普通遗传学实验-小麦单体
小麦基因育种实验报告

一、实验背景小麦(Triticum aestivum L.)作为全球最重要的粮食作物之一,其产量和品质的提高对于保障全球粮食安全具有重要意义。
近年来,随着分子生物学和基因工程技术的快速发展,小麦基因育种成为研究热点。
本实验旨在通过基因工程技术,将外源抗病基因导入小麦基因组,培育出抗病、高产的小麦新品种。
二、实验材料与方法1. 实验材料- 小麦品种:普通小麦品种“扬麦11号”- 抗病基因:来源于抗病小麦品种“抗病9号”的Rab基因- 重组质粒:含有Rab基因的重组质粒pUC19- 载体菌:大肠杆菌DH5α- 转化试剂:钙离子- 植物细胞培养基:MS培养基2. 实验方法1. 构建重组质粒- 将抗病基因Rab从抗病小麦品种“抗病9号”中克隆到载体质粒pUC19中,构建重组质粒pUC19-Rab。
- 转化大肠杆菌- 将重组质粒pUC19-Rab转化到大肠杆菌DH5α中,筛选阳性克隆。
- 植物细胞培养- 将阳性克隆提取质粒,电转化小麦愈伤组织,筛选阳性愈伤组织。
- 愈伤组织再生- 将阳性愈伤组织诱导分化再生小麦植株。
- 抗病性鉴定- 将再生植株接种白粉病菌,观察植株抗病性。
- 分子鉴定- 对抗病植株进行PCR扩增,检测Rab基因插入情况。
三、实验结果与分析1. 构建重组质粒成功构建了含有抗病基因Rab的重组质粒pUC19-Rab。
2. 转化大肠杆菌转化效率达到90%以上,获得阳性克隆。
3. 植物细胞培养成功诱导出阳性愈伤组织,再生出小麦植株。
4. 抗病性鉴定部分再生植株表现出较强的抗病性,抗病率约为60%。
5. 分子鉴定PCR扩增结果显示,部分再生植株中含有Rab基因。
四、实验结论本实验成功地将抗病基因Rab导入小麦基因组,获得了抗病、高产的小麦新品种。
这为小麦基因育种提供了新的思路和方法,有助于提高小麦产量和品质,保障粮食安全。
五、实验讨论1. 重组质粒构建成功,转化效率较高,表明实验方法可行。
2. 部分再生植株表现出较强的抗病性,说明抗病基因Rab已成功导入小麦基因组。
普通小麦株高的遗传分析

普通小麦株高的遗传分析作者:吴旭江程凯臧淑江张伯桥来源:《科学与信息化》2016年第34期摘要利用小麦扬麦9号和CI12633构建了184个重组自交系群体,选择双亲间具有多态的212个SSR标记绘制分子连锁图谱,图谱总长度1567.2cM,标记间平均距离8.2cM。
在3年9次试验条件下对株高性状进行鉴定,利用复合区间作图法监测到6个株高QTL,它们分别位于1D、2A、2B、3A和5A染色体上,其中位于2B染色体上的QTL来自品种CI12633,其余5个QTL均来自矮杆亲本扬麦9号,单个QTL能够解释4.13%-17.44%的表型变异,每个环境条件下检测到的所有QTL能解释29.46%-46.46%的表型变异,5A染色体上的QTL在9次试验环境下均能被检测出来,同时其效应也是最大的QTL,说明这个QTL能够在育种中被利用。
关键词普通小麦;株高;数量性状位点Mapping QTLs for Plant Height using a recombinant inbred lines in common wheatWU Xujiang1, CHENG Kai1, ZANG Shujiang1, ZHANG Boqiao1,2Lixiahe Regional Institute of Agricultural Sciences of Jiangsu/Key Laboratory of Wheat Biology and Genetic Breeding in the Middle and Lower Yangtze River (Ministry Agriculture), Yangzhou 225007, Jiangsu1; Jiangsu Collaborative Innovation Center for Modern Crop Production2Key words: common wheat; plant height; quantitative trait locus小麦株高对产量的形成有重要的影响,研究它的发育遗传规律有助于了解产量形成的机制。
一、小麦基因组DNA提取

