植物生理学实验技术(刘萍,李明军主编)思维导图
《z植物生理学试验》PPT课件

精选ppt
7
【方法与步骤】
取材1-2克新鲜植物材料洗净包在锡箔纸中,放入液氮或 低温冰箱中将细胞杀死→取出剪碎放入注射器溶冰,用加 压法将胞液挤出,存于Eppendorf管待测→取20微升待测 液于冰点渗透压计的测定管测定→提出探头,擦拭后可继 续下一个样品的测定。
【思考题】
冰点下降法与质壁分离法的原理有何不同?
精选ppt
38
仪器与用品
1、实验仪器 分光光度计,恒温水浴,分 析天平,烘箱,刻度试管,漏斗,活性 炭,酒精(20%)
2、实验试剂 酒精(20%),葡萄糖标准 液,葸酮试剂
3、实验材料 各种植物叶片、种子、果实
精选ppt
39
方法与步骤
1、可溶性糖的提取 干材料50mg→10ml刻度离心管(4ml80% 酒精)→水浴40min →离心 →上清液 +10mg活性炭 →脱色(80℃)30 min→定 容至10ml →过滤 →滤液
【仪器与用品】
广口瓶 温度计 酸式滴定管 干燥管 尼龙网制小篮 Ba(OH)2麝香草酚酞 5%乙醇 草酸(重结晶)
小麦种子
精选ppt
35
【实验步骤】
安排实验装置→放入萌发种子(同时用煮死的种子作 对照)于小篮内→滴定→
【思考题】
比较不同类型种子的呼吸强度。
精选ppt
36
实验9 可溶性总糖类的测定
精选ppt
22
[思考题]
植物营养必须的大量元素和微量元素有哪些 植物的大量元素和微量元素缺乏时的表现症状有什么 不同 如何做好植物的缺素实验
精选ppt
23
实验6 叶绿体色素的提取、分 离及理化性质的鉴定
精选ppt
24
【实验原理】
《植物生理学复习》课件

病原菌对植物生长的影响,以及植物对病原菌的响应和适应。
THANKS
感谢观看
植物的水分平衡
植物通过调节根系吸水和叶片蒸腾作 用来维持水分平衡,以适应不同的环 境条件。
植物对矿质元素的吸收与运
01
02
03
矿质元素的重要性
矿质元素是植物正常生长 和发育所必需的,包括氮 、磷、钾、钙、镁等。
矿质元素的吸收
植物通过根系吸收土壤中 的矿质元素,进入植物体 内的矿质元素会被运输到 各个组织器官。
抗旱性与环境
干旱环境对植物生长的影响,以及植 物对干旱环境的响应和适应。
植物的抗寒性
抗寒性概述
抗寒性机制
植物在寒冷环境中的适应能力,包括低温 诱导基因表达、冰晶形成等机制。
植物通过产生抗冻蛋白、增加细胞内糖分 等方式增强抗寒性。
抗寒性应用
抗寒性与环境
在农业上,培育抗寒性强的品种以提高作 物的越冬能力和适应性。
ቤተ መጻሕፍቲ ባይዱ矿质元素的运输
矿质元素在植物体内通过 木质部和韧皮部的运输系 统进行长距离运输,以满 足不同组织器官的需求。
植物对矿质元素的同化与利用
矿质元素的同化
01
植物将吸收的矿质元素合成自身所需的化合物,如蛋白质、核
酸、叶绿素等。
矿质元素的利用
02
植物利用合成的化合物进行各种生理生化反应,以维持正常的
代谢活动。
矿质元素缺乏与过量
03
缺乏或过量摄入矿质元素会对植物的生长和发育产生不利影响
,因此需要合理施肥以满足植物对矿质元素的需求。
04
植物的光合作用与呼吸作用
光合作用的概念与意义
概念
光合作用是植物利用光能将二氧化碳和水转化为有机物和氧 气的过程。
植物生理学植物的生殖和衰老 ppt课件

图 白花丹的雄性生殖单位
A.示两个精细胞间的连接(箭头处); B.示两个精细胞的异型性及其与 营养核间的联系
➢ 两个互相连接的精细胞与营养核作为一个功能团, 被称为雄性生殖单位(male germ unit,MGU)。
➢ (5)色素 成熟的花粉具有颜色,这是由于花粉外 壁中存在花色素苷、类胡萝卜素等色素的缘故。色 素具有招引昆虫传粉,防止紫外光对花粉的伤害等 作用。
➢ (6)矿质元素 花粉与其他植物组织一样,含有磷、 钙、钾、镁、硫、锰等多种矿质元素。不同植物中, 精细胞以及花粉管所含的元素种类与含量大体相似, 但花粉外壁中的元素组成有较大的差异。
➢雄性生殖单位中的营 养核和雌性生殖单位 中的助细胞在双受精 后消亡。
双受精的过程
➢ 助细胞释放Ca2+可能对细胞融合起重要的作用。
➢ 现已从玉米等植物中分离出精细胞和卵细胞, 在高Ca2+(5~50 mmol·L-1CaCl2)条件下, 可让精细胞和卵细胞在体外发生自动融合。
五、花粉和雌蕊的相互识别
➢ 蔗糖在花粉中含量很高,一般占可溶性糖的20%~25%。缺 乏蔗糖时花粉发育不良,如不育株的小麦花粉中就缺少蔗糖。
➢ (4)植物激素 花粉中含有生长素、赤霉素、细胞分裂素、 油菜素内酯和乙烯等植物激素。这些激素的存在对花粉的萌 发,花粉管的伸长以及受精、结实都起着重要的调节作用。 例如未授予花粉的雌蕊一般不能生长,这显然与缺少花粉激 素的作用有关。
图 小孢子发育示意图
各种各样的花粉
1.构造
1)壁物质
植物生理学课件-PPT课件

产生与发展
农政全书
齐民要术
陈旉农书
Hale Waihona Puke 王祯农书农政全书齐民要术
陈旉农书
王祯农书
农政全书
齐民要术
陈旉农书
王祯农书
产生与发展
●植物生理学诞生的三大标志:
▲J.von.Liebig(1840):创立矿质营养学说 ▲J.von.Sachs(1882) :撰写《植物生理学讲义》 ▲W.Pfeffer(1904):出版三卷本《植物生理学》巨著
研究内容
●生长发育生理
▲植物营养生长 ▲植物生殖生长 ▲植物衰老与脱落生理
研究内容
●逆境生理(stress Physiology)
▲抗旱机理 ▲抗涝机理 ▲抗寒机理 ▲抗热机理 ▲抗盐机理 ▲植物与生态环境保护
三、植物生理学的产生和发展
●植物生理学的奠基 ●植物生理学的诞生与成长 ●植物生理学的迅速发展
任务与展望
(二)植物生理学展望
(1)植物分子生理学(从生物大分子到复杂生命活动) (2)信号传递(实现生命整体性的重要环节) (3)代谢及其调节(生命活动的物质与能量基础) (4)植物环境生理(生命的协同进化与适应
五、学习植物生理学的方法
★充分认识本课程的重要性 ★重视基本概念、基本理论学习 ★理论联系实际 ★充分利用网络信息资源
产生与发展
●植物生理学的奠基
▲Van Helmont(1577-1644):柳树生长实验 ▲J.Woodward(1699) :发现植物对矿质营养的需求 ▲Priestley(1776):发现植物可以改善空气 ▲Ingenhousz(1779) :发现植物只有在光下才能净化空气 ▲T.de.Saussure(1767-1845):植物在光下利用CO2进行光合 ▲voisier(18世纪80年代):发现呼吸作用
高中生物学科思维导图(人教版必修二)-高中生物必修二思维导图

具有易于区分的性状,实验结果很容易观察和分析。
如:高茎和矮茎;圆粒和皱粒人工传粉过程 人工去雄→套袋隔离→人工授粉→再套袋隔离相关符号意义P :亲本; F 1:子一代; F 2:子二代;♀ :母本;♂:父本;×: 杂交; 性状 生物所表现出来的形态结构(双眼皮) 、生理特征( B 型血)和行为方式(左撇子)① 生物的性状是由遗传因子(基因)决定的 ②体细胞中遗传因子(基因)是成对存在的③ 形成配子时,成对的遗传因子(基因)彼此分离,分别进入不同的配子。
