谷氨酸受体
离子型谷氨酸受体途径-概述说明以及解释

离子型谷氨酸受体途径-概述说明以及解释1.引言1.1 概述概述离子型谷氨酸受体途径是神经递质谷氨酸通过作用在神经元膜上的离子通道蛋白而引发的一种信号传导机制。
离子型谷氨酸受体在神经元的兴奋性调节、突触传递以及神经元间的相互作用中发挥着重要作用。
本文将首先介绍离子型谷氨酸受体的定义与特点,然后探讨其在神经传导中的作用,最后分析离子型谷氨酸受体途径在疾病治疗中的潜在应用。
通过对离子型谷氨酸受体途径的系统阐述,有助于深入了解神经递质传导的机制,为相关疾病治疗提供新的思路和方法。
1.2 文章结构本文将首先介绍离子型谷氨酸受体的定义与特点,对其在神经传导中的作用进行详细探讨。
接着,将分析离子型谷氨酸受体途径在疾病治疗中的应用,并对其在这一领域的发展进行展望。
最后,总结离子型谷氨酸受体途径的重要性,并对其未来发展提出建议,以期为相关研究和临床应用提供参考。
}}}请编写文章1.2 文章结构部分的内容1.3 目的:本文旨在全面介绍离子型谷氨酸受体途径在神经系统中的重要作用,并探讨其在疾病治疗中的潜在应用。
通过深入研究离子型谷氨酸受体的定义、特点以及神经传导中的作用机制,我们将更好地了解神经递质的传导过程和神经元之间的相互作用。
同时,探讨离子型谷氨酸受体途径在治疗神经系统疾病如癫痫、帕金森病等方面的应用,将有助于拓展新的治疗途径和药物设计思路。
通过本文的阐述,希望读者能够深入了解离子型谷氨酸受体途径的重要性,并为未来疾病治疗的研究和发展提供参考和启示。
2.正文2.1 离子型谷氨酸受体的定义与特点:离子型谷氨酸受体是一类参与神经递质谷氨酸在神经元间传递的重要蛋白质通道,在神经系统中扮演着至关重要的角色。
这些受体通常由四个亚基组成,每个亚基都具有离子通道的特性,可以对荷带正电荷的离子(如钠、钾、钙等)进行选择性通透。
离子型谷氨酸受体主要分为NMDA受体、AMPA受体和kainate受体三类。
NMDA受体是最为研究广泛的一种离子型谷氨酸受体,其特点在于对于钙离子的通透性较高,参与了突触的可塑性、学习记忆等功能。
谷氨酸受体神经生物学
(二)AMPA&KA受体
AMPA受体激动可引起Na+内流,主要参与正常 的突触传递,形成突触后反应即EPSP.
(一)NMDA受体: 1.一般特征
(1)通道耦联的离子型受体、Ca++通道。 (2)通道被Mg++以电压依赖性方式阻断,该通道
受电压、化学因素双重控制。 (3)缓慢通道动力学特征。
2.受体-通道特征 (1) Zn2+(一) (2) H+(一)PH6.0完全阻断 (3)甘氨酸:Co-agonist (4)多胺:甘氨酸依赖性
图示海马的结构及产生LTP的示意图
海马分为三个区,由颗粒细胞构成的齿回(DG), 由锥体细胞构成的CA3区和CA1区,以及三条通路, 由嗅皮层发出到齿回的前穿质纤维(perforant path, pp),由齿回发出到CA3区的苔状纤维 (mossy fiber, mf)及由CA3区发出到CA1去的 Schaffer纤维/侧支.它们依次形成突触联系,而且 都是谷氨酸能突触.
AP4引起的突触前抑制,主要改变EPSP的频率 而不是幅度.
(2)突触前作用机制
突触前抑制主要通过激活PTX敏感的G-蛋 白实现其作用.
具体机制可能涉及不同的途径,如抑制突 触前Ca++电导, 激活K+通道而增加K+电流. 增加K+电流可以降低突触前膜的去极化,从 而减少Ca++内流量.另外,增加K+电流可以增 加突触前膜产生动作电位的阈值和降低动 作电位的峰值,从而减少递质的释放.
谷氨酸及NMDA受体与全身麻醉药

谷氨酸及NMDA受体与全身麻醉药谷氨酸是中枢神经系统中最主要、最广泛分布的神经递质之一。
在全身麻醉药的使用过程中,谷氨酸及其受体NMDA受体的作用机制相当复杂,对于全身麻醉药的临床效应有着重要的影响。
谷氨酸与NMDA受体NMDA(N-甲基-D-天门冬氨酸)受体是一种离子通道,广泛分布于中枢神经系统中。
NMDA受体包含四个亚基,可以通过离子通道的开放而发挥功能。
这种受体对于神经递质谷氨酸有着极为特别的作用——它仅在谷氨酸的存在下才能被激活。
因此,NMDA受体是谷氨酸在中枢神经系统中的一个重要途径。
在健康的神经系统中,NMDA受体的活性被认为具有很大的意义,其中包括了神经传递、神经发育和记忆等重要的生理功能。
但若是NMDA受体活性过高,不仅会损伤神经细胞,并可能引起中枢神经遗漏(即神经元在传递信号过程中阻断)和神经变性等严重后果。
麻醉药物中一些分子的作用机制就是针对NMDA受体进行抑制。
全身麻醉药对谷氨酸和NMDA受体的影响全身麻醉药对谷氨酸和NMDA受体均有一定的影响。
这些影响的具体效应包括:1. 抑制NMDA受体全身麻醉药不仅仅能够直接抑制NMDA受体的活性,还能够通过其他的途径间接影响谷氨酸的代谢和释放过程,从而抑制NMDA受体的活性。
研究表明,全身麻醉药一般对于大脑的NMDA受体做出的反应是不同的。
例如,丙泊酚等完全性麻醉药性质更类似于NMDA受体阻滞剂;而比如异丙酚则更类似于同时抑制突触前膜、突触后膜的电位差和衔接半透明物质的其他麻醉药。
2. 抑制谷氨酸的释放全身麻醉药中的一部分分子还可以抑制谷氨酸的释放,进而抑制NMDA受体的活性。
研究发现,异氟醚、恩氟烷和丙泊酚等麻醉药都可以通过抑制中枢突触前膜谷氨酸的释放来抑制NMDA受体。
3. 对NMDA受体的结构适应性但不同种类的全身麻醉药对于NMDA受体的影响是不同的,离子性麻醉药、挥发性麻醉药对应的是NMDA受体的不同区域,引起了受体结构改变。
谷氨酸受体

