第七章线粒体dna多态性85页PPT
合集下载
线粒体PPT课件
分布:通常分布于细胞生理功能旺盛的区域和需要能
量较多的部位,如肌细胞的肌纤维和精细胞的尾部;或 较为集中分布在有较多氧化反应底物的区域,如脂肪滴, 因为脂肪滴中有许多要被氧化的脂肪。
3
肌细胞的肌纤维和精子的尾部聚集较多的线粒体
线粒体包围着脂肪滴
线粒体的结构
光镜下线粒体结构
电镜下线粒体结构 4
线粒体的超微结构
5.线粒体的增殖与细菌一样——直接分裂。
20
21
五、线粒体的增殖----分裂
间壁分离
收缩分离
出芽分裂
22
23
生命活动中95%的能量来自线粒体
线粒体是糖、脂肪、氨基酸等能源物质最终氧 化释放能量的场所。
细胞氧化(细胞呼吸)
在特定的细胞器(主要是线粒体)内,在O2的参与 下,分解各种大分子物质,产生CO2,同时,分解代 谢所释放的能量储存于ATP中的过程,称为细胞氧 化、细胞呼吸或生物氧化。
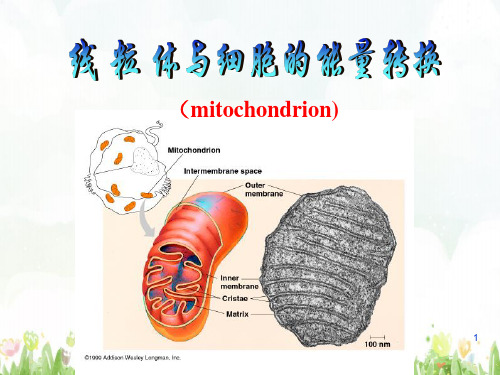
(mitochondrion)
1
一 线粒体的形态、数量和结构 形态:光镜下多数细胞的线粒体呈线状、粒状或杆状,形 态的差异与细胞类型及生理状态有关。
大小:细胞内较大的细胞器。一般直径:0.5—1.0um;长度:3um。在骨
骼肌细胞、胰脏外分泌细胞中可长达10~20μm,称巨线粒体。
2
成熟哺乳动物红细胞无线粒体。
一个线粒体内平均有5-10个mtDNA 双链环状的DNA分子,裸露不与组蛋白结合。 mtDNA结构紧密,编码37个基因。 mtDNA复制贯穿于整个细胞周期。
15
2种 编码 rRNA(12S和16S)基因 mtDNA 16569 bp 37个基因 22种 编码 tRNA基因 13种 编码 蛋白质基因
第七章线粒体的结构与功能PPT课件

1894年 ——Altmann —— 光镜 —— 生命小体 (bioblast) 1897年 —— Benda —— 线粒体(mitochondria)
一.线粒体的形态、大小和分布 形态:光镜: 线状、粒状、短杆状;有的圆形、哑铃形、星形;还有分枝状、环状等
*
线粒体的形态
光学显微镜下线粒体的形态
返回目录
*
化学渗透学说示意图
*
细胞氧化:在酶的催化下,氧将细胞内各种供能物质氧化而释放能量的过程。由于细胞氧化过程中,要消耗O2释放CO2和H2O所以又称细胞呼吸。
*
细胞氧化的基本过程
1、酵 解: 在细胞质基质内进行,反应过程不需要氧——无氧酵解
2、乙酰辅酶A生成: 线粒体基质内进行
3、三羧酸循环: 在线粒体基质内进行
线粒体的形态多种多样, 一般呈线状,也有粒状或短线状。细胞的生理状况发生变化时线粒体的形态亦将随之而改变。
*
线粒体的数量
同一类型细胞中,线粒体的数目是相对稳定的。 在不同类型的细胞中线粒体的数目相差很大。 生理活动旺盛的细胞(心肌细胞)线粒体多。
数百 ~ 数千个
3 105万个(有些卵母细胞)
特征酶:苹果酸脱氢酶
氧化还原酶 37%*源自线粒体各部分蛋白及酶的分布
线粒体的化学组成
线粒体的化学组分主要是由蛋白质、脂类和水份等组成
红色标注各部分的标志酶
*
线粒体:提供细胞95%以上的能量--- 细胞内的动力工厂 糖酵解:提供细胞少量的能量
细胞内的供能物质:主要糖类
*
知识回顾:真核细胞中的氧化作用 糖的氧化: 葡萄糖→细胞→ 胞质中分解为丙酮酸(不需要氧,糖酵解) ◆糖氧化成丙酮酸 ◆丙酮酸脱羧生成乙酰CoA ◆乙酰CoA进入三羧酸循环彻底氧化
一.线粒体的形态、大小和分布 形态:光镜: 线状、粒状、短杆状;有的圆形、哑铃形、星形;还有分枝状、环状等
*
线粒体的形态
光学显微镜下线粒体的形态
返回目录
*
化学渗透学说示意图
*
细胞氧化:在酶的催化下,氧将细胞内各种供能物质氧化而释放能量的过程。由于细胞氧化过程中,要消耗O2释放CO2和H2O所以又称细胞呼吸。
*
细胞氧化的基本过程
1、酵 解: 在细胞质基质内进行,反应过程不需要氧——无氧酵解
2、乙酰辅酶A生成: 线粒体基质内进行
3、三羧酸循环: 在线粒体基质内进行
