NaCl胁迫下流苏的电阻抗与电导分析
盐胁迫下植物激素水平的研究进展

盐胁迫下植物激素水平的研究进展张丽;罗孝明;蒙辉;袁远国【摘要】植物激素在一定程度上能够缓解盐胁迫对植物的伤害.结合盐胁迫对植物生长的影响,从盐胁迫下植物内源激素以及外源激素抵御盐胁迫的机理等方面作了简要分析,为选育抗盐品种提供参考.【期刊名称】《蔬菜》【年(卷),期】2017(000)003【总页数】4页(P29-32)【关键词】盐胁迫;植物激素;研究【作者】张丽;罗孝明;蒙辉;袁远国【作者单位】贵州省园艺研究所,贵州贵阳550006;贵州省园艺研究所,贵州贵阳550006;从江县农业局,贵州兴义557400;贵州省园艺研究所,贵州贵阳550006【正文语种】中文土壤盐渍化已经成为制约农业生产和生态发展的重要因素,普遍存在于世界各地。
我国的土壤盐渍化面积约3.6×107 hm2[1],并且随着生态环境的变化和不适宜的施肥方式导致的土壤盐渍化面积逐渐加大。
土壤盐渍化严重影响蔬菜的产量、品质以及经济效益,已成为农业生产的限制因素。
目前土壤盐渍化问题已经引起广泛关注,主要研究集中在抗盐品种的选育、植物激素处理、无土栽培等方面。
本文针对盐胁迫条件下植物激素水平的变化情况进行综述,以期为今后通过调节激素水平进行抗盐生理生化研究提供参考,有助于加快改良和恢复盐渍化土壤,使盐碱地块得以充分利用[2]。
1 盐胁迫对植物的伤害1.1 盐胁迫引起的膜质过氧化植物细胞质膜是具有选择吸收性的半透膜,对维护细胞内部环境和进行物质交换、能量和信息传递具有重要的作用。
细胞膜通常是最先感受胁迫伤害的结构[3]。
盐胁迫引起植物体内活性氧(ROS)的积累,ROS具有较强的氧化能力,能够破坏生物大分子的结构和性能,如引起酶失活、蛋白质降解、核酸结构破坏、碳水化合物分解等。
李斌等[4]研究表明,75 mmol/L NaCl胁迫使黄瓜幼苗叶片超氧阴离子(O2.-)产生速率、过氧化氢(H2O2)和丙二醛(MDA)含量以及电解质渗透率迅速升高,细胞膜受损,黄瓜幼苗生长受到抑制。
NaCl和Na2SO4胁迫对香樟幼苗生理特性的影响

。 - e r o d s e l g fCi n m on r a f2 y a l e d i s o n a r n c mp o a we e d t r i e . Th e u t h w e h twih t e e — n u h r r ee m n d e r s ls s o d t a t h n
dint e s,t e a tv te fp ot c i n y ms t e t d by Na r ghe ha ho et e t d by Na h c i ii so r e tvee z e r a e C1we e hi rt n t s r a e 2 SOd .The r s ls i ia e h tC. C T pho a s e i s p r o m a e o e a pt iiis u e a t s r s ,a a t e u t nd c t d t a (f Jl r e dl ng e f r nc d s m da ab lte nd r s l t e s nd s l
浓度 条 件 下 , 。 O NaS 。处 理 对 香 樟 的 伤 害 更 大 。
关 键 词 : 樟 ; 胁 迫 ; 盐 性 香 盐 耐
中图分 类号 :7 20 1 ¥ 9 . 3 .
文献 标 志码 : A
文 章 编 号 :0 17 6 ( 0 10 —0 00 1 0 —4 1 2 1 ) 60 3 — 5
西 北 林 学 院 学 报 2 1 。2 ( ) 0 3 O 1 6 6 :3 ~
J u n l f No t we tF r s r ie st o r a rh s o e ty Un v r i o y
Na 和 NaS 胁 迫 对 香 樟 幼 苗 生 理 特 性 的 影 响 C1 2 O4
NaCl胁迫对八棱海棠和山定子电导率和抗氧化酶活性的影响

En z y me S y s t e m i n Ma l u s r o b u s t a a n d Ma l u s b a c c a t a
L U Xi a o—y a n ,L V Xi n —mi n ,J I N P E NG ,RU S i —b o ,HU Ha i —f a n g ,Z HANG Do n g—y a
t i g a t e d a t f e r d i f f e r e n t c o n c e n t r a t i o n s o f N a C 1 t r e a t m e n t . 【 R e s u l t 】 O u r d a t a s h o w d t h a t t h e e l e c t r i c a l c o n d u c t i v i t y
用。
关键词 : N a C 1 ;八棱 海棠 ;山定子 ;电导率 ; 抗 氧化酶
中图 分 类 号 : ¥ 6 6 1 . 1 文献标识码 : A 文章编号 : 1 0 0 1— 4 3 3 0 ( 2 0 1 4 ) 0 2— 0 3 1 1— 0 7
Ef f e c t s o f Na Cl S t r e s s o n El e c t r i c a l Co nd u c t i v i t y a nd An t i o x i da nt
o f Ma l u s r o b st n a a n d Mal u s b a c c a t a p r e s e n t e d t h e i n c r e a s i n g t e nd e n c y wi t h t h e a d d i n g o f t h e c o nc e n t r a t i o n o f
NaCl胁迫对5种绿化植物幼苗生长和生理指标的影响及耐盐性综合评价

性 。通过主成分分析 和隶属函数法获得的 5种植物 的综合 评价值 D及 C。 均表 明 田菁和海 滨木槿对 N C 的耐性 a1
较强 , 其次为紫穗槐和决 明 , 光叶决明对 N C 的耐性最 差。 a1 关键词 : 化植 物 பைடு நூலகம் a 1 绿 N C 胁迫 ; 耐盐性 ;隶属函数法 ; 综合 评价 中图分类号 :Q 4 .8 7 86 0 9 5 7 ;¥2 .. 3 文献标志码 : A 文章编号 :17 — 8 5 2 1 ) 2 0 3 — 8 6 4 7 9 ( 0 1 0 - 0 5 0
6.8 a d 0 n 1 mg ・g b a c in 1 ri ai n y f n to a irg to me h d. An h tl r n e o v l n s o Na wa to d t e oe a c f f e p a t t Cl i s i e tfe n o r h nsv l v l ae y c rea in a a y i t d a d p i cp lc mpo n n lss d n iid a d c mp e e iey e au td b o r l t n lssmeho n rn i a o o ne ta ay i
cnai R t) o. anb a( e .Pi] n z r 幼苗 的 3 个生长指标和 6 个生理 指标 的变化进行 了测 定和 分析 , 采用 相关性分 析 、 主成分
分 析( C 与隶 属函数法相结合 的方法对 5种植 物对 N C 的耐性进 行 了鉴定 和综 合评 价 , 利用 回归 分析确定 P A) a1 并 了 5种植物 的生 长临界 N C 浓度 ( 。 。结果表 明 : 2~1 g・ N C 胁迫条件 下 , a1 ) 在 0m g a 1 5种植 物幼 苗的株 高生长 量和全株 干质量 总体上小于对照 , 总体 上随着 N C 质量浓度 的提高而降低。随 N C 质量浓度 的提高 , 且 a1 a1 5种植物 幼苗 的相对 电导 率 、 叶绿素含量 、 丙二醛含量 、 脯氨酸含量 、 溶性糖含量和 S D活性均受到不 同程度 的影 响 , 明 可 O 说
NaCl胁迫对马蔺生长及生理生化指标的影响

Efe t fNaCIsr s n o h a hy i l g c lbi c mia n x so rs l ta va .c ne ss fcso t e so gr wt nd p so o ia ・ o he c li de e fI i ace r hi n i
,
ZHANG n . u n,HUANG u—h n Mi g x a S ze ①
Abs r c :Ta i 1 ta t kng /2Ho g a d s l i n a ba i u t e s l i n, e fc so a ln out s o sc c l out ur o f t f1,2,3,4,6,8 a d 1 e n 0 g ・ L- 。Na sr s o s me r wt a d h soo i a. ic e c l n e e o rs a ta C1 te s n o g o h n p y il gc 1bo h mia i d x s f I i l ce Pa1 v r l. a .
wi ii g o C1 o c n r to t rsn fNa c n e ta in,h ihta d r o e t h e g n o t lngh,d y we g t o b v - run p r , r o n r ih s f a o e g o d a t o t a d wh l ln fI a ta v r hie ss s e ln e r a e g a u l oe pa to .1 ce a .c n n i e d i g d c e s r d al y.b th i h n r i h so b v — u eg ta d d y weg t fa o e
高, 马蔺幼苗 的株高 和根 长 、 地上部分和根及全株干质 量逐渐减 小 , 在 1g・ N C 胁 迫条件 下 , 高和 地上部 但 L a1 株 分及全株干质量均显著高于对照 。随 N C 质量浓 度 的提 高和胁 迫时 问的延长 , a1 马蔺 幼苗 叶片的相对 电导 率和丙 二醛 ( A) MD 含量 总体 上呈逐渐升高的趋势 ; 中 , 1~ L a 1 迫处理 7或 1 , 对电导率 有小 幅增 加 其 用 4g・ N C 胁 4d 相
NaCl和Na2CO3胁迫对黑麦草种子萌发及幼苗生理特性的影响

