遗传学 第06章 细菌的遗传分析
遗传学(王亚馥-戴灼华主编)课后习题答案

第2章孟德尔式遗传分析: 习题解1 题解a:(1) 他们第一个孩子为无尝味能力的女儿的概率是1/8;(2) 他们第一个孩子为有尝味能力的孩子的概率是3/4;(3) 他们第一个孩子为有尝味能力儿子的概率是3/8。
b:他们的头两个孩子均为品尝者的概率为9/16。
2 题解:已知半乳糖血症是常染色体隐性遗传。
因为甲的哥哥有半乳糖症,甲的父母必然是致病基因携带者,而甲表现正常,所以甲有2/3的可能为杂合体。
乙的外祖母患有半乳糖血症,乙的母亲必为杂合体,乙有1/2的可能为杂合体,二人结婚,每个孩子都有1/12的可能患病。
3 题解:a:该病是常染色体显性遗传病。
因为该系谱具有常显性遗传病的所有特点:(1)患者的双亲之一是患者;(2)患者同胞中约1/2是患者,男女机会相等;(3)表现连代遗传。
b:设致病基因为A,正常基因a,则该家系各成员的可能基因型如图中所示c:1/24 题解a:系谱中各成员基因型见下图b:1/4X1/3X1/4=1/48c:1/48d:3/45题解:将红色、双子房、矮蔓纯合体(RRDDtt)与黄色、单子房、高蔓纯合体(rrddTT)杂交,在F2中只选黄、双、高植株((rrD-T-))。
而且,在F2中至少要选9株表现黄、双高的植株。
分株收获F3的种子。
次年,分株行播种选择无性状分离的株行。
便是所需黄、双、高的纯合体。
6 题解:正常情况:YY褐色(显性);yy黄色(隐性)。
用含银盐饲料饲养:YY褐色→黄色(发生表型模写)因为表型模写是环境条件的影响,是不遗传的。
将该未知基因型的黄色与正常黄色在不用含银盐饲料饲养的条件下,进行杂交,根据子代表型进行判断。
如果子代全是褐色,说明所测黄色果蝇的基因型是YY。
表现黄色是表型模写的结果。
如果子代全为黄色,说明所测黄色果蝇的基因型是yy。
无表型模写。
7 题解: a:设计一个有效方案。
用基因型分别为aaBBCC、AAbbCC、AABBcc的三个纯合体杂交,培育优良纯合体aabbcc。
细菌的遗传分析 ppt课件

第六节 细菌的遗传分析
微生物作为遗传研究材料的优越性
ppt课件
15
按照细菌出现感受态的方式,可把转 化分为三种类型
自然转化(naturally occuring transformation):细 菌自发地出现感受态,如肺炎链球菌,流感嗜血杆菌, 枯草杆菌等。 人 工 诱 导 的 感 受 态 (artificially induced competence) :如 Ca2+ 诱导的大肠杆菌等发生的转 化。 原生质体转化(protoplast transformation):将DNA 分 子 连 同 PEG 一 同 加 入 原 生 质 体 , 造 成 细 胞 摄 取 DNA 。 还 可 以 用 电 穿 孔 法 (electroporation) 代 替 PEG , 用 高 压 脉 冲 电 流 在 细 胞 膜 上 击 成 小 孔 , 使 DNA 分子通过小孔而导入细胞,又称为电转化。可 适用于多种细菌,放线菌和真核细胞的转化。
结果与结论:
仍然出现原养型菌落。 从而表明互养并非原养型菌落出现的原因,而可能发生 了遗传重组。
ppt课件 26
转化作用及其排除
Lederberg 和 Tatum 曾 把 品系 A 的培养液经加热灭 菌,加入到 B 品系的培养 物中,未得到原养型菌落; 表明原养型菌落可能不是 由转化作用产生。 戴维斯(Dawis, 1950) 的 U 型管试验(结果没有得到原 养型细菌); 实验结论:细胞直接接触 是原养型细菌产生的必要 条件。 ppt课件
第二节细菌的遗传分析

2020/4/16
2
二、接合(conjugation)
• 在原核生物中,两个细胞在相互接触过程中, 遗传物质从一个个体转移到另一个个体的现象 称为接合。 输出遗传物质的个体称为供体(donor), 又称为“雄性”。接受外源遗传物质的个体称 为受体(receptor),又称为雌性。 E.coli(大肠杆菌)是遗传学研究中应用最 为广泛的细菌。野生型的E.coli可以在只含有盐 类和葡萄糖的简单培养基上生长。
2020/4/16
4
黎德伯格和塔特姆接合试验
2020/4/16
5
黎德伯格和塔特姆接合试验
• A和B均不能在基本培养基上生长,但若将 A和B在完全液体培养基上培养几个小时以 后再涂布在基本培养基上,就能长出一些 原养型(met+bio+thr+leu+)的菌落。细菌 的野生型又称为原养型。
• 这种原养型菌落的出现是由于营养上的互 补,还是由于两种不同类型细胞直接接触 而交换了遗传物质的结果呢?