谷氨酸受体种类谷氨酸受体分为两类:一类为离子型受体,包括:N-甲基-D-天冬氨酸受体(NMDAR)、海人藻酸受体(KAR)和α-氨基-3 羟基-5 甲基-4 异恶唑受体(AMPAR),它们与离子通道偶联,形成受体通道复合物,介导快信号传递;另一类属于代谢型受体(mGluRs),它与膜内G-蛋白偶联,这些受体被激活后通过G-蛋白效应酶、脑内第二信使等组成的信号转导系统起作用,产生较缓慢的生理反应。
离子型受体(1) NMDA 受体(NRs):其与突触的可塑性和学习记忆密切相关。
通过该受体本身、其共轭的离子通道及调节部位3 者形成的复合体而发挥功能,对Ca2+高度通透。
每个NMDA 受体上含有两个谷氨酸和两个甘氨酸结合识别位点,谷氨酸和甘氨酸均是受体的特异性激活剂。
到目前为止已克隆出5个亚基,NMDAR1、NMDAR2(A-D)其中NMDAR1 可单独形成功能性纯寡聚体NMDAR,但NMDAR2 亚基却不具备该功能。
有研究表明NMDAR可能是由NMDAR1 和NMDAR2 不同的亚基组成的一个异寡聚体。
(2) KA/AMPA 受体:它们也是受配基调控的离子通道,对Na+、K+有通透性,研究证明,一些受体亚型对Ca2+也有通透性。
AMPA 家族包括4 个结构极为相似的亚基GLUR1-4,各亚基的氨基酸序列的同源性高达70%。
由于氨基酸残基的疏水性分布,在靠近羧基端的部分构成4 个跨膜区。
AMPA、L-谷氨酸及KA 均可激活这类离子通道,并有AMPA 的高亲和力结合位点。
天然的AMPAR 是由这4 种亚基形成的五聚体。
每个单位的分子量为108kd。
AMPA 受体的4 种亚基在第4 个跨膜区上游均含有1 个由38 个氨基酸残基组成的特殊区段,该区存在2 个结构相似区,分别由受体基因上的2 个相临的外显子编码。
但各亚基的DNA 编码在翻译后要经过一些如:磷酸化、糖基化等修饰,这些修饰是通道功能的重要调节方式。
谷氨酸激活受体的机制

谷氨酸激活受体的机制1 引言谷氨酸激活受体(GluN)是N-甲基-D-天门冬氨酸(NMDA)受体中的一种亚型,具有重要的生理学和病理学作用。
该受体对学习、记忆、神经发育和神经退行性疾病等方面发挥着至关重要的作用。
近年来,对GluN功能和调节机制的研究引起了广泛关注。
2 谷氨酸激活受体的结构和功能GluN是一种离子通道受体,由四个亚基组成。
其中两个亚基是GluN1,另外两个亚基是GluN2、GluN3。
GluN1亚基是必须的,因为它向离子通道提供了游离位点。
GluN2和GluN3亚基则使得GluN亚型具有高度的选择性、调节性和发挥多种功能的能力。
GluN作为一种离子通道受体,对锌离子和镁离子有高亲和力。
其和镁离子之间的作用使GluN的活性受到抑制,这种抑制可以被钙离子和谷氨酸的结合解除。
GluN的活性被谷氨酸的结合所激活,使得钙离子可以进入细胞,从而触发细胞内的一系列生物学响应。
3 谷氨酸激活受体的调节机制GluN的活性可以通过多种方式进行调节。
最常见的是GluN亚型的组成方式,不同的亚型具有不同的功能和调控特性。
此外,GluN的活性还可以受到蛋白激酶、磷酸酶等蛋白质翻译后修饰的影响。
还有一些内源性物质,如阿尔茨海默氏病相关蛋白、五磷酸腺苷、氮一氧化物等,也可以对GluN的活性产生影响。
此外,GluN的活性还可以受到环境中的化合物、药物的影响。
例如,可卡因就是一种通过抑制GluN的活性,从而影响神经传递的典型药物。
4 结论总的来说,GluN是一个高度复杂的蛋白质。
它的活性被多种因素所调节,这些因素包括GluN亚型的组成方式、内源性调节因子、外源性化合物等。
对于GluN的研究已经深入,但人们对其调控机制的了解还有待加强。
通过深入研究GluN的功能和调控机制,有望为相关疾病的治疗提供有效的策略。
谷氨酸大脑中的重要兴奋性神经递质

谷氨酸大脑中的重要兴奋性神经递质谷氨酸(Glutamate)是一种重要的神经递质,在大脑中发挥着关键的兴奋作用。
它在神经传递过程中发挥着重要的作用,对维持大脑功能起着至关重要的作用。
本文将详细探讨谷氨酸在大脑中的功能及其重要性。
1. 谷氨酸的生物学角色谷氨酸是一种氨基酸,它是大脑中最主要的兴奋性神经递质之一。
它主要通过突触传递信息,并参与了大脑的学习、记忆和认知过程。
谷氨酸的产生与代谢是一个精密的调控系统,它在神经活动过程中保持了神经细胞之间的信息传递。
2. 谷氨酸的受体在大脑中,谷氨酸通过与其受体结合来产生兴奋性作用。
主要的谷氨酸受体包括NMDA受体、AMPA受体和kainate受体。
这些受体的活化能够引起离子通道的打开,从而电势改变和信号传递的增强,进一步加强神经元之间的通讯。
这些受体的功能在大脑的不同区域和神经元类型中有所不同,从而使谷氨酸的作用在不同的神经网络中得到精确调控。
3. 谷氨酸在学习和记忆中的作用谷氨酸在学习和记忆中发挥着重要的作用。
研究表明,谷氨酸通过与学习和记忆相关的脑区产生作用,增强神经元之间的突触连接,并参与了突触可塑性的调节。
此外,谷氨酸还影响了突触前后的各种信号转导途径,调节了胞内信号转导的平衡,从而参与了学习和记忆的正常进行。
4. 谷氨酸与神经系统疾病的关系谷氨酸在一些神经系统疾病中起到了重要的作用。
过高或过低的谷氨酸浓度与一些神经系统疾病的发生和发展密切相关。
例如,谷氨酸的过度释放与神经元损伤和细胞死亡有关,这在中风和帕金森病等神经系统疾病中发挥了重要作用。
5. 谷氨酸的调控机制为了维持大脑正常的功能,谷氨酸的水平需要得到精密的调控。
大脑中有多种机制来调节谷氨酸的释放和降解。
这包括通过转运蛋白和酶的调节来确保谷氨酸在突触间隙中的适当浓度,以及对谷氨酸受体的调制来确保谷氨酸的信号传递在适当的范围内。
综上所述,谷氨酸作为一种重要的兴奋性神经递质,在大脑中发挥着重要的作用。
谷氨酸受体的功能和结构

