_一对单显性大豆抗疫霉根腐病基因的遗传分析及定位 优先出版
大豆疫霉根腐病抗病基因分析及分子鉴定

wr o d l . D Pl e o y m to n e fr e i s s 恤 n t c tl l u s i r a v sa ne ec f iv t em e 击 o d rc o f o n t r o i l gt n ed h i e s s a . e P . s j o O e . s o y ea b ni n e t a r c t 1 n i o sat Pi y c lm a o el d o fg e et n og e n e比 l io t a n s h i P , T h e e r i s s t a n c e en g es ns i o y ea b nc nb a ep os t u 1 ed t a b s a do e nt e g h n f e rg o en et h e o . Ma y ’ I n y P l td n a i a e e s e r s i s 妞 n c e g e n si e cl n di u n g ve e s r a 1 P b t y o P h t h o ar r o tr o tr O e s i s t a n c e g e n e s i n s o y ea b nh a v eb ee nm a P edw p i t hm o1 ul c e rm a a r k e .U r i s gt n i g 1 1 t I yl i l 止 e dn l o 】 e ul c r a m r a k e st r od e t e tr c si e s t a 幻 c e en g n p 1 m o s a rc l u va i t s h r a s b e e nb c e o m i g ar n Pi a da n d e ec f iv t ew a y ri o f s d if t n ca i io t no fr e i s s a t n c e e g es n . T 卜 e o j b e iV t c e s ft o is h s t u d y er w et O d e t e mi r eP n h t y 0 p h t h o ar r O O t tr O r e i s s t a n c e g e n si e ns o es m o y e b nc a u1 t i v so ’ I a r1 i n e s , o t d i s c o v e r s 加v lr e e i s s l a n c eg ne e s , 即 dt os c r e e nr e i s S t a n t c u l t v i s o r a rl i n e s ob t eu S e d i d e r t c yf l o r i d e s ec s a o n r t 0 l nd a r e s i s a t n c e b e r e i d g. n R e a c t i o so n f l 2 l oy s ea b n lt U C i v so r a rl es n i c o l l e c t e d 丘 o ml Z P r o v l n c e s ol t Z s t r a i s n o f PS j o a e w e r e i d e n t i i f e d yu b si n g a od m i i f e d h y P o c o y t l i oc n ul io t a n Ch e t ni q u e . A m o n g
大豆疫霉根腐病部分抗性的遗传分析

关键词 : 大豆疫霉 ; 部分抗 性 ; 遗传分析
中图 分 类 号 :5 54 5 6 1 ¥6 .3 .5 文 献 标 识 码 : A 文章 编 号 : 0 9 8 (0 1 0 0 7 0 1 7— 04 2 1 )2— 19— 5 0
Ge ei a ayi o a t l ei a c oP t hh r o ei o b a n t n ls f ri s tn et o toas a sy e n c s p ar s p j n
(. 1 国家大豆改 良中心 , 南京农业大学农学院 , 作物遗传与种质创新 国家重点实验室 江苏 南京 20 9 ; 105
2 农 业 部病 虫 检 测 与 治 理 重 点 开 放 实 验 室 , 京 农 业 大 学 , . 南 江苏 南 京 2 0 9 ) 105
摘要 : 用苏 8 利 8一M 1× 沂 小黑 豆衍 生 的 16个 重 组 自交 家 系 ( J IX) 采 用 根 部 创 伤 接 种 方 法 接 种 大 豆 2 新 7 NRS , 疫 霉 菌 株 P 47, 究 大 豆 对 大 豆 疫 霉 根 腐 病 部 分抗 性 的 遗 传 机 制 。 结 果 表 明苏 8 69 研 8一M I对 大 豆 疫 霉 P4 7的 部 2 69 分 抗 性 是 由 2对 互 补 的 主基 因加 多基 因控 制 , 基 因 遗 传 率 为 7 .3 , 基 因遗 传 率 为 2 .9 。 主 41% 多 37%
2024中考生物二模试卷生物(湖南卷)

2024年中考第二次模拟考试(湖南省卷)生物(考试时间:60分钟试卷满分:100分)注意事项:1.答卷前,考生务必将自己的姓名、准考证号填写在答题卡上。
2.回答第Ⅰ卷时,选出每小题答案后,用2B铅笔把答题卡上对应题目的答案标号涂黑。
如需改动,用橡皮擦干净后,再选涂其他答案标号。
写在本试卷上无效。
3.回答第Ⅰ卷时,将答案写在答题卡上。
写在本试卷上无效。
4.考试结束后,将本试卷和答题卡一并交回。
第Ⅰ卷(选择题共50分)一、本题共25小题,每小题2分,共50分。
在每小题的四个选项中,选出最符合题目要求的一项。