第二节 细菌的遗传分析
细菌与细菌之间的遗传物质的交流 (拟有性过程)有四种不同的方式:
一、转化 二、接合(杂交) 三、性导 四、转导
2020/4/16
1
一、转化(Transformation)
• 细菌通过细胞膜摄取周围环境中DNA片 段,并通过重组将其整合到自身染色体 中的过程,称为转化。
当外源DNA进入宿主后,使宿主产 生新的表现型时就能测知转化的发生。
2020/4/16
10
F 因子的存在状态
2020/4/16
11
(二)F因子
• F因子处于自主状态时,可以不依赖宿主细胞 的染色体而独立复制(每个F+细胞只有一个F 因子)。据研究,F因子至少包含有15个基因, 其中有的基因控制F(或性)伞毛(F pillus) 的形成,F伞毛是F+细胞表面伸出的一种长附 属物。F+与F+之间互不理睬,但F+和F-一旦 相互接触,F伞毛就变成了两个细胞之间原生 质的通道,叫做结合管(conjugation tube)。 F+细胞中的F因子由结合管向F-传递,使F-变 成F+。
细菌的遗传分析试题答案

细菌的遗传分析试题答案一、选择题1. 细菌遗传物质的主要类型是什么?A. DNAB. RNAC. 蛋白质D. 糖类答案:A2. 在细菌中,哪种物质负责携带遗传信息?A. 质粒B. 染色体C. 噬菌体D. 细胞壁答案:B3. 细菌的基因重组通常通过哪种方式发生?A. 转化B. 转导C. 接合D. 所有以上答案:D4. 细菌的突变通常会导致什么结果?A. 抗药性增强B. 代谢速率改变C. 形态结构变化D. 所有以上答案:D5. 细菌的遗传分析中,哪种技术可以用来确定DNA序列?A. PCRB. 凝胶电泳C. 南方杂交D. 北方杂交答案:A二、填空题1. 细菌的染色体通常是________,并且可以在细胞分裂时被复制和传递给子代。
答案:环状双链DNA分子2. 在细菌中,________是一种小型的、环状的DNA分子,可以在细菌间进行水平基因转移。
答案:质粒3. 细菌的基因突变可能是由于________、化学物质或________引起的。
答案:紫外线辐射、自发突变4. 通过________技术,可以将细菌的DNA片段插入到载体中,用于基因克隆和表达。
答案:重组DNA技术5. 细菌的遗传分析中,________是一种用于检测特定DNA序列的技术,通过标记的探针与目标DNA的互补配对来实现。
答案:南方杂交三、简答题1. 简述细菌基因突变的类型及其可能的影响。
答案:细菌基因突变的类型包括点突变、插入突变和缺失突变。
点突变是指单个核苷酸的改变,可能导致氨基酸的改变或不影响蛋白质的功能。
插入突变和缺失突变则涉及一个或多个核苷酸的增加或减少,可能导致移码突变,从而影响蛋白质的结构和功能。
突变可能对细菌的生存和适应性产生重要影响,如抗药性的产生或代谢途径的改变。
2. 描述细菌接合的过程及其在遗传学研究中的意义。