谷氨酸受体的功能和结构谷氨酸受体是一种神经元受体,它在神经信号传递中发挥着重要的作用。
在人脑中,谷氨酸受体是最常见的受体之一,也是神经递质谷氨酸的主要递质受体。
谷氨酸受体的结构和功能已经被广泛研究,研究表明它在许多神经系统疾病的治疗上具有非常重要的潜力。
本文将介绍谷氨酸受体的功能和结构。
一、谷氨酸受体的功能谷氨酸是人体中最常见的神经递质之一,它是神经元之间信号传递的关键。
当神经元释放谷氨酸到突触间隙时,相关受体会被激活,此时信号转移到下一神经元,或者发挥其他作用。
谷氨酸受体是与谷氨酸作用的受体之一,它们的共同作用是促进神经信号的传递。
在神经系统中,谷氨酸受体经常被称为离子通道受体,因为它可以形成一个离子通道,领先于钙离子或钠离子等正离子进入胞内,从而产生神经信号传递的结果。
由于谷氨酸受体的这种离子通道特性,它可以调节神经元之间的连接强度,促进或抑制信号传递,从而影响大脑的感觉、运动、认知、情绪等方面的功能。
二、谷氨酸受体的结构谷氨酸受体分为两种类型:离子通道受体和嵌合型受体。
1、离子通道受体离子通道受体是图2所示的一种最简单的谷氨酸受体。
此类受体有一个离子通道,当谷氨酸结合受体时会打开通道,导致离子进入胞内。
离子通道受体通常在短时间内发挥作用,因为它们的响应速度非常快。
2、嵌合型受体与离子通道受体不同的是,嵌合型受体没有离子通道。
当谷氨酸结合嵌合型受体时,它会与受体特定的细胞内信号通路发生作用,从而产生特定的细胞反应。
嵌合型受体响应速度较慢,因此它们的作用会持续一段时间。
三、总结综上所述,谷氨酸受体是最常见的神经元受体之一,在神经信号传递中发挥着重要的作用。
它在神经系统中形成离子通道或与细胞内信号通路发生作用,从而产生不同的细胞响应。
谷氨酸受体的结构和功能已被广泛研究,研究表明它可以在许多神经系统疾病的治疗上发挥着非常重要的潜力。
在未来,人们希望能够深入了解谷氨酸受体的分子机制,以更好地理解它在神经元中的作用,从而设计更有效的治疗方法。
谷氨酸能神经元的亚型

谷氨酸能神经元的亚型谷氨酸是一种神经递质分子,它在神经系统中起着重要的作用。
在神经元中,谷氨酸主要通过激活不同的亚型受体来传递神经信号。
这些谷氨酸受体亚型在结构和功能上有所不同,下面将介绍几种常见的谷氨酸受体亚型及其作用。
1. NMDA受体NMDA受体是一种离子通道受体,具有高度选择性的对谷氨酸和N-甲基-D-天冬酰胺(NMDA)敏感。
该受体主要存在于中枢神经系统的突触间隙中,参与学习和记忆等认知功能的调节。
NMDA受体的激活需要同时存在谷氨酸、甘氨酸和膜电位的去极化,它的开放能够引起钙离子的内流,从而介导长时程突触可塑性的形成和维持。
2. AMPA受体AMPA受体是一种离子通道受体,对谷氨酸敏感。
在神经传递过程中,AMPA受体主要负责传递快速的兴奋性信号。
当AMPA受体激活时,钠离子会快速进入细胞内,引起神经元的去极化和兴奋。
AMPA受体在神经系统中广泛分布,参与了许多生理和病理过程,如学习记忆、神经发育和神经退行性疾病等。
3. kainate受体kainate受体也是一种离子通道受体,对谷氨酸和kainate敏感。
kainate受体与AMPA受体具有相似的离子通道结构,但其功能和调节机制有所不同。
kainate受体在中枢神经系统中广泛分布,参与了神经元的兴奋性传递和突触可塑性的调节。
研究发现,kainate 受体与癫痫、脑缺血和神经退行性疾病等疾病的发生密切相关。
4. metabotropic谷氨酸受体metabotropic谷氨酸受体是一类G蛋白偶联受体,与上述离子通道受体不同,它们通过激活细胞内信号转导途径来传递信号。
metabotropic谷氨酸受体分为三个亚型:mGluR1、mGluR2和mGluR3。
mGluR1主要参与兴奋性传递和突触可塑性的调节,mGluR2和mGluR3则主要参与抑制性传递和神经保护。
metabotropic谷氨酸受体在神经系统中广泛分布,参与了许多生理和病理过程,如情绪调节、药物成瘾和神经退行性疾病等。
谷氨酸受体及其与中枢神经系统功能相关性研究