一、小麦基因组DNA提取:1、用单籽粒粉碎机磨碎1~3粒小麦干种子,放入2ml离心管中,加900μlSDS提取液,50-60℃水浴45-60min,期间间歇震荡。
2、12000rpm,10min,吸取700μl上清液,加入等体积(700μl)酚-氯仿-异戊醇(25:24:1),于冰上颠倒混匀15min。
3、12000rpm,10min,吸取500μl上清液,加入0.6倍异丙醇(300μl)和1/10倍体积(50μl)NaAc(pH5.2),混匀后冰上静置15min,期间间歇震荡。
4、出现絮状DNA沉淀,10000rpm,10min。
5、弃上清,沉淀用70%乙醇漂洗2遍,然后用无水乙醇漂洗1遍,室温晾干,加100μl的1×TE(或双蒸水)溶解,过夜,10倍稀释,备用。
二、PCR反应体系(10 ul,用于SSR分析):终浓度10 ulWater 3.87ul10×buffer(Mg2+)1× 1.0uldNTP(2.5mM) 0.25mM 1.0ulTaq(5U/ul) 0.65U 0.13ulPrimer(2uM/ul/条) 0.4uM(/条) 2.0ulDNA 2.0ul三、PCR扩增条件(用于SSR分析):94℃预变性5min94℃变性1min60℃/58℃/55℃/50℃退火 1min 35个循环72℃延伸1min72℃延伸5min20℃保存PCR完成后,加变性双指示剂(DNA loading buffer)在PCR仪上95℃变性5~10min,然后立即置于冰上,4℃存准备点样电泳。
四、聚丙烯酰胺凝胶电泳准备工作:首先用去污剂清洗玻璃板(包括耳朵板和底板),凉干后,先用95%乙醇涂擦两块玻璃板,然后再用亲水硅烷(Binding silane)均匀涂擦底板,用疏水硅烷(Repel silane)于通风橱中均匀涂擦耳朵板,稍凉一会待玻璃表面变干,然后组装两块玻璃板(期间注意调节水平仪),开始灌胶,边灌边轻轻拍打玻璃表面,以防胶内产生气泡。
普通小麦发育类型的遗传

普通小麦发育类型的遗传本文旨在探讨普通小麦发育类型的遗传背景。
小麦是世界上人类最常食用的粮食作物,其发育类型受到遗传因素的影响。
以提高种子多样性为目标,近年来许多研究已经聚焦于种子发育类型,尝试识别不同植物品种中特定遗传因子的作用。
对于小麦而言,研究者已经识别出多个基因家族来控制种子发育类型。
其中,一些基因可以控制小麦的穗发育,如Dt1和Rht-B1;而另一些基因可以调节株高,如Rht-D1、Rht-B1a 和Rht-D1b。
此外,关键的生育期基因 TaGa20-A1也可以影响株高和穗长,但我们对其作用的定义仍然不明确。
另外,研究者也发现了可以控制小麦穗大小、种子大小和形状的基因,例如Gml1和Cml2。
这些基因通过调节外源激素水平,从而影响小麦所发育出的茎、叶和穗。
除此之外,除了发育本身需要的基因作用外,研究者还发现了一些可以抵抗外部环境因素对小麦发育的影响的基因,例如病毒抗性的Nb1和低温抗性的HvFTI。
总的来说,普通小麦发育类型的遗传构成复杂且丰富,受到许多不同的因素的调节。
研究发现的多个基因家族的功能,以及将其应用于植物育种,可以帮助我们在提高小麦收成和质量方面取得更进一步的进展。
本研究表明,小麦的发育类型受到多种因素的影响,包括基因、外部条件和环境因素。
在进行基因育种过程中,不同基因家族可以通过调节水平来控制穗发育、株高和穗大小、种子大小和形状。
为了更有效地改善小麦的收成和质量,育种者需要能够识别并调节多种发育相关因素,如气候条件、植物内部生理活动和遗传因子的作用。
利用这些信息,育种者可以将基因组技术和传统的育种技术有效地结合起来,从而获得更好的品质和产量。
此外,为了充分发挥植物发育类型的潜力,研究者还需要开展更多的研究,以识别遗传变异、病毒抗性机制以及其他影响植物发育的因素,以及这些因素之间的相互作用。
只有充分理解植物发育类型的遗传背景,才能实现更佳的品种育种,进一步提高小麦的收成和质量。
普通小麦-中间偃麦草杂种后代矮秆材料的细胞学鉴定的开题报告