答案:细菌接合是指两个细菌通过直接接触进行遗传物质的交换。
在这个过程中,一个细菌的质粒或染色体片段可以转移到另一个细菌中,从而实现基因的水平转移。
第六章细菌和噬菌体的遗传分析

18
五、细菌的遗传作图
(一)细菌的遗传重组 原核生物的遗传重组实质上是指受体中插入来自供体的遗传性不同 的DNA片段,并把这种DNA片段或它的复本整合为受体基因组的一部分。 受体的遗传重组可以通过三种途径来实现: 1、接合 供体细菌的DNA通过接合管进入受体细菌,实现基因重组。 2、转化 游离的细菌DNA片段被不同的细菌细胞(受体)吸收. 3、转导 一种细菌的DNA片段经过温和的或有缺陷的噬菌体传递给另一种细菌
2
霉菌菌落
3
大肠杆菌
4
二、细菌的突变型
(一)营养缺陷型 生理特性的突变包括:丧失合成某种营养物质能力的营养缺陷型。 (二)抗药突变型 抗性突变包括:抗药性或抗感染性。
5
三、细菌有性杂交
1964年Ledeberg和Tatum用大肠杆菌K,证明细菌有性杂交存在。 (一)大肠杆菌杂交试验
A bio-(生物素) met-(甲硫氨酸) thr+(苏) leu+(亮) thi+(VB1) B bio+(生物素) met+(甲硫氨酸) thr-(苏) leu-(亮) thi-(VB1) A、B都为营养缺陷型,在基本培养基上不能生长。
h+r × hr+
子代噬菌体基因型 h+r hr+ h+r+ hr 含B/B2培养基上噬菌斑 大半透明 小透明 小半透明 大透明 交换值=重组型噬菌体数/总噬菌斑数×100%= h+r++hr/总×100% =小半透明+大透明/总×100% (二)连锁图 T2快速溶菌突变型有多种,如ra、rb、rc都形成大噬菌斑,用不同类 型快速溶菌突变型与宿主范围突变型杂交,结果如下: 杂交组合 h+r hr+ h+r+ hr 重组值 h-r图距 h+ra×hr+ 34.0% 42% 12% 12% 24% 24 h+rb×hr+ 32.0% 56% 5.9% 6.4% 12.3% 12.3 h+rc×hr+ 39.0% 59% 0.7% 0.9% 1.6% 1.6 按图距h、ra、rb、rc有4种排列
(优选)细菌的遗传分析打印

• 若共转化的频率要比两个单个基因转化频率相乘积高的 话,那么这两个基因一定是紧密连锁的。
• 基因的顺序也可以通过共转化的结 果来分析确定,例如p+和q+常常共 转化,而q+和o+也常常共转化,但 基因p+和o+从未发生共转化,那么 这三个基因的顺序一定是p – q - o。某些处理过程可以诱导或加强感受态, 以 生大 长肠 后杆 期菌的为大例肠,杆用菌可Ca以2+(增如强Ca其C感l2)受处能理力对。数
按照细菌出现感受态的方式,可
把转化分为三种类型
1. 自然转化(naturally occuring transformation):细菌 自发地出现感受态,如肺炎链球菌,流感嗜血杆菌, 枯草杆菌等。