线粒体的形态多种多样, 一般呈线状,也有粒状或短线状。细胞的生理状况发生变化时线粒体的形态亦将随之而改变。
*
线粒体的数量
同一类型细胞中,线粒体的数目是相对稳定的。 在不同类型的细胞中线粒体的数目相差很大。 生理活动旺盛的细胞(心肌细胞)线粒体多。
数百 ~ 数千个
3 105万个(有些卵母细胞)
特征酶:苹果酸脱氢酶
氧化还原酶 37%*源自线粒体各部分蛋白及酶的分布
线粒体的化学组成
线粒体的化学组分主要是由蛋白质、脂类和水份等组成
红色标注各部分的标志酶
*
线粒体:提供细胞95%以上的能量--- 细胞内的动力工厂 糖酵解:提供细胞少量的能量
细胞内的供能物质:主要糖类
*
知识回顾:真核细胞中的氧化作用 糖的氧化: 葡萄糖→细胞→ 胞质中分解为丙酮酸(不需要氧,糖酵解) ◆糖氧化成丙酮酸 ◆丙酮酸脱羧生成乙酰CoA ◆乙酰CoA进入三羧酸循环彻底氧化
线粒体DNA多态性

mtDNA分析技术
mtDNA分析技术
基本技术:DNA测序
第一代荧光自动测序技术:HVR I 和 HVR II全控 制区(control region, CR) 第二代测序技术:全线粒体基因组
mtDNA分析技术
mtDNA分析适用检材
缺少毛根,毛球或附着组织的毛发,断裂的毛干 适用提取DNA方法:碱裂解法
瓶颈效应和复 制分离
下一代平均异 质性水平相差 较大
瓶颈效应指卵细胞 中的mtDNA拷贝, 只有数个能进入下 一代。
线粒体遗传疾 病的家族间临 床变异性
第 二
mtDNA多态性 节
mtDNA多态性
线粒体DNA分区
控制区(非编码区,D环):16024~576
绝大部分mtDNA多态性都集中在此区 高变区(high variabe regions, HVR)I和 II16024~16365,73~340 编码区
线粒体DNA多态性
贵州医科大学法医学院 任峥
主要内容内容
掌握
线粒体DNA的特点
线粒体DNA单倍型类群的概念和 意义
线粒体DNA分型的结果解释
主要内容
线粒体DNA与 线粒体DNA的 线粒体DNA分 核DNA的区别 多态性类型 型的命名
线粒体DNA的 线粒体DNA的
结构
分析技术
第
概述
一 节
mtDNA结构与功能
• mtDNA异质性
两种不同的mtDNA序列存在于同一个细胞, 组织或个体时称之为异质性(heteroplasmy)
表现:2个峰成分,次要峰占主峰40%以上, 可重复
• mtDNA异质性
出现几率:2%~8%(一代测序)
1~2个碱基:考虑异质性 两样本来自同一母系
线粒体 PPT

参与电子传递的电子载体有5种:黄素蛋白、细胞色素、泛 醌、铁硫蛋白和铜原子。
(3) 电子转运复合物及呼吸链
线粒体内膜呼吸链有4种复合物: 复合物I、
线粒体承担的能量转换实质是把H+跨膜电位差和质子浓度梯 度(pH差)形成的质子驱动力转换成ATP分子中的高能磷酸 键。
线粒体
有氧运动
细胞必须利用能量来维持它的正常生活
研究历史
1850年,德国生物学家第一个发现线粒体, 并推测这种颗 粒是由半透性的膜包被的。 1890, Altman 命名 Bioblast (生命小体)。 1898年 Benda 对线粒体进行命名。 1900年,Leonor Michaelis用染料Janus green对肝细胞进 行染色,发现线粒体具有氧化还原反应的作用。
γ亚基旋转的观察
ATP合酶的作用是可逆的合成ATP、水解ATP。ATP的水解, 利用ATP能量,使H+逆浓度梯度运输,可以建立起质子梯度。
6.3.2线粒体与细胞衰老
线粒体是细胞内自由基的源泉,正常情况下氧自由基可被线 粒体中的Mn2+-SOD所清除,机体衰老使SOD活性降低,氧 自由基积累在线粒体中,导致线粒体内膜参与能量转换的酶 系功能异常,引起细胞的衰老与死亡。
3 ATP合酶及结合变构机制
头部:F1偶联因子 (3α,3β,γ, ε, δ) 膜部:F0偶联因子:abc
a1b2c10-12
结合变构机制
在特定的时间,三个催化位点的构象不同 由于γ亚基的端部是高度不对称的,它的旋转引起β亚基3个 催化位点构象的周期性变化(L、T、O)