NaCl和Na2CO3胁迫对黑麦草种子萌发及幼苗生理特性的影响摘要:以黑麦草(lolium perenne)为试验材料,分别进行nacl (50、100、150、200 mmol/l)和na2co3(25、50、75、100 mmol/l)不同浓度胁迫处理,以加蒸馏水处理作对照,处理后研究nacl和na2co3胁迫下对黑麦草种子萌发、叶片相对电导率和丙二醛(mda)含量的影响。
结果表明:①在nacl和na2co3处理下,当na+浓度≤50 mmol/l时,对发芽率和mda含量没有显著影响,随着浓度的增加,特别是na+≥150 mmol/l时,黑麦草种子各项萌发生理指标和mda含量与对照相比均表现显著变化。
②就nacl和na2co3胁迫相比较,在相同na+浓度下,处于na2co3胁迫下的黑麦草种子幼苗叶片相对电导率及mda含量均明显高于nacl胁迫。
这说明nacl和na2co3胁迫均对黑麦草种子和幼苗造成了不同程度的伤害,但在相同na+浓度下,na2co3胁迫伤害程度大于nacl胁迫,因此说明黑麦草种子和幼苗对nacl的忍耐程度高于na2co3。
关键词:盐胁迫;黑麦草(lolium perenne);萌发;相对电导率;mda中图分类号:s543+.6 文献标识码:a 文章编号:0439-8114(2013)07-1613-03黑麦草(lolium perenne)品种caddieshack是冷季型草坪草中应用最广泛的草种之一[1]。
本试验采用人工控制盐分浓度的方法,设计了中性盐和碱性盐胁迫下对黑麦草种子的萌发特性及萌发过程中伴随的生理特性的影响,对其耐盐性作定量分析,旨在初步评价黑麦草的耐盐性,为黑麦草的种植推广、抗性品种的筛选和育苗工作提供理论依据,从而为更加合理充分利用盐碱地奠定基础。
1 材料与方法1.1 试验材料黑麦草品种caddieshack由江苏枫景花卉苗木种子基地提供。
1.2 试验方法1.2.1 种子发芽率、发芽势的测定将处理好的种子置于铺滤纸的培养皿中,分别用不同浓度的nacl和na2co3进行处理,以加蒸馏水处理为对照,nacl 胁迫分4个组:溶液分别为50、100、150、200 mmol/l(相应ph分别为7.10、7.12、7.14、7.15);na2co3胁迫分4个组:溶液分别为25、50、75、100 mmol/l(相应ph分别为10.32、10.64、10.83、10.93)。
nacl溶液 电导率计算

nacl溶液电导率计算摘要:1.钠离子和氯离子在水溶液中的电离2.电导率的定义和计算公式3.NACL 溶液的电导率计算实例4.影响电导率的因素正文:1.钠离子和氯离子在水溶液中的电离在水溶液中,许多化合物会解离成离子,这些离子可以导电。
例如,食盐(NaCl)在水溶液中会解离成钠离子(Na+)和氯离子(Cl-)。
这种解离过程增加了水溶液的电导率,使其能够导电。
2.电导率的定义和计算公式电导率(conductivity)是指在单位长度、单位截面积的导体中,通过的电流强度与电场强度之比。
其计算公式为:电导率(G)= 电流强度(I)/(电场强度(E)× 截面积(A))3.NACL 溶液的电导率计算实例假设我们有一个1 升(1000 毫升)的NACL 溶液,其中含有100 克(1 摩尔)的NaCl。
由于NaCl 在水溶液中完全电离,因此这个溶液中会有1 摩尔的Na+离子和1 摩尔的Cl-离子。
在25 摄氏度下,水的离子积(Kw)约为1.0×10^-14。
由于Na+和Cl-离子的浓度均为1 摩尔/升,我们可以计算出NACL 溶液中的电导率:G = (I / (E × A)) = (Kw × [Na+] × [Cl-]) / (L × F)= (1.0×10^-14 × 1 × 1) / (1 × 96485)≈ 1.04×10^-7 S/m因此,1 升1 摩尔/升的NACL 溶液的电导率约为1.04×10^-7 S/m。
4.影响电导率的因素电导率受到多种因素的影响,例如温度、溶液浓度、离子种类和溶液的压强等。
在一般情况下,随着溶液浓度的增加,电导率也会增加。
然而,当溶液浓度达到一定程度后,电导率可能会因为离子间的相互作用而减小。
盐胁迫对小麦幼苗生理生化特性的影响

盐胁迫对小麦幼苗生理生化特性的影响张军;王新军;于浩世【摘要】押为了明确盐胁迫对小麦幼苗生理生化特性的影响,采用水培方法,以兰黑粒小麦和小偃15小麦为材料,对其在不同浓度的NaCl处理下叶片中超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、丙二醛(MDA)含量、相对电导率、脯氨酸含量和可溶性蛋白质含量等生理指标进行测定。
结果表明:随着盐浓度的增加,兰黑粒小麦的SOD和POD活性先增后减,而小偃15小麦的SOD 和POD活性呈下降趋势;两种小麦的MDA含量、相对电导率和脯氨酸含量均随盐浓度的增加显著增大;可溶性蛋白质在2种小麦的变化趋势不尽一致。
2个供试品种均有一定的抗盐能力,兰黑粒小麦抗盐性相对较强。
%To study the effects of NaCl stress of wheat on its physiological and biochemical characteristics, Lanhei wheat and Xiaoyan 15 were taken as tested materials. The superoxide dismutase (SOD) activity, peroxidase (POD) activity, MDA content, relative conductivity, the proline content and the soluble protein content were investigated after different treatments. The results showed that under NaCl stress, compared with the each control, the activities of SOD and POD of Lanhei showed the trend of from increasing to decreasing, while the Xiaoyan15 showed the decreased; MDA content, relative conductivity, the proline content increased significantly as the stress decreased; whereas the trend of soluble protein was different between the 2 cultivars. Both of them had resistance to salt, and Lanhei wheat was a relatively higher salt-tolerant wheat.【期刊名称】《商洛学院学报》【年(卷),期】2015(000)004【总页数】4页(P59-62)【关键词】盐胁迫;兰黑粒小麦;小偃15小麦【作者】张军;王新军;于浩世【作者单位】商洛学院生物医药与食品工程学院/秦岭植物良种繁育中心,陕西商洛 726000;商洛学院生物医药与食品工程学院/秦岭植物良种繁育中心,陕西商洛726000;商洛学院生物医药与食品工程学院/秦岭植物良种繁育中心,陕西商洛726000【正文语种】中文【中图分类】S512作物产量的形成受多种因素的影响,其中干旱和盐碱造成的减产高达40%以上[1]。
植物细胞在盐胁迫下的膜透性研究

植物细胞在盐胁迫下的膜透性研究摘要:改善已经盐碱化的土壤的一种行之有效的方法是筛选抗盐碱,耐干旱的植物进行栽植。
作为抵御盐离子侵入细胞的第一道屏障,细胞膜的膜透性至关重要。
当植物处于盐胁迫的条件下时,其细胞内生理活动会受到盐离子的影响而发生改变,进而影响细胞膜的稳定性。
本文选取沙棘作为研究对象,首先对比分析了氯化钠和硫酸钠两种盐类的盐胁迫对样品细胞内保护酶含量的影响,分析了沙棘植物细胞在盐胁迫条件下的膜透性。
结果表明盐胁迫会使植物细胞内保护酶含量下降,从而导致植物细胞的膜透性增加,同时氯化钠盐胁迫对膜透性影响的程度显著强于硫酸钠。
关键词:植物细胞膜;盐胁迫;膜透性;保护酶;1引言随着近年来人类对耕作土壤的过度开发,许多地区的土壤已经出现了盐碱化的现象,这一问题直接影响到农作物的产量,对于人类发展的影响不可忽略[1]。
对于已经发生盐碱化的土壤,筛选出抗盐碱,耐干旱的植物进行栽植是一种行之有效的,改善盐碱化的方法。
在盐碱土壤中,植物所面临不利条件主要为盐胁迫,过高的盐浓度直接影响植物细胞内部的生理活动,进而影响植物的存活率。
作为抵御盐离子侵入细胞的第一道屏障,细胞膜的膜透性研究引起了学者的广泛关注[2]。
植物细胞膜透性对于植物抵抗过高盐浓度的伤害起到了至关重要的作用。
本文选取了沙棘作为研究对象,研究了植物细胞在盐胁迫下的膜透性,为筛选耐盐碱,抗干旱植物提供了依据。
2实验方法2.1 实验材料本文选用沙棘作为植物样本进行研究,进行实验的土壤为苗圃中的地表土,将土壤进行过筛后加入沙子,土壤和沙子的质量比为3:1。
选用的土壤中各种离子的含量如表1所示:表1 实验土壤中各种离子的含量2.2 实验过程将植物置于木桶中进行栽植,同种土壤质量为11kg,将两种盐类划分为四个浓度,分别为1.5,4.5,8.5,12.5g/kg,每种浓度的盐均分为三份相隔5d依次加入,盐胁迫20d后取样测试。
2.3 指标测试本实验所设计到的PDO (过氧化物酶),CAT (过氧化氢酶),膜透性分别采用愈创木酚法,紫外吸收法法和电导法进行测定3 结果分析图1 不同种类盐胁迫下植物细胞中保护酶含量的变化植物细胞中含有的保护酶可以将细胞内产生的超氧自由基和过氧化物消除,同时防止或减少羟基自由基的形成,有利于维持植物细胞膜的稳定存在。
玉米苗期对NaCl胁迫的响应与耐盐性调控机理的研究

玉米苗期对NaCl胁迫的响应与耐盐性调控机理的研究本研究对不同浓度NaCl胁迫下玉米幼苗的干重、渗透调节物质含量、保护酶活性、各器官离子的吸收与运转等生理生化特性进行研究,分析并探讨了盐胁迫对玉米幼苗的伤害及其耐盐机制,为进一步在分子水平上深入研究玉米耐盐性奠定了基础。
同时,在盐胁迫下,施用适量的外源物质钙、钾、磷、硅和NO供体硝普钠(sodium nitroprussideSNP),分析了其对玉米幼苗生理生化特性的影响,探讨了外源物质提高玉米耐盐性的作用机理,为提高植物的耐盐性提供一定的理论依据。
主要研究结果如下:1盐胁迫对玉米幼苗的伤害及其耐盐机制盐分胁迫包括渗透胁迫和离子胁迫,渗透胁迫导致吸水困难,离子胁迫造成生物膜破坏和生理紊乱。
盐胁迫对玉米幼苗的伤害及其耐盐机理主要有以下几个方面:1.1玉米幼苗具有一定的耐盐性,50mmol·L<sup>-1</sup>NaCl胁迫下,玉米幼苗干重略有增加,主要是根系干重显著增加。
随着NaCl浓度增加玉米幼苗各器官干重下降,干重下降较大的器官是生长叶和成熟叶叶片。
随NaCl浓度增加,各器官的含水量降低,含水量变化幅度为:根系>生长叶>成熟叶叶片>成熟叶叶鞘。
1.2 NaCl胁迫导致叶绿素含量降低,其中叶绿素a降低的幅度小于叶绿素b。
盐胁迫下玉米幼苗净光合速率、蒸腾速率、胞间CO<sub>2</sub>浓度、气孔导度降低,而水分利用率和气孔限制值上升,表明盐胁迫使玉米幼苗光合速率降低主要是由气孔因素引起的。
1.3 NaCl胁迫导致玉米幼苗细胞膜受到伤害,随NaCl浓度增加,叶片的电解质渗漏率增加,叶片和根系中MDA含量增加。
1.4 NaCl胁迫下,玉米幼苗叶片和根系中可溶性糖、可溶性蛋白、游离氨基酸和脯氨酸含量均增加,但叶片增加幅度大于根系。
1.5玉米幼苗在50 mmol·L<sup>-1</sup>、100mmol·L<sup>-1</sup>NaCl 胁迫下,叶片和根系中SOD、CAT、POD活性均增加。
200 mmoL/L NaCl胁迫对盐芥Na+含量的影响

