常见真核启动子及原核启动子特点精编版
常见真核启动子及原核启动子特点

常见真核启动子及原核启动子特点启动子是基因的调控序列,用于启动基因的转录过程。
真核核基因的启动子与原核启动子在结构和调控机制上有一些显著的差异。
常见的真核启动子包括TATA-box启动子、CAAT-box启动子、GC-box 启动子以及增强子和启动子相关顺反元件等。
相比之下,原核启动子主要包括启动子结构(包括RBS和启动子序列)和阻止子。
TATA-box启动子是真核启动子中最为常见的一类,其基本序列为TATAAAA。
TATA-box启动子位于转录起始点的上游区域,与转录因子TBP (TATA结合蛋白)结合,通过改变染色质的结构来帮助RNA聚合酶Ⅱ结合,起到促进基因转录的作用。
CAAT-box启动子基本序列为CCAAT,位于转录起始点的上游区域。
CAAT-box启动子常与转录因子CBF(CCAAT结合蛋白)结合,CBF与RNA 聚合酶Ⅱ发生相互作用,促进基因转录。
GC-box启动子的基本序列为GCGGGG,位于转录起始点的上游区域。
与TATA-box启动子和CAAT-box启动子不同,GC-box启动子常与转录因子Sp1结合,Sp1使染色质结构松弛,帮助RNA聚合酶Ⅱ结合并促进基因转录。
除了这些常见的启动子序列外,增强子也是一类重要的真核启动子,常用于增强基因的转录活性。
在基因转录过程中,转录因子能够结合到增强子上,进而与启动复合物相互作用,使基因转录增强。
和真核启动子不同,原核启动子主要包括启动子结构和阻止子。
启动子结构是几个具有一定保守序列的DNA序列是细菌RNA聚合酶与启动DNA 的结合位点。
与真核启动子相比,原核启动子相对简单,其基本序列也多样,通常包括TATAAT盒子和Pribnow盒子。
与此同时,原核启动子还包括RBS(ribosome binding site)结构,用于Ribosome的结合位置,进而参与翻译过程。
而阻止子是一种转录起始点之后的DNA序列,能够阻止RNA聚合酶进一步延伸合成RNA链。
DNA启动子概述

启动子概述启动子是DNA链上一段能与RNA聚合酶结合并能起始mRNA合成的序列,它是基因表达不可缺少的重要调控序列。
启动子是一段位于结构基因5’-端上游区的DNA序列,能活化RNA聚合酶,使之与模板DNA准确地结合,并具有转录起始的特异性。
基因的特异性转录取决于酶与启动子能否有效地形成二元复合物。
启动子分三类:启动子Ⅰ、启动子Ⅱ、启动子Ⅲ.只有启动子Ⅱ指导mRNA的转录。
真核生物启动子Ⅱ由两大部分组成:上游元件(upstream element)和启动子核心(core promoter)。
上游元件与转录的效率有关;启动子核心包括3部分:TATA盒、起始子(initinator)及下游元件(downstream element)。
TATA盒为转录调控因子包括各种调节蛋白的结合区,与转录起始位点的精确选择及转录有关,起始子是转录起始所必须,下游元件作用尚不清楚。
原核生物启动子区范围较小,包括TATAAT区(Pribnow区)及其上游的TTGACA区。
启动子是一段提供RNA聚合酶识别和结合位点的DNA序列,位于基因上游。
启动子具有如下特征:1序列特异性。
在启动子的DNA序列中,通常含有几个保守的序列框,序列框中碱基的变化会导致转录启动活性的改变。
2方向性。
启动子是一种有方向性的顺式调控元件,有单向启动子和双向启动子两类。
3位置特性。
启动子一般位于所启动转录基因的上游或基因内的前端。
处于基因的下4种属特异性。
原核生物的不同种、属,真核生物的不同组织都具有不同类型的启动没有启动子,基因就不能转录。
原核生物启动子是由两段彼此分开且又高度保守的核苷酸序列组成,对mRNA的合成极为重要。
启动子区域:(1)Pribnow盒,位于转录起始位点上游5—10bp,一般由6~8个碱基组成,富含A和T,故又称为TATA盒或—10区。
启动子来源不同,Pribnow盒的碱基顺序稍有变化。
(2)—35区,位于转录起始位点上游35bp处,故称—35区,一般由10个碱基组成。
原核真核启动子及BL21相关知识

感受态BL21、BL21(DE3)及BL21(DE3)pLysSBL21没有T7 RNA聚合酶基因, 因此不能表达T7启动子控制的目的基因, 但是可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)是λDE3溶原菌,带有T7 RNA聚合酶,可用于表达T7启动子控制的目的基因,也可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)pLysS含有pLysS质粒,一种与pET共存的表达T7裂解酶的质粒,可以减少目的基因的本底表达,提供更严紧的控制。