将α3β3γ固定在玻 片上,在γ亚基的 顶端连接荧光标记 的肌动蛋白纤维, 在含有ATP的溶液 中温育时,在显微 镜下可观察到γ亚 基带动肌动蛋白纤 维旋转。
(3) 电子转运复合物及呼吸链
线粒体内膜呼吸链有4种复合物: 复合物I、
线粒体承担的能量转换实质是把H+跨膜电位差和质子浓度梯 度(pH差)形成的质子驱动力转换成ATP分子中的高能磷酸 键。
线粒体
有氧运动
细胞必须利用能量来维持它的正常生活
研究历史
1850年,德国生物学家第一个发现线粒体, 并推测这种颗 粒是由半透性的膜包被的。 1890, Altman 命名 Bioblast (生命小体)。 1898年 Benda 对线粒体进行命名。 1900年,Leonor Michaelis用染料Janus green对肝细胞进 行染色,发现线粒体具有氧化还原反应的作用。
γ亚基旋转的观察
ATP合酶的作用是可逆的合成ATP、水解ATP。ATP的水解, 利用ATP能量,使H+逆浓度梯度运输,可以建立起质子梯度。
6.3.2线粒体与细胞衰老
线粒体是细胞内自由基的源泉,正常情况下氧自由基可被线 粒体中的Mn2+-SOD所清除,机体衰老使SOD活性降低,氧 自由基积累在线粒体中,导致线粒体内膜参与能量转换的酶 系功能异常,引起细胞的衰老与死亡。
3 ATP合酶及结合变构机制
头部:F1偶联因子 (3α,3β,γ, ε, δ) 膜部:F0偶联因子:abc
a1b2c10-12
结合变构机制
在特定的时间,三个催化位点的构象不同 由于γ亚基的端部是高度不对称的,它的旋转引起β亚基3个 催化位点构象的周期性变化(L、T、O)
将α3β3γ固定在玻 片上,在γ亚基的 顶端连接荧光标记 的肌动蛋白纤维, 在含有ATP的溶液 中温育时,在显微 镜下可观察到γ亚 基带动肌动蛋白纤 维旋转。
细胞生物学第七章 线粒体ppt课件

■ 两套遗传体系的协同性
通过离体实验发现两套 遗传体系的遗传机制不 同。 如放线菌酮是细胞质蛋 白质合成抑制剂,但是 对细胞器蛋白质的翻译 却没有作用。另外,一 些抗生素,如氯霉素、 四环素、红霉素等能够 抑制线粒体蛋白质的合 成,但对细胞质蛋白质 合成没有多大影响。 通过对转录的抑制研究, 发现线粒体基因转录的 RNA聚合酶也是特异 的(图)。
线粒体蛋白转运
图 线粒体蛋白转运的部位
分子伴侣(molecular chaperon)
概念:一类在序列上没有相关性但有共同功能的蛋白质,它 们在细胞内帮助其他含多肽的结构完成正确的组装,而且在 组装完毕后与之分离,不构成这些蛋白质结构执行功能时的 组份。 种类:伴侣素家族(chaperonin, Cpn)、热休克蛋白 家族 ( Hsp family )、 核质素、T 受体结合蛋白 (TRAP) 等 特征:1、分子伴侣对靶蛋白没有高度专一性,同一分子伴 侣可以促进多种氨基酸序列完全不同的多肽链折叠成为空间 结构、性质和功能都不相关的蛋白质。 2、它的催化效率很低。行使功能需要水解ATP,以改 变其构象,释放底物,进行再循环。 3、它和肽链折叠的关系,是阻止错误折叠,而不是促 进正确折叠。 4. 多能性(胁迫保护防止交联聚沉,转运,调节转录 和复制,组装细胞骨架) 5. 进化保守性
细胞生物学第七 章 线粒体
第一节、 线粒体的生物学特征
线粒体是能够在光学显微镜进行 观察的显微结构。 ● 1890年,德国生物学家 Altmann第一个发现线粒体。 ● 1897年对线粒体进行命名。 ● 1900年,Leonor Michaelis用 染料Janus green对肝细胞进行 染色,发现细胞消耗氧之后,线 粒体的颜色逐渐消失了,从而提 示线粒体具有氧化还原反应的作 用。
第7章 线粒体【生化课件(华中师大)】

( 二) 自主性的局限性 1. 基因组小
2. 合成的蛋白质种类少 线粒体中合成的多肽主要有细胞色素C氧化酶3个 线粒体中合成的多肽主要有细胞色素C氧化酶3 亚基.ATP合成酶中 的两个亚基NADH 合成酶中F NADH—C 亚基.ATP合成酶中F0的两个亚基NADH COQ还 原酶, 个亚基和细胞色素b 原酶,7个亚基和细胞色素b .