谷氨酸受体及其与中枢神经系统功能相关性研究谷氨酸受体是一类重要的神经递质受体,在中枢神经系统的运转中发挥着重要的作用。
它是一种离子通道和代谢型受体,具有广泛的分布和多功能性。
本文将探讨谷氨酸受体的结构、功能以及其与中枢神经系统功能的相关性。
一、谷氨酸受体的结构谷氨酸受体是由多个不同的亚单位组成的,包括GluN1、GluN2、GluN3、GluA1、GluA2、GluA3、GluA4、GluK1、GluK2、GluK3、GluK4、和GluK5。
在细胞膜上的谷氨酸受体由多个亚单位聚集到一起形成一个几乎完全嵌入膜中的结构。
这些亚单位包括外源性配体结合亚单位(即GluA和GluK)和内源性配体结合亚单位(即GluN)。
这些亚单位的组合和作用不仅决定了谷氨酸受体的通透性和灵敏性,还决定了其功能的独特性质。
二、谷氨酸受体的功能谷氨酸受体主要有两种功能,即离子通道和代谢型受体。
离子通道谷氨酸受体可以感受到外界的刺激而导致离子通道的开放和通透性增加,这些离子包括钙、镁、钠和氢等。
代谢型谷氨酸受体则不具有离子通道的功能,但它们能够通过激活细胞内的第二信使来介导细胞间信号的传递。
三、谷氨酸受体与中枢神经系统相关功能的研究谷氨酸受体与中枢神经系统相关的功能有很多,其中与神经发育不成熟所存在的高易损期相关的谷氨酸毒性具有广泛的关联。
由于谷氨酸过度释放导致的中毒已被证明会对中枢神经系统的发育产生极为不利的影响,在许多疾病中均会引起神经元的凋亡。
此外,谷氨酸受体还与许多其他与中枢神经系统功能相关的疾病有着密切的关系,例如阿尔茨海默病、帕金森氏病和抑郁症等。
四、谷氨酸受体的药物开发由于谷氨酸受体具有广泛的分布和多功能性,在许多疾病的治疗中都被认为是一个有前途的靶点。
目前已有一些药物开发出来,它们能够靶向不同类型的谷氨酸受体,从而减轻或治疗相关的疾病。
例如,在帕金森氏病的治疗中,一种被称为“棕色素减排素”(Nurr1)的蛋白质,被证明可以调节谷氨酸受体的活性,从而减少神经元的死亡。
细胞膜内谷氨酸受体的生物学功能及药物研制

细胞膜内谷氨酸受体的生物学功能及药物研制细胞膜内谷氨酸受体(intracellular glutamate receptor,iGluR)是一类位于与外界相接触的细胞膜内的离子通道,能够响应谷氨酸、激活细胞内钙离子通路,在生物体内发挥着重要的生物学功能。
细胞膜内谷氨酸受体分为三种亚型:N-甲基-D-天冬氨酸受体(NMDAR)、α-氨基-3-羧基-5-甲基异恶唑酸酯受体(AMPAR)和半NMDAR。
其中,NMDAR主要分布于神经组织和部分非神经组织中,具有相对较强的离子通导性和能够响应多种递质的特点;AMPAR和半NMDAR分别是NMDAR的主要辅助受体,能够调节神经元兴奋性和突触可塑性。
细胞膜内谷氨酸受体在生理和病理过程中发挥着重要的作用。
在中枢神经系统中,NMDAR和AMPAR是神经元的主要兴奋性受体,参与了学习记忆、痛觉传递、运动协调等功能的调节。
而在某些疾病中,细胞膜内谷氨酸受体也可能起到负面作用。
比如,在神经退行性疾病中,NMDAR亢活和氧化损伤造成的受体过度激活可能导致神经元坏死;在某些神经精神疾病中,神经元兴奋性受体的异常调节也被认为是其发病的重要机制之一。
针对细胞膜内谷氨酸受体在生物体内发挥的生物学功能和存在的相关病理生理作用,多种iGluR药物研制正在进行中。
目前,主要的iGluR药物可分为两类:一类是竞争性拮抗剂,能够与细胞膜内谷氨酸受体结合并保持通道关闭状态;另一类是非竞争性拮抗剂,能够与受体其他部位结合,从而影响其开放状态。
竞争性拮抗剂最早由AMPA、NMDA等天然毒素提取而来,它们能与细胞膜内谷氨酸受体结合,阻断钙离子的通道,从而减轻其过度激活引起的神经元损伤。
随着分子生物学技术的发展,人们开始研制更为特异、高效的竞争性拮抗剂,如现在广泛使用的5-羟基-2-甲基异恶唑酸(AMPA)受体选择性拮抗剂。
这些药物在控制神经系统疾病方面已取得初步的成功,但同时也带来了一些安全方面的难题,如高剂量使用会导致记忆障碍等副作用。
谷氨酸受体拮抗剂功能原理

谷氨酸受体拮抗剂功能原理英文回答:Glutamate receptor antagonists are a class of drugs that work by blocking the action of glutamate receptors in the brain. Glutamate is the primary excitatory neurotransmitter in the central nervous system and plays a crucial role in various physiological processes, including learning, memory, and synaptic plasticity. However, excessive glutamate activity can lead to neurotoxicity and is associated with various neurological disorders, such as epilepsy, Alzheimer's disease, and stroke.By antagonizing glutamate receptors, these drugs inhibit the binding of glutamate to its receptors, thereby reducing the excitatory effects of glutamate on neuronal activity. This helps to restore the balance between excitatory and inhibitory neurotransmission in the brain. There are several types of glutamate receptors, including NMDA receptors, AMPA receptors, and kainate receptors.Glutamate receptor antagonists can target specific subtypes of these receptors or act on multiple receptor subtypes.One example of a glutamate receptor antagonist is memantine, which is commonly used in the treatment of Alzheimer's disease. Memantine selectively blocks NMDA receptors, which are involved in learning and memory processes. By inhibiting the overactivation of NMDA receptors, memantine helps to improve cognitive function and reduce the symptoms of Alzheimer's disease.Another example is ketamine, which is known for its use as an anesthetic. Ketamine is a non-competitive antagonist of NMDA receptors and has been found to have rapid antidepressant effects in individuals with treatment-resistant depression. The exact mechanism of ketamine's antidepressant action is still under investigation, but it is believed to involve the modulation of glutamatesignaling and synaptic plasticity.Glutamate receptor antagonists can also have potential side effects. For example, blocking glutamate receptors canimpair normal synaptic transmission and affect cognitive function. Additionally, excessive inhibition of glutamate receptors can lead to sedation, dizziness, and other central nervous system side effects. Therefore, the use of glutamate receptor antagonists requires careful monitoring and dosage adjustment to minimize these adverse effects.Overall, glutamate receptor antagonists play a crucial role in the treatment of various neurological disorders by modulating glutamate signaling in the brain. Their ability to restore the balance between excitatory and inhibitory neurotransmission offers therapeutic potential for conditions such as Alzheimer's disease, epilepsy, and depression.中文回答:谷氨酸受体拮抗剂是一类通过阻断大脑中的谷氨酸受体来发挥作用的药物。
谷氨酸受体的结构与功能研究