1.在使用显微镜观察植物细胞时,要把低倍镜换成高倍镜应该转动如图中的()A.②B.③C.⑥D.⑦2.生物体在形成生殖细胞过程中,染色体数量要减少一半,玉米的体细胞中染色体数目为20条,它产生的精子中染色体数目是()A.10条B.20条C.10对D.20对3.被狗咬伤的患者,需要及时注射狂犬抗毒血清。
注射的物质和采取的措施及此免疫反应分别称为()A.抗体,控制传染源,非特异性免疫B.抗原,保护易感人群,特异性免疫C.抗体,保护易感人群,特异性免疫D.抗原,切断传播途径,非特异性免疫4.血液循环的动力来自心脏。
如图是哺乳动物心脏结构及与其相连的血管示意图,有关分析正确的是()A.②是主动脉,内流动着动脉血B.①是肺动脉,①中的血液流向③C.④和⑤之间有瓣膜,保证血液由④流向⑤D.血液循环包括体循环和肺循环,③是体循环的起点5.图所示的瓶子内装有正在萌发的种子,24小时后,把点燃的蜡烛放入瓶中,蜡烛熄灭,其原因是()A.种子进行呼吸作用,消耗了氧气B.种子进行蒸腾作用,放出了大量的水蒸气C.种子进行光合作用,放出了氧气D.种子进行呼吸作用,消耗了二氧化碳6.随着新冠的又一次来临,学校要求我们勤洗手,常通风,戴口罩,加强体育锻炼,这些措施属于()①控制传染源②切断传播途径③保护易感人群A.①②B.①③C.②③D.②7.2019年年底发现的新型冠状病毒,截至2021年6月1日,全球感染人数已经超过1.7亿,疫情形势依然十分严峻。
大豆资源抗疫霉根腐病基因分析

9 8 9年在东北大豆产区被发 国, 大豆疫霉最初于 1
3 ] 现[ , 随后在其它地区也发现了大豆疫霉的存 4 ~ 7 ] 在[ 。大豆疫霉是顽固的土传病原菌, 能够在大
收稿日期: 2 0 1 1- 0 1- 2 6 基金项目: 国家公益性 行 业 ( 农业) 科研专项( 3-2 0 ) ; 中国农业科学院作物科学研究所中央级公益性科研院所基本科研业务费专项 0 8 2 0 6 0 3 0 2- 0 6 ) ( 作者简介: 夏长剑( 1 9 8 5- ) , 男, 安徽太和人, 硕士研究生, 从事大豆抗疫霉根腐病研究, E- m a i l : x c h a n g j @y a h o o . c o m . c n 朱振东( 1 9 6 5- ) , 男, 湖南慈利人, 研究员, 博士, 研究方向为植物病害及抗病性, E- m a i l : z h u z d 1 1 5 @c a a s . n e t . c n 通讯作者:
[ 1 ] [ 1 6 ~ 1 8 ]
。但由
于大豆疫霉变异性较强, 抗病品种在田间出现新的 。因此, 不断发掘和利用 新的抗病基因, 对控制大豆疫霉根腐病、 保证大豆产 量及品质至关重要。本研究拟利用基因推导方法对 1 5 6份大豆资源的抗病基因进行分析, 旨在为抗病 育种提供有效抗源。
( N a t i o n a l K e yF a c i l i t yf o r C r o pG e n e t i c R e s o u r c e s a n dI m p r o v e m e n t / I n s t i t u t e o f C r o pS c i e n c e s ,C h i n e s e A c a d e m yo f A g r i c u l t u r a l S c i e n c e s ,B e i j i n g1 0 0 0 8 1 ,C h i n a ) A b s t r a c t :O n eh u n d r e da n df i f t ys i xs o y b e a na c c e s s i o n s w e r ei d e n t i f i e df o r t h e i r r e a c t i o n s t o 1 3p a t h o t y p e s o f P h y t o p h t h o r as o j a e u s i n g h y p o c o t y l i n o c u l a t i o nt e c h n i q u e i ng r e e n h o u s e . 3 1a c c e s s i o n s w e r e f o u n ds u s c e p t i b l e t o a l l 1 3p a t h o t y p e s ,w h e r e a s 1 2 5w e r er e s i s t a n t t oo n eo r m o r ep a t h o t y p e s ,a n dt h e s er e s i s t a n c ea c c e s s i o n s e l i c i t e d9 0 d i f f e r e n t r e a c t i o nt y p e s . P h y t o p h t h o r a r e s i s t a n c e g e n e s i nt h e r e s i s t a n c e a c c e s s i o n s w e r e p o s t u l a t e db y c o m p a r i n g t h e r e a c t i o nt y p e s o f d i f f e r e n t i a l s o y b e a nc u l t i v a r so r l i n e sp o s s e s s i n gk n o w na ss i n g l ep h y t o p h t h o r ar e s i s t a n c eg e n e . F i v er e a c t i o nt y p e s o c c u r r e di n9a c c e s s i o n s w e r ec o n s i s t e n t w i