癫痫的发病机制及研究进展(全文)
癫痫致病性基因突变对治疗选择影响的研究进展2024(全文)

癫痫致病性基因突变对治疗选择影响的研究进展2024(全文)摘要在过去十余年的研究中,对人类癫痫致病基因以及癫痫临床表现的分子机制的研究都取得了重大进展。
近年来,越来越多的研究表明,致病基因与临床表现的作用机制显著相关。
在不同类型基因突变的患者的抗癫痫治疗中,致病基因的早期识别对于抗癫痫药物的选择有指导意义。
该文系统总结了常见的癫痫致病基因,包括离子通道、细胞代谢和信号通路相关基因等,以及近年来与致病基因相对应的治疗靶点的研究进展。
随着研究的深入,特定基因缺陷及其作用机制为研究新型治疗方法提供了依据。
自1995年发现癫痫相关基因CHRNA4突变起,对癫痫相关基因的研究逐渐深入。
最初,突变的识别主要应用于儿童癫痫的诊断、预后和咨询,然而后续研究结果表明,特定基因突变的识别可以指导癫痫患儿的抗癫痫药物(antiepileptic drugs,AEDs)选择。
儿童癫痫突变基因的功能研究因此有了治疗意义,这是选择或研发针对特定分子缺陷治疗的关键步骤。
本文综述了常见癫痫致病性基因突变的类型,以及特定基因突变对应最适合药物的研究进展。
1 钠离子通道基因突变相关癫痫电压门控钠离子通道参与动作电位的产生和传播,拮抗钠离子通道是目前许多AEDs的主要作用机制。
常见突变的钠离子通道基因包括SCN1A、SCN1B、SCN2A、SCN3A、SCN8A和SCN9A等,这些基因的突变是一部分婴儿和儿童期患耐药遗传性癫痫的共同原因,其中最常见的三个突变基因为SCN1A、SCN2A及SCN8A。
1.1 SCN1A突变相关癫痫SCN1A是与癫痫密切相关的基因之一,而SCN1A相关癫痫的发作表现出显著的表型异质性。
典型疾病包括Dravet综合征和遗传性癫痫伴热性惊厥附加症,其症状表现为从发热性惊厥发作到严重的癫痫性脑病的显著异质性。
即使在具有相同致病基因突变的家族成员之间,临床表型也可能存在差异。
Dravet综合征主要致病基因突变为SCN1A杂合突变,这将导致钠离子通道功能缺失。
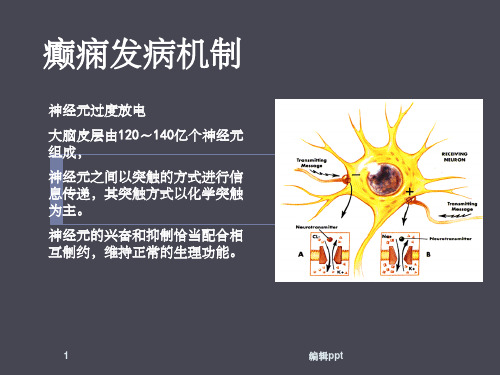
癫痫的发病机制

药物不良反应的观察与处理:抗癫痫药均
有多项不良反应,与血药浓度有关,多数不良反 应应为短暂性反应,缓慢减药即可明显减少,进 食时服药可减少恶心反应,将较大的1次剂量睡 前服用可减少镇静作用。严重特异反应如卡马西 平所致皮疹、肝损害,苯妥英钠所致神经系统损 害,苯巴比妥引起的智能、行为改变等应考虑减 药或停药。
1、一般护理原则 ①心理调适:患者应保持平衡心态,树立治疗信
心。 ②饮食调理:宜进食清淡、无刺激、富于营养的
食物,保持大便通畅,避免饥饿或过饱。 2、活动与休息 癫痫发作时与发作后均应卧床
休息,平时建立良好的生活习惯,劳逸结合, 保持充足的睡眠,减少精神和感觉刺激。
3、避免促发因素 癫痫的诱因有疲劳、饥饿、 缺睡、便秘、经期、饮酒、感情冲动、一过 性代谢紊乱和过敏反应。在日常生活中应尽 量避免这些可能的诱发因素。
1、护理目标: 病人在发作期或发作间期均无 重大安全事故发生,病人没有发生人身损害。
2、护理措施:1)发作期安护理:告知病人有 前驱症状时立即平卧;如果病人是在动态时 发作,陪伴者应抱住病人缓缓就地放倒,适 度扶住病人手脚,以防自伤及碰伤;切勿用 力按压病人抽搐的身体,以免发生骨折、脱 臼;将压舌板或筷子、纱布、手绢、小布卷 等置于病人口腔一侧上下臼齿之间,防止舌、 口唇和颊部咬伤;对于突然发病跌倒而易擦 伤的部位应用棉垫加以保护;癫痫持续状态、 极易躁动或发作停止后意识恢复过程中有短 暂躁动的病人均应专人守护,放置保护性床 档,必要时给予约束带适度约束。
(VPA) 肝功能异常
胰腺炎
托吡酯 震颤、头痛、头晕、胃肠道症状、精神症 (TPM) 状、体重减轻、小脑症
拉莫三嗪 头晕、嗜睡、恶心 (LTG)
儿童多见
基本资料:
癫痫的诊断与治疗研究