配子中只含有每对遗传因子(基因)中的一个结 论 控制果蝇白眼的基因在 X 染色体上,证明了萨顿假说的正确性一条染色体上有多个基因基因和染色体的关系基因在染色体上呈线性排列隐性性状(如豌豆的矮茎)等位基因位于同源染色体上 显性基因 决定显性性状的基因(用大写字母表示) 控制相对性状的基因隐形基因决定隐性性状的基因(用小写字母表示)性状分离 在杂种后代中,同时出现显性性状和隐性性状的现象 表现型 生物个体表现出来的性状,如豌豆的高茎和矮茎基因型 与表现型有关的基因组成, 如高茎豌豆的基因型是 DD 或 Dd,矮茎豌豆的基因型是 dd纯合子 基因组成相同的个体,如:DD 、 dd 、YYRR 、 yyrr杂合子基因组成不同的个体,如 Dd 、YyRr 、Yyrr 、yyRr 显性性状(如豌豆的高茎)分为相关概念实验过程及现象P 高茎×矮茎→F 1(全为高茎);F 1自交→F 2(高茎:矮茎 =3:1)相对性状 同种生物的同一种性状的不同表现类型豌豆的特点自花传粉、闭花受粉,自然状态下一般为纯种解释(提出假说)遗传因子 的发现分离定律的发现 过程及其内容自由组合定律的 发现过程及其内容萨顿的假说基因在染色体上摩尔根关于果蝇 的实验证据验证(演绎推理)④受精时,雌雄配子的结合是随机的设计测交实验: F 1 与隐性纯合子杂交,推测后代高茎:矮茎 =1:1 实验验证:在得到的 64株后代中, 30株是高茎, 34株是矮茎,比例接 近 1:1,验证了以上解释的正确性在杂合子的细胞中,位于一对同源染色体上的等位基因, 具有一定的独立性;分离定律内容 在减数分裂形成配子的过程中, 等位基因会随同源染色体的分开而分离, 分别进入两个配子中,独立地随配子遗传给后代实验过程及现象P 黄色圆粒×绿色皱粒→F 1(全为黄色圆粒);F 1自交→F 2实验过程及现象(黄色圆粒:黄色皱粒:绿色圆粒:绿色皱粒 =9:3:3:1)①纯种黄色圆粒和纯种绿色皱粒豌豆的基因型成分别是 YYRR 和yyrr,F 1 的基因型是 YyRr解释(提出假说)验证(演绎推理)自由组合定律内容②F 1形成配子时,每对基因彼此分离,不同对的基因自由组合。
人教版高中生物必修一思维导图(高清大图可以放大)(2021年-

人教版高中生物必修一思维导图一、绪论1. 生物学的定义与研究对象2. 生物学的分支学科3. 生物学的科学方法二、细胞的分子组成1. 细胞的结构与功能2. 细胞膜的组成与功能3. 细胞质基质与细胞器4. 细胞核的结构与功能三、细胞的代谢1. 细胞呼吸2. 糖酵解3. 三羧酸循环4. 电子传递链与氧化磷酸化5. 光合作用6. 碳循环与能量流动四、细胞的生命周期1. 细胞分裂2. 细胞周期3. 细胞分化与发育4. 细胞衰老与死亡五、遗传信息的传递1. DNA的结构与功能2. 基因的表达与调控3. 基因突变与进化4. 中心法则与遗传信息的复制六、生物的多样性1. 生物的分类与命名2. 生物的进化历程3. 物种的形成与灭绝4. 生物多样性的保护与利用七、生态学1. 生态系统的组成与功能2. 能量流动与物质循环3. 生物群落与生态位4. 生态平衡与人类活动八、人类健康与生物技术1. 生物技术在医学中的应用2. 生物技术在农业中的应用3. 生物技术在环境保护中的应用4. 生物技术在能源开发中的应用九、生物伦理与可持续发展1. 生物技术的伦理问题2. 生物多样性与可持续发展3. 人类活动对生物多样性的影响4. 生物伦理与人类福祉5. 生物技术的未来发展趋势人教版高中生物必修一思维导图一、绪论1. 生物学的定义与研究对象生物学是研究生命现象及其规律的自然科学,研究对象包括所有生物及其相互关系。
2. 生物学的分支学科生物学包括多个分支学科,如细胞学、遗传学、生态学、生物化学等。
3. 生物学的科学方法生物学研究采用实验、观察、比较等方法,以揭示生命现象的本质和规律。
二、细胞的分子组成1. 细胞的结构与功能细胞是生物体的基本单位,具有自我复制、代谢、遗传等功能。
2. 细胞膜的组成与功能细胞膜由磷脂双分子层和蛋白质组成,具有选择性通透性、物质运输、细胞识别等功能。
3. 细胞质基质与细胞器细胞质基质是细胞内的液态环境,细胞器包括线粒体、内质网、高尔基体等,各具特定功能。
生物必修一思维导图
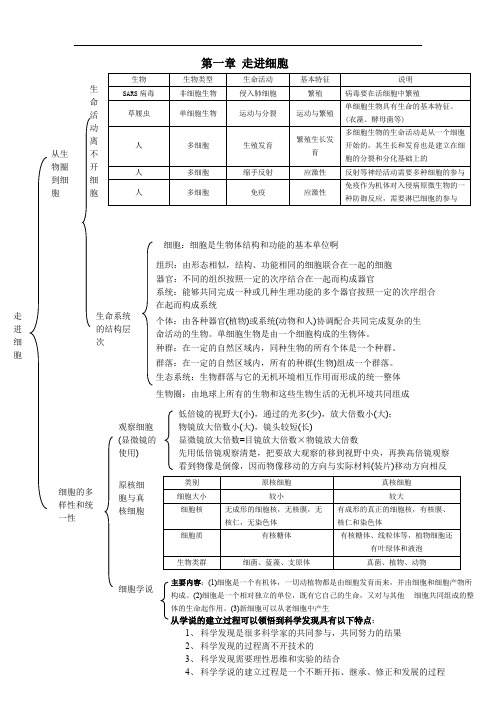
类别 细胞大小 细胞核
细胞质
生物类群
原核细胞 较小
无成形的细胞核,无核膜,无 核仁,无染色体
有核糖体
细菌、蓝藻、支原体
真核细胞 较大
有成形的真正的细胞核,有核膜、 核仁和染色体 有核糖体、线粒体等,植物细胞还
有叶绿体和液泡 真菌、植物、动物
细胞学说
主要内容:(1)细胞是一个有机体,一切动植物都是由细胞发育而来,并由细胞和细胞产物所 构成。(2)细胞是一个相对独立的单位,既有它自己的生命,又对与其他 细胞共同组成的整 体的生命起作用。(3)新细胞可以从老细胞中产生
组成元素:主要由 C、H、O、N 等元素组成,有些含有 S、Fe 等
R
相对分子质量:几千~100 万以上,属于大分子化合物
NH2 C COOH
基本单位:氨基酸,大约有 20 多种,
结构通式:
H
结构特点是至少含有一个氨
基(-NH2 和一个羧基(-COOH),并且都有一个有一个氨基(-NH2 和一个羧基(-COOH)连接在同一个碳原子上,将氨
基酸区别为不同的种类的依据是 R 基(侧链基团)。
形成过程:(1)脱水缩合:
R1
R2
R1
R2
生命活
NH2 C COOH + H2N C COOH
NH2 C CO HN C COOH
动的主 要承担 者—蛋 白质
H
H
H
H
H2O
肽键
二肽 (2)肽链:两(三)个氨基酸缩合的化合物叫二(三)肽,含有一(二)个肽键,脱掉
出型
主动运输:各种离子、
物质跨 膜运输 的方式
由高浓度到低浓度, 不要能量
由低浓度到高浓度,
植物生理学 植物生长物质PPT课件

自由态、结合态,结合态无活性,是储藏和运 输形式。
29
二 赤霉素的分布与运输
分布广:GA普遍存在于高等植物、蕨类、藻类、真菌、 细菌。
分布: 含量最高部位是植株生长旺盛部位。
运输方向: 双向运输。沿木质部向上,沿韧皮部向下。
30
GA表达 的荧光染 色,在根 尖,茎尖, 花等部位 含量多
前言
点击此处输入 相关文本内容
标题添加
点击此处输入相 关文本内容
点击此处输入 相关文本内容
2
植物生长素错当农药 晚稻疯长比人高
图2
图1
因用错农药而疯长的晚稻鹤立“稻”群,
十分醒目
3
利用生长物质调控石斛兰春节开花
图3 中国生产
图4 日本生产
4
❖ 植物生长物质(plant growth substances) 分为两类:植物激素和植物生长调节剂。
14
15
四 生长素的作用
1 促进营养器官的伸长。 促进伸长的最适浓度:茎>芽>根; 器官对IAA的敏感性,根>芽>茎。 促进效应以伸长区最为明显。
16
两面性:低浓 度的生长素促 进根、茎、芽 的生长,高浓 度则抑制其生 长。高浓度 2·4D(1000ppm) 杀死双子叶杂 草。 对IAA敏感性: 根>芽>茎
1 促进细胞分裂 细胞分裂素的主要生理功能就是促进细胞的分
裂。产生愈伤组织,使叶用蔬菜增产。 生长素、赤霉素和细胞分裂素都有促进细胞分
裂的效应,但它们各自所起的作用不同:
56
生长素只促进核的分裂(因促进了DNA的合成), 而与细胞质的分裂无关。 而细胞分裂素主要是对 细胞质的分裂起作用,所以,细胞分裂素促进细 胞分裂的效应只有在生长素存在的前提下才能表 现出来。
第一章绪论powerpoint 演示文稿 植物生理学的内容及相互关系

李比希指出,土壤中矿质养分的含 量是有限的,必将随着耕种时间的推移 而日益减少,因此必须增施矿质肥料予 以补充,否则土壤肥力水平将日趋衰 竭,作物产量将逐渐下降。这个主张即 著名的“归还学说”。它正确地指出了土 壤对植物营养的重要作用,从而促进了 田间试验、温室试验和实验室化学分析 的兴起以及化肥工业的发展,并为土壤 学的发展作出了划时代的贡献。
Plant physiology Wan-zuoxi Hubei Institute for Nationalities 2009
绪论
荷兰人医生兼炼金术士凡·海尔蒙(Van Helmont 1577-1644) 是最早用科学实验来探讨植物营养本质的人,他用柳树枝条做 试验,探索植物长大的物质来源。
Plant physiology Wan-zuoxi Hubei Institute for Nationalities 2009
绪论
到了1771年,氧的发现者英国英国化学家普里斯特利(J. Priestley 1733-1804)才发现绿色植物有净化空气的作用。
J. Priestley把老鼠放在密闭的玻璃钟罩里,不久老鼠便窒息而死, 其中的空气也失去助燃能力;但若在钟罩里放入绿色植物,经过几天, 钟罩里的空气能重新恢复助燃能力并支持老鼠的生存。后来,有人重复 普利斯特利的实验而得出相反的结果,即植物也能把空气变坏。
全套高中生物思维导图word清晰打印版

思维导图高中生物
快乐学习,一目了然
全套高中生物思维导图(清晰打印版)
•
•
•
•
•
•
•
•
•
例
•第2节生物膜的流动镶嵌
模型
•第3节物质跨膜运输的方
式
•第5章细胞的能量供应