t ht h er e a c t i o nt y p e s o f as i n g l eg e n e ,w h i l e 5r e a c t i o nt y p e s p r o d u c e db y 1 2a c c e s s i o n s w e r e c o n s i s t e n t w i t ht w o - g e n e c o m b i n a t i o n s . E i g h t y r e a c t i o nt y p e s e l i c i t e db y 1 0 4a c c e s s i o n s c o u l db en o v e l r e a c t i o nt y p e s . K e yw o r d s : S o y b e a ng e r m p l a s m ;P h y t o p h t h o r as o j a e ;R e s i s t a n c ei d e n t i f i c a t i o n ;G e n ep o s t u l a t i o n 由大豆疫霉菌( P h y t o p h t h o r as o j a eK a u f m a n n& G e r d e m a n n ) 引起的大豆疫霉根腐病( P y t o p h t h o r a r o o t a n ds t e mr o t , P R R ) 是严重影响大豆生产的毁灭 性病害之一
_一对单显性大豆抗疫霉根腐病基因的遗传分析及定位 优先出版

http: / / nauxb.njau.edu.cn doi: 10.7685 / j.issn.1000-2030.2015.04.002
*
( 南京农业大学大豆研究所 / 国家大豆改良中心 / 农业部大豆生物学与遗传育种重点实验室 /
Genetic analysis and mapping of a single dominant Phytophthora sojae resistance gene in soybean
GUO Na , HU Guanjun, ZHAO Jinming, HUANG Jing, SUN Jutao, LI Lihong, GAI Junyi, XING Han *
J] . 南京农业大学学报, 2015, 38( 4) : 532-537 郭娜, 胡冠军, 赵晋铭, 等. 一对单显性大豆抗疫霉根腐病基因的遗传分析及定位[
一对单显性大豆抗疫霉根腐病基因的 遗传分析及定位
胡冠军, 赵晋铭, 黄婧, 孙聚涛, 李丽红, 盖钧镒, 邢邯 郭娜,
作物遗传与种质创新国家重点实验室, 江苏 南京 210095) 摘要: [ 目的] 大豆疫霉根腐病是大豆毁灭性的病害之一, 防治该病最安全有效的方法是培育和利用抗性品种 。 迄今为止, Rps1k 等) 抗性有效。 因此, 已在大豆基因组上鉴定了 21 个大豆疫霉根腐病抗性基因, 但是在中国只有少数基因( 如 Rps1c、 。‘大方六月早’ 急需发掘新的大豆疫霉根腐病抗性基因, 以满足抗病育种的需要 对大豆疫霉菌的抗谱较广, 是目前筛选出 ‘大方六月早’ ‘矮脚早 ’ 的优异抗源。[ 方法] 采用下胚轴创伤接种方法进行抗病性鉴定, 以品种 为抗病亲本与品种 配置杂 ‘大方六月早 ’ 交组合获得 F2 ∶3 代家系群体, 并进行遗传分析, 通过 SSR ( simple sequence repeat ) 标记对 的抗性基因进行定 : ‘大方六月早 ’ 位。[ 结果] 该群体的抗性遗传分析表明 对大豆疫霉根腐病的抗性由 1 对主效基因控制, 抗病对感病表现 为显性, 暂命名为 RpsAH。使用 SSR 标记进行分子作图, 抗病基因 RpsAH 被定位在大豆 3 号染色体( 即 N 连锁群) 上, 距离 ‘大方六月早’ 标记 Sat_166 为 4.1 cM。[ 结论] 大豆品种 对大豆疫霉菌株 AH 的抗性由 1 个单显性基因控制, 该基因定位于 大豆分子遗传图谱 N 连锁群上。 关键词: 大豆; 大豆疫霉菌; 遗传分析; SSR 标记; 抗性基因 中图分类号: S332.2; S565.1 文献标志码: A 文章编号: 1000-2030( 2015) 04-0532-06
大豆对疫霉根腐病抗病性鉴定方法

1999年6月June1999中国油料作物学报Chinese journal of oil crop sciences第21卷第2期V ol.21N o.2大豆对疫霉根腐病抗病性鉴定方法朱振东 王晓鸣(中国农业科学院作物品种资源所 北京 100081)摘要 应用下胚轴伤口接种方法鉴定85份大豆种质对大豆疫霉菌1号生理小种的抗病性,在两次鉴定中有95.29%的大豆品种(系)表现一致的抗病或感病反应。
本方法所获鉴定结果稳定可靠,利用注射器接种可以极大地提高工作效率,建议作为我国鉴定大豆种质资源对大豆疫霉根腐病抗病性的基本方法。
关键词 大豆 大豆疫霉菌 胚轴伤口接种 抗病性鉴定由大豆疫霉菌(Phytophthora sojae)引起的大豆疫霉根腐病是严重影响大豆生产的破坏性病害之一,该病于1948年首先发生于美国的印第安纳州,目前已分布于世界各主要大豆生产区[1]。
1989年沈崇尧和苏彦纯在我国黑龙江、吉林和北京从大豆罹病植株上首次分离到大豆疫霉菌[2]。
随后,其他一些研究者也相继在黑龙江的许多地区分离到该菌并发现病原菌存在致病力分化[3~5]。
朱振东和王晓鸣将其分离的大豆疫霉菌分离物鉴定为1号生理小种[6]。
大豆疫霉菌已成为影响黑龙江大豆生产的重要病原菌之一[4]。
大豆对大豆疫霉菌的抗性由显性单基因控制,种植抗病品种是防治大豆疫霉根腐病的主要方法[1]。
因此,鉴定大豆疫霉根腐病发生区主要栽培大豆品种的抗病性,筛选直接用于生产的抗病品种是控制病害发生和蔓延的有效途径;同时迅速开展我国大豆种质资源的抗病性鉴定,为抗病育种提供丰富抗原,加速抗病育种的进展。