200 mmoL/L NaCl胁迫对盐芥Na+含量的影响摘要以耐盐模式植物盐芥为试材,分别测定了200 mmoL/L盐胁迫下其地上和地下部分Na+含量,从生理生化水平上探讨了盐芥的耐盐机理,为研究盐芥的耐盐特性奠定基础。
关键词NaCl胁迫;盐芥;耐盐性;影响土壤盐渍化已成为影响作物生长发育、产量和品质的一个重要因素,严重影响农业生产。
预计到2050年全球将有50%的耕地发生不同程度的盐渍化[1]。
盐芥(Thellungiella halophila)是拟南芥的近缘物种,是生长周期短、基因组较小的真盐生植物;并与拟南芥的遗传特性、生活特性相似,其基因组与拟南芥有很高的相似度[2],且具有比拟南芥更强的胁迫耐受性,已成为一种重要的非生物胁迫的模式材料。
该试验是测量盐芥根、叶在200 mmoL/L盐胁迫下Na+含量的变化,为研究盐芥的耐盐性提供借鉴。
1材料与方法1.1试验材料供试材料为盐芥(Thellungiella halophila Shandong eco-type),原种采自山东东营黄河入海口,试验所需种子为自繁。
试验用NaCl溶液用Hongland营养液配制。
1.2试验方法1.2.1盐芥种子处理。
在灭菌1.5 mL EP管中加入100~200粒盐芥种子;加70%酒精1 mL,处理1 min;吸出乙醇,加入1%NaClO溶液1 mL,涡旋10~15 min;倒出NaClO,用1 mL灭菌的DDW洗5~6遍,每次洗1~2 min;用少量灭菌的DDW重悬;吸取种子种于平皿中,用parafilm封口,放于4 ℃冰箱中7 d。
1.2.2盐芥种植。
将处理的种子放于温室中培养,至幼苗长出2片真叶时转移到水培条件下种植,用Hongland营养液浇灌。
生长温度为25 ℃,光照16 h,黑暗8 h。
1.2.3盐芥的盐处理。
盐芥生长90 d后,用200 mmoL/L NaCl溶液处理,处理的时间分别是0、0.5、1.5、3.0、5.0、9.0、12.0、24.0 h。
NaCl胁迫对葎草部分生理特性的影响

刘志华, 时丽冉 , 马 光. a 1 N C 胁迫对棒 草部分 生理特性的影响[ ] J .江苏农 业科 学, 1 , ( ) 36— 4 2 24 4 : 0 0 4 37
N C 胁迫对蓓草部分生理特性 的影 响 a1
刘 志华 ,时丽冉 ,马 光
10m o LN C 浓 度 下 , A 含 量 上 升 幅 度 较 大 ( < 0 m l a 1 / MD P
0 0 ) 表明植物对较轻 的胁迫具有一定抗性 。 .5 ,
相对 电导率( C 是反映 细胞 膜透性 的重要指标 。由图 E) 1 可看出 , 随着 N C 浓度 的升 高 , 料幼苗 叶片 的质膜透 性 a1 材 越来越大 。当 N C 浓度大于 5 o L时 , a1 0mm l / 已显示出明显的
图1 不 同浓度N C 处理对幼苗叶片质膜透性 的影响 aI
2 2 不 同浓 度 N C 处理 对 幼 苗 叶 片 丙二 醛含 量 的 影 响 . a1
将蓓草种 子用 0 1 H C 消 毒 1 i , 馏水 冲洗 干 .% gI 0m n 蒸 净, 自来水浸泡 1 , 0h 再播种于洗净 的沙基花盆 中; 用蒸馏水 浇灌 , 培养温度为( 5- ) 自然光照 ; 2 4 ℃, - 2 待幼苗展开真 叶后 , 改用 1 2H aln / ogad营养液培 养 , 2d浇灌 1次 营养 液。幼 每 苗生长 2周后开始 用 0 5 、0 、5 、0 mo LN C 处理 , 、0 10 10 20m l a 1 / 各种浓度的 N C 溶液 均用 营养液配制。处理 2周后 , a1 测定 幼 苗 叶片质膜透性、 丙二醛含量、 可溶性 糖含量和游离脯氨酸含 量 。每个处理 3个重复 , LD法进行差异显著性分析。 用 S
NaCl_胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响