�
1 化学渗透假说主要内容: : (1)电子传递链在线粒体内膜呈不对称分布. 有些组分不仅可以传递电子,而且可以起质子 泵的作用. (2)当电子在内膜中迂回传递时所释放的能量 将质子从内膜的基质侧泵至膜间隙,从而使膜 间隙的质子浓度高于基质,因而在内膜两侧形 成pH梯度和电位差. (3)在这个梯度的驱动下,质子穿过内膜上的 ATP合成酶流回基质,其能量促使ADP和Pi合 成ATP.
3 mt DNA复制,RNA转录和蛋白质翻译所 需的许多酶和蛋白质因子都是由核DNA 编码的.
五,线粒体蛋白质的合成与运输
( 一) 合成
导肽: 由核DNA编码的构成线粒体的蛋白质前体的N 末端含有识别线粒体膜上特异受体,指导蛋白 质在线粒体中定位的一段氨基酸序列,称为导 肽. 其特点是: (1)含丰富的带正电荷的氨基酸;(2)羟基 氨基酸含量也高;(3)几乎不含带负电荷的 氨基酸;(4)可形成双亲性的螺旋结构.
3线粒体上酶的分布
部位
外膜 膜间隙 内膜
基质
标志酶
单胺氧化酶 腺苷酸激酶
细胞色素氧化酶
主要酶
电子传递链
ATP合成酶
苹果酸脱氢酶
脂肪酸氧化酶系 三羧 酸循 环酶系(琥珀酸脱氢 酶除外)核酸和蛋白质合成酶系
三,线粒体的功能
(一)三羧酸循环: 三羧酸循环:
DNA多态性及其分析(共51张PPT)

反转重复序列,即旋转对称结构
一般为4~8个碱基对
例如 EcoRI 的识别序列:
5'-GA A T T C-3'
3'-C T T A A G-5’
BamHI 识别序列:
5’-GGATCC-3’
3’-CCTAGG-5’
2.核酸探针
由于人基因组DNA链很长,用某一限制性内切酶消化后,酶解
出来的各种不同长度的片段往往数量巨大,电泳后几乎是连续排 列的,无法进行分析比较。
扩增DNA的限制性酶切片段长度多态性(Amplification
一.RFLP分析技术
(一)基本原理
限制性酶切片段长度多态性技术,简称 RFLP技术(restriction fragment length
polymorphism,RFLP)
1. 限制性核酸内切酶(restriction endonuclease) 能够识别DNA双链上特异的碱基序列并能将之切断。 不同的限制性内切酶有各自特异的识别序列,一般 为4~8个碱基对 。
Y染色体——在哺乳动物的染色体中Y染色体似乎是
最少保守性的。
用不同的限制性内切酶和DNA探针,可以揭示出Y
染色体特异性片段的多态性。
四.线粒体的DNA多态性
线粒体DNA属于核外基因,表现为非孟德尔规律 遗传。
人mtDNA D环一端347bp的序列分析资料证明,两 个无关个体间,这一段mtDNA序列完全相同的可能 性只有0.27%(即1/370)。
采用合适的限制性内切酶 切出一系列长度的DNA片段
根据片段是否长度一致来推测其DNA序列是否相同 用标记好的相应的DNA探针与这些片段进行杂交 图谱反映出基因组DNA碱基序列组成是否存在差
异。
RFLP——通过核酸的限制性酶切片段长度多态性 来揭示DNA碱基序列组成的不同,因而称为限制性
医用细胞生物学》课件线粒体ppt演示课件

25
个人观点供参考,欢迎讨论!