和利用
•第1节降低化学反应活化
能的酶
•第2节细胞的能量“通
货”──ATli
•第3节ATli的主要来源
──细胞呼吸
•第4节能量之源──光与
光合作用
•第6章细胞的生命历程•第1节细胞的增殖
•第2节细胞的分化•第3节细胞的衰老和凋亡•第4节细胞的癌变
(选学)
•第5章基因突变及其他
变异
•第1节基因突变和基因重
组
•第2节染色体变异
•第3节人类遗传病
•第6章从杂交育种到基
因工程
•第1节杂交育种与诱变育
种
•第2节基因工程及其应用
•第7章现代生物进化理
论
•第1节现代生物进化理论
的由来
•第2节现代生物进化理论
的主要内容
定性
•第1节生态系统的结构
•第2节生态系统的能量流
动
的洗涤效果
•课题3 酵母细胞的固定
化
•专题5 DNA和蛋白质技
术
•课题1 DNA的粗提取与
鉴定
•课题2 多聚酶链式反应
扩增DNA片段
•课题3 血红蛋白的提取
和分离
•专题6 植物有效成分的
提取
•课题1 植物芳香油的提
取
•课题2 胡萝卜素的提取。
《生物》必修一思维导图(思维导图)
组成元素:主要由C、H、O、N等元素组成,有些含有S、Fe等
相对分子质量:几千~100万以上,属于大分子化合物
基本单位:氨基酸,大约有20多种
结构通式
第二章:组成细胞的分子 第三章:细胞的基本结构
生命活动的主要主要承担者-蛋白质
结构特点是至少含有一个氨基(NH2和一个羧基(COOH),并且都有一个有一个 氨基(NH2和一个羧基(COOH)连接在同一个碳原子上,将氨基酸区别为不同的 种类的依据是R基(侧链基团)。
探索历程
20世纪六十年代发现细胞膜并非是静态的
1970年细胞融合等实验表明细胞膜具有流动性 1972年桑格和尼克森提出流动镶嵌模型为大多数人所接受
生物膜的流动镶嵌模型
磷脂双分子层构成了膜的基本支架,具有流功性。蛋白质 分子有的镶在磷脂双分子层表面,有的部分或全部嵌入磷 脂双分子层中有的贯穿于整个磷脂双分子层。大多数蛋白 质分子也是可以运动的。
人
多细胞
缩手反射
人
多细胞
免疫
应激性 应激性
反射等神经活动需要多种细胞的参 与
免疫作为机体对入侵病原微生物的 种防御反应,需要淋巴细胞的参与
细胞:细胞是生物体结构和功能的基本单位啊
组织:由形态相似,结构、功能相同的细胞联合在一起的细胞
器官:不同的组织按照一定的次序结合在一起而构成器官
生命系统的结构层次
(3)空间结构
一条或几条肽链通过一定的化学键互相链接在一起,形成具有复杂空间结构的蛋 白质。高温、强酸强碱和重金属都会破坏蛋白质的空间结构。
结构的多样性:
组成蛋白质的氨基酸数目不同、氨基酸的种类不同、氨基酸排列顺序不同、多肽链的盘曲、折叠方 式及其形成的空间结构千变万化
植物生理学全课程讲义

植物生理学绪论一植物生理学的定义和内容研究植物生命活动规律和机理及其与环境相互关系的科学。
植物生命活动:从种子开始到形成种子的过程中所进行的一切生理活动。
植物生命活动形式:代谢过程、生长发育过程、植物对环境的反应植物生命活动的实质:物质转化、能量转化、信息转化、形态建成、类型变异1 物质转化体外无机物[H2O、CO2、矿质(根叶)]→体内有机物[蛋白质核酸脂肪、碳水化合物] →体外无机物[CO2 H2O]→植物再利用2 能量转化光能(光子)→电能(高能电子)→不稳定化学能(ATP,NADPH)→稳定化学能(有机物)→热能、渗透能、机械能、电能3 信息转化[1]物理信息:环境因子光、温、水、气[2]化学信息:内源激素、某些特异蛋白(钙调蛋白、光敏色素、膜结合酶)[3]遗传信息:核酸4 形态建成种子→营养体(根茎叶)→开花→结果→种子5 类型变异植物对复杂生态条件和特殊环境变化的综合反应植物生命活动的“三性”v植物的整体性v植物和环境的统一性v植物的变化发展性Ø植物生命活动的特殊性1 有无限生长的特性2 生活的自养性3 植物细胞的全能性和植株的再生能力强4 具有较强的抗性和适应性5 植物对无机物的固定能力强6植物具有发达的维管束植物生理学的内容1、植物细胞结构及功能生理﹕2、代谢生理:水分代谢、矿质营养、光合作用、呼吸作用等3、生长发育生理:种子萌发、营养生长生理、生殖生理、成熟衰老4、环境生理(抗性生理)以上的基本关系光合、呼吸作用→生长、分化水分、矿物质运输发育、成熟(功能代谢生理) (发育生理)↖↗环境因子(抗性生理)(温、光、水、气)二植物生理学的产生与发展(一)萌芽阶段(16以前世纪)*甲骨文:作物、水分与太阳的关系*战国时期:多粪肥田*西汉:施肥方式*西周:土壤分三等九级*齐民要术:植物对矿物质及水分的要求轮作法、“七九闷麦法”(1)科学植物生理学阶段1.科学植物生理学的开端(17~18世纪)1627年,荷兰 Van Helmont ,水与植物的关系1699年,英国Wood Ward,营养来自土壤和水18世纪,Hales,植物从大气获得营养1771年,英国Priestley发现植物绿色部分可放氧2年,瑞士 De Saussure,灰分与生长的关系2.植物生理学的奠基与成长阶段(19世纪)Ø1840年,德国Liebig建立矿质营养说。
植物生理学

• C)几个重要事件: 几个重要事件:
1)J.Priestley 在1771年发现光合作用; 年发现光合作用; 年发现光合作用 2)J.von Sachs于1882年编写《植物生理学讲 年编写《 于 年编写 义》; 3)W.Pfeffer于1904年编写出版《植物生理学》; 年编写出版《 于 年编写出版 植物生理学》; 4)我国植物生理学奠基人(李继侗、 罗宗洛、汤 我国植物生理学奠基人(李继侗、 罗宗洛、
幼苗生长→细胞分裂和增大 膜和壁的变化 激素及受体、 幼苗生长 细胞分裂和增大→膜和壁的变化 激素及受体、酶等 细胞分裂和增大 膜和壁的变化→激素及受体 (植物体 植物体) 植物体 (细胞 细胞) 细胞 (亚细胞 亚细胞) 亚细胞 (分子 分子) 分子
3 植物生理学与农业生产的关系
农业生产实践孕育了植物生理学,而植 农业生产实践孕育了植物生理学, 物生理学又是农业发展的基础,植物生理 物生理学又是农业发展的基础, 学的每一个重大成果都使得农业技术产生 重大变革。 重大变革。
植物生理学
主讲 李志刚 Email: lizhigangnn@
本课件根据曾广文、蒋德安主编《 本课件根据曾广文、蒋德安主编《植 物生理学》 中国农业科技出版社) 物生理学》(中国农业科技出版社) 制作。 制作。
绪论
教学目标
了解植物生理学的概念、 发展简史及其与农业生产的
关系
1 植物生理学的概念
2 植物生理学的发展简史及趋势
2.1 植物生理学的发展简史
A)古代植物生理学知识
• 《汜胜之书》中记载施肥可分基肥、种肥、追肥。“美 汜胜之书》中记载施肥可分基肥、种肥、追肥。 田之法绿豆为上” “曝使极燥”,“粪水溲种”,“七 田之法绿豆为上” 曝使极燥” 粪水溲种”,“七 九闷麦” 九闷麦”。 • 西欧、古罗马使用人和动物的排泄物、石灰、石膏和灰 西欧、古罗马使用人和动物的排泄物、石灰、 分作肥料。 分作肥料。
八棱海棠种子超低温保存中含水量对糖代谢的影响