第44卷第8期2024年4月生态学报ACTAECOLOGICASINICAVol.44,No.8Apr.,2024基金项目:新疆兵团人才发展专项(CZ00150102);国家自然科学基金项目(32360097)收稿日期:2023⁃06⁃16;㊀㊀网络出版日期:2024⁃01⁃29∗通讯作者Correspondingauthor.E⁃mail:Luzhuo@shzu.edu.cnDOI:10.20103/j.stxb.202306161284林晓华,卓一林,柳丽芳,施翔,卓露.NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响.生态学报,2024,44(8):3483⁃3491.LinXH,ZhuoYL,LiuLF,ShiX,ZhuoL.MembranestabilityofthedesertmossSyntrichiacaninervisMitt.duringNaClstress.ActaEcologicaSinica,2024,44(8):3483⁃3491.NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响林晓华,卓一林,柳丽芳,施㊀翔,卓㊀露∗石河子大学生命科学学院新疆植物药资源利用教育部重点实验室绿洲城镇与山盆系统生态兵团重点实验室,石河子㊀832000摘要:苔藓结皮作为生物土壤结皮演替的最高阶段和生物量的最主要贡献者,对维持荒漠地表稳定和改善微环境具有重要作用㊂齿肋赤藓SyntrichiacaninervisMitt.作为荒漠苔藓结皮的优势种,是不可多得的开展抗逆性资源研究的重要材料㊂研究对齿肋赤藓进行不同NaCl浓度(0㊁200㊁300和400mmol/L)和不同胁迫时间(处理1d㊁2d㊁3d㊁5d㊁7d和14d)处理,主要从结构特性和生理特性对其膜结构进行研究㊂结果表明:齿肋赤藓在中浓度NaCl胁迫(200mmol/L和300mmol/L)下均未观察到电导率显著变化,而随着胁迫时间的增加,细胞超微结构发生明显变化,特别在高浓度NaCl胁迫(400mmol/L)下,细胞发生严重的质壁分离,叶肉细胞部分发生死亡,且出现大量泡状结构,而叶绿体大部分解体,片层结构消失,淀粉粒数量增加且体积变大,但这一过程中没有膜损坏的直接证据㊂据此关于齿肋赤藓耐受NaCl胁迫的膜结构稳定性有2种可能推测:(1)齿肋赤藓具有特殊的形态和解剖特征,例如:细胞壁较厚,且存在角隅加厚现象,使其能够在恶劣的盐胁迫环境下生存,并保留细胞结构完好无损;(2)在NaCl胁迫过程中,快速且持续的增加可溶性糖( 组成性 保护物质)和游离脯氨酸,有助于提高细胞膜的稳定性㊂荒漠苔藓齿肋赤藓虽然没有被定义为盐生植物,但以上研究结果表明,齿肋赤藓可以作为一种新的模式物种展开关于盐胁迫方面的研究㊂关键词:盐胁迫;沙漠植物;齿肋赤藓;生理指标;透射电镜MembranestabilityofthedesertmossSyntrichiacaninervisMitt.duringNaClstress㊀LINXiaohua,ZHUOYilin,LIULifang,SHIXiang,ZHUOLu∗XinjiangProductionandConstructionCorpsKeyLaboratoryofOasisTownandMountain⁃basinSystemEcology,MinistryofEducationKeyLaboratoryofXinjiangPhytomedicineResourceUtilization,SchoolofLifeSciences,ShiheziUniversity,Shihezi832000,ChinaAbstract:ThemossSyntrichiacaninervisMitt.isoneofthedominantspeciesinthebiologicalsoilcrustsofdesert.IthaslongbeenthefocusofscientificresearchowingtotheecologicalvalueofS.caninervishasspecialsignificanceforresearchingonitssaltresistance.WeinvestigatedthechangesinmembranestructuresofS.caninervis,andbiochemicalandphysiologicalparametersundervaryingsaltconcentrationgradientsofNaCl(0,200,300,and400mmol/L)for1,2,3,5,7and14daysinthisstudy.Theresultsshowedthatittolerated200mmol/LNaClquitewell.Electronmicrographsofthefinestructureofthecellswereobtained,andnosignificantchangesinelectricalconductivitywereobservedatlowsaltconcentrations.However,majorchangesincellultrastructurewereobservedat300and400mmol/L.Additionally,osmoregulatorysubstancesandchlorophyllcontentreducedwithincreasingsaltconcentrationandtreatmenttime.Inconclusion,S.caninervishasspecialmorphologicalcharacteristics(e.g.thecellwall),whichcanenableittosurviveinharshsaltstressenvironmentsandpreservethecellstructure,whileremainingintactafterthereleaseofNaClstress;(2)DuringNaClstress,arapidandsustainedincreaseinsolublesugars( constitutive protectivesubstances)andfreeproline4843㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀contributestomembrane(Forexample,thecellwallisthickandthereiscornerthickeningphenomenon),enablingittosurviveinharshsaltstressenvironmentsandpreservethecellstructure,whileremainingintactafterthereleaseofNaClstress.Althoughitisnotecologicallyclassifiedasahalophyte,ourresultssuggestthatitcouldbeanovelplantmodelspeciestoexplorevariousaspectsofsaltstress.KeyWords:saltstress;desertmoss;SyntrichiacaninervisMitt.;physiologicalindex;transmissionelectronmicroscopy生物土壤结皮(BSCs)广布于古尔班通古特沙漠[1]㊁Negev沙漠[2]㊁Namib沙漠[3]和Mojave沙漠[4]等固定与半固定沙漠,在改变土壤肥力㊁提高土壤抗风蚀能力㊁改善荒漠生态环境等方面发挥重要的积极作用[1,5]㊂藓类结皮作为生物土壤结皮演替的最高阶段和生物量的最主要贡献者,能够在相对疏松的砂质中生长良好,在地下形成具有固着作用的假根,对维持荒漠地表稳定和改善微环境具有很强的重要作用[6]㊂在干旱地区生态系统中盐分与水分一样,对植物的生长发育㊁生存分布以及生理生态过程具有重要的影响㊂大量研究表明,生物结皮对盐分具有一定的耐受性,且不同的种类的生物结皮耐盐机理不同[7]㊂苔藓植物是最低等的高等植物,属于水生到陆生的一种过渡类型,陆地高浓度的氧含量㊁强烈的大气辐射以及剧烈变化的温度,催生了该类植物具有其他普通陆生植物无法比拟的综合抗逆性,分布在极端沙漠环境中的耐干苔藓齿肋赤藓SyntrichiacaninervisMitt.最为显著㊂目前,关于齿肋赤藓的研究主要集中在生态分布[8 9]㊁生态学作用[10 14],耐干胁迫的形态及生理响应[15 19],以及基因克隆与分子响应机制[20 22]等方面㊂研究区古尔班通古特沙漠降水量少,蒸发量高,是造成苔藓结皮遭受盐胁迫的重要原因,研究区土壤表层的高盐度又是影响齿肋赤藓生存的主要环境因素之一㊂然而,对齿肋赤藓耐盐方面的研究知之甚少,仅有的报道集中在表型结构方面,从膜结构稳定性角度探讨其耐盐机制的研究实属空白[23 25]㊂盐分的积累抑制了植物的生长,降低了其吸收水分和养分的能力,导致渗透胁迫[26]㊂高盐胁迫会破坏植物细胞的渗透和离子稳态以及光合作用,消耗细胞能量并导致氧化还原失衡[27]㊂为了适应盐胁迫,盐生植物进化出了各种形态㊁生理和生化的适应机制,使其能够在盐分环境中生存㊁繁殖㊁分布和完成生命周期[28 30]㊂根据植物在盐生基质上生长的能力可分为三种主要分类:专性盐生植物,仅在盐水基质中生长;兼性盐生植物,可以在含盐基质上生存,但在无盐或低盐浓度基质上生长发育更好;和生境无关的盐生植物,它们可以在含盐基质和无盐基质上成功生长[31]㊂然而,苔藓植物通常被认为是非盐生植物,但却可以在含盐量较高的机制上生存[32],比如:南极黄丝瓜藓PohlianutansLindb.具有独特的生理和遗传特征,使其能够高度适应极地高盐环境[33]㊂2005年,Zechmeister研究者[34]在奥地利潘诺尼的盐碱地调查发现64类物种,且全部为苔藓植物,其中包括:尖叶梨蒴藓Entosthodonhungaricus㊁小丛藓PottiaheimiiLindb等㊂大量研究表明,苔藓植物由于缺乏特殊的盐腺和盐囊泡等输导组织,使其耐盐生存策略不同于维管植物[35 36]㊂同时,苔藓植物具有生长快㊁易于培养㊁配子体相对发达,且具有超强的耐干㊁耐热㊁耐盐性等优势,暗示着该种将是另一个极具潜力的㊁可作为模式种进行耐盐机制研究的好材料[37]㊂因此,本文选取了古尔班通古特沙漠生物结皮藓类的优势种齿肋赤藓为研究对象,模拟不同盐浓度和胁迫时间,试图回答以下科学问题:1㊁荒漠苔藓齿肋赤藓是否具有某些特殊的形态结构或解剖特征适应不同强度的盐胁迫?2㊁在受到不同程度盐胁迫时,齿肋赤藓是否会产生某些 组成性 保护物质以维持膜的稳定性?从而探究齿肋赤藓膜结构稳定性在盐环境变化过程中的适应机制㊂对荒漠苔藓植物的耐盐机理进行深入解析,可为耐盐转基因农作物改良提供重要的理论依据和基因资源,也是目前植物种质资源开发与利用的主要趋势㊂1㊀材料与方法1.1㊀研究区概况古尔班通过特沙漠(44.119ʎ 46.209ʎN,84.319ʎ 90.009ʎE)位于新疆北部准噶尔盆地中部,是我国最大的固定㊁半固定沙漠,面积约为48800km2㊂年平均降水量约为70 150mm,年均蒸发量大于2000mm,年平均温度为5 7ħ,月温度在-20至30ħ之间,而夏季沙层表面瞬间极端高温达到80ħ以上㊂该沙漠中有梭梭(Haloxylonammdendro)㊁白梭梭(Haloxylonpersicum)㊁淡枝沙拐枣(Calligonumleucocladum)㊁双穗麻黄(Ephedradistachya)等小半乔木和灌木,并伴有短命和类短命植物的分布,另外,发育良好的生物土壤结皮以斑块状分布其中㊂1.2㊀实验材料和处理2022年4月,从古尔班通古特沙漠南缘腹地(44ʎ36N,88ʎ15E),利用五点取样法用自制的聚氯乙烯(PVC)取样器(直径=10cm,高度=2cm)采集一系生长良好,长势一致的齿肋赤藓生物结皮,带回实验室的材料在黑暗或昏暗条件下((20ʃ1)ħ,约20%的相对湿度)缓慢风干,并在两周内使用㊂实验前,利用水湿法采集进行消毒处理,然后放置于培养箱(23ħ白天/12ħ晚上)培养1周,从中选取高度约为1.5cm的单株植物个体作为实验材料㊂将选定的若干实验单株材料在流动的水下冲洗10min,去除砂及杂质,将干净的样品放在含有40mL双蒸馏水的培养皿(内径10cm)中,光照下((20ʃ1)ħ,80μmolm-2s-1)全覆水24h,直到全部恢复绿色㊂以5g齿肋赤藓植株为1个样本放置于培养皿中,每个处理有5个重复,向每个培养皿中加入7mLNaCl溶液,浓度分别为0㊁200㊁300和400mmol/L的进行处理㊂统计盐胁迫开始后1d㊁2d㊁3d㊁7d和14d样本的生长存活情况㊂1.3㊀相对电导率不同盐胁迫下齿肋赤藓膜渗透率利用DDS⁃307A数字电导仪(中国有限公司精密科学仪器有限公司)进行测量㊂将0.1g不同处理的样品在10mL蒸馏水中浸泡样品,室温放置12h,用电导仪将水和渗滤液的电导率测量,记为Ci,将试管盖上盖子,材料煮沸25min,冷却至室温,进行电导率的测量,记为Ct㊂相对电导率表示相对电解质泄漏L(%)或细胞膜的相对渗透率,并根据以下方程计算:L(%)=Ci-C0Ct-C0ˑ100%(1)其中,L是相对电解质泄漏,C0是蒸馏水中的基线电导,Ci是H2O和浸出物的电导,Ct是总电导㊂1.4㊀透射电镜观察选取不同盐浓度处理3d和14d的齿肋赤藓(茎尖约为0.5cm),每个处理选择5株长势一致的材料用于电镜样品的制备㊂将样品置于4%的戊二醛固定液中固定24h左右,然后用磷酸缓冲液(pH为7.2的磷酸盐缓冲液(PBS))进行洗涤,接着采用1%锇酸固定约为1.5h,再分别用浓度梯度为30%㊁50%㊁70%㊁80%㊁90%㊁95%和100%的乙醇进行逐级脱水处理,且每个梯度不少于20min㊂脱水处理完成后,依次用1ʒ1与4ʒ1的丙酮/乙醇,纯丙酮浸泡30min,用1ʒ1体积的丙酮/环氧树脂固定1h,1ʒ4体积的丙酮/环氧树脂固定3h,纯环氧树脂进行过夜(12h),Epon⁃812渗透包埋;在烘箱中逐渐升温至60ħ,放置12h;利用LKB⁃8800型超薄切片机进行切片,经醋酸铀-柠檬酸铅双重染色,用EM⁃1200EX透射电子显微(日本电子JEOL)镜进行后期扫描拍照㊂1.5㊀生理生化指标本研究选定的脯氨酸和可溶性糖2种主要的渗透调节物质,所用的方法分别是酸性茚三酮法和考马斯亮蓝法,其吸光度结果是由Lamda35紫外分光光度计(PE,Waltham,美国)测量所得[38,39]㊂叶绿素含量采用常规分光光度法测定,以95%乙醇作为空白,分别在649mm和665mm波长测定其吸光度,并计算叶绿素b含量和叶绿素a含量㊂提取物的总叶绿素浓度(μg/mL)使用描述的方程[40]计算㊂Ca+b=6.4534A665-17.232A649(2)其中,Ca+b是叶绿素a和b,A665是波长在665mm,A649是波长在649mm㊂1.6㊀统计分析使用SPSS10.0版本和GraphPadPrism8.0对数据进行分析㊂假设Pɤ0.05,通过双因素方差分析5843㊀8期㊀㊀㊀林晓华㊀等:NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响㊀(ANOVA)检验样本之间平均值差异的显著性,并通过Duncan多重比较检验估计和比较不同盐胁迫处理的显著性水平㊂2㊀结果分析2.1㊀不同浓度NaCl处理对齿肋赤藓表型特征的影响利用光学显微镜观察发现,四种不同盐浓度处理下齿肋赤藓的表型特征如图1所示,存在显著性差异㊂在200mmol/L的NaCl浓度下,齿肋赤藓表型无明显变化,实验处理14d时存活率为(98.0ʃ0.51)%,而在300mmol/L的NaCl浓度下处理3d时,植物体叶片表现出轻微的黄色,最终超过(60.81ʃ1.29)%的植株个体存活(表1)㊂随着不同浓度NaCl处理天数的增加,植株个体受胁迫程度越明显,在高浓度(400mmol/LNaCl)的盐胁迫5d时,齿肋赤藓个体会发生组织烧伤(色素沉着丧失),而进一步长期暴露在高盐的条件下会发生个体的死亡㊂图1㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓的表型图(T1 T14处理1 14d)Fig.1㊀ChangesintheappearanceofthedesertmossSyntrichiacaninervisMitt.underdifferentNaClconcentrations(T1 T14Treatment1 14day)表1㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓存活率Table1㊀SurvivalofthedesertmossSyntrichiacaninervisMitt.after14daysofexposuretoNaClconcentrations盐浓度ConcentrationofNaCl0mmol/L200mmol/L300mmol/L400mmol/L生存率Survivalrate/%100ʃ0.0098.0ʃ0.5160.81ʃ1.299.93ʃ1.032.2㊀不同浓度NaCl处理对齿肋赤藓电导率的影响盐胁迫导致质膜的选择透性发生变化,溶质外渗,细胞的膜结构和功能受到损伤㊂从图2可知,处理前3d齿肋赤藓植株体的电导率较为稳定,不同浓度处理之间不存在显著性差异(P>0.05),说明细胞膜结构相对较完整㊂而随着胁迫浓度和时间的延长,相对电导率整体呈上升趋势,胁迫期间的相对电导率在0.23% 0.41%之间,在200mmol/L时相对电导率的上升幅度较小,而最大电导率值(0.41ʃ0.02)出现在高浓度400mmol/L处理第14d㊂2.3㊀不同浓度NaCl处理对齿肋赤藓生理指标的影响不同浓度盐胁迫处理条件下,齿肋赤藓植株总叶绿素含量之间存在显著性影响(P<0.05),随盐浓度的增加总叶绿素含量降低(图3)㊂NaCl处理7d时,300mmol/L条件下齿肋赤藓植株体总叶绿素含量与对照(5505ʃ194)mg/g干重相比,显著降低至(3503ʃ643)mg/g干重,而高盐400mmol/L时,总叶绿素含量(937ʃ353)mg/g干重与对照相比降低了5倍㊂如图4所示,可溶性糖和脯氨酸含量表现出相似的变化趋势,在不同NaCl处理下显著升高,200mmol/L㊁300mmol/L和400mmol/L的盐胁迫处理14d分别为对照组的1.49和3.74,2.08和5.67,2.48和6.29倍㊂6843㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀图2㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓的电导率结果图(T1 T14处理1 14d)Fig.2㊀ChangeofrelativemembranepermeabilityunderdifferentNaClconcentrationsandexposuredurationofSyntrichiacaninervisMitt(T1 T14Treatment1 14day)不同字母表示不同处理之间存在显著性差异(P<0.05)㊀图3㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓的叶绿素含量的影响(T1 T14处理1 14d)Fig.3㊀ChangeoftotalchlorophyllcontentunderdifferentNaClconcentrationsandexposuredurationofSyntrichiacaninervisMitt(T1 T21Treatment1 14day)2.4㊀不同浓度NaCl处理对齿肋赤藓细胞超微结构的影响透射电镜观察发现,在不同盐浓度胁迫下齿肋赤藓的细胞学结构存在显著性差异㊂未处理(图5)的细胞叶绿体形态完整,长轴平行于相邻的细胞壁,具有丰富片层的类囊体(图5)㊂而随着盐浓度的升高,特别是400mmol/LNaCl处理3d和14d(图5),叶绿体形态整体膨胀至外膜破损,发生严重解体现象;基粒片层由最开始的清晰有序逐渐变形,片层紊乱㊁扭曲出现间隙,至最后片层结构几乎不可见;叶绿体内淀粉粒逐渐模糊至不可见,脂质体逐渐模糊不清㊂细胞核结构随着盐浓度的增加,由清晰可见逐渐变化为核膜边缘模糊,核仁逐渐模糊直至消失,核质严重外溢,最终细胞核破裂解体㊂线粒体结构在300mmol/L浓度时,内膜结构随着胁迫时间增长,逐渐消失甚至观察不到,嵴由清晰可见到无法观察消失㊂细胞原生质膜的结构受到损伤,细胞内电解质外渗,细胞膜变形严重至破裂,原生质体持续收缩并发生质壁分离(黑色箭头所示),细胞间隙扩大,细胞壁由完整光滑发生形变扭曲至粗糙破裂,与对照和200mmol/LNaCl处理中细胞膜紧贴细胞壁形成鲜明对比㊂由此可知,在300mmol/LNaCl胁迫下,齿肋赤藓微观形态结构发生变化,而400mmol/L胁迫下发生明显变化㊂3㊀讨论新疆地处亚欧大陆腹地,蒸发量大而降水量少,促使荒漠地表的盐分含量大幅度增加,不仅引起严重的生态问题,而且对当地经济发展造成巨大影响[13]㊂盐胁迫能够诱导植物产生多种生化反应,当盐分积累到一定7843㊀8期㊀㊀㊀林晓华㊀等:NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响㊀图4㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓的可溶性糖和脯氨酸含量的影响Fig.4㊀ChangeofsolublesugarcontentandFreeprolinecontentunderdifferentNaClconcentrationsandexposuredurationofSyntrichiacaninervisMitt(T1 T14Treatment1 14day)图5㊀在不同盐浓度和胁迫时间处理下荒漠苔藓齿肋赤藓的细胞超微结构图Fig.5㊀TransmissionelectronmicroscopyinleafcellsofSyntrichiacaninervisMitt.underNaClstressandrecoveryN:细胞核;CW:细胞壁;Chl:叶绿体;黑色箭头指示质壁分离程度会对植物体产生毒害作用[41 42]㊂本文基于结构和生理特性对盐胁迫下耐干苔藓齿肋赤藓膜结构稳定性的研究发现:盐胁迫下齿肋赤藓能够保证细胞壁的完整性和细胞膜的相对稳定性㊂尽管随着盐浓度和胁迫时间的增加,细胞超微结构变化,发生质壁分离的现象,但却没有直接证据表明细胞膜遭到损伤;另外,盐胁迫下渗透调节物质(可溶性糖和脯氨酸含量)的积累为齿肋赤藓细胞膜的稳定性提供了物质基础,很大程度提高了齿肋赤藓的耐盐能力,该实验结果与最初推测相符合㊂叶绿素是高等植物重要的光合色素,可作为植物耐盐性鉴定的主要生理指标[43]㊂本研究结果发现,在高盐胁后齿肋赤藓植株体的表型发生了明显变化,特别是高盐400mmol/LNaCl胁迫处理第5d,齿肋赤藓植株体出现黄褐色(图1),结合生理指标叶绿素含量也发生了显著下降(图3)㊂大量文献报道,叶绿体对盐胁迫较敏感,盐胁迫能够诱导叶绿体中产生过量的活性氧(ROS),从而导致叶绿体被膜和类囊体膜的解体,叶绿体结构破坏[44 45]㊂本研究材料齿肋赤藓在低浓度NaCl处理(200mmol/L)时,细胞的超微结构基本上不受影响,与银叶真鲜BryumargenteumHedw.和波叶仙鹤藓AtrichumundulatumHedw.(耐受50和100mmol/LNaCl)相比[46],表现出较高的耐盐性㊂而300mmol/LNaCl胁迫3d后,齿肋赤藓的叶绿体开始受到不同程8843㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀度的损伤,透射电镜发现其叶绿体基粒片层出现紊乱,松散变形加剧,部分类囊体模糊不清;400mmol/L叶绿体发生解体现象,外膜破损溶解加剧,基粒片层出现膨胀或断裂,片层结构几乎不可见㊂从而推测,高盐(400mmol/L)胁迫严重破坏了齿肋赤藓叶绿素的结构和功能,明显抑制了叶绿素合成酶的活性,进而打破叶绿素合成与降解之间的动态平衡,这与刘卫国等的研究结果较为符合[24]㊂另外,其他高等植物,如芦苇(Phragmitesaustralis(Cav.)Trin.exSteud)㊁枸杞(LyciumchinenseMiller)等在受到盐胁迫后叶绿体发生了解体现象,出现了大量的淀粉粒和嗜锇颗粒,这些现象是高盐胁迫下植株体受损伤的重要证据,都与本研究的结果一致[47 48]㊂据此推测,齿肋赤藓通过细胞体内积累大量能量物质来抵御胁迫伤害,可能是其应对环境胁迫重要的保护机制㊂另外,渗透调节物质是植物响应逆境胁迫中重要的生理指标,对维持植物细胞的渗透压㊁调节植物体内活性氧的平衡具有极为重要的作用[49]㊂已有的研究表明,在非生物胁迫中耐干苔藓可以通过调节植物体自身渗透调节物质的含量,缓解胁迫造成的生理代谢不平衡,维持正常渗透压,从而保障细胞的正常生理功能[50]㊂其中,可溶性糖和脯氨酸作为植物重要的渗透调节物质,能够通过增加渗透调节物质的含量,减轻植物对逆境的抵御能力[51]㊂该结果在本研究中也得到证实,齿肋赤藓具有较为完善的渗透调节能力以适应盐胁迫㊂与对照相比,高盐处理导致齿肋赤藓可溶性糖和脯氨酸含量的增加,进而提高细胞液的浓度和细胞的渗透压平衡,维持植株体的含水量,防止细胞损伤,以此在齿肋赤藓耐受高盐胁迫过程中起着更加稳定的保护作用㊂植物的细胞膜是活细胞与环境之间的界面和屏障,植物遭受高温㊁干旱㊁盐渍等不良环境会促使细胞膜发生不同程度的损伤,可以利用电导率法对细胞膜透性的变化规律进行研究[11]㊂本研究发现处理初期(T1 T2),不同盐浓度齿肋赤藓植株体的相对电导率之间不存在显著性差异(P>0.05)㊂随着胁迫时间的延长,低浓度NaCl处理的材料与对照相比材料相对电导率差异不显著,而高盐300mmol/LNaCl以上质膜的稳定性降低,电解质发生外渗,相对电导率提高㊂说明,齿肋赤藓细胞膜透性受高盐胁迫的影响,但在短时间盐胁迫中仍然能够保持膜的相对稳定性㊂综上所述,不同盐浓度的变化显著影响了荒漠苔藓齿肋赤藓的生理生化特征㊂在受到高盐胁迫时,齿肋赤藓植株体叶绿素含量显著下降,显微结构遭到破坏,但在高盐胁迫初期,细胞膜的完整性较好,相对电导率无显著性差异㊂由此可知,齿肋赤藓具有一定的耐盐性,能够适应200mmol/L以下的盐胁迫,并能够通过维持细胞膜的稳定性以及调控渗透调节物质的含量缓解胁迫造成的损伤㊂本文以结构特性和生理特性为研究基础,深入探讨了齿肋赤藓可适应的临界盐度以及相应的生理生化指标,该研究结果可以为进一步深入探讨齿肋赤藓耐盐机制提供理论依据㊂参考文献(References):[1]㊀ZhangYM,WangHL,WangXQ,YangWK,ZhangDY.ThemicrostructureofmicrobioticcrustanditsinfluenceonwinderosionforasandysoilsurfaceintheGurbantunggutDesertofNorthwesternChina.Geoderma,2006,132(3/4):441⁃449.[2]㊀KidronGJ,HerrnstadtI,BarzilayE.TheroleofdewasamoisturesourceforsandmicrobioticcrustsintheNegevDesert,Israel.JournalofAridEnvironments,2002,52(4):517⁃533.[3]㊀RiosADL,Garrido⁃BenaventI,LimónA,CasonED,Maggs⁃KöllingG,CowanD,ValverdeA.Novellichen⁃dominatedhypolithiccommunitiesintheNamibDesert.MicrobialEcology,2022,83(4):1036⁃1048.[4]㊀StarkL,McLetchieN,MishlerB.SexexpressionandsexdimorphisminsporophyticpopulationsofthedesertmossSyntrichiacaninervis.PlantEcology,2001,157(2):183⁃194.[5]㊀BelnapJ,PhillipsSL,MillerME,BelnapJ,PhillipsSL,MillerME.Responseofdesertbiologicalsoilcruststoalterationsinprecipitationfrequency.Oecologia,2004,141(2):306⁃316.[6]㊀ColeC,StarkLR,BonineML,McLetchieDN.Transplantsurvivorshipofbryophytesoilcrustsinthemojavedesert.RestorationEcology,2010,18(2):198⁃205.[7]㊀许文文,赵燕翘,王楠,赵洋.人工蓝藻结皮对沙区表层土壤酶活性及其恢复速率的影响.生态学报,2023,43(7):2856⁃2864.[8〛㊀卓露,林晓华,薛山,梁玉青,张卓文,李鸿彬,WoodAndrew,张道远.植物耐干机制研究进展.华中农业大学学报,2023,42(5):9843㊀8期㊀㊀㊀林晓华㊀等:NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响㊀0943㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀28⁃34.[9]㊀ZhangYM,ChenJ,WangL,WangXQ,GuZH.ThespatialdistributionpatternsofbiologicalsoilcrustsintheGurbantunggutDesert,NorthernXinjiang,China.JournalofAridEnvironments,2007,68(4):599⁃610.[10]㊀WuN,ZhangYM,DowningA,AanderudZT,TaoY,WilliamsS.Rapidadjustmentofleafangleexplainshowthedesertmoss,Syntrichiacaninervis,copeswithmultipleresourcelimitationsduringrehydration.FunctionalPlantBiology,2014,41(2):168⁃177.[11]㊀WuN,ZhangYM,DowningA,ZhangJ,YangCH.MembranestabilityofthedesertmossSyntrichiacaninervisMitt.duringdesiccationandrehydration.JournalofBryology,2012,34(1):1⁃8.[12]㊀ZhuoL,LiangYQ,YangHL,LiXS,ZhangYH,ZhangYG,GuanKY,ZhangDY.ThermaltoleranceofdriedshootsofthemossBryumargenteum.JournalofThermalBiology,2020,89:102469.[13]㊀张元明,曹同,潘伯荣.新疆古尔班通古特沙漠南缘土壤结皮中苔藓植物的研究.西北植物学报,2002,22(1):18⁃23.[14]㊀张元明,曹同,潘伯荣.干旱与半干旱地区苔藓植物生态学研究综述.生态学报,2002,22(7):1129⁃1134.[15]㊀LiY,WangZB,XuTH,TuWF,LiuC,ZhangYM,YangCH.ReorganizationofphotosystemIIisinvolvedintherapidphotosyntheticrecoveryofdesertmossSyntrichiacaninervisuponrehydration.JournalofPlantPhysiology,2010,167(16):1390⁃1397.[16]㊀ZhengYP,XuM,ZhaoJC,ZhangBC,BeiSQ,HaoLH.MorphologicaladaptationstodroughtandreproductivestrategyoftheMossSyntrichiacaninervisintheGurbantunggutDesert,China.AridLandResearchandManagement,2011,25(2):116⁃127.[17]㊀ZhuoL,ZhangYG,LiXS,YangHL,GuanKY,WoodAJ,ZhangDY.DifferentialfragmentregenerationinSyntrichiacaninervisMitt.fromtheGurbantunggutDesertofChina.JournalofBryology,2018,40(3):265⁃270.[18]㊀魏美丽,张元明.生物结皮中齿肋赤藓叶片细胞显微与亚显微结构特征.中国沙漠,2009,29(3):6.[19]㊀魏美丽,张元明.脱水对生物结皮中齿肋赤藓光合色素含量和叶绿体结构的影响.中国沙漠,2010,30(6):1311⁃1318.[20]㊀YangRR,LiXS,YangQL,ZhaoMQ,BaiWW,LiangYQ,LiuXJ,GaoB,ZhangDY.TranscriptionalprofilinganalysisprovidinginsightsintodesiccationtolerancemechanismsofthedesertmossSyntrichiacaninervis.FrontiersinPlantScience,2023,14:1127541.[21]㊀张一弓,张怡,阿依白合热木㊃木台力甫,张道远.异源过表达齿肋赤藓ScABI3基因改变拟南芥气孔表型并提高抗旱性.植物学报,2021,56(4):414⁃421.[22]㊀卓露,张道远,李小双,杨红兰,管开云.荒漠苔藓齿肋赤藓(Syntrichiacaninervismitt.)瞬时遗传转化方法建立.分子植物育种,2019,17(6):1913⁃1919.[23]㊀李春.盐胁迫下齿肋赤藓的适应机制研究[D].新疆:新疆大学,2016.[24]㊀刘卫国,丁俊祥,邹杰,林喆,唐立松.NaCl对齿肋赤藓叶肉细胞超微结构的影响.生态学报,2016,36(12):3556⁃3563.[25]㊀LiZ,WangN,ZhangX,ChengH,LiY.HighprecipitationandlowevaporationresultedinhighlakelevelsoftheJuyanzepaleolake,northwestChina,during34⁃26calkyrBP.ClimateResearch,2016,69(3):193⁃207.[26]㊀CᶄosicᶄM,Vujic㊅icᶄMM,SabovljevicᶄMS,SabovljevicᶄAD.EffectsofABAandNaClonphysiologicalresponsesinselectedbryophytespecies.Botany,2020,98(11):639⁃650.[27]㊀SilvaEN,RibeiroRV,Ferreira⁃SilvaSL,ViégasRA,SilveiraJAG.Comparativeeffectsofsalinityandwaterstressonphotosynthesis,waterrelationsandgrowthofJatrophacurcasplants.JournalofAridEnvironments,2010,74(10):1130⁃1137.[28]㊀FreemanJL,PersansMW,KenNM,AlbrechtC,PeerW,PickeringIJ,SaltDE.Increasedglutathionebiosynthesisplaysaroleinnickeltoleranceinthlaspinickelhyperaccumulators.ThePlantCell,2004,16(8):2176⁃2191.[29]㊀卓露,薛山,买买提明㊃苏来曼,李鸿彬,张道远.基于形态结构探讨新疆干旱地区真藓(Bryumargenteum)的环境适应性,生态学报,2023,43(21):8865⁃8874.[30]㊀YangJE,CaoY,YangZY,ZhangWM,SunLJ,LuCM.Morphological,physiologicalandbiochemicalresponsesofbiofuelplantEuphorbialathyristosaltstress.ActaAgriculturaeScandinavica,,2013,63(4):330⁃340.[31]㊀SabovljevicᶄM,SabovljevicᶄA.ContributiontothecoastalbryophytesoftheNorthernMediterranean:Aretherehalophytesamongbryophytes?,2007,13(3):131⁃135.[32]㊀AliA,YunDJ.Saltstresstolerance;whatdowelearnfromhalophytes?JournalofPlantBiology,2017,60(5):431⁃439.[33]㊀ZhangW,LiuSH,LiCC,ZhangPY,ZhangPY.TranscriptomesequencingofAntarcticmossundersaltstressemphasizestheimportantrolesoftheROS⁃scavengingsystem.Gene,2019,696:122⁃134.[34]㊀ZechmeisterHG.BryophytesofcontinentalsaltmeadowsinAustria.JournalofBryology,2005,27(4):297⁃302.[35]㊀LiangYQ,LiXS,ZhangDY,GaoB,YangHL,WangYC,GuanKY,WoodAJ.ScDREB8,anovelA⁃5typeofDREBgeneinthedesertmossSyntrichiacaninervis,conferssalttolerancetoArabidopsis.PlantPhysiologyandBiochemistry,2017,120:242⁃251.[36]㊀YangHL,ZhangDY,LiHY,DongLF,LanHY.EctopicoverexpressionofthealdehydedehydrogenaseALDH21fromSyntrichiacaninervisintobaccoconferssaltanddroughtstresstolerance.PlantPhysiologyandBiochemistry,2015,95:83⁃91.[37]㊀OliverMJ,VeltenJ,WoodAJ.Bryophytesasexperimentalmodelsforthestudyofenvironmentalstresstolerance:Tortularuralisanddesiccation⁃toleranceinmosses.PlantEcology,2000,151(1):73⁃84.[38]㊀BatesLS,WaldrenRP,TeareID.Rapiddeterminationoffreeprolineforwater⁃stressstudies.PlantandSoil,1973,39(1):205⁃207.[39]㊀LassouaneN,AïdF,LuttsS.WaterstressimpactonyoungseedlinggrowthofAcaciaArabica.ActaPhysiologiaePlantarum,2013,35(7):2157⁃2169.[40]㊀CarraPÓ.Photosyntheticpigmentsofalgae.Phycologia,1991,30(2):235.[41]㊀刘群,彭斌,田长彦,赵振勇,王雷,王守乐.8种盐生植物种子萌发特征与NaCl盐度的关系.生态学报,2023,43(17):7284⁃7293.[42]㊀颜佳倩,顾逸彪,薛张逸,周天阳,葛芊芊,张耗,刘立军,王志琴,顾骏飞,杨建昌,周振玲,徐大勇.耐盐性不同水稻品种对盐胁迫的响应差异及其机制.作物学报,2022,48(6):1463⁃1475.[43]㊀王玉祥,张博,王涛.盐胁迫对苜蓿叶绿素㊁甜菜碱含量和细胞膜透性的影响.草业科学,2009,26(3):53⁃56.[44]㊀孟凡娟,庞洪影,王建中,李淑艳,王彦杰.NaCl和Na2SO4胁迫下两种刺槐叶肉细胞叶绿体超微结构,2011,31(3):0734⁃0741.[45]㊀韦存虚,王建军,王建波,周卫东,孙国荣,梁建生.Na2CO3胁迫对星星草叶肉细胞超微结构的影响.生态学报,2006,26(1):108⁃114.[46]㊀BijeloviA,SabovljeviM,GrubisicD,KonjeviR.Phytohormoneinfluenceonthemorphogenesisoftwomosses(BryumargenteumHedw.andAtrichumundulatum(Hedw.)P.Beauv.).IsraelJournalofPlantSciences,2004,52(1):31⁃36.[47]㊀刘吉祥,吴学明,何涛,张文静.盐胁迫下芦苇叶肉细胞超微结构的研究.西北植物学报,2004,24(6):1035⁃1040.[48]㊀马晓蓉,杨淑娟,姚宁,王玲霞,马强,梁文裕.NaCl胁迫对宁夏枸杞叶和幼根显微及超微结构的影响.西北植物学报,2021,41(12):2087⁃2095.[49]㊀王娟,李德全.逆境条件下植物体内渗透调节物质的积累与活性氧代谢.植物学通报,2001,18(4):459⁃465.[50]㊀卓露.两种极端耐干苔藓抗逆性及其居群遗传分化研究[D].北京:中国科学院大学,2018.[51]㊀尹本丰,张元明.冻融过程对荒漠区不同微生境下齿肋赤藓渗透调节物含量和抗氧化酶活力的影响.植物生态学报,2015,39(5):517⁃529.1943㊀8期㊀㊀㊀林晓华㊀等:NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响㊀。
nacl溶液 电导率计算