向内突起形成嵴 内膜与嵴的内表面遍布着基粒
• 膜间腔:含多种可溶性酶、底物和辅助因子。
• 基质腔:含线粒体DNA,核糖体及各种酶类
19
五、线粒体的半自主性
• 线粒体具有自身的遗传物质mtDNA, 可编码37个基因,其中13个编码蛋白 质。
• 具自身的核糖体、有蛋白质合成系统
• mtDNA的复制、转录所需的酶、构成 线粒体核糖体的蛋白质都是由核DNA 编码、在细胞质中合成运进线粒体的。
• 线粒体是细胞的供能中心。
14
真核细胞中碳水化合物代谢俯瞰 15
三 羧 酸 循 环
16
化学渗透假说 17
基粒-合成ATP的关键部位
葡萄糖、氨基
酸等在基质腔
分解脱H+和电
子,电子通过
内膜传递促使
“H+”质子泵
入膜间腔,通
过基粒返回在
其头部合成ATP
基粒合成ATP的过程
18
线粒体的超微结构
• 外膜:光滑平整 • 内膜:高蛋白膜、电子传递链有关的酶类
同时线粒体遗传系统受控于细胞核遗传系同时线粒体遗传系统受控于细胞核遗传系2324间壁分离间壁分离收缩分离收缩分离出芽分裂出芽分裂25一一线粒体基因突变与疾病线粒体基因突变与疾病母系遗传及各种外界因素的影响母系遗传及各种外界因素的影响线粒体脑肌病线粒体脑肌病二线粒体中某些组分的治疗作用二线粒体中某些组分的治疗作用细胞色素细胞色素cc已被作为一氧化碳中毒新生已被作为一氧化碳中毒新生儿窒息肺功能不全高山缺氧心肌炎及儿窒息肺功能不全高山缺氧心肌炎及心绞痛的急救药或辅助药
• 细胞内95%以上的能量来自线粒体。
3
一、线粒体的形态
个人观点供参考,欢迎讨论!
向内突起形成嵴 内膜与嵴的内表面遍布着基粒
• 膜间腔:含多种可溶性酶、底物和辅助因子。
• 基质腔:含线粒体DNA,核糖体及各种酶类
19
五、线粒体的半自主性
• 线粒体具有自身的遗传物质mtDNA, 可编码37个基因,其中13个编码蛋白 质。
• 具自身的核糖体、有蛋白质合成系统
• mtDNA的复制、转录所需的酶、构成 线粒体核糖体的蛋白质都是由核DNA 编码、在细胞质中合成运进线粒体的。
• 线粒体是细胞的供能中心。
14
真核细胞中碳水化合物代谢俯瞰 15
三 羧 酸 循 环
16
化学渗透假说 17
基粒-合成ATP的关键部位
葡萄糖、氨基
酸等在基质腔
分解脱H+和电
子,电子通过
内膜传递促使
“H+”质子泵
入膜间腔,通
过基粒返回在
其头部合成ATP
基粒合成ATP的过程
18
线粒体的超微结构
• 外膜:光滑平整 • 内膜:高蛋白膜、电子传递链有关的酶类
同时线粒体遗传系统受控于细胞核遗传系同时线粒体遗传系统受控于细胞核遗传系2324间壁分离间壁分离收缩分离收缩分离出芽分裂出芽分裂25一一线粒体基因突变与疾病线粒体基因突变与疾病母系遗传及各种外界因素的影响母系遗传及各种外界因素的影响线粒体脑肌病线粒体脑肌病二线粒体中某些组分的治疗作用二线粒体中某些组分的治疗作用细胞色素细胞色素cc已被作为一氧化碳中毒新生已被作为一氧化碳中毒新生儿窒息肺功能不全高山缺氧心肌炎及儿窒息肺功能不全高山缺氧心肌炎及心绞痛的急救药或辅助药
• 细胞内95%以上的能量来自线粒体。
3
一、线粒体的形态
线粒体-课件(PPT-精)

Hot
Shock
Protein
热休克蛋白Hsp(主要的分子伴侣) 体外Hsp70 解折叠 腔內Hsp60、mHsp70 重折叠、组装
防止相互作用产生凝聚或错误折叠
识别蛋白质解折叠后暴露出的疏水面并与之结合
MPP 线粒体加工肽酶 PEP 加工增强性蛋白 磷酸转运蛋白
Processing Enhancing Protein Phospholipid Exchange Protein
电子转运复合物
破坏线粒体内膜后,可分离出4种膜蛋 白复合物。线粒体中氧化过程是由这四种 膜蛋白复合物相继作用来完成的。
电子转运复合物 复合物I(质子泵)
• 即NADH脱氢酶 • 催化1对电子从NADH传递给泛醌,每传一 递1对电子,伴随4个质子从基质转移到 膜间隙