㊀Guihaia㊀Dec.2023ꎬ43(12):2290-2299http://www.guihaia-journal.comDOI:10.11931/guihaia.gxzw202211060彭颖ꎬ朱梦婷ꎬ乔谦ꎬ等ꎬ2023.八棱海棠种子超低温保存中含水量对糖代谢的影响[J].广西植物ꎬ43(12):2290-2299.PENGYꎬZHUMTꎬQIAOQꎬetal.ꎬ2023.EffectsofwatercontentonsugarmetabolismofMalusˑrobustaseedcryopreservation[J].Guihaiaꎬ43(12):2290-2299.八棱海棠种子超低温保存中含水量对糖代谢的影响彭㊀颖1ꎬ朱梦婷1ꎬ乔㊀谦1ꎬ李㊀杏1ꎬ张㊀玥1ꎬ皮晓飞2ꎬ刘㊀燕1∗(1.北京林业大学园林学院ꎬ花卉种质创新与分子育种北京市重点实验室ꎬ国家花卉工程技术研究中心ꎬ城乡生态环境北京实验室ꎬ林木花卉遗传育种教育部重点实验室ꎬ北京100083ꎻ2.国家植物园ꎬ北京100093)摘㊀要:含水量是影响种子超低温保存效果的关键因素ꎬ而其作用机制尚不完全清楚ꎮ为探讨含水量对种子超低温保存生活力的影响途径ꎬ该研究以八棱海棠种子为材料ꎬ通过硅胶干燥法获得不同含水量的种子ꎬ测定超低温保存后种子生活力㊁糖含量及相关酶指标的变化并分析相关性ꎮ结果表明:(1)超低温保存15d后ꎬ不同含水量种子生活力不同ꎬ随着种子含水量的降低ꎬ种子生活力呈现先升高后降低的趋势ꎬ含水量为9.02%的八棱海棠种子生活力最高ꎬ为53.33%ꎻ超低温保存120d后ꎬ种子生活力随着含水量下降一直升高ꎬ含水量为6.40%生活力最高ꎬ为27.78%ꎮ这表明八棱海棠种子含水量对超低温保存后的生活力有明显影响ꎬ但受液氮保存时间影响ꎬ随着液氮保存时间的延长ꎬ最适含水量降低ꎮ(2)相关分析显示ꎬ超低温保存后种子含水量与生活力呈极显著负相关(r=-0.82)ꎻ与果糖和蔗糖含量㊁酸性转化酶㊁果糖激酶呈显著负相关ꎬ而种子萌发率与这些指标呈显著正相关ꎮ这表明种子含水量通过影响酸性转化酶活性而影响蔗糖和果糖含量ꎬ进而影响蔗糖代谢ꎬ响应低温和脱水胁迫ꎬ最终导致生活力差异ꎮ种子生活力还受到介导果糖激酶的果糖代谢影响ꎮ此外ꎬ海藻糖也是种子应对脱水和低温胁迫重要的调节物质之一ꎮ综上表明ꎬ八棱海棠种子不同含水量对其超低温保存后生活力有明显影响ꎬ并与蔗糖和果糖代谢差异有关ꎮ该研究为揭示种子超低温保存机制提供了理论参考ꎮ关键词:八棱海棠ꎬ种子超低温保存ꎬ含水量ꎬ糖代谢ꎬ代谢酶中图分类号:Q945㊀㊀文献标识码:A㊀㊀文章编号:1000 ̄3142(2023)12 ̄2290 ̄10EffectsofwatercontentonsugarmetabolismofMalusˑrobustaseedcryopreservationPENGYing1ꎬZHUMengting1ꎬQIAOQian1ꎬLIXing1ꎬZHANGYue1ꎬPIXiaofei2ꎬLIUYan1∗(1.BeijingKeyLaboratoryofOrnamentalPlantsGermplasmInnovation&MolecularBreedingꎬNationalEngineeringResearchCenterforFloricultureꎬBeijingLaboratoryofUrbanandRuralEcologicalEnvironmentꎬKeyLaboratoryofGeneticsandBreedinginForestTreesandOrnamentalPlantsofMinistryofEducationꎬSchoolofLandscapeArchitectureꎬBeijingForestryUniversityꎬBeijing100083ꎬChinaꎻ2.ChinaNationalBotanicalGardenꎬBeijing100093ꎬChina)收稿日期:2023-05-06基金项目:国家自然科学基金(31770741)ꎻ雄安新区科技创新专项(2022XAGG0100)ꎮ第一作者:彭颖(1998-)ꎬ硕士ꎬ主要研究方向为植物种质资源保存ꎬ(E ̄mail)pengying_hallo@163.comꎮ∗通信作者:刘燕ꎬ博士ꎬ教授ꎬ主要研究方向为植物种质资源保存ꎬ(E ̄mail)chblyan@163.comꎮAbstract:Watercontentisakeyfactorinfluencingtheseedcryopreservationꎬbutitseffectingmechanismisnotfullyunderstood.InordertoexploretheeffectandwayofwatercontentontheviabilityofseedcryopreservationonviabilityꎬMalusˑrobustaseedswithdifferentwatercontentswereobtainedbysilicagel.Theseedviabilityꎬsugarcontentandrelatedenzymeindexesbeforeandaftercryopreservationweremeasuredandcorrelationanalysisweredone.Theresultswereasfollows:(1)After15daysofcryopreservationꎬtheviabilityofseedswithdifferentwatercontentswasdifferentꎬandtheviabilityofseedsshowedatrendofincreasingandthendecreasingasthewatercontentofseedsdecreasedꎬandtheseedswith9.02%watercontenthadthehighestgerminationrateꎬat53.33%.Butafter120daysofcryopreservationꎬtheviabilityofseedsincreasedasthewatercontentdecreasedꎬandtheseedswith6.40%watercontentgotthehighestgerminationrateꎬat27.78%.ThisindicatesthatthewatercontentofMalusˑrobustaseedshasasignificanteffectontheviabilityaftercryopreservation.Itisaffectedbythestoragetimeinliquidnitrogenꎬandtheoptimumwatercontentdecreasesasthestoragetimeextendedinliquidnitrogen.(2)Correlationanalysisshowedthattherewasasignificantnegativecorrelationbetweenwatercontentandviability(r=-0.82).Therewassignificantnegativecorrelationwithfructoseandsucrosecontentꎬacidinvertaseꎬfructokinaseꎬwhileseedgerminationratewassignificantlypositivecorrelationwiththeseindexesꎬsuggestingthatwatercontentaffectsthecontentsofsucroseandfructosethrougheffectingtheactivityofacidconvertingenzymesꎬwhichinturnaffectssucrosemetabolismtorespondtolowtemperatureanddehydrationstressꎬultimatelyleadingtodifferencesinviability.Theseedviabilitywasalsoaffectedbyfructosemetabolismthroughfructosekinase.BesidesꎬalginatewasoneoftheimportantregulatorysubstancesofMalusˑrobustaseedinresponsetowaterandlowtemperaturestresses.InsummaryꎬwatercontenthasasignificanteffectontheviabilityofMalusˑrobustaseedscryopreservationthroughthecombinedeffectsofsucroseandfructosemetabolism.Thisstudyprovidesatheoreticalreferencetofurtherexplorethecryopreservationmechanismofseed.Keywords:Malusˑrobustaꎬseedcryopreservationꎬwatercontentꎬsugarmetabolismꎬenzymeofmetabolism㊀㊀八棱海棠(Malusˑrobusta)隶属于蔷薇科(Rosaceae)苹果属(Malus)(陆秋农和贾定贤ꎬ1999)ꎬ是重要的种质资源ꎬ其适应性和抗逆性均较强ꎬ既是优秀的观赏海棠ꎬ也是一种优良的苹果砧木(河北省农林科学院昌黎果树研究所ꎬ1986)ꎬ主要通过播种提供砧木苗ꎬ因此ꎬ其种子资源保存具有重要意义ꎮ超低温保存是种质永久保存最有潜力的技术ꎬ具有环保㊁方便㊁节约成本等诸多优点ꎬ受到广泛关注ꎮ目前ꎬ仅见八棱海棠在低温(-35~5ħ)贮藏的报道ꎬ瓶装密封在0~5ħ冰箱冷藏保存效果较好(何建斌等ꎬ2022)ꎮ因此研究八棱海棠种子的超低温保存ꎬ实现其种质资源的永久保存具有重要的应用价值ꎮ含水量是影响种子生活力的关键因素之一(Chmielarzꎬ2009ꎻWenetal.