nacl溶液电导率计算摘要:一、引言二、电导率的概念三、NaCl溶液的电导率计算1.公式2.影响因素四、实际应用1.电导率计测量2.在生活中的应用五、结论正文:一、引言电导率是描述溶液导电性强弱的一个物理量,它在化学、环境科学、生物科学等领域具有重要的意义。
本文将详细介绍如何计算NaCl溶液的电导率,并探讨影响电导率的因素以及实际应用。
二、电导率的概念电导率(σ)是指在单位长度、横截面积的导体内,通过单位时间内的电荷量(Q)与电场强度(E)之比。
它是一个无量纲的物理量,通常用来衡量溶液的导电能力。
三、NaCl溶液的电导率计算1.公式对于强电解质,如NaCl溶液,其电导率可以用Nernst-Einstein公式计算:σ= 0.0592 × (c × Z)^2其中,σ为电导率,c为溶液浓度(单位:mol/L),Z为离子的电荷数。
2.影响因素(1)溶液浓度:溶液浓度越高,电导率越大。
(2)离子电荷数:离子电荷数越大,电导率越大。
(3)温度:温度对电导率的影响较大。
通常情况下,温度升高,电导率增大。
四、实际应用1.电导率计测量电导率计是测量溶液电导率的仪器,通过测量电导率可以了解溶液的导电性能,从而对溶液的性质进行判断。
2.在生活中的应用电导率在生活有许多应用,例如:在饮用水处理过程中,需要控制水的电导率以保证水质;在电池制造过程中,需要监测电解液的电导率以调整电池性能等。
五、结论通过本文的介绍,我们了解了NaCl溶液的电导率计算方法以及影响电导率的因素。
电导率测定
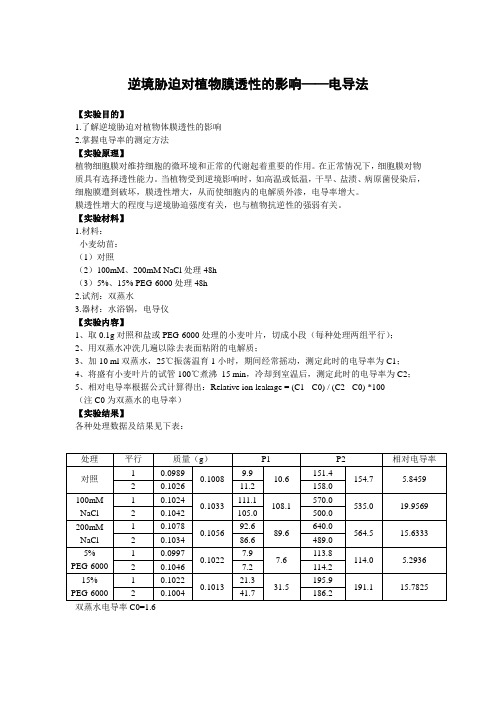
逆境胁迫对植物膜透性的影响——电导法【实验目的】1.了解逆境胁迫对植物体膜透性的影响2.掌握电导率的测定方法【实验原理】植物细胞膜对维持细胞的微环境和正常的代谢起着重要的作用。
在正常情况下,细胞膜对物质具有选择透性能力。
当植物受到逆境影响时,如高温或低温,干旱、盐渍、病原菌侵染后,细胞膜遭到破坏,膜透性增大,从而使细胞内的电解质外渗,电导率增大。
膜透性增大的程度与逆境胁迫强度有关,也与植物抗逆性的强弱有关。
【实验材料】 1.材料: 小麦幼苗: (1)对照(2)100mM 、200mM NaCl 处理48h (3)5%、15% PEG-6000处理48h 2.试剂:双蒸水3.器材:水浴锅,电导仪【实验内容】1、取0.1g 对照和盐或PEG-6000处理的小麦叶片,切成小段(每种处理两组平行);2、用双蒸水冲洗几遍以除去表面粘附的电解质;3、加10 ml 双蒸水,25℃振荡温育1小时,期间经常摇动,测定此时的电导率为C1;4、将盛有小麦叶片的试管100℃煮沸 15 min ,冷却到室温后,测定此时的电导率为C2;5、相对电导率根据公式计算得出:Relative ion leakage = (C1 - C0) / (C2 - C0) *100 (注C0为双蒸水的电导率) 【实验结果】各种处理数据及结果见下表: 双蒸水电导率C0=1.6处理 平行 质量(g ) P1 P2相对电导率 对照 1 0.0989 0.1008 9.9 10.6 151.4 154.7 5.8459 2 0.1026 11.2 158.0 100mM NaCl 1 0.1024 0.1033 111.1 108.1 570.0 535.0 19.9569 2 0.1042 105.0 500.0 200mM NaCl 1 0.1078 0.1056 92.6 89.6 640.0 564.5 15.6333 2 0.1034 86.6 489.0 5% PEG-6000 1 0.0997 0.1022 7.9 7.6 113.8 114.0 5.2936 2 0.1046 7.2 114.2 15%PEG-6000 10.10220.101321.3 31.5195.9 191.115.78252 0.1004 41.7186.2【实验讨论】1.从结果中可以看出,逆境胁迫会导致小麦叶片的膜透性增加,且胁迫程度越大,溶液电导率越大。
盐胁迫下1年生麻栎实生苗的生理变化