本文对下胚轴伤口接种鉴定大豆对大豆疫霉根腐病抗病性的方法进行评价,为建立我国统一的对该病抗病性鉴定方法提供参考。
1 供试材料供试大豆疫霉菌为41-1菌株,从黑龙江农科院合江地区农科所试验田罹病大豆植株上分离获得,已鉴定为1号生理小种[6]。
菌株用稀释V8培养基保存和培养。
一种大豆腐霉根腐病抗性资源精准鉴定方法[发明专利]
![一种大豆腐霉根腐病抗性资源精准鉴定方法[发明专利]](https://img.taocdn.com/s3/m/bdd9489df46527d3250ce01b.png)
专利名称:一种大豆腐霉根腐病抗性资源精准鉴定方法
专利类型:发明专利
发明人:毕影东,来永才,刘明,李炜,张劲松,喻德跃,刘淼,王玲,刘建新,邸树峰,杨光,樊超,刘凯,栾晓燕,侯国强,夏天
舒,孙兵
申请号:CN201910537491.6
申请日:20190620
公开号:CN110199711A
公开日:
20190906
专利内容由知识产权出版社提供
摘要:本发明提供一种大豆腐霉根腐病抗性资源精准鉴定方法,所述方法利用高温灭菌的米粒接种腐霉菌,然后用培养好的接种体接种大豆种质资源种子,采用蛭石为基质在温室培养,对所述植株的病害进行调查、分级并计算鲜重比,通过对植株鲜重比(Ratio of weight)和病害指数(Disease Index)进行方差分析和进行聚类分析,筛选抗性和感性资源,鉴定出对腐霉菌根腐病具有抗/耐性的新种质。
本发明确定了大豆资源腐霉菌根腐病抗性标准评价体系,优化了病菌培养基配方,创建了米粒接种体培养和接种方法,研发了病害指数计算方法,该方法简单、实用、高效,也适用于大豆其他卵菌病害抗性鉴定研究。
申请人:黑龙江省农业科学院耕作栽培研究所
地址:150086 黑龙江省哈尔滨市南岗区学府路368号
国籍:CN
代理机构:北京金智普华知识产权代理有限公司
代理人:张晓博
更多信息请下载全文后查看。
河南大豆新品系抗大豆疫霉根腐病基因鉴定

作物学报ACTA AGRONOMICA SINICA 2021, 47(2): 275 284 / ISSN 0496-3490; CN 11-1809/S; CODEN TSHPA9E-mail: zwxb301@DOI: 10.3724/SP.J.1006.2021.04105河南大豆新品系抗大豆疫霉根腐病基因鉴定张雪翠1,**孙素丽1,**卢为国2李海朝2贾岩岩1段灿星1朱振东1,*1中国农业科学院作物科学研究所, 北京 100081; 2河南省农业科学院经济作物研究所, 河南郑州 450002摘要: 大豆疫霉根腐病作为影响大豆生产的毁灭性病害之一, 对大豆生产威胁很大。
种植抗疫霉根腐病的大豆品种是控制该病害最有效的途径。
河南省位于我国黄淮夏大豆产区的腹地, 具有大豆疫霉根腐病发生的潜在威胁。
本研究的目的是对河南省新育成的大豆品系进行抗性鉴定和抗病基因分子标记检测, 以明确大豆新品系对大豆疫霉根腐病的抗性水平和抗病基因。
采用下胚轴创伤接种法对64个河南省培育的大豆新品系进行接种, 鉴定其对2个具有不同毒力的大豆疫霉分离物PsJS2和Ps41-1的抗性。
结果显示, 对分离物Ps41-1和PsJS2抗病的分别有35个和16个品系, 对Ps41-1和PsJS2为中间反应型的分别有16个和10个品系, 其中对2个分离物均抗病的有16个品系, 占鉴定品系的25%。
使用抗疫霉病基因RpsZheng共分离标记WZInDel11进行新品系的基因型鉴定发现, 对2个大豆疫霉分离物均抗病的16个品系中有13个含有标记WZInDel11, 对1个或2个大豆疫霉分离物表现为中间反应型的5个大豆品系, 分子检测结果表明, 其为杂合基因型, 这些品系中的纯合抗病单株可直接选育成纯合抗病品系用于抗病育种。
综合系谱分析结果推测, 有2个品系可能含抗疫霉根腐病基因RpsZheng, 2个品系可能含RpsYD29, 14个品系可能含有RpsZheng或其等位基因。
大豆对大豆疫霉根腐病抗性的遗传分析

大豆对大豆疫霉根腐病抗性的遗传分析孙石;赵晋铭;武晓玲;郭娜;王源超;盖钧镒;邢邯【期刊名称】《中国农业科学》【年(卷),期】2009(042)002【摘要】[目的]研究大豆对大豆疫霉根腐病完全抗性和部分抗性两种抗性的遗传方式. [方法]利用不同抗性类型的品种(系)配置4个杂交组合,在分别采用下胚轴伤口接种法和根部接种法接种大豆疫霉菌株PNJ1条件下,研究两种抗性的遗传模式.[结果]豫豆25和郑92116对大豆疫霉菌的抗性由一对显性单基因控制,General和Conrad对大豆疫霉菌的部分抗性由1对加性主基因+加性-显性多基因控制,F2代主基因和多基因遗传率分别为41.31%~74.84%和15.60%~50.36%,F2:3代主基因和多基因遗传率分别为54.21%~77.05%和13.52%~38.24%.大豆对疫霉根腐病完全抗性和部分抗性分别属于不同的遗传体系. [结论]两类抗性都有育种价值,并且在早世代选择是有效的.选育聚合有完全抗性和部分抗性的品种是使大豆获得疫霉根腐病高水平抗性和持久抗性的最佳途径.【总页数】7页(P492-498)【作者】孙石;赵晋铭;武晓玲;郭娜;王源超;盖钧镒;邢邯【作者单位】南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095;南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095;南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095;南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095;南京农业大学农业部病虫监测与治理重点开放实验室,南京,210095;南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095;南京农业大学大豆研究所/国家大豆改良中心/作物遗传与种质创新国家重点实验室,南京,210095【正文语种】中文【中图分类】S4【相关文献】1.