ꎬ2009)ꎬ大量研究表明ꎬ超低温保存后ꎬ只有在适宜的种子含水量时才能获得较高的萌发率(Generosoetal.ꎬ2019ꎻ张晓宁等ꎬ2020)ꎬ但其作用机制尚不完全清楚ꎬ一般认为含水量变化与细胞低温损伤 两因素假说 相关ꎬ即慢速冷冻会引起细胞逐渐脱水ꎬ发生溶质损伤ꎬ快速冷冻会引起细胞发生冰晶损伤(Mazuretal.ꎬ1972)ꎮ近年来研究也显示ꎬROS介导的氧化应激可能是含水量变化导致种子超低温保存活力降低的损伤机制之一(曹柏ꎬ2016ꎻ李萍ꎬ2018)ꎮ研究表明ꎬ在特定的逆境条件下ꎬ糖在压力感知㊁信号传递和胁迫介导的基因表达中发挥着关键作用ꎬ确保渗透调节反应㊁清除ROSꎬ并通过碳分配维持细胞能量状态(Saddheetal.ꎬ2021)ꎮ其中ꎬ糖含量与相关酶活性是较为敏感的指标ꎬ易受环境因素的影响ꎬ并对胁迫做出反应(方诗雯ꎬ2018)ꎮ不同含水量种子超低温保存中面临低温和脱水双重胁迫ꎬ而含水量是否会通过影响糖代谢进而影响超低温保存后种子的萌发尚未见报道ꎮ本文以八棱海棠种子为材料ꎬ通过研究超低温保存的适宜含水量ꎬ探究不同含水量种子超低温保存效果差异的生理基础ꎬ拟探讨以下问题: (1)含水量是否会影响八棱海棠种子超低温保存后的生活力ꎻ(2)含水量影响超低温保存生活力的作用机制ꎮ本研究为进一步提高种子超低温保存后的生活力提供依据ꎮ192212期彭颖等:八棱海棠种子超低温保存中含水量对糖代谢的影响1㊀材料与方法1.1供试材料八棱海棠种子购于河北省怀来县绿欣八棱海棠苗木基地ꎬ初始含水量为27.12%ꎬ沙藏60d后用于本试验ꎮ1.2方法1.2.1种子不同含水量的调制及测定㊀用硅胶干燥不同时间ꎬ获得不同含水量的种子ꎬ即W1(27.12%)㊁W2(20.32%)㊁W3(15.10%)㊁W4(9.02%)和W5(6.40%)ꎬ含水量随干燥时间的变化具体见图1ꎮ种子含水量用烘箱干燥法测定:取30粒种子ꎬ称重记为Aꎻ将种子放在称量皿中ꎬ称重记为Bꎻ一起放入105ħ烘箱中烘干至恒重ꎬ待冷却后测定种子和称量皿的总重ꎬ记为Cꎮ按照公式计算含水量ꎬ种子含水量=(B-C)/Aˑ100%ꎮ每个处理含30粒种子ꎬ设置3个重复ꎬ取均值ꎮ图1㊀八棱海棠种子含水量随干燥时间的变化Fig.1㊀ChangeofseedwatercontentofMalusˑrobustawithdryingtime1.2.2种子超低温保存及化冻方法㊀以100粒种子为一份ꎬ用锡纸包装后装入网兜ꎬ再投入液氮(liquidnitrogenꎬLN)分别保存15d和120dꎬ试验时快速取出ꎬ用40ħ水浴化冻5minꎬ备用ꎮ1.2.3种子萌发测定方法㊀将沙藏后的种子用清水洗净ꎬ均匀播于放有1层滤纸的一次性培养皿中ꎬ置于光照培养箱中:夜间温度20ħꎬ8hꎻ白天温度25ħꎬ16hꎻ相对湿度在60%以上ꎮ每天定时观察ꎬ记载发芽数ꎮ以胚根长度大于种子长度的1/2时视为发芽(李扬汉ꎬ1984)ꎮ按照下列公式计算生活力ꎮ发芽率(%)=发芽种子数/供试种子数ꎮ发芽势(%)=发芽高峰期萌发的种子数/供试种子数ꎮ发芽指数(GI)=ðGt/Dt(Gt是时间在t日的发芽数ꎬDt是对应的发芽天数)ꎮ每个处理30粒种子ꎬ重复3次ꎬ取均值ꎮ1.2.4糖代谢相关指标测定方法㊀可溶性糖和淀粉含量测定采用蒽酮法(刘萍和李明军ꎬ2016)ꎻ果糖含量测定采用果糖试剂盒(货号GT ̄2 ̄Y)ꎻ蔗糖含量测定采用蔗糖试剂盒(货号ZHT ̄2 ̄Y)ꎻ葡萄糖含量测定采用葡萄糖试剂盒(货号PT ̄2 ̄Y)ꎻ海藻糖含量测定采用海藻糖含量测试盒(货号HT ̄2 ̄Y)ꎻ山梨醇含量测定采用山梨醇含量测试盒(货号SC ̄2 ̄Y)ꎻ直链淀粉含量测定采用直链淀粉试剂盒(货号ZDF ̄2 ̄Y)ꎻ支链淀粉含量测定采用支链淀粉试剂盒(货号ZHDF ̄2 ̄Y)ꎻ蔗糖磷酸合成酶活性测定采用蔗糖磷酸合成酶试剂盒(货号SPS ̄2 ̄Y)ꎻ蔗糖合成酶(合成方向SS ̄Ⅱ)活性测定采用蔗糖合成酶(合成方向SS ̄Ⅱ)试剂盒(货号SSⅡ ̄2 ̄Y)ꎻ中性转化酶活性测定采用中性转化酶试剂盒(货号NI ̄2 ̄Y)ꎻ酸性转化酶活性测定采用酸性转化酶试剂盒(北京索莱宝科技有限公司ꎬ货号BC0560)ꎻ蔗糖合成酶(分解方向SS ̄Ⅰ)活性测定采用蔗糖合成酶(分解方向SS ̄Ⅰ)试剂盒(货号SSⅠ ̄2 ̄Y)ꎻ果糖激酶活性测定采用果糖激酶试剂盒(货号FRK ̄2 ̄Y)ꎮ以上试剂盒未标注来源的均来自苏州科铭生物技术有限公司ꎬ各种物质测定操作步骤按照试剂盒说明书进行ꎮ每处理重复3次ꎬ取均值ꎮ1.3数据处理与分析数据用Excel软件进行整理及图表制作ꎬ采用SPSS26.0软件进行单因素ANOVA分析和Origin进行相关性分析ꎮ2㊀结果与分析2.1不同含水量种子超低温保存后生活力变化种子含水量对超低温保存后种子生活力有明显影响ꎬ见表1ꎮ超低温保存15d后ꎬ随着含水量降低ꎬ生活力呈单峰变化ꎬ即呈现先升高再降低的趋势ꎬ含水量为9.02%(W4)的种子生活力显著高于其他含水量种子ꎬ发芽率为53.33%ꎬ发芽势为2922广㊀西㊀植㊀物43卷表1㊀八棱海棠种子含水量超低温保存后生活力的变化Table1㊀ChangesofMalusˑrobustaseedviabilityaftercryopreservation含水量Watercontent(%)发芽率Germinationrate(%)LN保存15d后KeepLNfor15dLN保存120d后KeepLNfor120d发芽势Germinationpotential(%)保存15d后KeepLNfor15dLN保存120d后KeepLNfor120d发芽指数Germinationindex保存15d后KeepLNfor15dLN保存120d后KeepLNfor120d27.12(W1)0.00ʃ0.00Ca0.00ʃ0.00Ba0.00ʃ0.00Ba0.00ʃ0.00Ba0.00ʃ0.00Ca0.00ʃ0.00Ba20.32(W2)6.67ʃ6.67Ca5.56ʃ1.92Ba5.56ʃ5.09Ba4.44ʃ1.92Ba3.46ʃ3.17Ca3.24ʃ1.17Ba15.10(W3)32.22ʃ5.09Ba23.33ʃ6.67Aa11.94ʃ8.35Ba17.78ʃ1.92Aa11.53ʃ4.47Ba12.22ʃ4.23Aa9.02(W4)53.33ʃ8.82Aa25.56ʃ6.94Ab33.33ʃ8.82Aa20.00ʃ5.77Aa20.52ʃ5.33Aa12.00ʃ3.01Aa6.40(W5)33.33ʃ3.33Ba27.78ʃ10.18Aa27.78ʃ5.09Aa23.33ʃ11.55Aa15.55ʃ0.99ABa13.27ʃ5.04Aa㊀注:不同大写字母表示相同处理组不同含水量种子差异显著(P<0.05)ꎻ不同小写字母表示相同含水量种子超低温保存15d和120d后差异显著(P<0.05)ꎮ㊀Note:Differentcapitallettersindicatesignificantdifferencesinseedswithdifferentwatercontentsinthesametreatmentgroup(P<0.05)ꎻdifferentlowercaselettersindicatethatseedswiththesamewatercontentaresignificantlydifferentafter15daysand120daysofcryopreservation(P<0.05).33.33%ꎬ发芽指数为20.52ꎻ自然含水量27.12%(W1)的种子保存后丧失生活力ꎻ较高含水量20.32%