盐胁迫下1年生麻栎实生苗的生理变化李勇;葛晓敏;唐罗忠;黄开栋;郑萌芳【摘要】采用室内盆栽法对1年生麻栎实生苗进行了盐(NaCI)胁迫试验,结果显示,土壤含盐量为0.4%和0.5%时,苗木叶片边缘会出现明显的失绿现象.高含量的盐处理会明显抑制麻栎新梢生长.土壤含盐量高于0.3%时,苗木叶片的含水量、叶绿素含量和光合速率均显著下降,而相对电导率显著升高.土壤含盐量较高时,苗木嫩叶中的N含量有所下降.苗木细根中的K含量会随着土壤含盐量的提高而降低,叶片中的K含量却呈相反趋势.当土壤含盐量低于0.3%时,苗木叶片中的Na含量处于较低水平,但高于0.4%时,叶片中的Na含量会显著升高;细根中的Na含量随着土壤含盐量的升高而升高.综合分析认为,土壤含盐量低于0.2%时,麻栎苗木能够正常生长;土壤含盐量高于0.3%时,麻栎苗木的生理会受到明显影响.【期刊名称】《江苏林业科技》【年(卷),期】2015(042)004【总页数】5页(P17-21)【关键词】麻栎;苗木;盐胁迫;生长;叶绿素;光合速率;相对电导率【作者】李勇;葛晓敏;唐罗忠;黄开栋;郑萌芳【作者单位】南方现代林业协同创新中心,南京林业大学,江苏南京210037;南方现代林业协同创新中心,南京林业大学,江苏南京210037;南方现代林业协同创新中心,南京林业大学,江苏南京210037;南方现代林业协同创新中心,南京林业大学,江苏南京210037;南方现代林业协同创新中心,南京林业大学,江苏南京210037【正文语种】中文【中图分类】S792.181盐碱土是陆地上分布最广的土壤类型之一,约占陆地总面积的25%。
仅在我国,盐碱地面积就有3 400万hm2以上[1]。
土壤盐渍化是影响植物生长和产量的重要环境因子[2-3],全世界约有1/3的土壤盐渍化,目前我国各类盐渍土总面积约为1亿hm2[4]。
土壤中过量的盐分会引起土壤理化性质发生改变,从而导致大部分植物生长受到影响、土地生产力下降[5]。
标准nacl溶液电导率