真核原核启动子原核:几个常用的启动子和诱导调控表达系统1.最早应用于的表达系统的是Lac乳糖操纵子,由启动子lacP + 操纵基因lacO + 结构基因组成。
其转录受CAP正调控和lacI负调控。
cUV5突变能够在没有CAP的存在下更有效地起始转录,该启动子在转录水平上只受lacI 的调控,因而随后得到了更广泛采用。
lacI产物是一种阻遏蛋白,能结合在操纵基因lacO 上从而阻遏转录起始。
乳糖的类似物IPTG可以和lacI产物结合,使其构象改变离开lacO,从而激活转录。
这种可诱导的转录调控成为了大肠杆菌表达系统载体构建的常用元件。
3.tac启动子是trp启动子和lacUV5的拼接杂合启动子,且转录水平更高,比lacUV5更优越。
4.trc启动子是trp启动子和lac启动子的拼合启动子,同样具有比trp更高的转录效率和受lacI阻遏蛋白调控的强启动子特性。
5.在常规的大肠杆菌中,lacI阻遏蛋白表达量不高,仅能满足细胞自身的lac操纵子,无法应付多拷贝的质粒的需求,导致非诱导条件下较高地表达,为了让表达系统严谨调控产物表达,能过量表达lacI阻遏蛋白的lacIq 突变菌株常被选为Lac/Tac/trc表达系统的表达菌株。
现在的Lac/Tac/trc载体上通常还带有lacIq 基因,以表达更多lacI阻遏蛋白实现严谨的诱导调控。
常见真核启动子及原核启动子特点
真核启动子EF1a常规表达用mRNA人延长因子1α来源的强哺乳动物表达启动子组成型表达水平十分稳定,与细胞类型无关PGK1 (人/小鼠)常规表达用mRNA磷酸甘油酸酯激酶基因来源的哺乳动物启动子组成型广泛表达,但可能因细胞类型而异。
由于甲基化或脱乙酰作用,倾向于抵抗启动子下调。
human beta actin常规表达用mRNAβ-肌动蛋白基因来源的哺乳动物启动子组成型无处不在,鸡的启动子常用与启动子杂交TRE常规表达用mRNA四环素响应元件启动子被四环素或者类似物诱导通常有本底表达Ac5常规表达用mRNA果蝇Actin 5c 基因来源的强昆虫启动子组成型果蝇表达系统的常用启动子CaMKIIa 光遗传学基因表达mRNACa2+/钙调蛋白依赖的蛋白激酶 II 启动子特异的用于中枢神经系统/神经元表达。
受到钙和钙调蛋白调节。
TEF1常规表达用mRNA酵母转录延伸因子启动子组成型与哺乳动物的EF1a 启动子类似ADH1常规表达用mRNA乙醇脱氢酶I的酵母启动子被乙醇抑制全长版本很强,促进高表达。
截短启动子是组成型的,表达较低。
Ubi常规表达用mRNA玉米泛素基因的植物启动子组成型在植物中促进高表达U6小RNA表达shRNA来源于人U6小核启动子组成型小鼠U6也使用,但效率略差。
常用的原核表达系统启动子T7lac高水平基因表达T7噬菌体来源的启动子加上lac 操纵子 几乎没有本底表达,需要T7 RNA 聚合酶,受到lac 操纵子的控制,可以被IPTG 诱导。
常用与pET 载体,受到lac 操纵子的严格调控Sp6体外转录/常规表达Sp6噬菌体来源的启动子 组成型, 需要SP6 RNA 聚合酶当用于体外转录的时候,专路方向有可能是正向的也可能是反向的,取决于启动子相对于目的基因的方向araBAD常规表达用阿拉伯糖代谢操纵子的启动子阿拉伯糖诱导 弱,常用与pBAD 载体。
适合于快速调控和低的本底表达lac 常规表达用Lac操纵子来源的启动子可以被IPTG或者乳糖诱导在常规的大肠杆菌中,lacI阻遏蛋白表达量不高,仅能满足细胞自身的lac操纵子,无法应付多拷贝的质粒的需求,导致非诱导条件下较高地表达,为了让表达系统严谨调控产物表达,能过量表达lacI阻遏蛋白的lacIq 突变菌株常被选为Lac/Tac/trc表达系统的表达菌株。
原核真核启动子及BL21相关知识
感受态BL21、BL21(DE3)及BL21(DE3)pLysSBL21没有T7 RNA聚合酶基因, 因此不能表达T7启动子控制的目的基因, 但是可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)是λDE3溶原菌,带有T7 RNA聚合酶,可用于表达T7启动子控制的目的基因,也可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)pLysS含有pLysS质粒,一种与pET共存的表达T7裂解酶的质粒,可以减少目的基因的本底表达,提供更严紧的控制。
真核原核启动子原核:几个常用的启动子和诱导调控表达系统1.最早应用于的表达系统的是Lac乳糖操纵子,由启动子lacP + 操纵基因lacO + 结构基因组成。
其转录受CAP正调控和lacI负调控。
cUV5突变能够在没有CAP的存在下更有效地起始转录,该启动子在转录水平上只受lacI 的调控,因而随后得到了更广泛采用。
lacI产物是一种阻遏蛋白,能结合在操纵基因lacO 上从而阻遏转录起始。