大豆疫霉根腐病部分抗性的遗传分析 [J], 武晓玲;赵晋铭;孙石;郭娜;王源超;盖钧镒;邢邯2.黑龙江省大豆品种对大豆疫霉根腐病抗性评价及抗性基因推导 [J], 徐鹏飞;王金生;姜良宇;李文滨;张淑珍;陈维元;吴俊江;李岑;邱丽娟;常汝镇3.野生大豆种质资源对大豆疫霉根腐病抗性评价 [J], 靳立梅;徐鹏飞;吴俊江;李文滨;邱丽娟;常汝镇;陈维元;于安亮;王金生;南海洋;陈晨;韩英鹏;陈艳秋;丁广洲;张淑珍4.黑龙江省主要栽培大豆品种(系)对大豆疫霉根腐病的多抗性评价 [J], 朱振东;王化波;王晓鸣;武小菲5.栽培大豆种质资源对大豆疫霉根腐病的抗性评价 [J], 张淑珍;徐鹏飞;吴俊江;李文滨;邱丽娟;常汝镇;陈维元;于安亮;王金生;靳立梅;陈晨;南海洋;陈艳秋;丁广洲因版权原因,仅展示原文概要,查看原文内容请购买。
大豆品种(系)对疫霉根腐病抗性研究
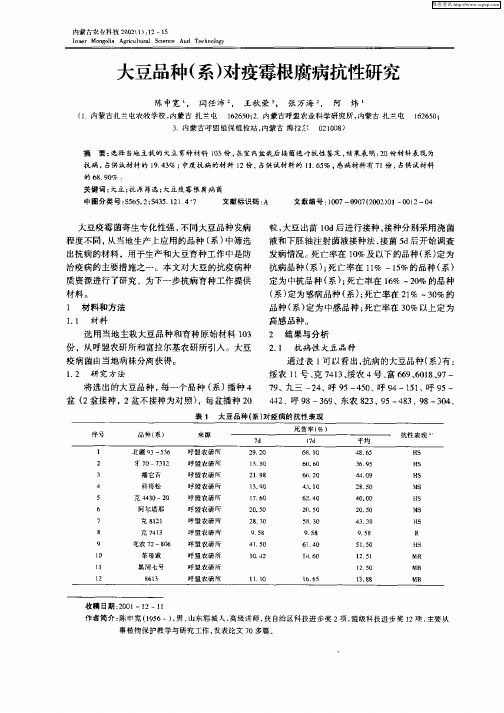
定 为 中抗 品 种 ( )死 亡率 在 1% ~ 0 的品 种 系 ; 6 2% ( ) 系 定为感 病 品种 ( )死 亡率在 2 % 一3% 的 系 ; 1 0 品种 ( ) 系 定为 中感 品种 ; 亡率 在 3 % 以上 定为 死 0
治疫病的主要措施之一。本文对大豆的抗疫病种 质资源进行了研究 ,为下一步抗病 育种工作提供
维普资讯
1 期
陈 申 等: 宽 大豆品种 ( 对痤霉根腐病 抗性 研究 系)
9 8—2 8 9 9 、 8—5 8 9 3 、 7—1 8 9 2 、 7—2 l1s和 9 5、n2 2—
3 结论 与讨 论
2 5共 2 份 , 5, 0 占试验材料的 1 .3 94 %。
大豆疫霉菌寄生专化性强 , 不同大豆品种发病
程 度不 同 , 当地生产 上应 用的 品种 ( ) 从 系 中筛选 出抗 病 的材 料 ,用于 生产 和大豆育 种 工作 中是 防
粒, 大豆 出苗 1d后进行 接种 , 0 接种分 别采用 浇 菌
液和下胚轴注射菌液接种法 , 接菌 5 后开始调查 d 发病情况。 死亡率在 1%及 以下的品种( 定为 0 系) 抗病品种 ( ; 系)死亡率在 1% ~1%的品种( 1 5 系)
2 2 中度抗 病品 种 .
13份大 豆材料 筛选结 果表 明 ,供试 材料 中 0 感病 品种 ( ) 7 份 ,占试 验材 料 的 6 0 , 系 为 1 89 % 抗病 品种 ( ) 2 系 为 O份 ,占试 验材料 的 1.3 , 94% 中间类型 为 l ,占试 验 材料 的 1 5 , 过 2份 1 % 通 6
大豆疫霉根腐病抗性的遗传分析及基因定位的初步研究的开题报告

大豆疫霉根腐病抗性的遗传分析及基因定位的初步
研究的开题报告
题目:大豆疫霉根腐病抗性的遗传分析及基因定位的初步研究
研究背景:
大豆是我国主要的农作物之一,也是世界上最为重要的粮食作物之一。
然而,大豆疫霉根腐病是造成大豆种植业损失较大的一种病害。
目前,对于大豆疫霉根腐病的抗性机制和遗传基础还知之甚少。
因此,研
究大豆疫霉根腐病抗性的遗传分析及基因定位,对于提高大豆种植的抗
病性和产量具有重要意义。
研究内容:
本研究将选取一批具有不同抗性水平的大豆材料,通过遗传分析和
基因定位技术,识别出与大豆疫霉根腐病抗性相关的基因。
具体内容为:
1. 收集大豆疫霉根腐病抗性材料并测定其抗病性水平;
2. 选取抗病性最强和最弱的两组材料进行遗传分析;
3. 利用分子标记和基因定位技术对两组材料进行基因组扫描和基因
定位;
4. 对基因定位结果进行验证。
研究意义:
本研究可为大豆疫霉根腐病抗性的遗传机制和分子调控机理提供理
论基础,并为大豆种植业的发展提供有价值的技术支持。
此外,本研究
还将有助于大豆产业的绿色化发展,减少由于病害而导致的农业失利,
提高农业生产效益。
大豆品种早熟18抗疫霉根腐病基因的SSR分子标记

大豆品种早熟18抗疫霉根腐病基因的SSR分子标记姚海燕;王晓鸣;武小菲;肖炎农;朱振东【期刊名称】《植物遗传资源学报》【年(卷),期】2010(11)2【摘要】大豆品种早熟18是抗疫霉根腐病的有效抗源。
本研究鉴定和分子标记早熟18的抗疫霉根腐病基因,以期为该品种的有效利用及分子辅助育种奠定基础。
以感病大豆品种Williams与早熟18杂交建立分离群体。
抗性遗传分析表明,早熟18对大豆疫霉菌抗性由1个显性单基因控制,该基因被定名为RpsZS18。
SSR标记连锁分析表明,RpsZS18位于大豆分子遗传连锁群D1b上的SSR标记Sat_069和Sat_183之间,与这两个标记的遗传距离分别为10.0 cM和8.3 cM。
RpsZS18是D1b连锁群上鉴定的第一个抗疫霉根腐病基因。
【总页数】5页(P213-217)【关键词】大豆;早熟18;疫霉根腐病;抗病基因;SSR标记【作者】姚海燕;王晓鸣;武小菲;肖炎农;朱振东【作者单位】中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程;华中农业大学植物科学技术学院【正文语种】中文【中图分类】S513【相关文献】1.与大豆疫霉根腐病抗性基因Rps1-c连锁的分子标记研究 [J], 曲娟娟;任桂萍;许修宏2.大豆品种绥农10抗疫霉根腐病遗传分析及抗病基因的SSR标记 [J], 于安亮;王欣;姜良宇;徐鹏飞;王金生;张淑珍;吴俊江;李文滨;陈维元;李宁辉;范素杰3.大豆抗疫霉根腐病基因SSR标记 [J], 刘丽君;孙欣;薛永国;杨喆4.用SSR标记分析抗疫霉根腐病大豆品种(系)的遗传多样性 [J], 杜青;朱振东;肖炎农;王晓鸣;武小菲5.大豆抗花叶病毒及耐疫霉根腐病的SSR标记分析 [J], 卢双勇;韩英鹏;滕卫丽;李文滨因版权原因,仅展示原文概要,查看原文内容请购买。