普通小麦-中间偃麦草杂种后代矮秆材料的细胞学鉴定的开题报告一、研究背景小麦是世界上最重要的粮食作物之一,其生产与发展一直是各国政府十分重视的问题。
当前,国内外普遍主张开展小麦品种改良工作,其中以矮秆、高产、抗病害、高品质、耐旱耐涝等性状为改良目标。
因此,对小麦的细胞遗传学进行深入研究显得尤为必要。
二、研究意义小麦细胞遗传学研究对于推动小麦品种改良研究、提高小麦品种的适应性、增强小麦的产量和品质、改善和保护生态环境等方面具有重要的意义,而其中杂种育种是一个重要手段之一。
本研究针对普通小麦(中间偃麦草杂种后代矮秆材料)的细胞遗传学进行鉴定,可为了解杂交后代的细胞遗传学特性与变异规律、指导育种选优、提升小麦品种的适应性与产量等方面提供重要理论依据和科学支持。
三、研究内容与目标本研究主要针对普通小麦(中间偃麦草杂种后代矮秆材料)进行细胞学鉴定,具体研究内容包括:小麦遗传物质的形态结构与细胞形态特征分析,细胞遗传学方法分析以及细胞遗传学特征鉴定等方面。
旨在深入研究普通小麦(中间偃麦草杂种后代矮秆材料)的细胞遗传学特征,为小麦品种的改良与发展提供科学依据。
四、研究方法本研究将采用细胞遗传学研究方法,主要包括显微镜、细胞质染色、染色质显微镜、核型分析等技术手段,对样品进行细胞形态学检测、细胞核型分析等内容。
五、研究计划本研究拟于2021年1月开始,至2022年6月结束,大致时间安排和任务如下:1. 2021年1-3月:细胞样品的采集和制备。
2. 2021年4-8月:细胞样品的形态结构和细胞形态特征分析。
3. 2021年9月-2022年1月:细胞遗传学方法分析。
4. 2022年2-6月:细胞遗传学特征鉴定。
六、预期成果本研究的预期成果包括:对普通小麦(中间偃麦草杂种后代矮秆材料)的细胞学鉴定进行深入研究,明确其细胞遗传学特征与变异规律;提供数据支持,指导小麦育种选优,为生产提供科学依据;并取得相关研究成果的科研论文发表。
普通小麦籽粒性状的主基因+多基因遗传模型分析

普通小麦籽粒性状的主基因+多基因遗传模型分析的报告,
600字
小麦籽粒性状的主基因+多基因遗传模型分析报告
本报告尝试分析小麦籽粒性状的主要基因及其多基因遗传模型。
从小麦籽粒性状丰度及质量方面来看,有不同程度影响的基因有新垣(Xyn)家族基因、赤霉素耐性基因(En)家族基因、
膨胀性基因(Ex)家族基因、糊精基因(Gluin)家族基因及
其他相关基因。
在本报告中,我们将使用多基因遗传模型对小麦籽粒性状进行分析,以更好地了解不同基因家族对小麦籽粒性状的影响情况。
我们将基于多个数据集,包括植物基因组学数据、表型数据以及分子标记数据,对小麦籽粒性状进行遗传分析。
通过对基因家族及其基因表达水平进行分析,发现Xyn家族
基因在小麦籽粒性状中扮演了主要角色,其丰度和质量都有显着的影响。
En家族基因也发挥了重要作用,尤其是对小麦籽
粒质量的影响,Ex家族基因对小麦籽粒性状的影响最大,其
贡献率甚至可以达到90%,而Gluin家族基因则对小麦籽粒性
状影响最小。
将多基因遗传模型应用于小麦籽粒性状,可以有效地确定重要基因家族及其如何影响小麦籽粒性状,从而指导后续小麦种子性状调控技术的开发。
本报告对小麦籽粒性状的主要基因及其多基因遗传模型提供了有价值的信息,为小麦籽粒性状精准调控提供了重要依据。
遗传学综合复习题_整理版