电子转运复合物 复合物II
• 即琥珀酸-辅酶Q还原酶 • 琥珀酸脱氢酶有两种,一种是以泛醌作为 受体的,另一种是作用于所有受体。 • 催化从琥珀酸来的1对低能电子经FAD和 Fe-S傳递给泛醌,使FADH2上的电子通过 还原泛醌进入呼吸链中。
电子载体
• 细胞色素 一种带有含铁血红素辅基而对可见光 具有特征性强吸收的蛋白。 血红素中的铁通过Fe3+和Fe2+两种状态 的变换,传递单个电子。
电子载体
• 铁硫蛋白 一类含非血红素铁的蛋白质。 在铁硫蛋白分子的中央结合的是铁和硫, 称为铁硫中心。最常见的是在蛋白质的中 央含有2个铁原子和2个硫原子或含有4个铁 和4个硫,分别称为[2Fe-2S]和[4Fe-4S]。 也是靠Fe3+和Fe2+两种状态的转换传递电子。
相原和
都 是 疏 水 性 分 子
都 具 有 氧 化 还 原 作 用
第七章线粒体dna多态性

由于缺乏组蛋白的保护,线粒体亦缺乏 DNA 损 伤修复系统,mtDNA的突变率较高。
人mtDNA是一个长为16,569 bp的双链闭合环状分子, 外环含 G 较多,称重链 (H 链 ) ,内环含 C 较多,称轻链 (L 链)。 mtDNA双链闭合环状在降解检材中不易被核酸外切 酶影响,在法医检材中能保持mtDNA完整性。
线粒体遗传系统仍需依赖于细胞核遗传系统
细胞分裂时,突变型和野生型mtDNA发生 分离,随机地分配到子细胞中,使子细 胞拥有不同比例的突变型mtDNA分子。
• 在连续的分裂过程中,异质性细胞中突变 型mtDNA和野生型mtDNA的比例会发生漂 变,向同质性的方向发展。
表型的高度多态(polymorphism) mtDNA突变所产生的表型极为多态。 在一个家系中,患病个体的表现可以完 全不同;有的以血管病变为主,如中风、 冠心病等;有的表现为不同程度的肌无 力;有的以精神症状为主;有的以眼科 疾病为突出表现。发病的年龄差异也很 大。
COII
D
mtDNA-STR
主要集中在D-loop区;核心序列一般在10bp 以下,常见的是CA重复(nt514-nt523)和 polyC (nt568-nt573和nt303-nt309). CA重复:重复次数在10次以下。
• 近期研究表示线粒体DNA中某些重复序列 与疾病有关。如线粒体DNA拷贝数大大低 于正常,可引起致死性婴儿呼吸障碍。 实际人类mtDNA的长度不总是16569bp,
在法医学上常用的位点 • 序列多态性(Sequence polymorphism)
主要集中在D-loop区(法医学价值所在) HV1:nt16024~nt16365,342bp; HV2:nt 73~nt 340,268bp HV3:nt 438~nt 574,137bp
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
mtDNA与核常染色体DNA的比较
mtDNA
核常染色体DNA
结构
闭合、环状子,无组蛋白 线状,形成核小体
大小
大小为16,569bp
约30亿bp
数目 编码
50~几千个拷贝 /细胞 约有1,100bp非编码区
遗传方式 母系遗传
1个拷贝 /细胞
基因组大部分为非编 码区
孟德尔方式遗传
重组
不发生重组,单倍型遗传 发生重组
ND5 L2 S2 H ND4
ND4L
E ND6
“16,569” bp Q
Light (L) strand
OL
A
N C
Y
L1
ND1 Coding Region
I M
ND2
W
R
S1
ND3 G
COI
COIII
ATP6 ATP8
COII D
K
9-bp deletion
mtDNA-STR
主要集中在D-loop区;核心序列一般在10bp 以下,常见的是CA重复(nt514-nt523)和 polyC (nt568-nt573和nt303-nt309).