癫痫的诊断与治疗研究一、简介癫痫是一种常见的神经系统疾病,其临床表现为反复发作的癫痫发作。
癫痫作为一种确诊困难、治疗难度大的疾病,其病因、发病机制、诊断及治疗一直是神经科学领域的研究热点,本文旨在探讨癫痫的诊断及治疗研究进展。
二、病因据研究表明,癫痫的病因十分复杂,其发病机制涉及到遗传、外伤、感染、药物等多个方面。
其中最常见的病因为脑部结构异常、遗传基因突变、神经元传导异常等。
脑部结构异常包括先天性脑结构异常、肿瘤、外伤、炎症、缺血性损伤等。
遗传基因突变包括离子通道、神经递质及其受体、细胞凋亡相关基因等。
神经元传导异常包括神经元兴奋性增高、抑制性下降等。
三、诊断1. 临床表现癫痫的常见症状为突然发作的抽搐或失神,这些症状具有很强的周期性,且在不同病例中表现也不尽相同,可能是瞬时的,也可能持续数分钟。
在癫痫发作期间,患者表现为面色苍白、出汗、强直阵挛、口吐白沫等症状。
2. 辅助检查在临床上,可以通过多种方法进行癫痫的诊断:神经生理检查(EEG)、脑磁图(MEG)、脑电图(MRI)、脑PET、脑CT等。
其中最常用的检查方法为EEG,临床上多采用24小时或48小时动态监测,可帮助寻找到患者是否存在癫痫发作。
这些辅助检查对癫痫的诊断具有重要的作用。
四、治疗1. 药物治疗目前,癫痫的治疗主要采用药物治疗,包括传统的苯妥英酸、卡马西平、丙戊酸等以及新一代的药物如加巴喷丁、拉莫三嗪等,针对不同类型的癫痫发作,采用药物的选择也会有所不同,治疗时间长短因人而异。
药物治疗虽然具有很好的效果,但是也存在着治疗反复、难以根治的问题。
2. 手术治疗对于药物治疗无效或不能耐受的患者,可以考虑手术治疗。
手术治疗主要用于局限性癫痫,包括脑内肿瘤、脑积水等情况。
手术治疗可减少癫痫发作的发生,提高患者生活质量。
3. 其他治疗方法在药物治疗和手术治疗无效的情况下,患者还可以尝试其他治疗方法,如神经刺激治疗、间歇性电疗等,这些治疗方法通常针对特定的癫痫类型。
儿童癫痫的研究进展

儿童癫痫的研究进展儿童癫痫是一种常见的神经系统疾病,其特征是反复发作的脑部异常放电,导致患儿出现意识丧失,抽搐以及其他脑功能异常表现。
目前,关于儿童癫痫的研究一直在不断进行,包括其发病机制、诊断、治疗等方面的研究。
本文将介绍儿童癫痫研究的进展情况。
一、发病机制的研究儿童癫痫的发病机制非常复杂,目前尚未完全清晰。
不过,研究人员已经对其发病机制有了一定的认识。
研究发现,癫痫的发生与大脑神经元的失去稳定状态和过度兴奋有关。
此外,许多遗传因素也与儿童癫痫的发病有关。
如国外学者近期的一项研究显示有些孩子会出现突变基因,从而影响神经元的传递,进而引发癫痫。
二、诊断的研究儿童癫痫的诊断一直是临床医生和医疗工作者关注的重点。
现在,通过神经成像技术(如脑电图、核磁共振等)以及一些生物标志物的检测,可以对儿童癫痫进行更加准确的诊断。
此外,还可以通过对孩子的不同症状进行评估,制定科学合理的治疗方案,有助于提高儿童癫痫的治疗效果。
三、治疗的研究儿童癫痫的治疗是一个复杂的过程,目前包括一些药物治疗、手术治疗以及其他治疗方式。
随着研究的不断深入,针对儿童癫痫的治疗方案也在不断更新。
研究发现,药物治疗是最常见的治疗方法,但是药物治疗存在一定的不足。
在一些情况下,儿童癫痫患者可能需要手术治疗。
手术治疗主要包括大脑切除术、神经调节器、植入癫痫电极等。
此外,还有一些新型的治疗方法,如外科手术、干细胞治疗和脑深部电刺激等。
这些方法都有望成为有效的儿童癫痫治疗手段。
总之,儿童癫痫的研究包括其发病机制、诊断、治疗等方面的研究。
通过对儿童癫痫的全面了解,我们可以加深对儿童癫痫的认识,并进一步完善儿童癫痫的治疗方案,为儿童的身心健康做出更大的贡献。
同时,还需要我们关注和加强儿童癫痫的防治工作,推动儿童癫痫的早期诊断、早期干预,让更多的孩子远离这种疾病的困扰。
2024癫痫发作预测的研究进展