2和3:工程转化(engineered transformation)
转化过程
图7-13 细菌转化的机制
(三) 共同转化与遗传图谱绘制
• 通过转化可以测定基因的连锁、基因的排列顺序 以及图距。原理如下:
• 如果在供体的染色体上有两个离得很远的基因a+和b+, 我们将会发现它们总是在不同的DNA片段上,这样如果 有一个a+b+供体和ab受体,那共转化(cotransformation) 即两个基因同时转化的概率是由两个基因单独转化的概 率的乘积。若每个基因的转化频率是10-3的话,那么这 两个基因同时被转化的频率应为10-3×10-3=10-6。
突变型的筛选
例如: 营养缺陷型的筛选: u.v
野生型细菌
CM 完全培养基: MM 基本培养基:
影印培养
影印实验(replica plating )
Joshua Lederberg & Esther Lederberg(1952)
遗传学_ 细菌和病毒的遗传分析_
1180 + 418 + 685 +107 +11940 +3660
100% = 2390 100% =13% 17990
trp2
tyr
34
his2
13 tyr1
his
40
trp
八、转导(transduction)
⚫ 普遍性转导(Generalized transduction)
转导是以噬菌 体为媒介,将 外源基因携带 入细菌,使受 体细胞发生遗 传重组的方式。
a、b间发生交换
单性状的转化子
a、b间不发生交换
双性状的转化子
七、转化作图的原理
细菌两连锁基因的交换率
=
单性状转化子的数 单性状转化子数+共转化的转化子数
100%
表7-1 枯草芽孢杆菌trp2+ his2+ tyr1+(供体)× trp2- his2- tyr1-(受体)的转化实验 座位转化子类型
噬菌体的遗传分析
一、细菌和病毒的遗传分析
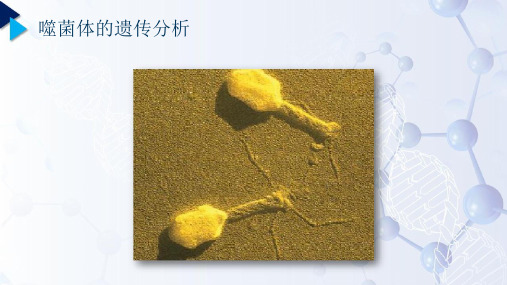
7-1 T4噬菌体的电镜照片
二、病毒对遗传学研究的贡献
1952年 Hershey & Chase的同位素示踪试验
证明T4病毒的遗传物质 是脱氧核糖核酸(DNA) 【1969年诺贝尔奖】
二、病毒对遗传学研究的贡献
1956年Fraemkel Conrat的烟草花叶病毒的重建试验
滑,可致病)
粗糙型R菌株 (无荚膜,菌落粗
糙,不致病)
三、转化现象的发现——Griffth的肺炎双球菌实验
IIR菌株不致病 IIIS菌株致病
灭活的IIIS菌株不致病 灭活的IIIS菌株的某种物 质使IIR菌株发生性状改 变,变成致病的IIIS菌株
细菌的遗传分析

Question
• 我们已知在F+×F-杂交中,几乎所有F-细菌变 为F+, F+×F-→F+;
• 而在Hfr ×F-杂交中,尽管出现高频重组,但F- 细菌很少转变为F+细菌。这个问题使遗传学家感 到迷惑不解。?