谷氨酸受体的结构与功能研究谷氨酸受体是一类广泛存在于中、外周神经系统中的受体,它们的结构和功能研究至关重要。
在过去的几十年里,越来越多的科学家和研究者致力于谷氨酸受体的研究,以发现这些受体的生物学意义和治疗上的潜力。
1. 谷氨酸谷氨酸是大脑中最普遍的兴奋性神经递质。
它产生于神经元的突触前端,并与谷氨酸受体结合引发神经传递。
然而,由于谷氨酸过量的释放及其与谷氨酸受体结合所造成的神经兴奋效应,在许多神经性疾病中会发挥负面作用。
这也促使了谷氨酸受体的研究。
2. 谷氨酸受体的分类谷氨酸受体分为两类,即离子型谷氨酸受体和代谢型谷氨酸受体。
离子型谷氨酸受体由离子通道构成,分为AMPA、NMDA和Kainate受体;而代谢型谷氨酸受体是由G蛋白耦联受体和酶联受体组成的,主要有mGluR受体。
3. 谷氨酸受体的结构AMPA和Kainate受体是单体,NMDA受体是异二聚体。
离子型谷氨酸受体的构象在不同的状态下也有所变化,如AMPA受体可分为关闭状态、部分开启状态和全开启状态。
MGlur受体则是七跨膜结构,分为8个亚型,常见亚型包括mGluR1、mGluR2、mGluR3 、mGluR5等。
4. 谷氨酸受体的功能离子型谷氨酸受体的激活将导致细胞内钙离子浓度增加,从而调节神经元的兴奋和抑制。
NMDA受体具有重要的突触可塑性效应,例如长时程增强(LTP)和长时程抑制(LTD)等。
于是,针对离子型、代谢型谷氨酸受体的调节也成为治疗多种神经性疾病的方法。
5. 谷氨酸受体在神经性疾病中的作用谷氨酸受体与多种神经性疾病的发生和发展有关。
例如,研究表明,失衡的AMPA受体激活与癫痫发作有关;NMDA受体功能障碍可导致神经退行性疾病如帕金森病、阿尔茨海默病等。
抑制离子型谷氨酸受体和调节代谢型谷氨酸受体可以成为多种神经性疾病的治疗策略。
6. 谷氨酸受体在新药研发中的应用当今,随着对神经性疾病的认识的不断深入,谷氨酸受体也成为了研发新型神经科学药物的重要靶点。
谷氨酸受体及其药物研究

谷氨酸受体及其药物研究谷氨酸受体(Glutamate receptor)是神经系统中最常见的神经元通路转导分子之一,它参与了神经元信号的传递和神经刺激的响应。
谷氨酸受体有两种类型:离子通道型谷氨酸受体和代谢性谷氨酸受体。
离子通道型谷氨酸受体是神经元的电气活动和突触递质释放过程的重要调节因子,是神经元传递信息的主干路线之一。
目前已有许多研究针对谷氨酸受体的药物研究,今天我将介绍一些最近谷氨酸受体及其药物研究的进展。
一、谷氨酸受体介绍1、离子通道型谷氨酸受体介绍离子通道型谷氨酸受体(iGluRs)是神经元膜上的离子通道蛋白,它们由4个亚基组成,包括了NMDA型谷氨酸受体、α-氨基-3-羧基-5-甲基异恶唑酸受体(AMPA-R)和卡巴拉钦受体(Kainate-R)三个基本家族。
其中NMDA型谷氨酸受体是一种高度特异性的钙离子敏感离子通道,在神经元活动中发挥了关键的调节作用,其功能异常与多种神经系统疾病密切相关。
AMPA-R 和Kainate-R则对神经元电气和化学信号传导调节也非常重要。
2、代谢性谷氨酸受体介绍代谢性谷氨酸受体(mGluR)是除了离子通道型谷氨酸受体之外的别一种主要类型的谷氨酸受体,它们主要通过细胞内信号转导的方式参与神经活动和神经元发育等生命过程。
由于他们的影响可能与兴奋性相对保持平衡,他们成为临床意义相对比较重要的靶点。
二、药物开发进展在谷氨酸受体上,有不少的药物研发正在进行。
这些药物的机制多样,在调节谷氨酸受体的功能上发挥了特定的作用,以下是几个药物的主要机制及其应用。
1、甘氨酸受体(mGluR3)激动剂甘氨酸受体(mGluR3)的激动剂在某些神经系统疾病的治疗中得到了广泛关注。
一项发表于美国神经学杂志上的研究显示,一种名叫NGP1-01的新型甘氨酸受体激动剂能显著地改善药物未能控制的精神分裂症患者的不良症状。
这个研究为研发新型甘氨酸受体激动剂打开了一扇门。
2、AMPA型谷氨酸受体正向调节剂AMPA型谷氨酸受体正向调节剂被认为是一种当今最有希望的抗抑郁药物,它能够增强神经元之间的联系,提高神经元之间的通信效率和强度,从而改善抑郁症患者。
谷氨酸受体

谷氨酸受体分为两类:一类为离子型受体,包括:N-甲基-D-天冬氨酸受体(NMDAR)、海人藻酸受体(KAR)和α-氨基-3 羟基-5 甲基-4 异恶唑受体(AMPAR),它们与离子通道偶联,形成受体通道复合物,介导快信号传递;另一类属于代谢型受体(mGluRs),它与膜内G-蛋白偶联,这些受体被激活后通过G-蛋白效应酶、脑内第二信使等组成的信号转导系统起作用,产生较缓慢的生理反应。
离子型受体(1) NMDA 受体(NRs):其与突触的可塑性和学习记忆密切相关。
通过该受体本身、其共轭的离子通道及调节部位3 者形成的复合体而发挥功能,对Ca2+高度通透。
每个NMDA 受体上含有两个谷氨酸和两个甘氨酸结合识别位点,谷氨酸和甘氨酸均是受体的特异性激活剂。
到目前为止已克隆出5个亚基,NMDAR1、NMDAR2(A-D)其中NMDAR1 可单独形成功能性纯寡聚体NMDAR,但NMDAR2 亚基却不具备该功能。
有研究表明NMDAR可能是由NMDAR1 和NMDAR2 不同的亚基组成的一个异寡聚体。
(2)KA/AMPA 受体:它们也是受配基调控的离子通道,对Na+、K+有通透性,研究证明,一些受体亚型对Ca2+也有通透性。
AMPA 家族包括4 个结构极为相似的亚基GLUR1-4,各亚基的氨基酸序列的同源性高达70%。
由于氨基酸残基的疏水性分布,在靠近羧基端的部分构成4 个跨膜区。
AMPA、L-谷氨酸及KA 均可激活这类离子通道,并有AMPA 的高亲和力结合位点。
天然的AMPAR 是由这4 种亚基形成的四聚体。
每个单位的分子量为108kd。
AMPA 受体的4 种亚基在第4 个跨膜区上游均含有1 个由38 个氨基酸残基组成的特殊区段,该区存在2 个结构相似区,分别由受体基因上的2 个相临的外显子编码。
但各亚基的DNA 编码在翻译后要经过一些如:磷酸化、糖基化等修饰,这些修饰是通道功能的重要调节方式。
离子型谷氨酸受体功能的多样性是通过不同亚基组装、选择性基因结合和转录前mRNA 的编辑等方式来实现的。
谷氨酸受体G蛋白