2024癫病发作预测的研究进展摘要癫病发作的不可预测性严重影响癫病患者的安全、就业能力和生活质量。
一个可靠的癫病发作预测系统能够在发作前至少几分钟提供警告,以便患者能及时采取适当行动。
近年来,脑电图、心电图、周期节律分析、影像学、可穿戴设备等技术的应用为基千多指标精准预测癫病发作提供了新的见解。
为了更好地将癫病预测从研究挑战发展为临床现实,文中就最新的癫病发作预测的研究进展进行综述,并对该领域的未来发展方向提出建议。
癫病是全世界最常见的慢性神经系统疾病之一,影响全球约6900万患者[ 1 ]。
癫病发作存在突发性和不可预测性,容易导致患者溺水、烫伤、骨折、猝死等致残致死性情况。
癫病患者发生意外事故猝死的风险是普通人群猝死的27倍,在神经系统疾病中仅次于脑卒中[ 2 I 3 ]。
为了降低癫病突发带来的严重疾病负担,尤其是伴随着近年来机器学习(machine learning入可穿戴设备(wear.able devices, WD)和可植入设备等领域的发展,对癫病发作机制研究的日益深入以及新型生物标志物的发现,基千各类指标的癫病发作预测研究成为了热点和重点。
我们以"seizure prediction""seizure forecasting""seizure anticipation”作为英文关键词检索PubMed、Embase及Web of Science数据库,尽可能选择5年内发表的相关文献,主要针对癫病患者的脑电图(electroencep h alogram入心电图(electrocard i ogram入周期节律、影像、血液等多维度特征对该领域的研究进展进行综述,旨在对该领域的未来发展方向从临床角度提出建议。
一、基千电生理信号的癫病发作预测研究1脑电图:脑电图是癫病发作预测研究中的最常用、最高效的工具之一。
有研究者发现其可以预测未来几天内的癫病发作,是近年来该领域最重大的突破之一[ 4 / 5 ]。
癫痫研究新进展

癫痫研究新进展一、概述癫痫作为一种常见的神经系统疾病,其发病率在全球范围内均呈上升趋势。
随着医学技术的不断进步和科研工作的深入,癫痫研究取得了显著的进展。
本文将重点介绍癫痫研究的最新成果和发展趋势,包括癫痫的发病机制、诊断技术、治疗方法以及预防策略等方面的内容。
在发病机制方面,研究者们通过大量的基础研究和临床试验,逐渐揭示了癫痫发病的复杂过程。
遗传因素、环境因素以及脑内神经元结构和功能的异常等多种因素相互作用,共同导致了癫痫的发生。
这些研究成果为我们深入了解癫痫的发病机理提供了重要的理论依据。
在诊断技术方面,随着医学影像技术和电生理技术的不断发展,癫痫的诊断手段也日益丰富和精准。
高分辨率的磁共振成像(MRI)和正电子发射断层扫描(PET)等技术能够精确地定位癫痫病灶,为手术治疗提供准确的指导。
脑电图(EEG)等电生理技术也在癫痫的诊断中发挥着重要作用,能够实时监测脑电活动,为癫痫的发作预测和干预提供有力支持。
在治疗方法方面,癫痫的治疗手段日趋多样化和个体化。
除了传统的药物治疗外,手术治疗、神经调控技术以及基因治疗等新型治疗手段也逐步应用于癫痫的治疗。
这些新方法在提高癫痫的治疗效果、减少并发症以及改善患者生活质量等方面显示出巨大的潜力。
在预防策略方面,随着对癫痫发病机制的深入了解,预防策略也逐渐从单一的药物预防向多元化、综合性的方向发展。
通过加强健康教育、改善生活习惯、降低环境污染等措施,可以有效地降低癫痫的发病率和复发率。
针对不同类型的癫痫患者,制定个性化的预防方案,有助于更好地控制癫痫的发作。
癫痫研究新进展为我们提供了更加全面和深入的认识,为癫痫的诊断和治疗提供了更加精准和有效的手段。
随着科研工作的不断深入和技术的不断创新,相信癫痫的诊疗水平将得到进一步提升,为更多患者带来福音。
1. 癫痫的定义与流行病学概况这一古老而又复杂的神经系统疾病,一直以来都吸引着医学界的广泛关注。
它俗称“羊角风”或“羊癫风”,是一种由大脑神经元突发性异常放电引发的慢性疾病,这种异常放电会导致短暂的大脑功能障碍。
癫痫的病因及治疗现状