遗传学综合复习题1.试用“显性假说”、“超显性假说”的遗传理论解释杂种优势产生的原因,为什么在生产上不代?利用杂种F2答:○1显性假说的核心是杂种优势源自杂种非等位基因的互补作用,认为多数显性基因比隐性基因更有利于个体生长发育,不同自交系杂交,双亲的显性基因集中到了杂种中产生了互补作用,从而导致杂种优势。
超显性假说认为杂种优势来源于双亲异质结合所引起的等位基因互作,这一假说认为等位基因之间无显隐性,杂合等位基因的互作作用大于纯和等位基因间的作用.。
○2具有杂种优势的F1代一般都表现近交衰退现象。
根据遗传的基本规律,由于杂合体的基因分离和重组,F2群体必将出现基因型的分离和性状表现的分离。
因此,F2与F1相比较,在生长势、生活力、繁殖力、抗逆性等方面都会表现下降。
并且2个亲本的纯和程度越高,性状差异越大,F1表现的杂种优势越强,F2的衰退现象也越加明显。
BY:lhq 2.经典遗传学与分子遗传学发展阶段的基因概念的异同。
综合所学遗传学知识,试述生物发生遗传变异的主要途径。
答:经典遗传学认为基因是一个最小的单位,不能分割,既是结构单位,又是功能单位。
分子遗传学在精细的微生物遗传分析中查明,基因并不是不可分割的最小单位,而是远为复杂得多的遗传和变异的单位。
在遗传学中,基因的概念对应的是一种性状,分子遗传学中基因对应的是一个功能蛋白,后来又发展到与一个功能肽段相对应。
现在对基因的定义是一段含有遗传信息的核酸链,包括编码序列和调控序列,还包括RNA。
基因突变: 由于核酸序列发生变化,包括缺失突变、定点突变、移框突变等,使之不再是原有基因的现象。
例如:父母双方都是褐色的牛生出白色的牛.基因重组: 造成基因型变化的核酸的交换过程。
包括发生在生物体内(如减数分裂中异源双链的核酸交换)和在体外环境中用人工手段使不同来源DNA重新组合的过程。
例如:对于豌豆,D代表高茎基因,d代表矮茎基因,父母双方基因型均为Dd的高茎豌豆产生的基因型为dd的矮茎豌豆。
普通遗传学课后习题解答

普通遗传学课后习题解答第⼀章遗传的细胞学基础(p32-33)4.某物种细胞染⾊体数为2n=24,分别指出下列各细胞分裂期中的有关数据:(1)有丝分裂后期染⾊体的着丝点数。
(2)减数分裂后期I染⾊体着丝点数。
(3)减数分裂中期I的染⾊体数。
(4)减数分裂末期II 的染⾊体数。
[答案]:(1)48;(2)24;(3)24;(4)12。
[提⽰]:如果题⽬没有明确指出,通常着丝点数与染⾊体数都应该指单个细胞或细胞核内的数⽬;为了“保险”(4)也可答:每个四分体细胞中有12条,共48 条。
具有独⽴着丝点的染⾊体才称为⼀条染⾊体,由复合着丝点联结的两个染⾊体单体只能算⼀条染⾊体。
5.果蝇体细胞染⾊体数为2n=8,假设在减数分裂时有⼀对同源染⾊体不分离,被拉向同⼀极,那么:(1)⼆分⼦的每个细胞中有多少条染⾊单体?(2)若在减数分裂第⼆次分裂时所有的姊妹染⾊体单体都分开,则产⽣的四个配⼦中各有多少条染⾊体?(3)⽤n 表⽰⼀个完整的单倍染⾊体组,应怎样表⽰每个配⼦的染⾊体数?[答案]:(1)两个细胞分别为6 条和10 条染⾊单体。
(2)四个配⼦分别为3条、3 条、5条、5 条染⾊体。
(3)n=4 为完整、正常单倍染⾊体组;少⼀条染⾊体的配⼦表⽰为:n-1=3;多⼀条染⾊体的配⼦表⽰为:n+1=5。
[提⽰]:正常情况下,⼆价体的⼀对同源染⾊体分离并分配到两个⼆分体细胞。
在极少数情况下发⽣异常分配,也是染⾊体数⽬变异形成的原因之⼀。
6. ⼈类体细胞染⾊体2n=46,那么,(1)⼈类受精卵中有多少条染⾊体?(2)⼈的初级精母细胞、初级卵母细胞、精⼦、卵细胞中各有多少条染⾊体?[答案]:(1)⼈类受精卵中有46 条染⾊体。
(2)⼈的初级精母细胞、初级卵母细胞、精⼦、卵细胞中分别有46 条、46 条、23 条、23条染⾊体。
7.⽔稻细胞中有24条染⾊体,⼩麦中有42条染⾊体,黄⽠中有14条染⾊体。
理论上它们各能产⽣多少种含不同染⾊体的雌雄配⼦?[答案]:理论上,⼩稻、⼩麦、黄⽠各能产⽣=4096、=2097152、=128 种不同含不同染⾊体的雌雄配⼦。
遗传学练习题