• 序列多态性(Sequence polymorphism)
主要集中在D-loop区(法医学价值所在) HV1:nt16024~nt16365,342bp; HV2:nt 73~nt 340,268bp HV3:nt 438~nt 574,137bp
• 长度多态性(Length polymorphism)
mtDNA突变可以影响线粒体功能,引起ATP合成障碍, 导致疾病发生,但实际上基因型和表现型的关系并非 如此简单。突变型mtDNA的表达受细胞中线粒体的异 质性水平以及组织器官维持正常功能所需的最低能量 影响,可产生不同的外显率和表现度。
瓶颈效应:
卵细胞中有100000左右拷贝的mtDNA,在受精卵形成时, 只有数个拷贝卵细胞的mtDNA进入到受精卵,这个现象称 为瓶颈效应。
PCR-RFLP法 PCR-SSO杂交法 PCR—DNA自动测序
目前分析mtDNA多态性最常用,也是最准 确的方法。
mtDNA单倍型
在分子进化的研究中,线粒体单倍型或单倍群是一组类似的 单倍型,它们有一个共同的单核苷酸多态性祖先。单倍群 由相似的单倍型组成,所以可以从单倍型来预测单倍群。 而单核苷酸多态性分析被用来确认单倍型。单倍群以字母 来标记,并且以数字和一些字母来补充。 与Y染色体DNA单倍群区别。
的拷贝数不同所产生的,其突出特征是限制性内切酶识别 位点本身的碱基没有发生改变,改变的只是它在基因组中 的相对位置。
mtDNA多态性的应用: 1.线粒体DNA与遗传性疾病的研究。 2. 生物进化学者分析人类线粒体DNA与其他
物种的对比,旨在确定它们的亲缘关系。
3. 在法医学上用于个体识别。
mtDNA单核苷酸多态性
mtDNA突变所产生的表型极为多态。 在一个家系中,患病个体的表现可以完 全不同;有的以血管病变为主,如中风、 冠心病等;有的表现为不同程度的肌无 力;有的以精神症状为主;有的以眼科 疾病为突出表现。发病的年龄差异也很 大。
突变率比较高
主要分布 控制区(D-环区)- 含有启动子和重链复制的起点
跨距约1100bp,个体间存在较大差异
CA重复:重复次数在10次以下。
• 近期研究表示线粒体DNA中某些重复序列 与疾病有关。如线粒体DNA拷贝数大大低 于正常,可引起致死性婴儿呼吸障碍。
实际人类mtDNA的长度不总是16569bp, 可能会有10bp左右的增加。
mtDNA多态性分析技术
• 长度多态性:AMP-FLP分析技术 • 序列多态性
高拷贝和异质性
mtDNA在一个细胞中具有数百次拷贝,因此mtDNA 突变比例可以在0到100%之间变化, 异质性:一两个不同的mtDNA序列存在于一个个体的细 胞、组织和不同的个体时。
• 阈值效应(threshold effects) 对于异质性的mtDNA突变,细胞可以承受正常
的mtDNA减少直到一个阈值时细胞才调亡或细胞的功 能受损。
主要为转换(90%):嘧啶转换 HV-I 80%
HV-II 20%
少数是顛换(10%): C→A 或 A→C
插入或缺失----长度多态性
poly—C:nt16184-nt16193 ( HV-I )
CA 重复:nt514-nt523 ( HV-III )
mtDNA多态性的类型
• 1.点的多态性 表现为DNA链中发生单个碱基的突变,且突变导致一个原有 酶切位点的丢失或形成一个新的酶切位点。Southern杂交 即可诊断。 • 2.序列多态性 ①由于DNA 顺序上发生突变如缺失、重复、插入所致。 ②由于高变区(highly variable region )内串联重复顺序
由于缺乏组蛋白的保护,线粒体亦缺乏DNA损 伤修复系统,mtDNA的突变率较高。
人mtDNA是一个长为16,569 bp的双链闭合环状分子, 外环含G较多,称重链(H链),内环含C较多,称轻链(L 链)。 mtDNA双链闭合环状在降解检材中不易被核酸外切 酶影响,在法医检材中能保持mtDNA完整性。
第七章 线粒体DNA多态性
学习目的
1 了解mtDNA的结构与功能 、线粒体基因组;
2 掌握mtDNA多态性 、mtDNA单核苷酸多态性 、 mtDNA 的串联重复序列多态性 、mtDNA单倍群;