癫痫的病因及治疗现状癫痫是一种常见的神经系统疾病,其主要特征是反复发作性的全身或局限性肌肉痉挛,有时还伴随着意识丧失或行为异常等症状。
大多数患者的发病年龄在20岁以下,由于其病因复杂,治疗难度较大,对患者身心健康造成了不小的影响。
本文将探讨癫痫的病因及治疗现状。
一、病因癫痫的病因是多种原因综合作用的结果,包括遗传、脑部异常结构或损伤、化学物质或药物中毒、感染、缺氧、电解质紊乱等因素。
其中,遗传因素在癫痫病因中占据了重要地位。
据研究表明,癫痫患者的家庭中有癫痫历史者比正常人群高出3~4倍,同卵双生子的癫痫发病率也比异卵双生子更高。
此外,一些特定基因突变与某些癫痫类型有关。
二、分类根据发作部位和性质可将癫痫分为部分性癫痫和全面性癫痫。
部分性癫痫,即为发作在大脑的一部分,主要表现为肌肉痉挛、双下肢或半身不随意或意识丧失等症状。
全面性癫痫,即为发生脑部不同区域同时发作,最常见的就是Grand Mal,表现为全身强直-阵挛痉挛以及意识丧失。
三、治疗癫痫的治疗主要包括药物治疗、手术治疗和非药物治疗。
药物治疗是目前最常见的治疗方式,常用的抗癫痫药物包括苯妥英、卡马西平、丙戊酸等,瞬时抗癫痫药物主要有利多卡因。
这些药物可以通过控制神经系统的兴奋水平来降低癫痫发作的频率和强度。
但是,由于每种药物的适应症及生理作用不同,所以需要根据病情进行选择。
手术治疗主要适用于药物治疗无效的患者。
手术治疗包括溶栓、创伤性脑损伤、颅内或脑部肿瘤、大型脑囊肿等手术,这些手术可以切除或减轻引起癫痫的原因,从根本上解决癫痫问题。
非药物治疗主要包括按摩疗法、针灸、气功、瑜伽等。
这些治疗方式主要通过刺激或调节神经系统来起到控制发作的作用,虽然非药物治疗方式的有效性仍需进一步的研究和实践,但它依然为那些无法耐受抗癫痫药物的患者提供了另一种治疗选择。
总之,癫痫是一种需要长期持续的治疗疾病,它的治疗需要多学科的协同配合。
虽然现代医学在癫痫治疗上取得了很大进步,但是对于某些难治性癫痫患者而言,仍需求助于其他方面的治疗手段,比如心理治疗和健康生活方式等。
癫痫发病机制

A当前最流行的看法认为其发病体制是大脑神经元过分放电而惹起.最近几年来癫痫发病体制研究遇到宽泛重视,并获得必定进展.现综述以下.1 电解质与癫痫1.1 钙离子;实验研究发现癫病发生前或发生中细胞外离子浓度变化最显然的是钙离子降低.钾离子高升.学者们认为这类细胞外钙离子的减少是因为突出后钙离子被摄入,钙离子内流所致,所以钙离子内流在癫痫发生中起侧重要的作用。
1.2 钠离子:研究表示.伴热性惊厥的浑身性癫痫与电压依靠性钠通道基因 SCNIA和 SCNIB 的突变有关。
不论是钠通道活性增添仍是降低,均可导魏伴热性惊厥的浑身性瘴痛的发生。
1.3 氯离子:检查研究表示 CLcN2 基因的突变所惹起的氧离子通道功能改变与少儿失神癫痛、青少年失神癜瘸’青少年肌阵挛性癫痈和觉悟期浑身强直阵挛发生性癫痫有关.2 神经递质与癫痫2.1 谷氩酸受体:实验研究中发现.在动物模型中使用谷氨酸能激动剂。
可惹起癫痫的发作.在癫痫病人皮质内,跟着搬痫活动的发生,有谷氨酸的显然增添。
谷氨酸受体可分为离子型和代谢型两大类。
2.1.1 离子型谷氪酸受体 (iGluRs):iGluRs 主要包含 N 一甲基一 D 门冬氨酸 (NMDA)、n 一氨基羟甲基恶唑丙酸 (AMPA)和海人酸 (KA)受体,此中 NMDA 受体可分为 7 种亚型 (NR],NR2A~D.NR3),近期研究发此刻电刺激和匹罗卡平致癫模型中,分别阻断 NR2A 和 NR2B亚单位均能显然减少癫痫连续状态惹起的神经元细胞死亡.2.1.2 代谢型谷氨酸受体 (mGluRs):当前 mGluRs 对癫瘸的作用和扩散有重要作用体制尚无定论。
2.2GABA受体:实验研究发现, GABA受体参加了癫痫的发生和发展.中枢神经系统 GABA含鼍降低是神经细胞过分喜悦、引诱同步放电,致使癫痫发生的重要原由之一。
GABA 受体包含 GABAa、GABAb、 GABAc三型, GABAe受体与癫痫的关系当前还没有知.2.2.1GABAa 受体:多项研究均证明喜悦 GABAa 受体能克制癜痫发生,而克制 GABAa 受体则会引发癫痫。
癫痫的自身免疫性病因研究进展(完整版)