中断杂交实验 (Interrupted-mating experiment)
Wollman 和 Jacob进行中断杂交实验:
细菌的遗传分析
概述
• 细菌、放线菌和蓝细菌等均属于原核生物(prokaryotes)。 • 主要特征:没有核膜,其核基因组是由一个裸露的环状
DNA分子构成,称为拟核。细胞内没有以膜为基础的 细胞器,也不进行典型的有丝分裂和减数分裂。 • 细菌是单细胞生物,结构简单,繁殖能力强,分布广, 世代周期短,个体数量多,在正常条件下,完成一个世 代仅20 min, 较容易诱变和筛选各类型突变。 • 细菌不仅是许多病毒的宿主细胞,而且有自身的遗传特 性,又易于培养建立纯系,长期保存,成为遗传学研究 的常用实验材料。
Hfr : thr+ Leu+ azir tonr Lac+ gal+ strs ×
F- :thr- Leu- azis tons Lac- gal- strr
azi:叠氮化钠; ton:噬菌体T1; str:链霉素; Lac:乳糖; gal:半乳糖
结果发现Hfr的未选择性标记基
因进入F-所需时间: • 9分钟时:
细菌的细胞结构:简单 (原核生物) • 基本结构: 细胞壁 (cell wall), 细胞膜 (cell membrane); 拟核 ( nucleoid ),核糖体 (ribosome), 细胞质 (cytoplasm),内含物等;
• 特殊结构: 一定条件下具有的结构 e.g. 荚膜 (capsule) 和鞭毛 (flagella)
细菌的遗传分析-1

(二)、突变型的筛选 二、
选择培养法: 选择培养法:
是根据菌株在基本培养基和选择培养基上的生长表现确 是根据菌株在基本培养基和选择培养基上的生长表现确 基本培养基 定菌株的突变型, 原养型和营养缺陷型或对某一抗生素的 定菌株的突变型,如原养型和营养缺陷型或对某一抗生素的 敏感型和非敏感型(抗性型 ; 敏感型和非敏感型 抗性型); 抗性型
人工诱变
哈工大哈工大-遗传学
第六章 细菌的遗传分析
营养缺陷突变) 影印法(营养缺陷突变)
人工诱变 完全培养基
印迹
基本培养基 +aaA
基本培养基
哈工大哈工大-遗传学 第六章 细菌的遗传分析
基本培养基 +aaB
原核生物遗传物质转移的方式: 原核生物遗传物质转移的方式: 接合( 接合(conjugation) ) 转化( 转化(transformation) ) 转导(transduction) 转导( )
哈工大哈工大-遗传学 第六章 细菌的遗传分析
因子的三种状态: ⑶. E.coli 与F 因子的三种状态: 因子, ①.没有F因子,即F-; 没有 因子 因子, ②.一个自主状态F因子,即F+; 一个自主状态 因子 因子, ③.一个整合到宿主染色体内的F因子,即Hfr。 一个整合到宿主染色体内的 因子 。
哈工大哈工大-遗传学 第六章 细菌的遗传分析
(三)、F 因子与高频重组品系 1. F 因子
供体和受体的性别差异,是由F因子引起的 供体和受体的性别差异,是由 因子引起的 因子 因子:致育因子(性因子),是一种附加体。 ),是一种附加体 ⑴.F 因子:致育因子(性因子),是一种附加体。 携带F因子的菌株称为供体菌或雄性, 表示。 携带 因子的菌株称为供体菌或雄性,用F+表示。 因子的菌株称为供体菌或雄性 未携带F因子的菌株为受体菌或雌性, 表示。 未携带 因子的菌株为受体菌或雌性,用F-表示。 因子的菌株为受体菌或雌性 ⑵.F 因子的组成: 因子的组成: 染色体外遗传物质,环状 染色体外遗传物质,环状DNA; ; 40~60个蛋白质基因; 个蛋白质基因; 个蛋白质基因 2~4个/细胞 雄性内 。 个 细胞 雄性内)。 细胞(雄性内
遗传学复习要点

遗传学复习要点0.细菌的遗传分析F因子将供体细胞的基因导入受体,形成部分二倍体的过程叫性导或F-导。