谷氨酸受体:G 蛋白、致癫痫作用、麻醉药及其相互作用一、摘要谷氨酸受体可分为两种类型:离子型谷氨酸受体和G蛋白偶联受体 (GPRC) 或亲代谢型受体 (mGluRs). 我们的研究证实,激活大脑皮质神经元 mGluRs 1受体引起的兴奋作用可以导致癫痫。
近年来研究也揭示麻醉药具有可以调节G蛋白受体的功能。
例如:局麻药利多卡因可以抑制mGluR 兴奋介导的致癫痫作用。
其它研究表明麻醉药抑制GPRC 调节的呼吸道平滑肌收缩作用(Sakihara 等, 2004) 。
麻醉药与G蛋白受体间的相互作用机制尚未完全阐明。
初步研究发现,麻醉药抑制Gα或Gβγ亚单位激活的信号转导蛋白如磷脂酶C(Pabelick 等, 2001)、蛋白激酶C Kamatchi et al., 2001)及离子通道(Yamkage 1992)。
一些证据也表明对GPRC复合体直接的麻醉作用(Nietgen 等, 1998)涉及了在Gα亚单位核苷酸结合位点上对核苷酸交换的直接抑制作用 (Pentyala 等, 1999, Striff 等 2003)。
麻醉药也具有结合细胞膜上GPRC的作用,提示其对异源三聚体G蛋白偶联受体具有抑制作用,而与G蛋白内在活性无关(Ishizawa et al., 2000)。
本文阐述兴奋性GPRC特别是mGluRs引起癫痫的信号机制。
这些可为讨论麻醉药的作用提供背景资料。
二、与癫痫有关的两种主要脑电波癫痫神经冲动放电是大脑皮层神经元的大量同步发放(McCormick & Contreras, 2001)。
有两种同步放电类型:第一种是短时放电(少于500ms),在癫痫患者偶尔出现,无典型临床表现,因此称作“发作间尖峰”(Zifkin & Cracco, 1990)。
第二种是长时放电(数秒) ,与癫痫的临床发作与持续有关,也被称作“发作放电”。
在离体海马脑片,可以通过药理学实验方法来诱导出这两种类型的同步放电。
谷氨酸受体拮抗剂功能原理

谷氨酸受体拮抗剂功能原理英文回答:Glutamate receptor antagonists are a class of drugsthat work by blocking the action of glutamate receptors in the brain. Glutamate is the most abundant excitatory neurotransmitter in the central nervous system and plays a crucial role in various brain functions, including learning, memory, and synaptic plasticity.By antagonizing glutamate receptors, these drugs can inhibit the effects of glutamate and reduce its excitatory activity. This can be beneficial in certain conditionswhere excessive glutamate activity is thought to contribute to pathological processes, such as epilepsy, neurodegenerative disorders, and chronic pain.For example, in the treatment of epilepsy, excessive glutamate release can lead to overexcitation of neurons and the generation of seizures. Glutamate receptor antagonists,such as memantine, can block the effects of glutamate and help prevent the occurrence of seizures.In neurodegenerative disorders like Alzheimer's disease, there is evidence of excessive glutamate release and excitotoxicity, which can contribute to neuronal damage and cognitive decline. Glutamate receptor antagonists, such as riluzole, can help reduce glutamate-mediated excitotoxicity and potentially slow down the progression of the disease.In chronic pain conditions, glutamate receptors in the spinal cord play a crucial role in transmitting pain signals. By blocking these receptors, drugs like ketamine can help alleviate pain and provide relief to patients.Overall, the functional principle of glutamate receptor antagonists lies in their ability to block the action of glutamate and reduce its excitatory effects. This can have therapeutic benefits in conditions where excessiveglutamate activity is involved.中文回答:谷氨酸受体拮抗剂是一类通过阻断大脑中的谷氨酸受体作用的药物。
离子型谷氨酸受体的发现

离子型谷氨酸受体的发现
离子型谷氨酸受体是一类神经递质受体,在神经系统中起着重
要的作用。
这些受体最早是在20世纪80年代被发现的。
离子型谷
氨酸受体分为两类,一类是NMDA受体,另一类是非NMDA受体(包
括AMPA受体和kainate受体)。
NMDA受体是由N-甲基-D-天冬氨酸(NMDA)激活的离子通道受体,它对神经元的兴奋性起着重要作用,参与学习和记忆等功能。
AMPA受体和kainate受体也参与了神经递
质的传递和神经元的兴奋性调节。
离子型谷氨酸受体的发现是通过多个研究团队的努力和合作完
成的。
早期的研究主要集中在对离子型谷氨酸受体在离体大脑切片
中的电生理学特性进行分析,随后通过分子生物学技术,成功克隆
了NMDA受体和非NMDA受体的基因。
这些工作为进一步研究离子型
谷氨酸受体的功能和调控机制奠定了基础。
离子型谷氨酸受体的发现对神经科学领域产生了深远的影响。
它的发现不仅揭示了神经递质在神经元间传递的分子机制,也为神
经系统疾病的治疗提供了新的靶点。
随着对离子型谷氨酸受体的研
究不断深入,人们对其在神经系统发育、学习记忆、神经退行性疾
病等方面的作用有了更深入的理解,为相关疾病的治疗和药物研发
提供了新的思路和途径。
总的来说,离子型谷氨酸受体的发现是神经科学领域的重要里程碑,它的发现和研究为我们理解神经系统的功能和疾病提供了重要的启示,也为相关疾病的治疗和药物研发提供了新的方向和可能性。
谷氨酸受体家族的结构与功能研究

