失活。
二、切除修复
• 切除修复是指在一系列酶的作用下,将DNA分子 中受损伤部分切除,然后以另一条完整的互补链 为模板,重新合成切去的部分,是DNA恢复正常 结构的过程。
5´ 3´
T
G
GATC
GATC
3´ 5´
CH3
dNTPs
第五章 DNA损伤修复和基因突变基因突变

•
•
3 基因突变
• 一个正常的生物体叫作野生型(wild type, WT) • 如果DNA发生改变,就会使生物体的某 些性状有所改变,这种改变了性状的生 物体相对于正常的生物体来说,就成为 突变体(mutant)。
• 所有的组织都有可能随机地与环境反应产生 突变,这种突变叫做自发突变(spontaneous mutagenesis)。 • 自发突变的发生率对每个组织来说都是特征 性的,这种特征是背景水平(background lever)的突变。 • 使用了诱变剂的突变叫诱发突变(induced mutagenesis)。
•
¾2.1.2错配修复系统
• 错配修复(mismatch repair)对DNA复制忠 实性的贡献率达10-2-10-3 ,DNA子链中的错 配几乎完全被修正,这充分反映了母链的 重要性。 • 该系统识别母链的根据来自Dam甲基化酶, 它能使位于5‘GATC序列中腺苷酸的6N位甲基 化。
• 一旦复制叉通过复制起始位点,母链就会 在开始DNA合成前的几秒至几分钟内被甲 基化。 • 只要两条DNA链上碱基配对出现错误,错 配修复系统就会根据“保存母链,修正子 链”的原则,找出错误碱基所在的DNA链, 并在对应于母链甲基化腺苷酸上游鸟苷酸 的5’位置切开子链,再根据错配碱基相对 于DNA切口的方位修复路径,合成新的子 链片段。
次黄嘌呤
Lac I,复制平均错误率10-9 DNA复制中的错误水平10-7—10-11 研究发现有一些基因的突变可以大 大提高整个基因组其它基因的突变率,这些 基因被称为增变基因(mutator genes)。
多个碱基改变造成的突变
转座子插入、重组错误
•
突变如何对表型产生影响
同义突变(synonymous mutation)指没有改变 产物氨基酸序列的密码子变化,与密码子的简 并性有关。 错义突变 ( missense mutation )指碱基序列的 改变引起了产物氨基酸序列的改变。 无义突变(nonsense mutation或null mutation)指 某个碱基的改变使代表某种氨基酸的密码子变 为蛋白质合成的终止密码子,导致肽链合成过 早终止。
基因突变与DNA损伤修复

06
了物种的基因的相对稳定性。
07
突变的性质
③可逆性 1) 野生型基因 突变型。 2) 回复突变率一般低于正突变率。 3) 真正的回复突变很少发生。常被第二个位点的突变所抑制,抑制因子突变(suppressor mutation)。 4)回复突变的有无是区别基因突变与染色体缺失或重复的标志。
真菌营养缺陷型的检出-菌丝过滤法
A
分生孢子→诱变处理→基本培养基 ↙ ↘ 未萌发孢子 萌发菌丝 培养 去除(过滤) ↙ ↓ ↘ 少数 死亡 营养 野生型 孢子 缺陷型 (去除) ↘ ↙ 营养培养基
B
果蝇突变体的检测
果蝇X连锁隐性突变的检出
ClB法 ClB品系:l—隐性致死基因;B—棒眼 C—交换抑制因子(B-l倒位);
错义突变: 碱基序列的改变引起了产物氨基酸序列的改变。
有些错义突变严重影响蛋白质活性甚至完全无活性,从而影响了表现型。如果该基因是必须基因,则称为致死突变。
有些错义突变不影响或基本上不影响蛋白质的活性,不表现明显的性状变化,称为中性突变。
③按其发生的原因:
自发突变(spontaneous mutation):在自然情况下发生的突变。
移码突变:增加或减少一个或几个碱基对的突变,引起密码编组的移动。 eg:Hb Wayne是138位UCC失去一个C,使α链的合成不在原来应该终止的地方停止,一直到146位合成精氨酸后才终止,从而使α链延长。 缺失和重复突变 插入突变 (转座子)
脱嘌呤:碱基和脱氧核糖间的糖苷键受到破坏,从而引起一个鸟嘌呤或腺嘌呤从DNA分子上脱落下来。 脱氨基:如胞嘧啶C脱氨基后变成尿嘧啶U。U=A,结果G-C→A-T。
13.5.3 错配修复系统
修复杂种DNA错配碱基及基因转变。
基因突变类型、损伤修复-文档资料

2009年江苏省初赛
48.有人对某一种生物的两个不同个体的同一个基因 的部分序列进行了测定,结果见下表:
根据上表的数据,你认为在第二个个体身上发生的突 变为: A.错义突变 B.缺失突变 (答案:D) C.沉默突变 D.无义突变
2019年江苏省初赛
80.一个访问地球的外星人被发现与我们具有相同的遗传密码。
G C
C G
C G
DNA
G C
C G
A T
DNA
C
GGCG NhomakorabeaU
mRNA
mRNA
精
氨基酸
精
氨基酸
错义突变:三联体密码子发生突变导致蛋白质中原 来的氨基酸被另一种氨基酸取代。 无义突变:三联体密码子突变为终止密码子。它导 致翻译提前结束而使产物失活。
2. 移码突变 是指:由一个或多个非三整倍数的核苷酸插入 或缺失,而使编码区该位点后的三联体密码子阅 读框架改变,导致后向氨基酸都发生错误。 通常该基因产物会完全失活;如出现终止密码 子则使翻译提前结束。 3. 密码子的缺失或插入 是指:决定一个氨基酸的三个核苷酸同时缺失 或增加。这样多肽链中氨基酸数目会减少或增加, 但密码子的读框不变。
2009年全国联赛
39. 下列关于SOS反应的描述,哪项是正确的?
A. 专用于碱基切除修复
B. 可以完全修复DNA的损伤
C. 专用于嘧啶二聚体的修复 D. 是准确性差的修复方式
答案:D
2009年江苏初赛 36.将来自两个不同的着色性干皮病患者的内表皮细 胞进行融合,发现融合细胞对紫外线的反应像正常人 一样。此结果说明: A.两个病人发生突变的基因是相同的 B.两个病人的细胞不能产生互补 C.紫外线列于健康的人是安全的 D.至少有两个不同的基因参与表皮细胞内的DNA核 苷酸切除修复