遗传学练习题普通遗传学课程考试试题⼀⼀、名词解释(每⼩题2分, 共10分)1、花粉直感2、QTL3、单倍体4、连锁群5、狭义遗传率⼆、填空题(每空1分,共20分)1、保持平衡状态的群体应是⼀个______随机交配________群体。
2、表现显性的⼩麦单体1A,与表现隐性的双体杂交,若F1中的单体表现_____显性_____,则说明有关基因不在1A染⾊体上。
3、某DNA的核苷酸中,A的含量为30%,则G的含量为__20%________。
4、⼩麦和⿊麦是两个近缘种。
在⼩麦中常发⽣籽粒颜⾊的突变。
根据基因突变的__平⾏性_______可推测⿊麦中也可能发⽣⼦粒颜⾊的突变。
5、花粉母细胞经过减数分裂形成四个⼩孢⼦,每个⼩孢⼦的核再经过第⼀次有丝分裂后形成两个⼦核。
其中⼀个核移到细胞质稀疏的⼀端,发育成____⽣殖核______。
另⼀个核移到细胞质稠密的⼀端,发育成_____营养核_____。
6、将外源基因导⼊植物细胞的主要⽅法有____农杆菌转化法__________和_____基因枪__________。
7、性状伴随Y染⾊体的遗传属_____限性_______遗传。
8、⽟⽶的两对同源染⾊体,分别载有Aa和Bb基因。
这杂种细胞有丝分裂末期到达⼀极的染⾊体含_________AaBb___________基因,到达另⼀极的染⾊体含_____ AaBb ____________基因。
9、植物杂种优势常利⽤三系配套,这三系指___不育系________、____保持系_____和____恢复系_________。
10、已知两对独⽴遗传的⾮等位基因A,a与B,b,并具有互作关系。
如果基因型、表现型如下:基因型表现型A_B _ 甲性状A_bb ⼄性状aaB_ ⼄性状若aabb表现丙性状,则基因____A____对基因_____B___为____积加____作⽤。
11、普通⼩麦(AABBDD)×提莫菲维⼩麦(AAGG)的F1在减数分裂前期I可形成_____7______个⼆价体和____21_______个单价体。
普通遗传学复习题及答案