乳糖的类似物IPTG可以和lacI产物结合,使其构象改变离开lacO,从而激活转录。
这种可诱导的转录调控成为了大肠杆菌表达系统载体构建的常用元件。
3.tac启动子是trp启动子和lacUV5的拼接杂合启动子,且转录水平更高,比lacUV5更优越。
4.trc启动子是trp启动子和lac启动子的拼合启动子,同样具有比trp更高的转录效率和受lacI阻遏蛋白调控的强启动子特性。
5.在常规的大肠杆菌中,lacI阻遏蛋白表达量不高,仅能满足细胞自身的lac操纵子,无法应付多拷贝的质粒的需求,导致非诱导条件下较高地表达,为了让表达系统严谨调控产物表达,能过量表达lacI阻遏蛋白的lacIq 突变菌株常被选为Lac/Tac/trc表达系统的表达菌株。
现在的Lac/Tac/trc载体上通常还带有lacIq 基因,以表达更多lacI阻遏蛋白实现严谨的诱导调控。
真核生物三类启动子

真核生物三类启动子(总3页) -本页仅作为预览文档封面,使用时请删除本页-真核生物启动子有三类,分别由RNA聚合酶Ⅰ、Ⅱ和Ⅲ进行转录。
类别Ⅰ(class Ⅰ)启动子:只控制rRNA前体基因的转录,转录产物经切割和加工后生成各种成熟rRNA。
类别Ⅰ启动子由两部分保守序列组成:核心启动子(core promoter):位于转录起点附近,从-45至+20;上游控制元件(upstream control element,UCE):位于-180至-107;RNA聚合酶Ⅰ对其转录需要2种因子参与:UBF1:一条M为97000的多肽链,结合在上述两部分的富含GC区;1个TBP,即TATA结合蛋白(TATA-binding protein,TBP);SL1:一个四聚体蛋白,含有 3个不同的转录辅助因子TAFⅠ;在SL1因子介导下RNA聚合酶Ⅰ结合在转录起点上并开始转录。
类别Ⅱ(class Ⅱ)启动子:类别Ⅱ启动子涉及众多编码蛋白质的基因表达的控制。
该类启动子包含4类控制元件:基本启动子(basal promoter):序列为中心在-25至-30左右的7 bp保守区,TATAAAA/T,称为TATA框或Goldberg-Hogness 框。
与RNA聚合酶的定位有关,DNA双链在此解开并决定转录的起点位置。
失去TATA框,转录将在许多位点上开始。
起始子(initiator):转录起点位置处的一保守序列,共有序列为:P y P y ANT(A)P y P yP y为嘧啶碱(C或T),N为任意碱基,A为转录的起点。
DNA在此解开并起始转录。
上游元件(upstream factor):普遍存在的上游元件有CAAT框、GC框和八聚体(octamer)框等。
CAAT框的共有序列是GCCAATCT,GC框的共有序列为GGGCGG和CCGCCC,八聚体框含有8bp,共有序列为ATGCAAAT;应答元件(response element):诱导调节产生的转录激活因子与靶基因上的应答元件结合。
启动子——精选推荐

启动⼦在遗传学中,启动⼦(promoter)是指⼀段能使特定基因进⾏转录的脱氧核糖核酸(DNA)序列。
启动⼦可以被RNA聚合酶辨认,并开始转录合成RNA。
在核糖核酸(RNA)合成中,启动⼦可以和调控基因转录的转录因⼦产⽣相互作⽤,控制基因表达(转录)的起始时间和表达的程度,包含核⼼启动⼦区域和调控区域,就像“开关”,决定基因的活动,继⽽控制细胞开始⽣产哪⼀种蛋⽩质。
启动⼦位于控制基因表达的调控序列中、基因转录起始位点的上游(DNA反义链的5′⽅向),长约100~1000个碱基对。
启动⼦本⾝并⽆编译功能,但它拥有对基因翻译氨基酸的指挥作⽤,就像⼀⾯旗帜,其核⼼部分是⾮编码区上游的RNA聚合酶结合位点,指挥聚合酶的合成,这种酶指导RNA的复制合成。
因此该段位的启动⼦发⽣突变(变异),将对基因的表达有着毁灭性作⽤。
完全的启动⼦称为规范序列。
⽬录1 启动⼦元件2 启动⼦序列2.1 原核⽣物启动⼦2.2 真核⽣物启动⼦3 结合4 与启动⼦功能变异有关的疾病5 参考⽂献启动⼦元件[编辑]启动⼦代表⼀些重要的元件可以与其他调节区域(如增强⼦、沉默⼦、边界元件或绝缘⼦)合作⼀致,以主导基因转录的⽔平。
由于启动⼦⼀般都是在基因的上游,启动⼦所在的位置或是转录起始点会由+1开始编号。
上游的位置所以都是由+1逆数的负数,例如-100就是位置100的上游碱基对。
以下是各种启动⼦:核⼼启动⼦是引发转录的必要部分及转录起始点,位置约为-35。
且是RNA聚合酶的结合位点及⼀般转录因⼦结合位点。
近端启动⼦是基因的近端序列上游,包括⼀些基本的调控元件,位置约为-250,且是特定转录因⼦结合位点。
远处启动⼦是基因的远处序列上游,包括⼀些额外的调控元件,影响⼒较近端启动⼦弱。
它是在上游更远的位置(但不是位置性的增强⼦或调控区域),是特定转录因⼦结合位点。