谷氨酸受体家族的结构与功能研究谷氨酸受体(Glutamate Receptor)是一类离子通道蛋白,分为两类:离子型谷氨酸受体(Ionotropic Glutamate Receptor,iGluR)和代谢型谷氨酸受体(Metabotropic Glutamate Receptor,mGluR)。
iGluR是一类离子通道受体,主要分为N型、K型、AMPA型和δ型四个亚型;mGluR是一种G蛋白偶联受体,与G蛋白相互作用,并通过二级信使调节蛋白磷酸化等反应。
这类受体在人类大脑功能方面扮演着重要角色,包括了记忆、学习、辨别和控制行为等方面。
因此,对于谷氨酸受体家族的结构以及功能进行深入研究和探索十分必要。
1. 谷氨酸受体的结构iGluR的结构主要由三部分组成:N端区(N-terminal domain,NTD)、大约约50nm长的膜外外间环节(extracellular ligand-binding domain,LBD)、M2膜间环节以及C肝区(C-terminal domain,CTD)。
这些结构组成了iGluR的功能中心部分,其中的LBD有机会在神经递质神经元前释放到突触小泡中。
相比之下,mGluR的结构相对简单,也更加复杂。
mGluR的结构主要由七个跨膜结构组成,每一个跨膜结构又细分为若干个水相区域:这些水相区域并非是单一的,可能是对应多个对接小分子的活性位点。
2. 谷氨酸受体的功能研究表明,谷氨酸受体在人类大脑中具有很多不同的功能,包括感觉传递、动作调节和记忆形成等方面。
这些功能需要通过与其他分子和蛋白质进行互动和关联来完成。
在功能上,iGluR是一种离子通道受体,而mGluR则与G蛋白相互作用。
在iGluR的两个亚型中,N型和K型亚型分别被发现与负责学习和认知能力、挫折调节、行为状况和成瘾等方面的神经细胞活动相连系,而AMPA型iGluR则被找到与感觉性细胞时相连系。
mGluR作为G蛋白偶联的受体,在各种人类大脑功能方面都扮演着不可替代的角色。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(2)突触前作用机制
突触前抑制主要通过激活PTX敏感的G-蛋白实现其作用。 具体机制可能涉及不同的途径,如抑制突触前Ca2+电导, 激活K+通道而增加K+电流。增加K+电流可以降低突触前膜 的去极化,从而减少Ca2+内流量。另外,增加K+电流可以 增加突触前膜产生动作电位的阈值和降低动作电位的峰值, 从而减少递质的释放。
(二) AMPA&KA受体 AMPA受体激动可引起Na+内流,主要参与正常 的突触传递,形成突触后反应即EPSP。
KA受体激动后的主要作用是引起细胞内Ca2+浓
度的变化,即通过Ca2+信号系统来发挥作用。
谷氨酸受体在昆虫方面的有关研究:
超高效和高选择性是未来农药发展的主要方向。由于 离子通道和神经受体的复杂性和其在不同生物体内表现的 药理性质的差异,为活性高、选择性强的新农药的创制提 供了可能。而目前全球最畅销杀虫剂的3大作用靶标就是
3 受体和组氨酸门控离子通道同属于半胱氨酸环 配体门控离子通道超家族,其结构如下图所示
图1������ 半胱氨酸环受体的共同特征 Fig. 1������ G enera l features o f cy s-loo p recepto rs [ 9 ] A: 受体的侧面图, 两个C 表示高度保守的二硫键桥, NT 表示传递介质的结合位 点, 1、2、3 和4 表 示4个跨膜区; ������ B: 主要由M 2 跨膜区形成孔道的内腔。
流,产生迅速的兴奋性神经传导,这个过程使神经
元细胞膜去极化。但Kainate受体的生理功能目前还
不清楚,而AMPA受体的功能己比较清楚。
AMPA受体有3个结合位点
第一个是谷氨酸结合位点,AM-PA的拮抗剂如托吡 酯和YM9OK结合于此位点; 第二个为非竞争性结合位点,如吡拉西坦结合于 此,使谷氨酸的兴奋性上调,这在记忆和认知功 能方面有重要的生理作用; 第三个结合位点是在离子通道上,能介导多种昆 虫毒素。
一、代谢型谷氨酸受体作用特点(Metabatropic receptors, mGluRs)
GroupⅠ GroupⅡ GroupⅢ mGluR1 PLC1↑ DHPG mGluR5 mGluR2 cAMP↓ ACPD mGluR3 mGluR4 cAMP↓ L-AP4 mGluR6 PPG mGluR7 L-SOP mGluR8 CPCCOET MPEP EGLU CPPG MAP4
一、代谢型谷氨酸受体作用特点 (一) 突触定位
mGluRs 的作用与它的突触分布关系密切。即mGluRs若 是在突触前分布,其作用主要是调节递质的释放,而分布
在突触后则作用是产生突触后效应,即EPSP或IPSP。多数
情况来看,I型mGluRs分布在突触后,而II型和III型mGluRs
主要分布在突触前,一些区域仍然有II型mGluRs分布在突
触后,但III 型mGluRs基本都分布在突触前。
一、代谢型谷氨酸受体作用特点
(二)生物学作用
1. 突触后作用 抑制K+电导,降低K+电流,增加细胞内Ca2+浓度 易化AMPA受体的作用(PKC&Ca2+依赖性)
易化NMDA受体的作用(PKC依赖性)
2. 突触前作用
最早发现的是AP4的突触抑制作用,之后发现了其它的 mGluRs激动剂同样也有抑作用制,主要是II&III型mGluRs 被激 动后发挥的抑制作用。
其中Ⅰ型mGluRs能激活磷脂酶C,产生三磷酸肌醇,使 Ca2+从胞内钙库释放,引起神经细胞兴奋和增加神经细胞 的敏感性,Ⅱ、Ⅲ型mGluRs能抑制腺苷酸环化酶,减少
cAMP的生成,从而抑制谷氨酸的释放。
目前谷氨酸受体系统与药物作用的靶点的研究主要集