标准nacl溶液电导率
标准NaCl溶液的电导率可以根据其浓度来确定。
NaCl是常见的电解质,其溶液的电导率与浓度呈正相关关系。
以下是一些常见浓度下的NaCl溶液电导率的大致范围:
1.0.01MNaCl溶液的电导率约为1.3-1.5mS/cm。
2.0.1MNaCl溶液的电导率约为12-14mS/cm。
3.1MNaCl溶液的电导率约为120-140mS/cm。
这些数值仅作为参考,实际的电导率可能会受到温度、离子活动度等其他因素的影响。
此外,使用不同的测量设备和方法也可能导致稍有不同的结果。
如果需要更准确的电导率数值,建议参考相关的化学手册、实验室方法或标准化组织的指南。
这些资源可以提供更详细和准确的NaCl 溶液电导率数据。
1/ 1。
氯化钠标准电导率计算公式

氯化钠标准电导率计算公式电导率是描述溶液中电解质浓度的重要参数,它是电解质溶液导电能力的一个指标。
而氯化钠是一种常见的电解质,因此氯化钠标准电导率计算公式是很重要的。
本文将介绍氯化钠标准电导率计算公式的推导过程和应用。
首先,我们来看一下电导率的定义。
电导率(κ)是指单位长度内单位横截面积的溶液的电导能力,通常用西门子/厘米(S/cm)来表示。
电导率与电解质溶液中的电解质浓度密切相关,一般来说,电导率随着电解质浓度的增加而增加。
对于氯化钠溶液来说,其电导率可以通过以下公式来计算:κ = λ× c。
其中,κ表示电导率,λ表示电解质的等当电导率,c表示电解质的浓度。
接下来,我们来看一下氯化钠标准电导率计算公式的推导过程。
首先,我们需要知道氯化钠的等当电导率。
氯化钠在无限稀释时的等当电导率可以通过实验测定得到,一般来说,氯化钠在25摄氏度下的等当电导率为7.6S/cm。
其次,我们需要知道氯化钠的浓度。
浓度是指单位体积内溶质的质量或摩尔数,通常用克/升或摩尔/升来表示。
对于氯化钠溶液来说,其浓度可以通过溶质的质量或摩尔数与溶剂的体积之比来计算。
最后,我们可以将氯化钠的等当电导率和浓度代入电导率计算公式中,就可以得到氯化钠溶液的电导率了。
举个例子,假设我们有一个氯化钠溶液,其浓度为0.1摩尔/升,那么根据上述公式,该溶液的电导率为:κ = 7.6 S/cm × 0.1 mol/L = 0.76 S/cm。
通过这个简单的例子,我们可以看到氯化钠标准电导率计算公式的应用。
当然,实际情况中可能会有更复杂的情况,比如溶液中可能还含有其他电解质,或者溶液的温度不是25摄氏度等因素,这时候就需要对公式进行修正或者使用更为复杂的计算方法。
除了上述的计算方法外,还有一种常用的方法是通过电导率计来直接测量氯化钠溶液的电导率。
电导率计是一种专门用来测量溶液电导率的仪器,它可以直接给出溶液的电导率数值,非常方便快捷。
濒危植物长叶红砂适应盐胁迫的生理生化机制研究