(W2)的种子发芽率仅为6.67%ꎬ显著低于含水量15.10%(W3)的32.22%和6.40%(W5)的33.33%ꎮ超低温保存120d后ꎬ各含水量种子的生活力均下降ꎬ保存15d时生活力最高的W4种子生活力下降最大ꎬ发芽率由53.33%降至25.56%ꎬ已与W3和W5种子无显著差异ꎮ随着含水量的降低ꎬ种子生活力一直升高ꎬ含水量为6.40%(W5)的种子生活力最高ꎬ发芽率为27.78%ꎬ发芽势为23.33%ꎬ发芽指数为13.27ꎮ2.2种子含水量在超低温保存15d后糖含量的变化液氮保存15d后ꎬ不同含水量八棱海棠种子超低温保存后可溶性糖总量的变化见图2:aꎮ具有最高生活力的W4种子(含水量9.02%)ꎬ其可溶性糖总量显著低于其他各组ꎬ但种子含水量降至6.40%(W5)时ꎬ可溶性糖含量升高且与其他高含水量种子没有显著差异ꎮ进一步测定不同含水量八棱海棠种子超低温保存后葡萄糖含量的变化ꎬ结果见图2:bꎮ与可溶糖含量变化趋势不同ꎬ自然含水量种子(W1)的葡萄糖含量显著低于其他组ꎬ生活力较高的W4种子含量较低ꎬ但与其余含水量种子之间差异不显著ꎮ不同含水量八棱海棠种子超低温保存后蔗糖含量的变化不大ꎬ结果见图3ꎮ仅液氮保存后丧失生活力的自然含水量W1种子蔗糖含量显著低于其他含水量ꎬ而其他含水量种子之间差异不显著ꎻ从变化趋势看ꎬ低含水量的W3~W5种子蔗糖含量要高于较高含水量的W1和W2种子ꎮ超低温保存后ꎬ随着种子含水量降低ꎬ其果糖含量升高ꎬ高含水量W1和W2种子与较低含水量W3~W5种子差异显著ꎻ虽然含水量W4的种子果糖含量最高ꎬ但与W3和W5差异不显著ꎮ不同含水量之间的海藻糖和山梨醇含量变化趋势一致ꎬ高含水量W1和W2种子的含量显著低于较低含水量ꎬ随着含水量降低ꎬ均呈升高趋势ꎬW5种子海藻糖和山梨醇含量最高ꎬ与W3和W4种子的海藻糖含量差异显著ꎬ而山梨醇含量差异不显著ꎮ不同含水量八棱海棠种子超低温保存后淀粉含量的变化见图4ꎮ八棱海棠种子中支链淀粉含量明显高于直链淀粉含量ꎬ约为其5倍ꎮ随着含水量的降低ꎬ淀粉含量和直链淀粉含量均呈先升高再降低的趋势ꎮ液氮冻后丧失生活力的W1种子的直链淀粉含量显著低于其他含水量ꎬ各含水量间的支链淀粉含量未见明显差异ꎮ2.3种子超低温保存后生活力、含水量与糖含量指标的相关性分析超低温保存15d后ꎬ生活力㊁含水量与糖含量指标的相关性分析结果见图5ꎮ含水量与生活力呈极显著负相关关系ꎬ相关系数为0.82ꎬ还与蔗糖㊁果糖㊁葡萄糖㊁海藻糖和直链淀粉呈显著负相关ꎮ生392212期彭颖等:八棱海棠种子超低温保存中含水量对糖代谢的影响不同大写字母表示相同处理组不同含水量种子差异显著(P<0.05)ꎬ下同ꎮDifferentcapitallettersindicatesignificantdifferencesinseedswithdifferentwatercontentsinthesametreatmentgroup(P<0.05)ꎬthesamebelow.图2㊀不同含水量八棱海棠种子超低温保存15d后可溶性糖总糖和葡萄糖含量的变化Fig.2㊀ChangesofsolublesugarandglucosecontentsindifferentwatercontentsofMalusˑrobustaseedsafter15daysofcryopreservation图3㊀不同含水量八棱海棠种子超低温保存15d后蔗糖㊁果糖㊁海藻糖和山梨醇含量的变化Fig.3㊀ChangesofsucroseꎬfructoseꎬtrehaloseandsorbitolcontentsindifferentwatercontentsofMalusˑrobustaseedsafter15daysofcryopreservation活力与蔗糖呈显著正相关ꎬ相关系数为0.59ꎬ与果糖和海藻糖呈极显著正相关ꎬ相关系数分别为0.83和0.72ꎬ蔗糖与果糖和海藻糖也呈显著正相关ꎮ15d液氮保存表明ꎬ种子含水量可以影响蔗糖和果糖代谢ꎬ追踪测定液氮保存120d后不同含水量种子的生活力(表1)㊁蔗糖和果糖含量变化及相关代谢酶活性关系ꎮ2.4种子含水量对超低温保存120d后糖代谢相关指标的影响2.4.1八棱海棠不同含水量种子超低温保存120d后蔗糖和果糖含量变化㊀各个含水量的种子在液氮保存120d后的蔗糖含量均显著低于保存15d图4㊀不同含水量八棱海棠种子超低温保存15d后淀粉含量的变化Fig.4㊀ChangesofstarchcontentindifferentwatercontentsofMalusˑrobustaseedsafter15daysofcryopreservation后的蔗糖含量ꎬ结果见图6ꎮ低含水量(W3~W5)种子的蔗糖和果糖含量也高于高含水量(W1~W2)种子ꎬ并且保存120d后生活力最高的W5种子的蔗糖和果糖含量也显著高于其他含水量ꎮ2.4.2超低温保存120d后蔗糖代谢酶活性变化㊀八棱海棠不同含水量种子超低温保存120d后蔗糖合成相关酶的变化见图7:aꎬ蔗糖磷酸合成酶(sucrosephosphatesynthetaseꎬSPS)和蔗糖合成酶(合成方向)[sucrosesynthetase(synthesis)ꎬSS ̄Ⅱ]活性变化均没有一定规律性ꎮW2种子SPS活性显著高ꎬ而W3显著低ꎬ高含水量的W1种子与低含水量的W4和W5没有显著差异ꎻ高含水量的4922广㊀西㊀植㊀物43卷∗表示显著相关(P<0.05)ꎻ∗∗表示极显著相关(P<0.01)ꎮSS.可溶性糖ꎻSuc.蔗糖ꎻGlc.葡萄糖ꎻFru.果糖ꎻTre.海藻糖ꎻSbt.山梨醇ꎻAum.淀粉ꎻAse.直链淀粉ꎻAin.支链淀粉ꎮ∗indicatessignificantcorrelations(P<0.05)ꎻ∗∗indicatesextremelysignificantcorrelations(P<0.01).SS.SolublesugarꎻSuc.SucrosꎻGlc.GlucoseꎻFru.FructoseꎻTre.TrehaloseꎻSbt.SorbitolꎻAum.AmylumꎻAse.AmyloseꎻAin.Amylopectin.图5㊀八棱海棠种子超低温保存15d后生活力、含水量与糖含量的相关性Fig.5㊀CorrelationbetweenviabilityꎬwatercontentandsugarcontentofMalusˑrobustaseedsafter15daysofcryopreservation图6㊀八棱海棠种子不同含水量超低温保存120d后蔗糖和果糖含量的变化Fig.6㊀ChangesofsucroseandfructosecontentsindifferentwatercontentsofMalusˑrobustaseedsafter120daysofcryopreservation自然含水量W1种子的SS ̄Ⅱ活性最高ꎬ但与低含水量组W3和W5组无显著差异ꎮ八棱海棠不同含水量种子超低温保存120d后蔗糖分解相关酶的变化见图7:bꎮ3种酶活性均没有随种子含水量变化呈规律性变化ꎮ超低温保存120d后ꎬ各含水量种子的蔗糖中性转化酶(neutralinvertaseꎬNI)和蔗糖合成酶(分解方向)[sucrosesynthetase(decomposition)ꎬSS ̄Ⅰ]活性差异不明显ꎮ生活力最高的W5种子的酸性转化酶(acidinvertaseꎬAI)显著高于其他含水量种子ꎬ生活力随液氮保存时间下降最明显的W4种子AI活性最低ꎬ但W1~W4之间没有显著差异ꎮ八棱海棠不同含水量种子超低温保存120d后ꎬ蔗糖分解类酶活性高于蔗糖合成类酶活性ꎬ说明在八棱海棠中主要以蔗糖分解为主要代谢方向ꎮ2.4.3超低温保存120d后果糖激酶活性变化㊀不同含水量种子果糖激酶(fructokinaseꎬFRK)活性变化结果见8ꎮ液氮保存120d后ꎬFRK活性变化呈单峰变化ꎬ低含水量W3~W5种子的FRK活性显著高于高含水量W1~W2种子ꎬ生活力下降最大的W4种子FRK活性值最大ꎮ2.4.4八棱海棠种子超低温保存120d后生活力、含水量与糖含量和相关酶指标的相关性分析㊀超低温保存120d后ꎬ种子含水量㊁生活力和糖及其酶相关性见图9ꎮ相关分析显示与液氮保存15d相同ꎬ含水量与生活力呈极显著负相关ꎬ相关系数为0.85ꎬ含水量还与蔗糖㊁果糖含量呈负相关ꎬ生活力与蔗糖和果糖含量呈显著正相关ꎬ与AI活性和FRK活性也呈显著正相关ꎬ说明含水量通过影响蔗糖分解成果糖影响了种子生活力ꎬ酸性转化酶在其中发挥了重要作用ꎬ同时果糖代谢也影响了种子的生活力ꎮ3㊀讨论八棱海棠种子可以实现超低温保存ꎬ并且含水量对超低温保存效果有重要影响ꎮ本研究中八棱海棠种子从自然含水量27.12%逐渐降为9.02%时ꎬ在液氮保存15d后ꎬ种子发芽率呈逐渐升高趋势ꎬ此结果与马尾松(Pinusmassoniana)(张晓宁等ꎬ2020)㊁多花芍药(Paeoniaemodi)(周好ꎬ2021)和鸡蛋果(Passifloraedulis)(Generosoetal.