中在两个方面:①谷氨酸神经递质的拮抗剂,目前还在动物 实验阶段,尚未获得与临床的相关性。②Ⅱ、Ⅲ型mGluRs 的激动剂:在动物实验中被证明具有神经保护、抗惊厥、 抗焦虑作用,推测可能和它们能减少谷氨酸能神经递质的 释放有关。
(1) Zn2+(一)
(2) H&#ist (4)多胺:甘氨酸依赖性 非甘氨酸依赖性 (5)Mg2+电压依赖性阻断通道,-70mV基本不开放,去极化后 Mg2+与通道亲和力降低并移出通道。意义:使通道受化学, 电压双重控制。 (6)特异性阻断剂 AP5,受体阻断剂,NMDA受体 antagonist; 非特异性阻断剂MK801,通道阻断剂 blocker
NMDA受体是在所有已知的神经递质受体中最独特
的一种,因为它必须在两种激动剂谷氨酸和甘氨酸同时
存在,并且细胞膜去极化时才能被激活。
在NMDA受体上另外还有Mg2+、PCP、Zn2+、H+等 调节结合位点。Mg2+、PCP的结合位点位于离子通道内 部。
PCP、MK801,氯胶酮或美金刚在NMDA受体激 动剂使离子通道开放时很易通过离子通道,使离 子通道阻塞,所以是NMDA受体的非竞争性拮抗 剂,在静息电位时(约-70mv),NMDA受体的离 子通道被Mg2+阻断,即使有谷氨酸与甘氨酸在结 合位点上,也能阻止离子流动,只有细胞膜去极 化时,Mg2+的阻断作用才消失,使Ca2+流入细胞 内。在细胞膜轻微去极化至约-5Omv时,Mg2+的 阻断作用减少,离子通道部分开放。
Ca2+ 与 NMD A反 应的 关系
NMDA受 体 -通道 被Mg2+ 抑制
甘氨酸 加强 NMDA 受体的 电反应
突触后两种电反应,即快反应和慢反应(4.2ms,81.8ms)
谷氨酸能突触及胶质细胞的作用模式图
3.生理作用
(1) 参与突触传递 (2) 参与突触传递可塑性 刺激schaffer侧支引起CA3区兴奋,若给高频刺激 (条件刺激), 再给测试刺激将引起强烈反应, 而且持续时间延长,即突触递长时程增强(longterm potentiation, LTP)。
(1) 作用特征
AP4发挥突触前抑制时不改变突触后神经元的电学特征, 如输入阻抗, 膜电位等。 AP4既能抑制AMPA受体激动的作用,同时也能抑制 NMDA受体激动的效应,意味着突触前释放的递质减少了, 而非突触后效应所致。 AP4发挥突触抑制作用时,外源性谷氨酸对突触后受体的 激动作用不受影响。 AP4可以加强双脉冲刺激引起的突触前抑制作用,后者是 突触前抑制的标志。 AP4引起的突触前抑制,主要改变EPSP的频率而不是幅度。
而且这种抑制作用与浓度相关:
100μmol/L的伊维菌素( ivermectin)可导致保幼激素 的合成速度下降61.5%,但低浓度伊维菌素 (0.1和 1.0μmol/L)对保幼激素的合成没有影响; 10μmol/L的鹅膏蕈氨酸能使保幼激素的合成速度下 降23.4%,高浓度(30~100μmol/L)则没有影响; 而高浓度的木防己苦毒素能够促进保幼激素的合成。
IGluRs主要分布在无脊椎动物的中枢神经和神经 肌肉的连接处,对控制吞咽、运动、感知等可能起着关 键作用,但目前人们对IGluRs的生理功能了解还甚少 。 Liu等在蜚蠊Diplopterapunctata的咽侧体内发现了
IGluRs新的功能,即调控保幼激素的合成。研究发现, 经谷氨酸处理后的咽侧体细胞膜电位出现去极化和超极 化;药理学研究表明,经鹅膏蕈氨酸( ibotenic acid)诱 导的膜超极化可以被木防己苦毒素(picrotoxin)所抑制; 激动剂激活的IGluRs通道能够抑制保幼激素的合成速 度,
二、离子型受体(Ionotropic Receptor, iGluR) : 二、离子型谷氨酸受体作用特点 (一)NMDA受体:
1.一般特征
(1)通道耦联的离子型受体、Ca2+通道。 (2)通道被Mg2+以电压依赖性方式阻断, 受电压、化学因素双重控制。 (3)缓慢的通道动力学特征。
该通道
2. 受体-通道特征
谷氨酸受体
代谢型谷氨酸受体: 通过信号转导机制改
谷氨酸 受体的 分类
变细胞内生化进程,介导多种细胞功能。
亲离子型谷氨酸受体: 能引起细胞膜
对特定离子通透性的改变进而改变膜 电位。
①结构
GluRs与乙酰胆碱(ACh)、r-氨基丁酸(GABAa,
GABAc)、甘氨酸( Gly )受体、5-羟色胺( 5-HT )
一、代谢型谷氨酸受体(mGluRs)
mGluRs没有离子通道,但能通过G蛋白与一
系列第二信使的级联反应来介导谷氨酸兴奋性毒
性。mGluRs可分为3个亚型8个亚单位,Ⅰ型包括
mGluR1和mGluR5,Ⅱ型包括mGluR2和
mGluR3,Ⅲ型包括mGluR4、mGluR6 、
mGluR7 、GluR8。
谷氨酸受体
制作人:张莹
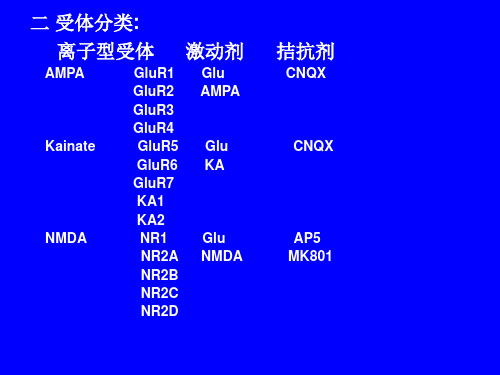
谷氨酸受体 谷氨酸受体的种类 谷氨酸受体的有关研究
谷氨酸
谷氨酸(glutamate,Glu)是代谢中间物是脊 椎动物和无脊椎动物中枢神经系统中一种最 重要的兴奋性神经递质,主要分布于大脑皮 质、海马、小脑和纹状体,在学习、记忆、神 经元可塑性及大脑发育等方面均起重要作用。
研究证实,Glu的神经递质作用是通过兴 奋性氨基酸受体而实现的,受体活性的变化、 受体数目的增减都会对突触效能产生明显的 影响。在生理状态下,谷氨酸是神经细胞间 信息传导的重要媒介,但在病理条件下,谷 氨酸通过兴奋谷氨酸受体介导神经毒性作用。
激动剂
Glu AMPA
拮抗剂
CNQX
Kainate
Glu KA
CNQX
NMDA
Glu NMDA
AP5 MK801
AMPA与Kainate受体一般只允许单价离子通过,
而NMDA受体允许Na+和Ca2+的通过。AMPA与
Kainate受体都与几个不同的亚单位,共同形成一个