F 因子整合进细菌染色体→[Hfr] → F’→与F-接合→ 产生部分二倍体。
F’和λd颗粒不同,它加进了细菌的基因,并不减少本身的基因。
F’因子也没有蛋白质外壳包装的问题,所以长度不为包装所限制。
细菌的转化和转导作图:转化:没有噬菌体作介导,由DNA直接转入受体细胞的过程,称为转化。
细菌的转导与作图转导:以病毒作为载体把遗传信息从一个细菌细胞传到另一个细菌细胞。
转导分为一般性和特殊性转导转导病毒产生的频率非常低。
由于噬菌体外壳蛋白决定噬菌体附着细胞表面的能力,因此,这种噬菌体颗粒仍然具有侵染性。
它感染细菌细胞,并将其内含物-细菌的DNA片断注入其中。
进入的DNA片段可以和寄主细胞DNA发生重组,形成遗传结构发生重组的细菌细胞-转导体。
②共转导频率与图距的关系式1966年,T.T Wu (Harvard University)得到了一个共转导频率与从接合实验中得到的图距相连系的数学表达式:(4)局限性(特异性)转导与作图由温和噬菌体进行的转导叫做局限性转导(specialized transduction)。
该噬菌体DNA整合进细菌染色体中时,都占有一个特定的位置,所以只转移细菌染色体的特定部分。
细菌同源重组的特点细菌的转化、接合和转导重组都是同源重组。
细菌中的重组发生在一个完整的环状双螺旋DNA分子与一个单链或双链DNA分子片段之间,而且没有相对应的(相反的)重组子。
重组发生在单链DNA片段和完整的双链DNA之间,且供体单链与受体DNA之间结合形成一段异源双链区,最后结果取决于错配修复。
无重组发生:校正切除的是异源双链区中的属原供体单链的核苷酸。
若无修复校正作用,则该细菌分裂后产生两个细胞,一个是受体的基因型,另一个是重组体的基因型。
高效率标记:有些遗传标记在转化中很少发生校正作用,或校正切除几乎总是在受体DNA上,因此转化频率较高,这类遗传标记称为~。
医学:细菌的遗传分析和基因定位

质粒和转座子
除了染色体,细菌中还可 能含有质粒和转座子等可 移动遗传元件。
基因密度和结构
细菌基因组中的基因密度 较高,且基因结构相对简 单,通常不含内含子。
基因表达调控
转录调控
细菌通过调节转录起始和转录终止来控制基因表 达。
翻译调控
细菌通过调节翻译起始和翻译终止来控制蛋白质 合成。
适应性调控
细菌在应对环境变化时,会迅速调整基因表达以 适应新环境。
医学细菌的遗传分析和基因定位
contents
目录
• 细菌遗传学基础 • 细菌遗传分析技术 • 基因定位技术 • 医学中细菌遗传和基因定位的应用 • 未来展望与挑战
01 细菌遗传学基础
细菌基因组结构
01
02
03
环状染色体
细菌的基因组通常由一个 环状染色体组成,其大小 通常在数百万至数千万碱 基对之间。
因功能研究和基因克隆等。
04 医学中细菌遗传和基因定 位的应用
病原菌的遗传特征分析
病原菌的遗传特征分析有助于了解病 原菌的传播途径、变异规律和致病机 制,为疾病的预防和治疗提供科学依 据。
通过全基因组测序等技术手段,可以 全面揭示病原菌的基因组结构和变异 情况,为快速诊断和有效控制疾病提 供支持。
抗生素抗性的遗传基础
抗生素抗性的遗传基础研究有助于发 现新的抗生素药物靶点,为开发新型 抗生素提供理论支持。
通过研究病原菌对不同抗生素的抗性 机制,可以了解抗性基因的传播方式 和抗性进化规律,为制定有效的抗感 染治疗方案提供依据。
疾病与基因变异的关系研究
疾病与基因变异的关系研究有助于发现新的疾病易感基因和致病基因,为疾病的 预测、预防和治疗提供新思路。
公平获取资源
细菌的遗传分析

(六)大肠杆菌的染色体呈环状
从上表中可以看出,转移顺序的差异是由于各Hfr之间转移的原点(O)和转移的方向不同所致。
该实验说明F因子和细菌DNA都是环状的,F因子插入环状染色体的不同位置形成不同的转移原点和转移方向。
*
(六)大肠杆菌的染色体呈环状
*