大豆根腐病研究进展

成 瑢等:大豆根腐病研究进展
· 59 ·
发生最重[2]。近年来 ,随着大豆种植面积不断扩大 ,大 豆重茬和迎茬情况严重 ,根腐病病情越来越难控制 。 笔者主要综述大豆根腐病的病原物生物学特征、发病 规律、抗病基因的克隆与应用,以及大豆根腐病的各种 防治方法介绍 ,以期为大豆根腐病的综合防控提供参 考依据。 1 大豆根腐病流行规律及其病原菌危害特征 1.1 大豆根腐病发病流行规律
基金项目:湖南农业大学“神农学者”计划;湖南农业大学青年科学基金“大豆 PI437654 抗孢囊线虫基因的克隆与功能分析”(14QN19)。 第一作者简介:成瑢,女,1990 年出生,湖南蓝山人,硕士研究生,研究方向:植物与微生物分子互作研究。通信地址:410128 湖南省长沙市芙蓉区 湖 南农业大学植物保护学院,E-mail:crl5423@。 通讯作者 :刘世名,男,教授,博士生导师,博士,研究方向 :植物病理学。通信地址 :410128 湖南省长沙市芙蓉区 湖南农业大学植物保护学院, E-mail:smliuhn@。 收稿日期:2015-10-16,修回日期:2015-12-22。
12大豆疫霉菌大豆疫霉菌为土传真菌易在有水沉积或湿度很大的土壤中引发大豆根腐病该病原菌主要通过土壤病残体及种子表皮内的卵孢子进行传播卵孢子在植株残体和土壤内能存活多年休眠卵孢子能在休闲病田中存活长达4年4
中国农学通报 2016,32(8):58-62 Chinese Agricultural Science Bulletin
· 60 ·
中国农学通报
表 1 大豆根腐病抗病基因
基因名称
Rps1
Rps2
Rps3
Rps4 Rps5 Rps6 Rps7 Rps8 Rps9 Rps10 RpsZS18 RpsSN10 RpsYB30 RpsYu25 RpsUN1 RpsUN2 RpsJS RpsYD29 RpsSu RpsAH 未命名
大豆品种(系)抗疫霉根腐病基因推导

大豆品种(系)抗疫霉根腐病基因推导陈晓玲;朱振东;王晓鸣;肖炎农;武小菲【期刊名称】《中国农业科学》【年(卷),期】2008(041)004【摘要】[目的]推导大豆品种(系)的抗疫霉根腐病基因,为病害防治筛选有效抗病品种(系).[方法]用下胚轴创伤接种法鉴定124个品种(系)对12个大豆疫霉菌菌株的反应型,通过基因推导方法分析品种(系)的抗病基因.[结果]124个品种(系)对12个菌株共产生51个反应型,13个品种(系)产生的反应型分别与几个含有已知抗病基因品种(系)的反应型相同,33个品种(系)产生的反应型符合一些两个已知抗病基因组合的反应型,这些品种(系)可能含有已知抗病基因或基因组合;52个品种(系)共产生37个既不同于任何含有单个已知抗病基因品种(系)的反应型也不同于两个已知抗病基因组合的反应型,它们可能含有新的抗病基因或基因组合.[结论]鉴定的大豆品种(系)普遍存在对疫霉根腐病的抗性,但抗病品种(系)数量和抗性水平存在地区问差异,同一地区多数抗病品种的遗传背景和抗性水平相似.【总页数】8页(P1227-1234)【作者】陈晓玲;朱振东;王晓鸣;肖炎农;武小菲【作者单位】中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程,北京100081;华中农业大学植物科技学院,武汉430070;中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程,北京100081;中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程,北京100081;华中农业大学植物科技学院,武汉430070;中国农业科学院作物科学研究所/国家农作物基因资源与基因改良重大科学工程,北京100081【正文语种】中文【中图分类】S4【相关文献】1.中国47个小麦新品种(系)苗期抗叶锈基因推导 [J], 袁军海;刘太国;陈万权2.黑龙江省大豆品种对大豆疫霉根腐病抗性评价及抗性基因推导 [J], 徐鹏飞;王金生;姜良宇;李文滨;张淑珍;陈维元;吴俊江;李岑;邱丽娟;常汝镇3.甘肃省50个主要小麦品种(系)苗期抗条锈基因推导及成株期抗病性分析 [J], 曹世勤;尚勋武;张勃;李明菊;徐世昌;骆惠生;金社林;贾秋珍;黄瑾;金明安4.黄淮麦区34个小麦主栽品种(系)抗条锈病基因推导 [J], 黄亮;刘太国;刘博;高利;罗培高;陈万权5.70份小麦品种(系)抗条锈病基因推导及检测 [J], 徐默然;吕晓欢;王凤涛;蔺瑞明;冯晶;徐世昌因版权原因,仅展示原文概要,查看原文内容请购买。
辽宁省沈阳市沈河区2023-2024学年八年级上学期期末生物试卷

辽宁省沈阳市沈河区2023-2024学年八年级上学期期末生物试卷(解析版)一、单项选择题(本题共12小题,每小题的四个选项中,只有一个最符合题意,请将你的选项直接涂黑在答题卡上,每小题1分,共12分)1.(1分)在山阴面或溪流附近,常见一类植物:叶子背面有褐色的孢子囊群,有不发达的输导组织()A.藻类植物B.蕨类植物C.苔藓植物D.裸子植物2.(1分)买麻藤是一种裸子植物,但因其茎和叶的特征以及成熟后能结出“红果”,常被误认为是被子植物。
下列相关叙述正确的是()A.买麻藤具有六大器官B.“红果”是买麻藤的种子C.其茎、叶中无输导组织D.“红果”外面有果皮3.(1分)有一种体形细长的动物,判断它是环节动物还是线形动物的主要依据是()A.身体是否分节B.是否依靠触手上刺丝捕食食物C.是否有肛门D.能否自由生活4.(1分)爬行动物比两栖动物更适应陆地生活的原因是()①体表覆盖角质鳞片或甲②肺发达③生殖和发育摆脱了对水的依赖④卵表面有坚韧的卵壳⑤皮肤裸露A.①②B.①②③④C.④⑤D.②③④5.(1分)现A、B、C、D四种动物它们的分情况是:A和B同“目”不同“科”,B和C 同“科”不同“属”,D和A不在同一“目”中()A.A和B B.D和A C.B和C D.都一样6.(1分)目前世界自然保护联盟“濒危物种红色名录”中评估的物种达到138374种。