癫痫的自身免疫性病因研究进展(完整版)癫痫是最为常见的慢性神经系统疾病之一,影响了全球范围约6 900万患者[1 ]。
即使在规范使用抗癫痫发作药物(anti-seizure medications,ASM)的前提下,仍有约30%患者的癫痫发作控制不理想。
对癫痫疾病强调在病因层次进行管理,而“免疫性”因素已经被列入癫痫病因之一[2 ]。
“自身免疫性癫痫(autoimmune epilepsy)”的概念于2002年的国际自身免疫大会上首次提出,表明自身免疫因素在部分癫痫患者中的重要性[3 ]。
2017年,国际抗癫痫联盟(International League Against Epilepsy,ILAE)出版的癫痫定义和分类指南中,正式将“免疫性”列为癫痫的六大病因(结构性、遗传性、感染性、代谢性、免疫性、未知)之一;“免疫性”病因的患者可能受益于免疫治疗,与其他病因的癫痫患者在治疗方式上存在不同,从而成为研究热点[4 ]。
近年来,“自身免疫性癫痫”这一概念被大量应用,有很多学者对于在自身免疫性脑炎的背景下使用“癫痫”一词提出质疑[5 , 6 ]。
针对这一问题的存在,ILAE自身免疫和炎症特别工作组于2020年提出了两个新概念定义:继发于自身免疫性脑炎的急性症状性痫性发作(acute symptomatic seizures secondary to autoimmune encephalitis,ASSAE)和自身免疫相关癫痫(autoimmune-associated epilepsy,AAE)[7 ]。
2023年在Epilepsia上发表的一篇专家述评,对神经抗体相关癫痫的专业术语进一步澄清,将AAE修改为自身免疫性脑炎相关癫痫(autoimmune encephalitis-associated epilepsy,AEAE),有助于明确此类癫痫发作和脑炎之间的关系[8 ]。
我们通过检索文献,对ASSAE和AEAE这两个诊断实体的定义、流行病学、病因、临床表现、诊断、治疗和预后进行综述,以期能为神经科医生在临床上处理和应对自身免疫性病因有关的癫痫提供参考。
癫痫的发病机制研究进展

[ 关键词 ]癫痫/ 发病机制 ;离子通道 ;神 经递质 ;神 经胶 质细胞
[ 中图分 类号 ] R 4 . 721
[ 文献标识码 ] A
[ 文章 编号 ] 10 24 (0 2 0 O 1 — 3 0 8— 3 4 2 1 ) 2一 1 1 0
d i 0 36 /.sn 10 2 4 .0 2 0 .2 o:1 .9 9 ji .0 8— 34 2 1.0 0 1 s
为 明确 。 1 1 钠离 子通 道 . 19 9 8年 ,Waae 在一 个大 的 lc 等
中[。 。 。
13 钙 离 子 通 道 钙 内流 与 阵 发 性 去极 化 漂 移 、 .
家 系研究 中发 现 伴 热 性 惊 厥 的 全 身 性 癫 痫 ( E S G F +) 是 由于 色 体 1q3 1位 点 上 的 基 因 S N B 9 1. C 1 ( 编码 电压 门控 钠通道 B 1调节 亚基 ) 的点 突变 所 致 。S N B基 因的点 突变导 致 N 通 道 B 基 细 C 1 a 亚 胞外 免 疫球 蛋 白折 叠 结 构 域 中 的半 胱 氨 酸 残 基 被 色 氨酸 残基取 代 , 以致 影 响 了 B亚基 对 o亚 基 动 【 力 学 的调 节 功 能 ,导 致 N 通 道 的反 复 开放 ,从 a 而 引起 神经元 持 久过 度 兴奋 J 。离 体研 究 也 表 明 , 当p 1亚 基 和 仅亚 基 在 体 外 共 同 表 达 时 ,p 亚 基 的突变 可 明显 延 长 稳 态 条 件 下 的神 经 元 的 去 极 化
等情 况下 钠 钾 离 子 泵 的功 能 紊 乱 ,钾 离 子 由胞 内 移 向胞外 ,神 经 元 兴 奋 性 增 高 ,发 生 长 时 间 去 极
颞叶癫痫的病因及发病机制的研究进展

载 脂 蛋 白 E ( pl ortnE,A OE 基 因 A oi po i p e P ) 家族 一 直被 研 究 阿 尔 茨 海 默 症 的学 者 所 关 注 ,携 带有 A O e 因 者 比非 携 带 者 T E发 病 要 早 1 P E 4基 L O 年左 右 ,动 物 实 验 证 据 也 说 明它 的 出现 会 削 弱 大
含 量 的 钙 平 衡 调 节 基 因 1 (a im h mes s cl u o ot i c as
个 家 系 的所 有 成 员 中 发 现 ,说 明 了 遗传 的 异 质
性 。所 以 学 术 界 目前 都认 同 T E是 由多 种 基 因 与 L 多种 环 境 因素共 同作用所 产 生 的结果 。 1几 种 被 公 认 的 导 致 T E 的 家 族 性 基 因 : . L 20 0 2年 ,K lcio [ 常染 色体 显 性 部 分 性 癫 a hkv等 2 a 1 在 痫伴 听觉 先 兆 ( D E F 的家 系 种发 现 了 神经 胶 A PA ) 质 瘤 失 活蛋 白 1 ( uie c ,gi at a d1 1 cn h lmancvt , e i r o i i e L I1 ,并 表 明 L I G ) G 1只在 神 经 元 迁徙 或 皮 质 的重 组 中起 作 用 翻 ,大 部 分 的 L I 因 突 变 导 致 蛋 白 G I基 质 断 裂 、功 能 丧 失 是 可 能 的 潜 在 发 病 机 制 。
颞 叶癫痫 (e p r b plpy L ) 是 主 T m oa l ee i s ,T E lo e
2易 感性 基 因 :对 于 占临床 患 者绝 大 多 数 的散 . 发 性 T E,易 感 基 因不 仅 在 其 发 病 过 程 中 发 挥 一 L
癫痫发病机制的研究进展