ꎬ2019)等种子一样ꎬ含水量需要降到一定的水平(分别为6.1%㊁8.87%和10%)才能在液氮中保存依然有较高生活力ꎬ可能是由于生物组织含水量降低时会形成玻璃态ꎬ而含水量较高的植物进行592212期彭颖等:八棱海棠种子超低温保存中含水量对糖代谢的影响图7㊀八棱海棠种子不同含水量超低温保存120d后蔗糖代谢酶活性变化Fig.7㊀ChangesofsucrosemetabolismenzymeactivityindifferentwatercontentsofMalusˑrobustaseedsafter120daysofcryopreservation图8㊀八棱海棠种子不同含水量超低温保存120d后果糖激酶(FRK)活性变化Fig.8㊀Changesoffructokinase(FRK)activityindifferentwatercontentsofMalusˑrobustaseedsafter120daysofcryopreservation超低温保存时ꎬ通常易在细胞内形成冰晶ꎬ破坏细胞结构ꎬ从而导致细胞死亡ꎮ八棱海棠种子在自然含水量状态下ꎬ液氮冻存15d和120d后生活力均丧失ꎬ本研究还发现随着液氮冻存时间的延长ꎬ最适含水量也会随之降低ꎮ近年来研究发现ꎬ参与糖代谢过程中的蔗糖㊁葡萄糖和果糖具有信号分子的功能ꎬ在植物遇到逆境的情况下ꎬ这些可溶性糖类直接或间接对植物进行调控ꎬ对逆境的胁迫做出应激反应(Windetal.ꎬ2010)ꎮ尽管有些植物在低温胁迫下糖含量会增加ꎬ但不同植物可能会积累不同的糖类物质ꎮ例如ꎬ耐低温的番茄(Solanumlycopersicum)品种在低温(4ħ)胁迫时会积累果糖和葡萄糖(齐红岩等ꎬ2012)ꎻ稻(Oryzasativa)在低温(平均温度4.6∗表示显著相关(P<0.05)ꎻ∗∗表示极显著相关(P<0.01)ꎮSuc.蔗糖ꎻFru.果糖ꎻSPS.蔗糖磷酸合成酶ꎻSS ̄Ⅱ.蔗糖合成酶(合成)ꎻNI.中性转化酶ꎻAI.酸性转化酶ꎻSS ̄Ⅰ.蔗糖合成酶(分解)ꎻFRK.果糖激酶ꎮ∗indicatessignificantcorrelations(P<0.05)ꎻ∗∗indicatesextremelysignificantcorrelations(P<0.01).Suc.SucroseꎻFru.FructoseꎻSPS.SucrosephosphatesynthetaseꎻSS ̄Ⅱ.Sucrosesynthetase(synthesis)ꎻNI.NeutralinvertaseꎻAI.AcidinvertaseꎻSS ̄Ⅰ.Sucrosesynthetase(decomposition)ꎻFRK.Fructokinase.图9㊀八棱海棠种子超低温保存120d后生活力㊁含水量与糖含量和相关酶的相关性Fig.9㊀CorrelationbetweenviabilityꎬwatercontentandsugarcontentandrelatedenzymesofMalusˑrobustaseedsafter120daysofcryopreservationħ)下积累可溶性总糖㊁还原性糖和蔗糖(Turhan&Erginꎬ2012)ꎮ在本研究中ꎬ液氮保存15d后ꎬ低含水量(W3~W5)种子的蔗糖和果糖含量均高于高含水量(W1~W2)种子ꎬ但此时活力最高的W4种子的可溶性糖和葡萄糖含量相比W3和W5种6922广㊀西㊀植㊀物43卷子较低ꎮ蔗糖和果糖与含水量和生活力均相关ꎬ推测含水量可能通过影响可溶性糖中的蔗糖和果糖含量而影响生活力ꎬ适宜的含水量有利于八棱海棠种子蔗糖和果糖含量的积累ꎬ增加抗低温能力ꎬ需要进一步验证ꎮWang等(2000)研究表明ꎬ叶片中蔗糖合成酶和转化酶活性的增强可加快蔗糖的快速循环ꎬ有利于蔗糖的积累ꎬ以抵抗干旱胁迫的影响ꎮSPS催化蔗糖合成ꎬSS活性与蔗糖的分解与合成有关ꎬ但目前不同文献对这两种参与蔗糖代谢的相关酶在胁迫下的变化规律并不完全一致ꎮ例如ꎬ苗期夜间低温处理会导致番茄叶片的SS和SPS活性受到抑制(Wang&Liꎬ2011ꎻ刘玉凤等ꎬ2011)ꎬ小麦(Triticumaestivum)受低温胁迫后ꎬ不同器官SS和SPS基因的表达变化不同ꎬ分蘖中SS基因表达逐渐降低ꎬ叶片中SS基因表达先升高后降低ꎬ而SPS在叶片和分蘖中均表现为先升高再降低(Zengetal.ꎬ2011)ꎮ蔗糖转化酶则参与蔗糖的分解ꎬ在植物体内ꎬ果糖可以通过蔗糖合成酶(分解方向)和蔗糖转化酶的作用下分解蔗糖而产生ꎬ本研究中发现果糖含量与蔗糖含量呈显著正相关ꎬ同时蔗糖和果糖与AI活性也具有显著相关性ꎬ说明八棱海棠种子超低温保存过程中蔗糖主要通过AI降解成分子量更小的果糖ꎬ供机体吸收ꎬ提高渗透调节能力以响应水分和低温胁迫ꎮ这与孙汉青(2018)的研究结果一致ꎬ即干旱条件可以提高苹果(Maluspumila)果实中AI活性ꎬ导致果实中蔗糖含量下降ꎬ果糖㊁葡萄糖含量上升ꎬ缓解了干旱对植物的影响ꎬ但本研究中SPS和SS两种酶的活性与含水量以及生活力之间的相关性不显著ꎬ仅SPS与蔗糖相关ꎬ可能是糖含量与糖代谢酶活性之间有较为复杂的关系ꎬ错综复杂地调控着糖代谢ꎬ有待更多的研究数据解开诸多困惑ꎮ果糖磷酸化是果糖进入代谢途径的第一道生化反应ꎬ其中起作用的高效酶是FRKꎬ在调节细胞中的果糖浓度及响应环境胁迫中发挥着关键的作用(叶香媛和周文彬ꎬ2021)ꎬ在盐胁迫和干旱胁迫下ꎬFRK基因分别在玉蜀黍(Zeamays)(Zörbetal.ꎬ2010)和向日葵(Helianthusannuus)幼苗叶片(Fuldaetal.ꎬ2011)中表达量上调ꎬ此外ꎬFRK可能还在缺氧条件下对水稻幼苗碳水化合物代谢发挥重要作用(Guglielminettietal.ꎬ2006)ꎬ本研究也表明果糖激酶与超低温保存后生活力显著正相关ꎬ暗示果糖激酶在低温胁迫中发挥了一定作用ꎮ海藻糖是一种双糖ꎬ可作为逆境胁迫下的信号和保护性物质(Suprasannaetal.ꎬ2016)ꎬ保持膜的完整性ꎬ清除活性氧(Zulfiqaretal.ꎬ2019)ꎮ孙汉青(2018)研究发现海藻糖参与了苹果果实中糖代谢的调节ꎬ维持了干旱条件下植物正常生长发育ꎬ缓解干旱对植物的影响ꎬ外源添加海藻糖还可以缓解胁迫对植物的影响ꎮ本研究发现ꎬ低含水量种子的海藻糖含量要高于高含水量种子ꎬ并且海藻糖与含水量和生活力均呈显著相关ꎬ说明海藻糖是八棱海棠种子应对水分和低温胁迫重要的调节物质之一ꎬ其具体影响方式还有待进一步研究ꎮ山梨醇是木本蔷薇科植物中主要的光合产物㊁同化物运输形式和可溶性的贮藏性碳水化合物主要的兼容溶质与抗氧化剂ꎬ作为小分子细胞相容物质ꎬ可以抵御生物和非生物胁迫(Noiraudetal.ꎬ2001)ꎮ在短期干旱胁迫下ꎬ山梨醇含量随干旱程度的加大而明显增加ꎬ其合成关键酶活性和基因表达也显著增加(张钊ꎬ2016)ꎬ本研究中也发现随含水量的降低ꎬ八棱海棠种子山梨醇含量有所上升ꎮBensari等(1990)曾提出水分胁迫下淀粉含量的变化是植物抵御胁迫的内在因素的假说ꎬ一般认为淀粉降解㊁可溶性糖含量升高有利于抵抗外界环境胁迫ꎮ4ħ低温下ꎬ甘薯(Dioscoreaesculenta)中淀粉合成关键基因表达显著降低ꎬ而水解酶编码基因表达量显著升高(崔鹏等ꎬ2022)ꎻ而林敏卓(2012)研究表明ꎬ低温胁迫时南极冰藻(Chlamydomonassp.)