造成野生动物濒临灭绝的主要原因是()A.自然灾害B.这些动物易感染病毒C.天敌过多D.人类活动对生态环境的破坏7.(1分)蔬菜上喷洒蜕皮徽素(促使昆虫加快蜕皮速度的化学物质)能减少菜粉蝶等害虫对蔬菜的危害,原因是蜕皮激素能()A.缩短幼虫期B.缩短蛹期C.缩短成虫期D.破坏害虫的取食器官8.(1分)蟾蜍是一种两栖动物,从它身上提取的蟾酥及蟾衣是我国紧缺的药材。
以下对蟾蜍生殖和发育的描述中,错误的是()A.发育过程为变态发育B.蝌蚪先生出后肢,再生出前肢C.幼体和成体在形态结构方面存在差异D.雌雄抱对后将受精卵排入水中9.(1分)科学家发现了一个决定睾丸发育的关键基因,如果缺少该基因,胎儿则不能发育出睾丸。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
收稿日期: 2014-12-02 基金项目: 国家公益性行业( 农业) 科研专项( ( 20130301 江苏省自然科学基金项目( BK20130684 ) ; 教育部长江学者和创新团队发展计划项目( PCSIRT13073)
* Email: guona@ njau.edu.cn。 作者简介: 郭娜, 讲师, 博士, 主要从事大豆抗病性研究, 教授, 博导, 主要从事大豆育种工作, 通信作者: 邢邯, 025-84399526, Email: hanx@ njau.edu.cn。
J] . 南京农业大学学报, 2015, 38( 4) : 532-537 郭娜, 胡冠军, 赵晋铭, 等. 一对单显性大豆抗疫霉根腐病基因的遗传分析及定位[
一对单显性大豆抗疫霉根腐病基因的 遗传分析及定位
胡冠军, 赵晋铭, 黄婧, 孙聚涛, 李丽红, 盖钧镒, 邢邯 郭娜,
作物遗传与种质创新国家重点实验室, 江苏 南京 210095) 摘要: [ 目的] 大豆疫霉根腐病是大豆毁灭性的病害之一, 防治该病最安全有效的方法是培育和利用抗性品种 。 迄今为止, Rps1k 等) 抗性有效。 因此, 已在大豆基因组上鉴定了 21 个大豆疫霉根腐病抗性基因, 但是在中国只有少数基因( 如 Rps1c、 。‘大方六月早’ 急需发掘新的大豆疫霉根腐病抗性基因, 以满足抗病育种的需要 对大豆疫霉菌的抗谱较广, 是目前筛选出 ‘大方六月早’ ‘矮脚早 ’ 的优异抗源。[ 方法] 采用下胚轴创伤接种方法进行抗病性鉴定, 以品种 为抗病亲本与品种 配置杂 ‘大方六月早 ’ 交组合获得 F2 ∶3 代家系群体, 并进行遗传分析, 通过 SSR ( simple sequence repeat ) 标记对 的抗性基因进行定 : ‘大方六月早 ’ 位。[ 结果] 该群体的抗性遗传分析表明 对大豆疫霉根腐病的抗性由 1 对主效基因控制, 抗病对感病表现 为显性, 暂命名为 RpsAH。使用 SSR 标记进行分子作图, 抗病基因 RpsAH 被定位在大豆 3 号染色体( 即 N 连锁群) 上, 距离 ‘大方六月早’ 标记 Sat_166 为 4.1 cM。[ 结论] 大豆品种 对大豆疫霉菌株 AH 的抗性由 1 个单显性基因控制, 该基因定位于 大豆分子遗传图谱 N 连锁群上。 关键词: 大豆; 大豆疫霉菌; 遗传分析; SSR 标记; 抗性基因 中图分类号: S332.2; S565.1 文献标志码: A 文章编号: 1000-2030( 2015) 04-0532-06
第4期
郭娜, 等: 一对单显性大豆抗疫霉根腐病基因的遗传分析及定位
533
由大豆疫霉菌( Phytophthora sojae Kaufmann & Gerdemann ) 引起的大豆疫霉根腐病 ( Phytophthora root [1 ] rot of Soybean , PRR) 是大豆生产上毁灭性的病害之一。1948 年, 在美国印第安纳州首次报道该病害 , 随
*
( 南京农业大学大豆研究所 / 国家大豆改良中心 / 农业部大豆生物学与遗传育种重点实验室 /
Genetic analysis and mapping of a single dominant Phytophthora sojae resistance gene in soybean
GUO Na , HU Guanjun, ZHAO Jinming, HUANG Jing, SUN Jutao, LI Lihong, GAI Junyi, XING Han *
( Soybean Research Institute / National Center for Soybean Improvement / Key Laboratory of Biology and Genetics and Breeding for Soybean, Ministry of Agriculture / State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China) Abstract: [ caused by Phytophthora sojae is a destructive disease on soybean Objectives]Phytophthora root and stem rot ( PRR ) , throughout the world. Utilization of resistance varieties is the most economical and environmentally safe method for controlling the disease. Up to now, 21 resistance genes have been identified in soybean. However, only a few genes, such as Rps1c and