启动⼦规范序列的⽤途⼀般都是有问题的,且可引致对启动⼦序列的误解。
在规范序列中,转录因⼦结合位点在特定细胞情况下有⼀个单独的序列会与蛋⽩质牢固地结合。
真核生物三类启动子

真核生物启动子有三类,分别由RN A 聚合酶Ⅰ、Ⅱ和Ⅲ进行转录。
类别Ⅰ(class Ⅰ)启动子:只控制rR NA 前体基因的转录,转录产物经切割和加工后生成各种成熟rRN A 。
类别Ⅰ启动子由两部分保守序列组成:核心启动子(c ore promo t er ):位于转录起点附近,从-45至+20;上游控制元件(u pstr e am contr ol eleme n t ,UCE ):位于-180至-107;RNA 聚合酶Ⅰ对其转录需要2种因子参与:UBF1:一条M 为97000的多肽链,结合在上述两部分的富含G C 区;1个TBP ,即TA TA 结合蛋白(TA TA-bindi n g prote i n ,TBP );SL1:一个四聚体蛋白,含有 3个不同的转录辅助因子T AF Ⅰ;在SL1因子介导下R NA 聚合酶Ⅰ结合在转录起点上并开始转录。
类别Ⅱ(class Ⅱ)启动子:类别Ⅱ启动子涉及众多编码蛋白质的基因表达的控制。
该类启动子包含4类控制元件:基本启动子(b asal promo t er ):序列为中心在-25至-30左右的7 bp 保守区,T A TAA AA/T ,称为TAT A 框或Go l dber g -Hogne s s 框。
与RNA 聚合酶的定位有关,DNA 双链在此解开并决定转录的起点位置。
失去TAT A 框,转录将在许多位点上开始。
起始子(initi a tor ):转录起点位置处的一保守序列,共有序列为:Py PyA NT(A)P y P yPy 为嘧啶碱(C 或T ),N 为任意碱基,A 为转录的起点。
DNA 在此解开并起始转录。
上游元件(upstr e am facto r ):普遍存在的上游元件有C A A T 框、G C 框和八聚体(octam e r )框等。
真核生物三类启动子

真核生物启动子有三类,分别由RNA 聚合酶Ⅰ、Ⅱ和Ⅲ进行转录。
类别Ⅰ(class Ⅰ)启动子:只控制rRNA 前体基因的转录,转录产物经切割和加工后生成各种成熟rRNA 。
类别Ⅰ启动子由两部分保守序列组成:核心启动子(core promoter):位于转录起点附近,从-45至+20;上游控制元件(upstream control element ,UCE ):位于-180至-107;RNA 聚合酶Ⅰ对其转录需要2种因子参与:UBF1:一条M 为97000的多肽链,结合在上述两部分的富含GC 区;1个TBP ,即TATA 结合蛋白(TATA-binding protein ,TBP );SL1:一个四聚体蛋白,含有 3个不同的转录辅助因子TAF Ⅰ;在SL1因子介导下RNA 聚合酶Ⅰ结合在转录起点上并开始转录。
类别Ⅱ(class Ⅱ)启动子:类别Ⅱ启动子涉及众多编码蛋白质的基因表达的控制。
该类启动子包含4类控制元件:基本启动子(basal promoter ):序列为中心在-25至-30左右的7 bp 保守区,TATAAAA/T ,称为TATA 框或Goldberg-Hogness 框。
与RNA 聚合酶的定位有关,DNA 双链在此解开并决定转录的起点位置。
失去TATA 框,转录将在许多位点上开始。
起始子(initiator ):转录起点位置处的一保守序列,共有序列为:P y P y ANT(A)P y P yP y 为嘧啶碱(C 或T ),N 为任意碱基,A 为转录的起点。
DNA 在此解开并起始转录。
上游元件(upstream factor ):普遍存在的上游元件有CAAT 框、GC 框和八聚体(octamer )框等。
CAAT 框的共有序列是GCCAATCT ,GC 框的共有序列为GGGCGG 和CCGCCC ,八聚体框含有8bp ,共有序列为ATGCAAAT ;应答元件(response element ):诱导调节产生的转录激活因子与靶基因上的应答元件结合。
02真核基因的启动子 11PPT

启动子是一段特定的直接与RNA聚合 酶及其转录因子相结合、决定基因转录起 始与否的DNA序列。不同的启动子对RNA 聚合酶的亲和力不同,所结合的反式作用 因子(trans-acting factors)也不同,因此, 基因转录活性也很不相同。
Ⅰ.典型的原核启动子有四大要素:
转录起始位点,-10区,-35区和-10区与-35区之间 的间隔。
▪ 多数II类启动子有一个被称为TATA盒的共有序列,通常处于-30区, 相对于转录起始位点的位置比较固定。TATA盒存在于所有真核生物 中,TATA盒是一个保守的七碱基对,其序列为:
▪ 也有一些II类启动子不含有TATA盒,这样的启动子称 为无TATA盒其序列为 CAT(A为起始位点)。
-10区是一个6碱基保守序列(常以-10为中心)。 T80A95t45A60A50T96,有助于DNA的解链。
-35区是另一个6碱基保守序列(常以-35为中心)。 T82T84G78A65C54a45
研究表明,当-10区和-35区中心位置相距16-18bp 时,该启动子的功能较强;相距较近或较远时,起始 转录的功能都相应减弱。
Ⅱ.真核基因启动子
真核生物有3类RNA聚合酶,负责转录3类不同的启动子。
由RNA聚合酶I负责转录的rRNA基因,启动子(I类) 比较单一,由转录起始位点附近的两部分序列构成。第一 部分是核心启动子(core promoter),由-45—+20位核 苷酸组成,单独存在时就足以起始转录。另一部分由170—-107位序列组成,称为上游调控元件,能有效地增 强转录效率。
由RNA聚合酶Ⅲ负责转录的是5SrRNA、tRNA和某 些核内小分子RNA(snRNA),其启动子(Ⅲ类)组成 较复杂,又可被分为三个亚类。两类5S rRNA和tRNA基 因的启动子是内部启动子(internal promoter),位于转
真核生物三类启动子

真核生物启动子有三类,分别由RNA 聚合酶Ⅰ、Ⅱ和Ⅲ进行转录。
类别Ⅰ(class Ⅰ)启动子:只控制rRNA 前体基因的转录,转录产物经切割和加工后生成各种成熟rRNA 。
类别Ⅰ启动子由两部分保守序列组成:核心启动子(core promoter):位于转录起点附近,从-45至+20;上游控制元件(upstream control element ,UCE ):位于-180至-107;RNA 聚合酶Ⅰ对其转录需要2种因子参与:UBF1:一条M 为97000的多肽链,结合在上述两部分的富含GC 区;1个TBP ,即TATA 结合蛋白(TA TA-binding protein ,TBP );SL1:一个四聚体蛋白,含有 3个不同的转录辅助因子TAF Ⅰ;在SL1因子介导下RNA 聚合酶Ⅰ结合在转录起点上并开始转录。
类别Ⅱ(class Ⅱ)启动子:类别Ⅱ启动子涉及众多编码蛋白质的基因表达的控制。
该类启动子包含4类控制元件:基本启动子(basal promoter ):序列为中心在-25至-30左右的7 bp 保守区,TA TAAAA/T ,称为TATA 框或Goldberg-Hogness 框。
与RNA 聚合酶的定位有关,DNA 双链在此解开并决定转录的起点位置。
失去TATA 框,转录将在许多位点上开始。
起始子(initiator ):转录起点位置处的一保守序列,共有序列为:P y P y ANT(A)P y P yP y 为嘧啶碱(C 或T ),N 为任意碱基,A 为转录的起点。
DNA 在此解开并起始转录。
上游元件(upstream factor ):普遍存在的上游元件有CAAT 框、GC 框和八聚体(octamer )框等。
CAAT 框的共有序列是GCCAATCT ,GC 框的共有序列为GGGCGG 和CCGCCC ,八聚体框含有8bp ,共有序列为ATGCAAA T ;应答元件(response element ):诱导调节产生的转录激活因子与靶基因上的应答元件结合。
7-启动子与增强子及转录-7

在RNA聚合酶作用下发生第一次聚合反应 5’-pppG -OH + NTP – 5’ -pppGpN – OH + ppi • 4、释放
合成8-9nt的RNA片段后, σ 因子脱落,导致核心酶构象改变
转录起始复合物
封闭复 合物
RNA聚合酶全 酶+启动子 (DNA处于双链 状态)
转录起始点:在多数情况下为嘌呤,常见的序列为CAT
Pribnow框(-10区):RNA聚合酶牢固结合位点。保守序列的中心位于转录
起始上游的-10bp处。原核生物的共有保守序列为TATAAT,称为TATA框。 (1) RNA pol紧密结合; (2) 形成开放启动复合体; (3) 使RNA pol定向转录。 Sextama框(-35区):RNA聚合酶识别位点。保守序列的中心位于转录起始 上游的-35bp处。原核生物的共有保守序列为TTGACA,RNA聚合酶σ 因子可以识 别该位点。 (1) 为RNA pol的识别位点。
RNA polⅡ启动子结构
启动子Ⅱ最为复杂,它和原核的启动子有很多不同: (1)有多种元件:TATA框,GC框,CATT框等; (2)结构不恒定; (3)它们的位置、序列、距离和方向都不完全相同; (4)有的有远距离的调控元件存在,如增强子; (5)这些元件常常起到控制转录效率和选择起始位点的作用; (6)不直接和RNA pol结合; (7)需多种转录因子介入。
转录复合体
TAFs
TFIIA
TFIIB
TFIIF Pol II
TFIIE RNA pol Ⅱ的转录起始
第四节 原核生物RNA的合成