淀粉合成酶的基因表达明显上调ꎮ本研究中随着含水量降低ꎬ淀粉总量有先上升再下降的趋势ꎬ但淀粉含量与可溶性糖含量的变化相关程度不高并且不与含水量和生活力显著相关ꎮ可能是因为可溶性糖和淀粉含量的变化是一个动态过程ꎬ未在此阶段显示出明显的对应关系ꎮ由于材料来源所限ꎬ本研究获得的八棱海棠种子是已经过沙藏60d的种子ꎮ从超低温保存技术原理分析ꎬ采用未经过沙藏的处于深休眠期的种子液氮保存后生活力应该更高ꎬ后续可以进一步试验确定适宜采收期和超低温保存的适宜含水量ꎬ深休眠种子液氮贮存后对休眠解除的影响等问题也值得进一步研究ꎮ792212期彭颖等:八棱海棠种子超低温保存中含水量对糖代谢的影响4㊀结论八棱海棠种子含水量对超低温保存后生活力有明显影响ꎬ但也受液氮保存时间影响ꎬ随着液氮保存时间的延长ꎬ最适含水量降低ꎮ种子含水量通过影响酸性转化酶活性ꎬ将蔗糖降解为分子量更小的果糖ꎬ以响应脱水和低温胁迫ꎬ从而影响种子生活力ꎻ种子生活力还受到通过果糖激酶的果糖代谢影响ꎮ此外ꎬ海藻糖也是八棱海棠种子应对脱水和低温胁迫重要的调节物质之一ꎮ参考文献:BENSARIMꎬCALMESJꎬVIALAG.1990.Waterdeficitanddistributionofphotosyntheticcarbonbetweensucroseandstarch[J].ActaOecolꎬ11(6):843-855.CAOBꎬ2016.EffectsofcryopreservationonphysiologicalcharacteristicsofIrisensataseedsandestablishmentofregenerationsystem[D].Harbin:NortheastForestryUniersity.[曹柏ꎬ2016.超低温保存对玉蝉花种子生理特性影响及再生体系建立[D].哈尔滨:东北林业大学.]ChangliFruitResearchInstituteꎬHebeiAcademyofAgricultureandForestrySciencesꎬ1986.Hebeifruitannalsvol.3:hebeiappleannals[M].Beijing:ChinaAgriculturePress.[河北省农林科学院昌黎果树研究所ꎬ1986.河北省果树志第三卷:河北省苹果志[M].北京:中国农业出版社.]CHMIELARZPꎬ2009.Cryopreservationofdormanteuropeanash(Fraxinusexcelsior)orthodoxseeds[J].TreePhysiolꎬ29(10):1279-1285.CUIPꎬZHAOYRꎬYAOZPꎬetal.ꎬ2022.Starchphysicochemicalpropertiesandexpressionlevelsofanabolismkeygenesinsweetpotatounderlowtemperature[J].ChinAgricSciꎬ55(19):3831-3840.[崔鹏ꎬ赵逸人ꎬ姚志鹏ꎬ等ꎬ2022.低温对甘薯淀粉理化特性及代谢关键基因表达量的影响[J].中国农业科学ꎬ55(19):3831-3840.]FANGSWꎬ2018.StudiesonthemechanismofcarbohydratemetabolisminseedsofGinkgobilobal[D].Shenyang:ShenyangAgriculturalUniversity.[方诗雯ꎬ2018.银杏种子后熟过程中碳水化合物代谢机制研究[D].沈阳:沈阳农业大学.]FULDASꎬMIKKATSꎬSTEGMANNHꎬetal.ꎬ2011.Physiologyandproteomicsofdroughtstressacclimationinsunflower(HelianthusannuusL.)[J].PlantBiol(Stuttg)ꎬ13(4):632-642.GENEROSOALꎬCARVALHOVSꎬWALTERRꎬetal.ꎬ2019.Mature ̄embryocultureinthecryopreservationofpassionfruit(PassifloraedulisSims)seeds[J].SciHorticꎬ256:108638.GUGLIELMINETTILꎬMORITAAꎬYAMAGUCHIJꎬetal.ꎬ2006.DifferentialexpressionoftwofructokinasesinOryzasativaseedlingsgrownunderaerobicandanaerobicconditions[J].JPlantResꎬ119(4):351-356.HEJBꎬLIXXꎬYANGYHꎬ2022.EffectsofseedstorageconditionsongerminationofMalusˑrobustaseeds[J].HebeiForSciTechnolꎬ(1):46-47.[何建斌ꎬ李晓霞ꎬ杨沿辉ꎬ2022.八棱海棠种子储存条件对发芽的影响[J].河北林业科技ꎬ(1):46-47.]LIPꎬ2018.StudyoncryopreservationofthreegermplasmresourcesofLonicerajaponica[D].Harbin:NortheastForestryUniversity.[李萍ꎬ2018.金银忍冬3种种质资源超低温保存的研究[D].哈尔滨:东北林业大学.]LIYHꎬ1984.Botany[M].Shanghai:ShanghaiScienceandTechnologyPress.[李扬汉ꎬ1984.植物学[M].上海:上海科学技术出版社.]LINMZꎬ2012.Molecularcloningandfunctionalanalysisofanti ̄freezegenesfromChlamydomonassp.ice ̄l[D].Zibo:ShandongInstituteofLightIndustry.[林敏卓ꎬ2012.南极冰藻Chlamydomonassp.ice ̄l低温胁迫相关基因的克隆和功能验证[D].淄博:山东轻工业学院.]LIUPꎬLIMJꎬ2016.Plantphysiologyexperiments[M].2nded.Beijing:SciencePress.[刘萍ꎬ李明军ꎬ2016.植物生理学实验[M].2版.北京:科学出版社.]LIUYFꎬLITLꎬJIAOXHꎬ2011.Effectsofshort ̄termsub ̄lownighttemperaturetreatmentandrecoveryonphotosynthesisandsucrose ̄metabolismoftomatoleaves[J].JHorticꎬ38(4):683-691.[刘玉凤ꎬ李天来ꎬ焦晓赤ꎬ2011.短期夜间亚低温及恢复对番茄光合作用和蔗糖代谢的影响[J].园艺学报ꎬ38(4):683-691.]LUQNꎬJIADXꎬ1999.Chinesefruittreezhiappleroll[M].Beijing:ChinaForestryPublishingHouse.[陆秋农ꎬ贾定贤ꎬ1999.中国果树志苹果卷[M].北京:中国林业出版社.]MAZURPꎬLEIBOSPꎬCHUEHꎬ1972.Atwo ̄factorhypothesisoffreezinginjury.Evidencefromchinesehamstertissue ̄culturecells[J].ExpCellResꎬ71(2):345-355.NOIRAUDNꎬMAUROUSSETLꎬLEMOINERꎬ2001.IdentificationofamannitoltransporterꎬAgMaT1ꎬinceleryphloem[J].PlantCellꎬ13(3):695-705.QIHYꎬJIANGYYꎬHUALJꎬ2012.Responsestoshort ̄termlownighttemperatureofsucrose ̄metabolizingindomesticatedtomatoandwildrelative[J].JHorticꎬ39(2):281-288.[齐红岩ꎬ姜岩岩ꎬ华利静ꎬ2012.短期夜间低温对栽培番茄和野生番茄果实蔗糖代谢的影响[J].园艺学报ꎬ39(2):281-288.]SADDHEAAꎬMANUKARꎬPENNASꎬ2021.Plantsugars:8922广㊀西㊀植㊀物43卷。