濒危植物长叶红砂适应盐胁迫的生理生化机制研究薛焱;王迎春;王同智【摘要】以濒危盐生植物长叶红砂(Reaumuria trigyna)幼苗为材料,研究了不同浓度NaCl溶液(0、100、200、300和400 mmol/L)处理30 d对其生长和生理生化指标的影响,以分析长叶红砂的耐盐生理机制.结果表明:(1)100和200 mmol/L NaCl处理时,长叶红砂鲜重和干重均显著增加,但随着盐浓度继续增加,长叶红砂幼苗生长受到抑制,且地上部受到的抑制大于根部,显示长叶红砂适宜生长的NaC1浓度是200 mmol/L. (2)随NaCl胁迫浓度的升高,长叶红砂的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)呈下降趋势,胞间CO2浓度(C1)呈上升趋势,说明光合速率的下降使利用CO2的能力下降,胞间积累了大量的CO2,且盐处理浓度越高量越大.(3)随NaCl胁迫浓度的升高,长叶红砂幼苗Na+、Cl含量增加,可溶性糖、脯氨酸、游离氨基酸及可溶性蛋白等有机渗透调节物质的合成增加.研究认为,长叶红砂是通过调节叶片Na+ 、Cl-以及有机渗透调节物质含量来提高其耐盐能力.%Reaumuria trigyna seedings were treated with NaCl of different concentrations (0,100,200,300 and 400 mmol/L) for 30 days. A study about its growth,physiological and biochemical mechanisms in different NaCl concentrations showed: (1)Under 100 and 200 mmol/L NaCl stress,the fresh weight and dry weight increased significantly than that of control. Growth of R. Trigyna was inhibited with increasing NaCl concentrations, showing that the suitable NaCl concentration of R. Trigyna seedlings growth was 200 mmol/L. (2)Pn,Tr and Gs decreased and C,increased,showing decrease of photosynthesis rate causing reduction of utilization rate and intracellular CO2 accumulation which was higher whenNaCl concentration was higher. (3)Osmotic adjustments of R. Trigyna mainly relied on Na+ and Cl- ,soluble sugar,free amino acid,proline and soluble protein. This study discovered that R. Trigyna could improve its salt tolerance ability by adjusting Na+ and Cl- content and osmosis regulating substances in leaves.【期刊名称】《西北植物学报》【年(卷),期】2012(032)001【总页数】7页(P136-142)【关键词】长叶红砂;盐胁迫;生长;光合特性;渗透调节【作者】薛焱;王迎春;王同智【作者单位】内蒙古大学生命科学学院,呼和浩特010021;包头医学院,内蒙古包头014060;内蒙古大学生命科学学院,呼和浩特010021;内蒙古大学生命科学学院,呼和浩特010021【正文语种】中文【中图分类】Q945.79土壤盐渍化是世界性的资源环境和生态问题,盐渍化土地的开发与利用已成为人们关注的热点。
氯化钠导电性

氯化钠导电性
氯化钠(NaCl)是一种盐,是一种由氯原子和钠原子共同组成的化合物,是一种常见的无机盐。
它有着大量的应用,也是一种重要的化学原料。
本文将重点讨论氯化钠的导电性。
氯化钠本身是一种导电物质,它是由氯离子(Cl-)和钠离子(Na+)组成的,这些离子对电荷有吸引力,并以此作为其共同的导电机制。
由于氯离子具有负电而钠离子具有正电,因此,当电荷在氯化钠中流动时,离子会传播电荷,从而使氯化钠具有导电性。
此外,由于氯化钠中的离子和电荷的结合力比较弱,在一定的温度和压力下极易被电场分离,使氯化钠的导电性得以最大程度的发挥。
因此,氯化钠可以用作电路中的负载元件,例如,可以用作电阻元件,在实际应用中,氯化钠也可以用作电池液中的电解质,以提高电池的性能。
另外,氯化钠也可以被利用来测量室温下水的电导率,因为水本身不久导电,但加入一定数量的氯化钠后,其电导率就会变高。
因此,氯化钠可以被用来大致估计水中悬浮物的含量,也可以用来研究水的电催化特性。
此外,氯化钠的导电性还可以用来研究其他物质的导电性,由于氯化钠本身的导电性可以作为一个基准,可将其他物质的导电性与之进行比较。
总的来说,氯化钠是一种导电物质,由氯离子(Cl-)和钠离子(Na+)组成,具有很强的电荷吸引力,可以用作元件、离子液体、
电池液等电子材料。
通过测量水的电导率,它还可以用来研究水的电催化特性以及其它物质的导电性。
另外,在电路设计中,氯化钠也可以被用来构建电路的电阻元件,以此提高电路的性能。
因此,氯化钠一直是服务于电子学领域的重要原料,具有众多的应用前景。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第4 7卷 第 1 2期
2011年 1 月 2
林
业
科
学
Vo . 1 47. 1 No 2 De - 2 0 1 c.. 1
S EN lA CI I '
ቤተ መጻሕፍቲ ባይዱ
S LVAE I
S N【 I CAE
N C 胁 迫 下 流 苏 的 电阻抗 与 电导分 析 a1
Elc r c lI pe a c n e t o y e Le ka e A n l sso e t i a m d n e a d El c r l t a g a y i f Chi e e Frn e r e n s ig te
( h o a t u eu u )u d rNa te s C in n h sr ts s n e CIS r s
姚 琳 张 钢
( 北农 业 大 学 园 艺学 院 河 保 定 0 10 ) 7 0 1
摘 要 : 以 流苏 2年 生 苗 木 为 材 料 , 定 其 叶片 与 茎 在 N C 胁 迫 ( ,. % ,. % , . % ) 的 电 阻 抗 图 谱参 数 测 a1 00 4 0 8 12 下 和 电导 率 的 变 化 , 析 电阻 抗 图谱 参 数 与 电导 率 间 的 相 关 性 , 分 以讨 论 在 N C 处 理 下 用 电 阻 抗 图谱 法 研 究 流 苏 受胁 aI
价 流 苏 叶 和 茎 N C 胁 迫 的最 佳 参 数 。 a1
关 键 词 : 流 苏 ; a 1 迫 ;电 阻抗 图谱 ; 导 率 NC胁 电
中 图 分 类 号 : 7 84 ; 67 1 ¥ 1 . 3 ¥ 8 . 文 献 标 识 码 :A 文 章 编 号 :1 0 — 4 8 2 1 ) 2— 0 0— 9 0 1 7 8 (0 1 1 0 6 0
迫 程 度 的有 效 性 。结 果 表 明 : ) 着 N C 浓 度 的 升 高 , 1随 a1 叶片 电导 率 显 现 逐 渐 升 高 或 先 升 高 后 下 降 的趋 势 , 的 表 茎
现 不 明显 ;随 着胁 迫 时 间 的延 长 叶片 和 茎 电导 率 , 整体 均显 现 逐 渐 升 高 或 先 升 高 后 降 低 的趋 势 。 流 苏 茎 的 耐 盐性 强 于 叶 片 。2 随 着 N C 浓 度 的 升 高 , 苏 叶片 和 茎 的高 频 电 阻 率 r胞 内 电 阻 率 r 和 弛 豫 时 间 呈 现 先 增 大 后 减 ) a1 流 、 . 小 或 逐 渐 减 小 的特 征 ;随着 胁迫 时 间 的 延 长 叶 片 和 茎 的 各 处 理 的 电 阻 抗 图谱 参 数 均 在 胁 迫 1 5天 时 出 现 最 低 值 相关 分析 表 明 :流 苏 对 0 8 以上 N C 浓 度 胁 迫 敏 感 , 电阻 抗 图 谱 参 数 可 以 表 征 流 苏 受 N C 胁 迫 的 程 度 , 用 .% aI 用 al 且 ES 数 胞 内 电 阻率 r、 I参 高频 电 阻率 r 表 征 叶 耐 盐 特性 , 胞 内 电阻 率 r、 豫 时 间 来 表 征 茎 耐 盐 特 性 。 r为 评 来 用 l 弛 .
r l tv l c r l t e k g ft e l a e n r a e r d a l r i c e s d f sl n h n d c e s d Ho e e h t msd d e a i e e e to y e l a a e o e v s i c e s d g a u ly o n r a e i t a d t e e r a e . h r y w v rt e se i n to v o s r s o s so h e k g o s l sr s e .W i n r a i g d r t n o e sr s ,t e r l t e e e t l t e k g o b i u e p n e ft e l a a e t a t te s s t i c e s n u a i ft te s h e a i l c r y e l a a e h o h v o
o hn s r g t e( ho a tu e ss ev sa dse n e a I t s 0 4 , . % a d 1 2 )( tra f ieeF i er C in nh srt u )lae n tmsu d rN C r s( . % 0 8 C n e u se n . % wa s e
Abta t src :
T eeet cli e a c pcrso y( S aa tr a drlt eeet lt la a ela e n tms t h lcr a mp d n ese t c p EI)p rmees n eai lcr ye ek g e vsa ds i o v o e o
Ya n Z a g Ga g o Li h n n ( ol e f H r c l r , g i l rl nvr t o bi B o i 7 C l g ot ut e A r ut a i s y fHee e o i u c u U e i a dn 0 0 ) g 1 1 O
p rmeesa drlt eee t lt la ae h eut so e h t )W i h n raigi o c nrt no a 1 h aa tr n eai lc oye ek g .T ersl h w dta:1 v r s t teice s cn e t i fN C ,te h n n ao