• 起始 • 延长 • 终止 • 后加工
一、转录起始
• 1、结合 RNA聚合酶全酶通过σ 因子与启动子结合,形成闭合复合物
常见真核启动子及原核启动子特点
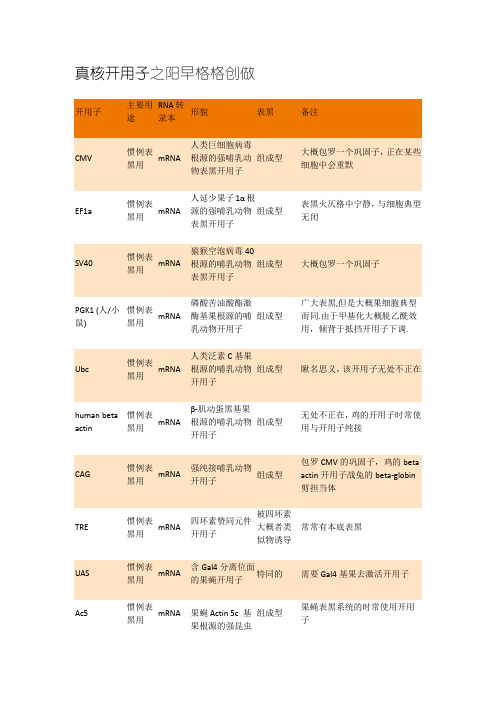
真核开用子之阳早格格创做EF1a惯例表黑用mRNA 人延少果子1α根源的强哺乳动物表黑开用子 组成型表黑火仄格中宁静,与细胞典型无闭PGK1 (人/小鼠) 惯例表黑用mRNA 磷酸苦油酸酯激酶基果根源的哺乳动物开用子组成型广大表黑,但是大概果细胞典型而同.由于甲基化大概脱乙酰效用,倾背于抵挡开用子下调.human beta actin 惯例表黑用mRNA β-肌动蛋黑基果根源的哺乳动物开用子组成型无处不正在,鸡的开用子时常使用与开用子纯接TRE惯例表黑用 mRNA 四环素赞同元件开用子被四环素大概者类似物诱导 常常有本底表黑Ac5惯例表黑用mRNA 果蝇Actin 5c 基果根源的强昆虫组成型果蝇表黑系统的时常使用开用子开用子CaMKIIa光遗传教基果表黑 mRNA Ca2+/钙调蛋黑依好的蛋黑激酶 II 开用子特同的 用于中枢神经系统/神经元表黑.受到钙战钙调蛋黑安排.TEF1 惯例表黑用 mRNA 酵母转录蔓延果子开用子 组成型与哺乳动物的EF1a 开用子类似ADH1惯例表黑用 mRNA乙醇脱氢酶I 的酵母开用子 被乙醇压造齐少版本很强,促进下表黑.截短开用子是组成型的,表黑较矮.Ubi惯例表黑用 mRNA 玉米泛素基果的动物开用子 组成型正在动物中促进下表黑U6 小RNA 表黑 shRNA 根源于人U6小核开用子组成型小鼠U6也使用,但是效用略好.时常使用的本核表黑系统开用子T7体中转录/惯例表黑T7噬菌体根源的开用子 组成型, 需要T7 RNA 散合酶当用于体中转录的时间,博路目标有大概是正背的也大概是反背的,与决于开用子相对付于手段基果的目标 T7lac下火仄基果表黑 T7噬菌体根源的开用子加上lac 把持子险些不本底表黑,需要T7RNA 散合酶,受到lac 把持子的统造,不妨被IPTG 诱导. 时常使用与pET 载体,受到lac 把持子的庄重调控Sp6体中转录/惯例表黑Sp6噬菌体根源的开用子 组成型, 需要SP6 RNA 散合酶 当用于体中转录的时间,博路目标有大概是正背的也大概是反背的,与决于开用子相对付于手段基果的目标 araBAD 惯例表黑用阿推伯糖代开把持子的开用子阿推伯糖诱导强,时常使用与pBAD 载体.符合于赶快调控战矮的本底表黑lac惯例表黑用 Lac 把持子根源的开用子 不妨被IPTG 大概者乳糖诱导正在惯例的大肠杆菌中,lacI 阻拦蛋黑表黑量不下,仅能谦脚细胞自己的lac 把持子,无法草率多拷贝的量粒的需要,引导非诱导条件下较下天表黑,为了让表黑系统宽紧调控产品表黑,能过量表黑lacI 阻拦蛋黑的lacIq 突变菌株常被选为Lac/Tac/trc 表黑系统的表黑菌株.当前的Lac/Tac/trc 载体上常常还戴有lacIq 基果,以表黑更多lacI 阻拦蛋黑真行宽紧的诱导调控.pL 下火仄基果表黑Lambda噬菌体根源的开用子温度安排常常战温度敏感的cI857 压造子拆配使用。
表达载体之原核VS真核

酿酒酵母表达系统常用启动子
1)糖酵解途径中关键酶的强启动子,受葡萄糖诱导:
甘油醛-3-磷酸脱氢酶基因GAPDH
磷酸甘油激酶基因PKG
乙醇脱氢酶基因ADH
2)半乳糖激酶启动子(GAL1)
半乳糖诱导、葡萄糖抑制
GAL8
GAL4
UAS
GAL1
GAL7
GAL1
A、 GAL1、GAL7和 GAL10基因连锁在一起,独立转录,共同调控
3、甲醇酵母表达系统操作原理
宿主株与标记基因 甲醇酵母系统的整合事件 胞内表达与分泌表达
甲醇酵母系统宿主
二大宿主系统主要特点
项目 最适温度 毕赤酵母 30℃ 汉森酵母 37℃
最适pH值
甘油阻遏 糖基化 高密度发酵
4.5
是 部分过度 100g/L
4.5
否 较正常 100g/L
二大宿主系统主要选择标记
表 达 载 体 之 原 核 真 核
VS 万 佩
原 核 真 核
都原 是核 我表 们达 表体 达系 重与 组真 基核 因表 蛋达 白体 的系 重, 要亦 载如 体阴 ,阳 也, 各相 有辅 优相 劣成
目的基因
构建
转化
培养,诱导表达载体重组载体表达菌株
“基因——载体——菌株——培养”
常见启动子 λpL启动子: trp启动子: trc启动子: 热诱导启动子 化学诱导启动子 trp+lac杂交启动子
(一) 酿酒酵母表达系统
酿酒酵母系统启动子 酿酒酵母分泌系统 酿酒酵母糖基化系统 酿酒酵母表达系统存在的问题
1、酿酒酵母启动子
起始 位点 mRNA
40-120bp
DAS
编码序列
UAS
真核生物的启动子