氨酸和谷氨酰胺循环是大脑中谷氨酸保持平衡的一个重要
知与癫痫关系密切的氨基酸类物质有谷氨酸、天冬氨酸、 1一氨基丁酸及甘氨酸等。其中研究较多的是谷氨酸(gluta— mate,Glu)和7一氨基丁酸(^y—amino
butyric
回收循环机制,谷氨酸降解酶和谷氨酰胺合成酶在谷氨酸
和谷氨酰胺循环中发挥了重要作用|3】。在许多颞叶癫痫患
复极化过程的调节,决定着动作电位的发放频率和幅度。
KCNQ作为编码电压门控性钾离子通道的基因家族,目前 已经发现5个型KCNQl.KCNQ5,其突变或缺失与癫痫 的发生有密切的联系[28]o研究证实,KCNQl突变能够引 发癫痫”…。电压门控钾离子通道基因-KCNQ2和KCNQ3 突变可诱发良性家族性新生儿惊厥(benign
组成,与G蛋白偶联,调节细胞膜上离子通道和酶的活 性。其中,mGluRl和mGluR5参与了癫痫发作过程。研究 者发现,mGluRl可以启动细胞内某些依赖于蛋白质合成 的信号通路,进而致使海马区神经微环路保持长时间持续 性放电状态,最终诱发癫痫发作9I。mGtuRl和谷氨酸运输
钙离子通道与癫痫有密切关系118-19]。
familial neonatal
GABAa受体亚基Ot 1,B 3,12以及8的突变都与不同表
型的癫痫有密切联系,1 2亚基突变可导致伴热性惊厥的
全身性癫痫发作”2。1”。GABAb受体是G蛋白偶联的跨膜受 体,能产生缓慢而持久的突触反应,它也是临床上治疗癫 痫药物作用的靶点[14l。GABAb受体激活可以开启依赖于G 蛋白的细胞信号通路,进而导致细胞功能的长时间改变,
1.3275.R.20140401.1745.005.html
网络出版时间:2014—04—01
癫痫的发病机制(1)
离子通道异常
静息电位稳定性
神经元单
+
反复兴奋环路
离子分布异常
个动作电位
突触兴奋性
高度同步化的动
突触抑制性
作电位爆发
癫痫
3
编辑ppt
发病情况 门诊和急诊 人群发病率 人群患病率 全国约有患者 我市约有患者
12%-28% 2.5-3.5/万 35-80/万 700-900万 14000-32000人
遗传: 父母亲直接遗传极少见
种以上的药物治疗
国际癫痫发作分类
部分性发作
部分性发作 继发全身性发作
简单部分性发作
复杂部分性发作
全身性发作
失神发作 婴儿痉挛症 失张力发作 全身性强直阵挛发作 肌阵挛发作 阵挛发作 强直发作 Akinetic
部分性发作
源于数量有限的神经元,通常限于同侧 大脑半球,整个发作过程中始终可保持 局限化,亦可进一步扩散到全脑范围
近亲中增加2-6%患病率
特殊类型
定位
影响功能
全身性癫痫伴发热 19q13.1 钠通道
良性新生儿惊厥 20q13.3 钾通道
夜间额叶癫痫
20q13.2 钙通道
年龄: <6月 <20岁 >20岁
与发作类型相关 多有脑部器质改变 占原发性癫痫60% 多见继发性癫痫
癫 特痫发性及癫痫癫痫(id综iop合ath征ic e的pil国epsy际) 分类
16
编辑ppt
复杂部分性发作
精神运动性癫痫;伴意识障碍;病灶多在颞叶
特殊感觉性发作:幻嗅、幻视、幻听等 记忆障碍发作:熟悉感、陌生感、失真实感 情感障碍发作:恐惧、焦虑、不安、愤怒、忧郁、
欣快 思维障碍发作:妄想、强迫思维 自动症
癫痫发生机制的细胞生物学研究

癫痫发生机制的细胞生物学研究癫痫是一种常见的神经系统性疾病,它的特点是反复发作性的脑部异常放电引起的症状,如抽搐、意识丧失、失语、幻觉等。
在目前已知的成千上万的癫痫患者中,研究人员一直在努力解决癫痫发生机制的本质问题。
癫痫的发生机制极其复杂,涉及神经元的兴奋性和抑制性调节、离子通道的变化、神经传递物质的释放等多种生理和病理学的问题,不仅需要多学科的综合研究与探索,在细胞生物学领域方面更有其深层次的研究意义。
我们可以从以下几个角度来探讨癫痫发生机制的细胞生物学研究。
1. 神经元兴奋性和抑制性的调节及其在癫痫中的作用神经元的兴奋性和抑制性是影响癫痫的重要因素之一。
信号传递的过程中,神经元之间会出现不均衡的兴奋和抑制,从而导致癫痫发作。
研究发现,神经元通常会分化为两类,即兴奋性神经元和抑制性神经元。
其中,兴奋性神经元在神经元网络中有着重要的作用,能通过释放兴奋性神经传递物质来协调神经元网络的活动。
抑制性神经元则能通过释放抑制性神经传递物质来平衡神经元的活动,从而保持神经元网络的稳定。
当神经元活动受到抑制性神经元过度影响或兴奋性神经元不足影响时,会出现神经元活动不协调的情况,导致癫痫的发生。
这种现象在近几年的癫痫细胞生物学研究中得到了广泛的关注和探究。
2. 离子通道变化与癫痫发生的关系离子通道的活性调节对于神经元的活动起着至关重要的作用。
神经元的膜电位和细胞内外的离子浓度差异,能够依靠离子通道的开放关键来调节,进而实现神经元的信号传递。
在癫痫发生时,离子通道的活性发生了变化,导致神经元的兴奋性和抑制性发生了错位。
此时,过度的兴奋性会导致神经元的过度放电,难以被抑制,从而使得癫痫的发作。
离子通道变化与癫痫发生的关系在研究癫痫的过程中起着至关重要的作用。
通过不断地探究和解剖离子通道的本质,可以更好地理解癫痫发生的机制。
3. 神经传递物质的释放和癫痫的发生神经传递物质的释放也是影响癫痫发作的重要因素之一。
神经元活动和信号传递过程中会涉及到各种神经传递物质的释放,这些神经传递物质会影响神经元的兴奋性和抑制性,从而影响癫痫的发生。
中医对癫痫病因病机分析