Rps1k, were effectively resistant to populations of P. sojae in China, so to mine new resistance genes is necessary for the disease. Soybean cultivar ‘Dafangliuyuezao’ has broad spectrum resistance to P. sojae, and is an elite resistance source for Phytophthora root rot of soybean. בAijiaozao ’ [ Methods] A population of F 2 ∶3 lines derived from the cross of ‘Dafangliuyuezao ’ was developed for resistance genetic analysis and resistance gene mapping. Using hypocotyl inoculation method at the seedling stage in the glasshouse, the reaction to P. sojae isolate AH( virulence type 2, 3a, 3b, 4, 5) , Results] The segregation ratio in the population in 156 F2 ∶3 lines were identified. [ fit into 1 ∶2 ∶1 for homozygous resistant, segregating and homozygous susceptible, showing that the resistance of ‘Dafangliuyuezao ’ to P. sojae strain AH is controlled by a single dominant gene, with the temporary name of RpsAH. On the basis of linkage analysis with SSR markers, RpsAH was located on molecular linkage group N, and flanked by Sat_166 with genetic distances 4.1 cM. [ Conclusions] The resistance to Phytophthora root rot is controlled by a single dominant gene, and the resistance gene RpsAH in ‘Dafangliuyuezao’ was mapped on molecular linkage group N. Keywords: soybean; Phytophthora sojae; genetic analysis; SSR markers; resistance gene
[2 ] 后在全世界范围内呈扩大蔓延趋势 。在美国, 由该病造成的经济损失每年超过 2.5 亿美元 。 在中国, 不 [3-6 ] , 同大豆产区相继发现大豆疫霉根腐病 可以在大 且呈加重趋势。大豆疫霉根腐病是一种土传性病害 ,
豆的任何生育期进行侵染并造成严重危害 。选育和种植抗病品种是防治该病最经济有效的方法 , 抗性的 [7-8 ] 。 遗传分析及抗病基因的定位是抗病育种的关键 大豆对大豆疫霉根腐病的完全抗性是单基因控制的质量性状抗病性 , 可抵御病害的扩展, 是当前抗病 [9-10 ] 。 到目前为止, Rps2、 育种中广泛利用的抗病性类型 已经鉴定的大豆疫霉根腐病抗性位点有 Rps1、 Rps3、 Rps4、 Rps5、 Rps6、 Rps7 和 Rps8 等[11-18]。 Rps1 ( 包 含 5 个 等 位 基 因 Rps1a、 Rps1b 、 Rps1c、 Rps1d 和 Rps1k) 与 Rps7 连锁, 定位在 3 号染色体( LG N) 上; Rps2 定位在 16 号染色体 ( LG J ) 上; Rps3 ( 包含 3 个等 Rps3b 和 Rps3c) 与 Rps8 连锁, 位基因 Rps3a、 定位在 13 号染色体 ( LG F ) 上; Rps4 和 Rps5 均与 Rps6 连锁, Rps10、 这 3 个抗性基因都定位在 18 号染色体( LG G) 上。最近, 研究人员又发现几个新的抗性基因 : Rps9、 RpsYu25、 RpsUN1 、 RpsUN2 、 RpsYD29 、 RpsJS 等[19-25]。 其中 Rps9、 RpsYu25、 RpsUN1、 RpsYD29 与 Rps1 连锁或 等位, 均定位在 3 号染色体( LG N) 上; RpsUN2 与 Rps2 连锁, 定位在 16 号染色体 ( LG J ) 上; Rps10 定位在 17 号染色体( LG D2) 上; RpsJS 定位在 18 号染色体( LG G) 上, 与 Rps4 和 Rps6 连锁或者等位。 然而, 大豆 疫霉的变异频率很快, 特别是在抗病品种的选择压力下, 新的毒力小种很容易产生和积累, 使得抗病品种 在推广 8 ~ 15 年后就会“丧失 ” 原有的抗性。 并且研究人员发现在国外鉴定的抗病基因中 , 只有 Rps1c 和 Rps1k 的抗性在中国有效[26]。因此, 迫切需要发掘新的大豆疫霉根腐病抗性基因 。 ‘矮脚早’ ( P 2 ) 杂交产生的 F 2 ∶3 代家系为材料, ‘大方六月早’ ( P1) 与 本研究以亲本 明确大豆疫霉根腐 , SSR ( simple sequence repeat ) , 病完全抗性的遗传规律 利用 标记对抗病位点进行定位 为抗性基因克隆和 分子标记辅助抗病育种奠定基础 。