三、性导(sexduction) (一)F’因子 整合到细菌中的F因子也可以重新离开染色体,成为独立的环。这个过程是整合的逆过程,称为环出(looping out)。 F因子在环出过程中并不是完全准确无误的,往往连同部分染色体片段一同离开。 部分染色体DNA与F DNA的杂合环称为F’因子。
*
(四)细菌的交换过程
这样,重组后的F-细菌不再是部分二倍体,而是单倍体,得到的重组体的类型只有一个,而不是两个,相反的重组体是不能存活的(例如有++,没有――)。
*
(五)用中断杂交技术作连锁图
Wollman和Jacob用中断杂交实验了解接合过程中基因转移的顺序和时间,从而绘制出连锁图。
根据供体基因进入受体细胞的顺序和时间绘制连锁图的技术,称为中断杂交技术。
*
(一)杂交实验
1946年,Leaderberg和Tatum发现E.coli可以通过接合交换遗传物质。选用两个不同营养缺陷型的E.coli菌株,A和B。A菌株需要在基本培养基中补充甲硫氨酸(met)和生物素(bio) ,B菌株需要在基本营养培养基上补充苏氨酸(thr)和亮氨酸(leu)才能生长。采用多营养缺陷型是为了防止回复突变干扰试验结果。
*
黎德伯格和塔特姆接合试验
*
黎德伯格和塔特姆接合试验
A和B均不能在基本培养基上生长,但若将A和B在完全液体培养基上培养几个小时以后再涂布在基本培养基上,就能长出一些原养型(met+bio+thr+leu+)的菌落。细菌的野生型又称为原养型。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
细菌的重组
Hfr菌株的进入F-菌株的不 完全的基因组(外基因 子exogenote)与F-的 染色体(内基因子 endogenote )之间发 生交换。
结论
A、B菌株有细胞接触,并发生了重组。 不同品系的大肠杆菌可以杂交(接合),并 进行基因重组。
杂交时的贡献
Hayes发现,两个菌株在杂交时贡献不 一样
A(strr)与B(strs) A(strs)与B(strr) 在不含str的基本培养基上都能得到菌落 但在含str的基本培养基上,只有A(strs)与
分解代谢功能的突变型 catabolic functional mutants
抗性突变型 resistant mutants
大肠杆菌基因符号的命名
Demerec 法则
规定: 表型(首字母大写,正体) Gal+ ,Gal-;Tetr,Tets
基因型 (斜体) gal+ gal- ;tetr tets
mal 麦芽糖不能利用 man 甘露糖不能利用 met 甲硫氨酸缺陷型
mtl 甘露糖醇不能利用 pdx 吡多醇缺陷型 phe 苯丙氨酸缺陷型 pro 脯氨酸缺陷型 pur 嘌呤缺陷型 pyr 嘧啶缺陷型 rha 鼠李糖不能利用 str 链霉素抗性 thi 硫胺缺陷型 thr 苏氨酸缺陷型 ton phageT1抗性 trp 色氨酸缺陷型
第六章 细菌的遗传分析
本章重点: 细菌杂交 转化和转导
学时数: 3-4学时
细菌的细胞和基因组
细菌细胞
比较小,有细胞壁 拟核(nucleoid) 无真核细胞特有的细胞器
细胞分裂就是生长繁殖
细菌的分裂
细菌的基因组
细菌染色体
细菌染色体大多为裸露的环状闭合DNA双链,没有 组蛋白和其他蛋白质结合,也不形成核小体结构。
附加体episome:有些质粒除了可在染色体外
自我复制外,还可以整合到细菌染色体上,成 为细菌染色体的一部分,随着细菌染色体的复 制而复制。
F因子的结构
F因子
F+通过性伞毛与F-结合,传递遗传物质。 F-得到F因子变成F+; F+失去F因子变成F-。 F+ F-杂交通常只转移F因子,染色体上基因 转移的频率很小,不到10-6
(这种结构有利于外源DNA的插入。)
E.coli基因组概况
E.coli的染色体为闭合双链环状DNA,长约 1333um.