中医对癫痫病因病机分析2010-05-04 16:24⼀、病因学分析(⼀)七情失调,多责惊恐 痫病本⾝即是⼀种发作性神志异常的脑病疾患。
历代医家在对情志因素致痫的病因论述中,多重点强调惊恐致痫的作⽤。
在《素问·举痛论》中就有'巩则⽓下 '、'惊则⽓乱'的论述。
⽽《素问·奇病论》则云'其母有所⼤惊,⽓上⽽不下,精⽓并居,故令⼦发为巅疾也'。
隋·巢元⽅《诸病源候论·⼩⼉杂病诸候·惊痫候》⽈'惊痫者,起于惊怖⼤啼,精神伤动,⽓脉不⾜,因惊⽽作痫也。
'《寿世保元·痫证》⽈:'盖痫疾之原,得之惊,或在母腹之时,或在有⽣之后,必因惊恐⽽致疾。
盖恐则⽓下,惊则⽓乱,恐⽓归肾,惊⽓归⼼。
并于⼼肾,则肝脾独虚,肝虚则⽣风,脾虚则⽣痰,蓄极⽽通,其发也暴,故令风痰上涌⽽痫作矣。
可见在七情失调、情志过极的诸情志因素中,以惊恐最为常见和重要。
由于突受⼤恐,致使⽓机逆乱,进⽽损伤脏腑,肝肾受损则易致阴不敛阳⽽⽣热⽣风,脾胃受损则精微不布,痰浊内聚。
经久不化,遇逢诱因,则痰浊或随⽓逆,或随⽕炎,或随风动,蒙闭脑窍,发为癫痫。
⼜《景岳全书·杂症谟·癫狂痴呆》指出⼩⼉痫证'有从胎⽓⽽得者,有从⽣后受⽽得者,盖⼩⼉神⽓尚弱,惊则肝胆夺⽓⽽神不守舍,舍空则正⽓不能⽴⽽痰邪⾜以乱之。
'说明⼩⼉脏腑娇嫩,元⽓未充,神⽓怯弱,或素蕴风痰,复因惊恐,⽽正⽓不能主,则痰邪作乱发为癫痫,补充了惊恐等情志因素致痫多基于正⽓不⾜的内在因素,同时也反映了惊恐致痫更以⼩⼉为多。
(⼆)先天因素,尤重孕产 综合⽂献分析,中医学明确指出先天因素在癫痫病发⽣中的作⽤,并多从孕妇调养失当和胎⼉发育不良两个⽅⾯说明痫病的先天原因。
《素问·奇病论》云:'⼈⽣⽽有病巅疾者……,病名为胎病,此得之在母腹中时,其母有所⼤惊,⽓上⽽不下,精⽓并居,故令⼦发为巅疾也。
癫痫的发病机制与治疗