3 了解mtDNA分型命名 、 mtDNA分析技术 (法医mtDNA 分型 、人类线粒体全基因组DNA分析 );
线粒体DNA 惟一的细胞核外DNA,携带有编码蛋白质和RNA的基因 • 结构:双链16569bp,
其中一条为重连,一条 为轻连。基因编码物 各不相同。 基因中不含非编码序列, 多数情况下,几乎不 含终止密码。 转录与翻译均在线粒体中进行。 DNA不与组蛋白结合 部分密码子不同于核基 因组密码子。
特征(1)碱基结构紧凑、简洁:以最少碱基数编码尽量 多的蛋白质和RNA ♦ 基因间无间隔序列 ♦ 基因内无内含子、前导序列、带帽序列和终 止信号 ♦ 相邻基因甚至有碱基的重叠 (2)不存在同源基因间的重组与交换 结果:mtDNA所有序列变异呈单倍型特性 卵细胞
遗传漂变:源自世代遗传过程中随机抽样引发的随机波动。 单倍型的比例来源:基因突变和遗传漂变。
人类线粒体DNA单倍型类群
人类线粒体DNA单倍型类群是遗传学上依据 线粒体DNA差异而定义出来的单倍群。可 以追溯母系遗传的人类遗传。
4 掌握mtDNA分型的特殊问题 。
DNA的分布
主要在染色体上
(所以说,染 色体是DNA的主 要载体)97.5%
细胞核遗传
细胞质内
2.5%
细胞质遗传
生物的遗传
前言
线粒体电镜照片
光镜下线粒体外型
多样,一般为颗粒状, 电镜下的主要特征是双 层膜且有嵴。线粒体中 分布着三羧酸循环系统 和电子传递系统,是细 胞氧化磷酸化反应产生 ATP的场所。此外,线 粒体是核外唯一含有遗 传效应物质的细胞器。
1981年Anderson等人完成最初的剑桥序列,样本主要是欧 洲血统的个体(这些样本包括一部分HeLa细胞和牛的序 列参入在内)。再次分析序列更正了最初序列,确定了11 处错误。现在以修正序列为准。
实际线粒体序列共有16569bp。为了保持历史数据,由一个 缺失占据一个位置。
在法医学上常用的位点
半自主性的细胞器
线粒体基因组能够单独进行复制、转录及合成蛋白质, 但这并不意味着线粒体基因组的遗传完全不受核基因的控制。 线粒体自身结构和生命活动都需要核基因的参与并受其控制, 说明真核细胞内尽管存在两个遗传系统,一个在细胞核内, 一个在细胞质内,各自合成一些蛋白质和基因产物,造成了 细胞核和细胞质对遗传的相互作用。
自身复制---单向复制:置换环复制或D-环(D-loop)复 制
特点:不对称的复制, H链和L链各有一个复制起点,先复制H链, H链复制到达L链起始点后,L链开始复制
D-环:新合成第三条H链姊妹链 ,序列与L链互补 H链复制起点附近( 520-700 ),高变异
转录功能--两条链同时转录合成RNA--对称转录
◆细胞器和细胞质颗粒中的遗传物质,统称 为细胞质基因组(plasmon)。
第一节 mtDNA的结构和功能
一、mtDNA的结构特征
线粒体是细胞质中独立的细胞器,也是动物细胞 核外唯一的含有DNA的细胞器。
1981 年 , 剑 桥 大 学 的 Anderson 小 组 测 定 了 人 mtDNA的完整DNA序列,称为“剑桥序列”。
线粒体遗传系统仍需依赖于细胞核遗传系统
细胞分裂时,突变型和野生型mtDNA发生
分离,随机地分配到子细胞中,使子细
胞拥有不同比例的突变型mtDNA分子。
• 在连续的分裂过程中,异质性细胞中突变 型mtDNA和野生型mtDNA的比例会发生漂 变,向同质性的方向发展。
表型的高度多态(polymorphism)
高 变 区 HV-I
29 ~ 408号碱基
HV-II 15996 ~ 16401号碱基
HV-III
438 ~ 574号碱基
主要原因 A : mtDNA没有组蛋白的保护
B : DNA聚合酶缺乏3’-5’外切酶校正功能
C : 缺乏DNA损伤的修复体系
D : mtDNA极少或不受来自选择压力的影响
突变类型 碱基的错配---序列多态性
突变率
高
低
全序列获得 1981年, Aderson序列(剑 2019年草图 桥序列)
二 mtDNA
二、mtDNA的遗传学特征
1. mtDNA为母系遗传 2 mtDNA高拷贝和异质性
3 mtDNA复制分离和瓶颈效应 4. mtDNA的阈值效应