2001年10月15日完成了E.coli K12菌株的基因组全 序列测定。总共4639221 bp, 4279个蛋白质编码 基因,115个编码rRNA和tRNA的基因。 (GenBank编号:U00096)
- 表示缺陷型或突变型 相同表型的不同基因突变(基因座)则在3字母后加 大写字母,如metA, metB,……
细菌中常用的突变型基因符号
ara 阿拉伯糖不能利用 arg 精氨酸缺陷型 att 原噬菌体附着点 ade 腺嘌呤缺陷型 azi 叠氮化钠抗性 bio 生物素缺陷型 cys 胱氨酸缺陷型 gal 半乳糖不能利用 his 组氨酸缺陷型 lac 乳糖不能利用 leu 亮氨酸缺陷型 lys 赖氨酸缺陷型
tsx phageT6抗性 tyr 酪氨酸缺陷型 xyl 木糖不能利用
大肠杆菌的突变型的筛选
分解功能突变型
Media中加入呈色底物,利用分解产物与底 物的反应或不反应
抗性突变型
Media中加入抗生素或phage
合成功能突变型(营养缺陷型)
用影印培养或点种法
影印培养
点种法
诱变后培养在完全培养基上,缺陷型和野生 型都能生长
野生型从何而来?
回复突变? × 转化? 营养互补? 基因重组?
突变率一般为2×10-7 , 两个基因同时突变4 ×10-14 三个基因同时突变8 ×10-21
U型管试验
滤器孔径极小,保证细胞不能通过, 避免细胞接触。
分别培养A、B菌株,通气保证培 养液物质交换。 分别涂平板, 都没有菌落生长。
高频重组
研究中发现,有一个菌株与F-杂交时, F很少转变成F+,但后代中重组子频率很 高,达到10-2, 这一菌株被称为高频重组菌株(high frequency recombination strain,Hfr菌株)。
Hfr细菌重组的特点
F因子通常最后进入F-菌株, F-菌株得到 的往往是染色体片段,仍然为F-菌株。 部分二倍体(partial diploid)
E.coli基因组注 释图(部分) 彩框表示编码 蛋白质的基因。
大肠杆菌的突变型及筛选
细菌可以在人工培养基上培养,形成的群体称为菌落。 colony
基本培养基和完全培养基;
野生型(原养型)和突பைடு நூலகம்型
Wild-type
mutant
枯草芽孢杆菌的菌落
大肠杆菌的突变型
合成代谢功能的突变型 Anabolic functional mutants
原点(origin)
细菌染色体之外的能独立增殖的环状DNA, 含形成性伞毛的基因。又称性因子(sex factor)或致育因子(fertility factor),可 以转移。约6×104bp,约为E.coli染色体 的2%
质粒、附加体
质粒plasmid:指染色体外的遗传物质,可以
自我复制,并在细胞分裂时分配到子细胞中。
B(strr)能得到菌落,说明遗传物质是单向转 移的过程
大肠杆菌的性别
在杂交中提供遗传物质的A菌株相当于雄 性,记作F+,因为在细胞中有F因子
接受遗传物质的B菌株相当于雌性,记作 F-,在细胞中没有F因子
致育基因
F因子 (fertility gene)
F factor/element
配对区域(pairing region)
点种于含单一营养物质的基本培养基上
如果在所有平板上都 能生长,是野生型; 只在其中一个平板上 生长的,就是所加营 养物质的缺陷型
大肠杆菌的杂交
接合(conjugation)
细菌的杂交 A
B 完全液体培养基
基本固体培养基平皿
1946,Lederberg和Tatum用 A菌株 met- bio- thr+ leu+ thi+ B菌株 met+bio+ thr- leu- thi做混合培养,涂基本培养基平板,长出了几 十个菌落,发生频率为10-7 表明met+ bio+ thr+ leu+ thi+组合到同一细胞中