Rasmussen综合征(部分性连续性癫痫)早期出现肌阵挛, 伴进行性弥漫性神经系统损害表现
部分性发作持续状态
临床表现及分类
部分性发作持续状态
临床表现及分类
边缘叶性癫痫持续状态 常表现意识障碍(模糊)与精神症状又称精神运动性癫 痫状态, 常见于颞叶癫痫。 偏侧抽搐状态伴偏侧轻瘫 多发生于幼儿, 表现一侧抽搐伴发作后一过性或永久性同侧肢体瘫痪。
治疗
1
癫痫持续状态 GTCS-自动症 ↓ ↓
2
在给氧防护的同时, 防止跌伤\自伤与伤人 从速制止发作。 防止窒息与舌咬伤 防止骨折与脱臼
3
01
从速控制发作, 同时给予支持与对症治疗
02
保持呼吸道通畅
03
纠正酸碱平衡\电解质紊乱
发作先兆 是指在大发作前数秒钟内病人出现的幻觉、错觉、自动症或局部肌肉阵挛抽动等症状,而且在大发作后,常能回忆起昏迷前所出现的症状
癫痫综合征 是指在特定的年龄、不同病因或促发条件下,某些临床表现和体征通常固定地组合在一起所出现的痫性疾病。
自动症 是指在癫痫发作的过程中或发作之后,病人的意识尚处于混浊状态时所出现的一些或多或少的不自主、无意义、无目的的刻板样动作,清醒后不能回忆。临床表现形式多样。
间歇期护理
护理措施
及时补充维生素和微量元素:抗癫痫药能引起维生素K、叶酸、维生素D和钙、镁等物质的缺乏。
饱餐与饥饿:一次服用大量甜食后,大量的糖分进入血液,会激发胰腺分泌过多的胰岛素,从而使血糖很快下降,血糖过低导致脑的能量供应不足而促发癫痫发作。同样,饥饿也会使癫痫容易发作。
饮料:大量饮用或饮用太浓的茶、咖啡同样可诱发癫痫。因为这些饮料中或多或少的含有中枢兴奋性物质,使抗发作能力降低、诱发癫痫发作。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
癫痫的发病机制及研究进展(全文)
癫痫的研究者们从电生理、离子及离子通道、分子生物学、免疫等多个角度探索癫痫的发病机制,获得了丰富的资料。
癫痫的神经电生理
神经细胞动作电位中的兴奋及抑制性突触后电位与癫痫发生密切相关。
兴奋性突触后电位(EPSP)是以兴奋性氨基酸的谷氨酸,也可能有天门冬氨酸作为递质引起的突触后电位,电刺激内嗅皮质到海马齿状颗粒细胞的谷氨酸能梨状细胞层可提高动物神经元的兴奋性,直至癫痫发作。
在动物模型中使用谷氨酸能激动剂,可引起癫痫发作。
还有人发现在癫痫病人皮质内,随着癫痫活动的发生,谷氨酸明显增加,谷氨酸的兴奋作用促成了癫痫的发生和发展。
抑制性突触后电位(IPSP)是由v-氨基丁酸(GABA)介导的突触后电位,GABA是中枢内重要的抑制性神经递质。
癫痫病人GABA神经元的正常抑制功能有明显的损伤。
研究发现,癫痫病灶内GABA神经元明显减少,在癫痫敏感的脑组织内,GABA受体的量也有降低。
癫痫的离子变化
在癫痫和癫痫发作的模型中发现发作前或发作中细胞外离子浓度变化最明显的是钙离子降低,钾离子升高。
病性放电中细胞外钙离子减少和钾离子增多是同步变化的。
电休克诱导的癫痫中发现新皮质和海马细胞外钾离子比正常基线上升4倍,而钙离子浓度在新皮质降低到0.45mmol/L,在海马CAl区降低到0.7mmol/L。
钾离子的增多是由于钾外流所致,钾外流依赖于去极化后内流的钙离子激活钙离子依赖性钾通道。
钙离子通道阻滞剂可预防细胞外钙离子的减少,抑制癫痫发作。
癫痫与苔藓纤维发芽
苔藓纤维,即齿状回颗粒细胞的轴突,在正常情况下投射到海马CA3区的辉层。
研究表明,颞叶癫痫病人以及海藻酸、毛果芸香碱和电点燃诱导的癫痫模型,均存在广泛的苔藓纤维发芽现象。
目前电生理研究证实,海藻酸致惊大鼠海马苔藓纤维发芽可重建兴奋性环路,
并形成正反馈,因而导致癫痫形成。
癫痫的分子生物学
基因突变单基因突变很少,仅有小部分癫痫和癫痫综合征以单基因突变的方式遗传。
目前发现的有良性家族性新生儿惊厥(BFNC)和常染色体显性遗传夜间发作性额叶癫痫。
许多原发性癫痫呈现出复杂的多基因遗传,即由许多微效累加基因和某些环境因素共同作用而引起的遗传病,如儿童和青少年失神癫痫、青少年肌阵挛痫、原发性全身强直阵挛性癫痫、原发性部分性癫痫等。
在多基因遗传中研究最多的是青少年肌阵挛癫痫。
染色体异常染色体畸变均有可能导致癫痫的发生。
如Downs综合征,是由于染色体21三体所致,其中5%
`~10%的病人出现癫痫发作。
染色体13、18或22三体的病人,25%~50%将会出现癫痫发作,染色体4p部分缺失(Wolf-Hirschhorn综合征)将导致70%左右的病人出现癫痫发作,但染色体5p缺失很少伴有癫痫发作。
线粒体突变线粒体基因的突变常常通过母亲遗传,部分病人可表现为癫痫发作,如肌阵挛性癫痫伴破碎红纤维(MERRF)是一种性连母系遗传病,mtDNA的tRNAIys基因中A8844G突变是MERRF最常见的原因。
癫痫的免疫学异常
目前,国内外已用多种免疫技术成功地造出了癫痫动物实验模型,同时在临床研究中亦已证实了癫痫患者在治疗前后确实存在有多种免疫异常。
学者们认为各种原因引起的脑损伤破坏了血脑屏障,导致脑组织抗原释放入血循环,暴露在机体的免疫系统之下,成为异种抗原,启动免疫反应而产生了抗脑组织抗体。
抗脑组织抗体又通过损伤的血脑屏障进入脑,作用于突触,封闭抑制性受体,减少抑制性冲动,导致癫痫发作。
癫痫的发病机制研究方面取得了丰富的研究成果,但其仍不十分清楚,结论尚不统一,还有待进一步研究。
注:本文为网友上传,不代表本站观点。