真核生物的启动子由于真核生物中有三种不同的RNA聚合酶,因此也有三种不同的启动子,其中以启动子Ⅱ最为复杂,它和原核的启动子有很多不同:(1)有多种元件:TATA 框,GC框,CATT框,OCT等;(2)结构不恒定。
有的有多种框盒如组蛋白H2B;有的只有TATA框和GC框,如SV40早期转录蛋白,(3)它们的位置、序列、距离和方向都不完全相同,(4)有的有远距离的调控元件存在,如增强子;(5)这些元件常常起到控制转录效率和选择起始位点的作用;(6)不直接和RNA pol结合。
转录时先和其它转录激活因子相结合,再和聚合酶结合。
(一)Ⅱ类基因的启动子和调控区Ⅱ类基因的启动子由核心元件和上游元件组成。
核心元件包括TATA框和转录起始位点附近的启始子(initiator,Inr)。
在起始点一般没有同源序列,但mRNA的第一个碱基倾向A,另一侧翼由Py组成(在原核启动子的CAT起始序列也有这种情况),称为起始子(initiator),一般由P Y2CAPY5构成,位于-3~+5,可能提供RNA pol Ⅱ识别。
无论TATA是否存在,Inr对于启动子的强度和起始位点的选择都是十分重要的。
现已分离纯化了与Inr特异结合的蛋白质因子。
1.核心元件TATA框合又称Hogness框,Goldberg-Hogness框,俚语称为金砖(Goldbrick),其一致序列是:T85A97T93A85A63A83A50,常在起始位点的上游-25左右,相当于原核的-10序列。
但-10是不可缺少的,而真核启动中也有的缺乏TATA框。
其作用是:(1) 选择正确的转录起始位点,保证精确起始,故也称为选择子(selector),当有的基因缺少TATA框时,可能由Inr来替代它的这一作用,如鼠的脱氨核苷转移酶(Tdt)基因就没有TATA框,但有17bp的Inr;(2)影响转录的速率。
TATA框的8bp的保守序列一般都是由A.T对组成,少数情况在其中的两个位点上由G.C对取代了A.T,可见它是较容易打开。
原核生物启动子的概念和组成特点

原核生物启动子的概念和组成特点研究了这么久原核生物启动子的概念和组成特点,总算发现了一些门道。
先来说说原核生物启动子的概念吧。
我觉得它就像是一场表演的开场信号。
在原核生物的基因表达这个大舞台上,启动子就是那个告诉演员(基因)可以开始表演(转录)的指令。
你想啊,就像一场音乐会,没有开场音乐或者主持人的开场话语,歌手也不知道什么时候该上台唱歌对吧,原核生物的基因也是一样,没有启动子发信号,就不知道什么时候该把自己的遗传信息转录成RNA。
说到它的组成特点呢,这里面就很有趣了。
原核生物启动子一般有几个关键的部分。
其中有个部位叫转录起始位点,这个我觉得特别神奇,它就像是比赛的起跑线一样,转录就从这个点开始。
比如说一群跑步运动员,转录起始位点就是他们听到枪声出发的那个位置。
还有个叫-10区的地方,也叫Pribnow框。
这个区的特点就是它那里的核苷酸序列很特殊,像是一个特殊的密码。
我就一直在想为啥要有这么个特殊的地方呢,它是不是就像一个门锁一样,有了这个特殊的序列,转录相关的酶才能识别并且结合在这儿,就像一把特殊的钥匙开一把特殊的锁那样。
再有就是-35区了。
我琢磨着这-35区和-10区可能就像是接力赛的两个接力点。
-35区也许就先把信号传递出去,然后-10区接力,继续引导转录的开始。
它们的间隔距离也很关键呢。
对了,我之前还以为这个距离是随便的呢,后来发现不是,这个距离必须要合适才行。
就跟跑步的时候,两个接力队员之间的距离不能太长也不能太短,合适的距离才能顺利交接接力棒。
我还想啊,原核生物启动子的组成特点肯定还有很多我们没发现的小奥秘呢。
在研究的过程中,我还发现有时候不同的原核生物,虽然大的组成部分是相似的,但是在一些细节的核苷酸序列上可能会有变化。
这就又像是不同的音乐会可能开场信号会有一些细微的差别一样。
这到底会对转录有多大的影响呢,这也是我一直比较困惑的地方。
不过我相信随着研究的深入,会发现更多关于原核生物启动子这个神秘家伙的秘密的。
原核真核启动子及BL21相关知识
感受态BL21、BL21(DE3)及BL21(DE3)pLysSBL21没有T7 RNA聚合酶基因, 因此不能表达T7启动子控制的目的基因, 但是可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)是λDE3溶原菌,带有T7 RNA聚合酶,可用于表达T7启动子控制的目的基因,也可以表达大肠杆菌启动子lac、tac、trc及trp控制的基因。
BL21(DE3)pLysS含有pLysS质粒,一种与pET共存的表达T7裂解酶的质粒,可以减少目的基因的本底表达,提供更严紧的控制。
真核原核启动子原核:几个常用的启动子和诱导调控表达系统1.最早应用于的表达系统的是Lac乳糖操纵子,由启动子lacP + 操纵基因lacO + 结构基因组成。
其转录受CAP正调控和lacI负调控。
cUV5突变能够在没有CAP的存在下更有效地起始转录,该启动子在转录水平上只受lacI 的调控,因而随后得到了更广泛采用。
lacI产物是一种阻遏蛋白,能结合在操纵基因lacO 上从而阻遏转录起始。
乳糖的类似物IPTG可以和lacI产物结合,使其构象改变离开lacO,从而激活转录。
这种可诱导的转录调控成为了大肠杆菌表达系统载体构建的常用元件。
3.tac启动子是trp启动子和lacUV5的拼接杂合启动子,且转录水平更高,比lacUV5更优越。
4.trc启动子是trp启动子和lac启动子的拼合启动子,同样具有比trp更高的转录效率和受lacI阻遏蛋白调控的强启动子特性。
5.在常规的大肠杆菌中,lacI阻遏蛋白表达量不高,仅能满足细胞自身的lac操纵子,无法应付多拷贝的质粒的需求,导致非诱导条件下较高地表达,为了让表达系统严谨调控产物表达,能过量表达lacI阻遏蛋白的lacIq 突变菌株常被选为Lac/Tac/trc表达系统的表达菌株。
现在的Lac/Tac/trc载体上通常还带有lacIq 基因,以表达更多lacI阻遏蛋白实现严谨的诱导调控。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
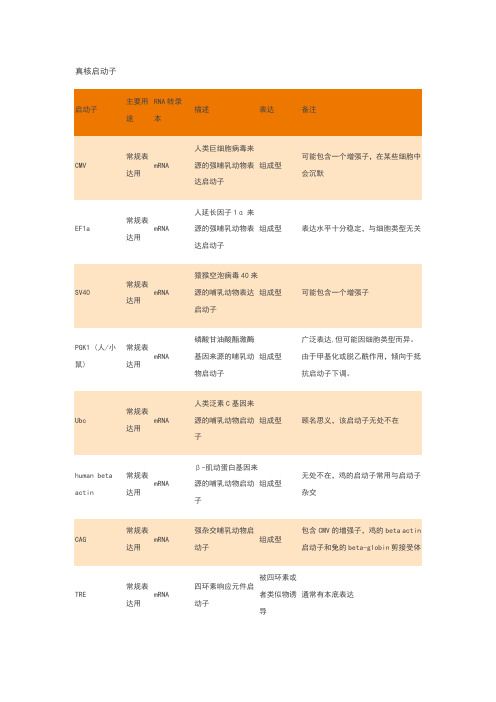
常见真核启动子及原核
启动子特点
集团企业公司编码:(LL3698-KKI1269-TM2483-LUI12689-ITT289-
真核
启动子
EF1a
常规
表达用
mRNA 人延长因子
1α来源的强哺乳动物表达启动子 组成型?
表达水平十分稳定,
与细胞类型无关
PGK1(人/小鼠)
常规
表达用
mRNA 磷酸甘油酸
酯激酶基因
来源的哺乳
动物启动子
组成型? 广泛表达,但可能因
细胞类型而异。
由于
甲基化或脱乙酰作
用,倾向于抵抗启动
子下调。
humanbetaactin 常规
表达用
mRNA
β-肌动蛋白基因来源的哺乳动物启动子
组成型?
无处不在,鸡的启动
子常用与启动子杂交
TRE 常规
表达
用
mRNA
四环素响应
元件启动子
被四环
素或者
类似物
诱导
通常有本底表达
Ac5常规
表达
用
mRNA
果蝇Actin5c
基因来源的
强昆虫启动
子
组成型?
果蝇表达系统的常用
启动子
CaMKIIa 光遗
传学
基因
表达
mRNA?
Ca2+/钙调蛋
白依赖的蛋
白激酶II启
动子
特异的
用于中枢神经系统/
神经元表达。
受到钙
和钙调蛋白调节。
GAL1,10常规
表达
用
mRNA
酵母双向启
动子
被半乳
糖诱
导,被
葡萄糖
抑制
可以单独使用或一起
使用。
受GAL4和
GAL80调节。
TEF1常规
表达
用
mRNA
酵母转录延
伸因子启动
子
组成型?
与哺乳动物的EF1a
启动子类似
ADH1常规
表达
用
mRNA
乙醇脱氢酶I
的酵母启动
子
被乙醇
抑制
全长版本很强,促进
高表达。
截短启动子
是组成型的,表达较
低。
Ubi 常规
表达
用
mRNA
玉米泛素基
因的植物启
动子
组成型?在植物中促进高表达
U6小
RNA
表达
shRNA
来源于人U6
小核启动子
组成型?
小鼠U6也使用,但效
率略差。
常用的原核表达系统启动子
T7lac 高水
平基
因表
达
T7噬菌体
来源的启
动子加上
lac操纵
子
几乎没有本底
表达,需要
T7RNA聚合
酶,受到lac
操纵子的控
制,可以被
常用与pET载体,受到lac操纵子
的严格调控
IPTG 诱导。
Sp6
体外
转录/常规表达
Sp6噬菌体来源的启动子 组成型,需要
SP6RNA 聚合
酶
当用于体外转录的时候,专路方向
有可能是正向的也可能是反向的,
取决于启动子相对于目的基因的方
向
araBAD 常规
表达
用
阿拉伯糖代谢操纵子的启动子
阿拉伯糖诱导
弱,常用与pBAD 载体。
适合于快速
调控和低的本底表达
lac
常规表达用 Lac 操纵子来源的启动子 可以被IPTG
或者乳糖诱导
在常规的大肠杆菌中,lacI 阻遏蛋白表达量不高,仅能满足细胞自身的lac 操纵子,无法应付多拷贝的质粒的需求,导致非诱导条件下较
高地表达,为了让表达系统严谨调
控产物表达,能过量表达lacI 阻遏
蛋白的lacIq 突变菌株常被选为
Lac/Tac/trc 表达系统的表达菌株。
现在的Lac/Tac/trc 载体上通常还带有lacIq 基因,以表达更多lacI 阻遏蛋白实现严谨的诱导调控。
Ptac
常规表达用 Lac 和trp 启动子杂交而成 与lac 启动子
的调节方式类似
转录水平高,比lacUV5优越
pL
高水
平基
因表
达?
Lambda 噬菌体来源的启动子
温度调节?
通常和温度敏感的cI857抑制子搭配使用。