一、单项选择题1. 下列关于遗传学研究任务的各项描述中,错误的是【...A. 阐明生物遗传和变异的现象及其表现的规律B. 探索遗传和变异的原因及其物质基础,揭示其内在规律C. 证明生物性状表现受遗传因素控制,与环境因素无关D. 指导动植物、微生物的育种实践,提高医疗卫生水平 2. 达尔文的进化理论认为【A. 环境条件是生物变异的根本原因B. 自然选择是生物变异的根本原因C. 生殖隔离是生物进化的主要动力D. 自然选择是生物进化的主要动力3. 原核细胞结构较简单,下列结构中原核生物所不具备的结构是【 A. 细胞壁 B. 细胞膜 C. 核糖体 D. 染色体4. 观察鉴定染色体数目和形态的最佳时期是细胞分裂【 A. 间期B. 前期C. 中期D. 后期5. 真核细胞的染色质主要化学组成包括【A.RNA 、组蛋白和非组蛋白B.RNA 、组蛋白、非组蛋白和少量DNAC.DNA 、组蛋白和非组蛋白D.DNA 、组蛋白、非组蛋白和少量RNA6. 细胞分裂中期,染色体【A. 呈松散状态分散于细胞核中B. 呈松散状态分散于细胞质中C. 收缩到最粗最短分布于细胞核中D. 收缩到最粗最短分布于赤道板上 7. 一对同源染色体【A. 在细胞分裂间期配对平行排列B. 来源于生物同一亲本C. 形态和结构相同D. 总是带有相同的基因 8. 栽培大麦(2n=14)有丝分裂中期细胞内具有的染色单体数目为【A. 14条B. 7条C. 28条D. 42条9. 减数分裂前期I 可被分为5个时期,依次是【A.细线期一粗线期一偶线期一双线期一终变期B.细线期一偶线期一粗线期一双线期一终变期C. 终变期一细线期一粗线期一偶线期一双线期D. 偶线期一双线期一细线期一终变期一粗线期 10. 在形成性细胞的减数分裂中,染色体数目的减半发生在【 A.前期I B.前期II C.后期I D.后期II11. 高等植物的10个花粉母细胞可以形成40个【 A. 胚囊B. 卵细胞C. 花粉粒D. 精核12. 生物的主要遗传物质是【普通遗传学复习题及答案】】】】】】】】】】】A. DNAB. RNAC. 蛋白质D. 酶13.紫外线诱发生物突变最有效波长和DNA最大吸收紫外线光谱分别是【】A. 280nm 与280nmB. 280nm 与260nmC. 260nm 与280nmD. 260nm 与260nm14.分析某核酸样品,其碱基比率为27% A、23% C、27% T、23% G,该核酸是【】A.单链DNAB.双链DNAC.单链RNAD.双链RNA15.DNA半保留复制的含义是新合成的DNA分子中【】A.保留亲代DNA分子一半的碱基B.保留亲代DNA分子一半的核糖C.保留亲代DNA分子一条单链D.保留亲代DNA分子半条双链16.反转录酶催化的过程是【】A. DNA f RNAB. RNA^DNAC. rRNA f DNAD. tRNA f mRNA17.遗传信息传递的中心法则表明【】A. 细胞内遗传信息传递是单向的B.细胞内DNA只有通过自我复制产生C.细胞内RNA分子只能由DNA转录产生D.细胞内蛋白质只能由RNA指导合成18.下列关于遗传密码特征的描述中,哪一个是错误的【】...A.三个碱基决定一个氨基酸B.真核生物内显子的存在表明遗传密码可以是非连续的C.多数氨基酸可以由两种或两种以上的密码子编码D.病毒的遗传密码和人类核基因的遗传密码是通用的19.生物在繁殖过程中,亲子代之间传递的是【】A. 不同频率的基因B. 不同频率的基因型C. 亲代的性状D. 各种表现型20.豌豆种皮色中有灰色和白色,灰色(G)对白色(g)为显性。
小麦单株穗整齐度的遗传性观察
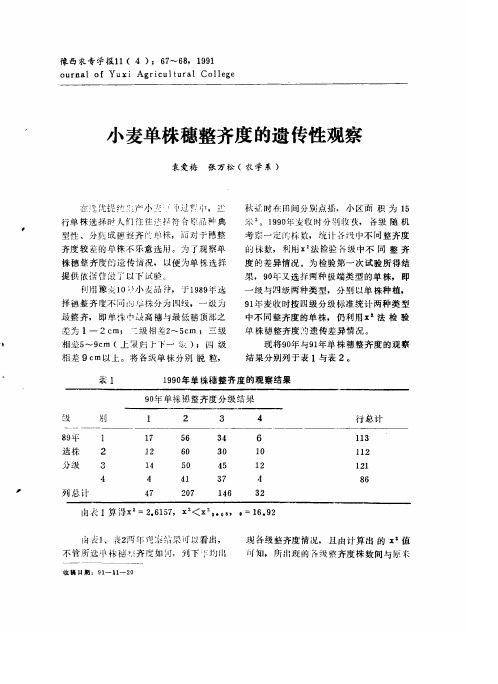
2
102 67
J. 山 1 ó 孟 J .ó ù 怪 n U An U 1
4
人 1 。 曰 , O
3 行总计
J
2
4
2
25
l
0 30
株分 级
列 总计
1
0 3
x
Z
1 69
由表 2 算 得
=
, 4 7 15
。
x
Z
<
二
2
。。
。。 ,
。
=
7
。
81
所选 的 各 级 间 无 相 关 ( 而 发生 变 化
。
x
Z
<
x :。。。。 Nhomakorabea)
,
即
况 下 单 株 发育 良好 分 孽 成 穗较 多
,
,
,
,
往往
所 出 现 的 各 级 株 数并不 随原来 级 数 的 不 同 这 表 明单 株穗 整齐度不 遗传
。
会 出 现 分 粟穗 与主 茎 穗 或 分孽穗 之 间 穗层 不整 齐 的现 象 的整 齐情 况
。
但 这 并不 影 响下 代 单 株 穗
与表
2
。
哀
株 穗 整 齐 度 的 观 察结 果
0 年 单株 穗 整 齐度 分 级 结果 9 别
8 9平
月 口 任 Q O J
1 4 1 4 1 4 2 7
2
尸 三 L 一 任 八 O 工 O 八 甘 0 , 凡 八 U 匕 n U
3
月 . 八 0 上 月 . 任 任 ó 任 1 月 I t J 3 3
l
o
p r n
e n
t
一个春小麦单体材料的初步鉴定

版I 6o第二次减数分裂后期、末期每个同 I ) ,
源染色体的两个染色体分开, 并趋向两极; 而游 离的染色体行动迟缓,处于细胞中间的赤道板
附近 ( 图版 1 7 , 1 )最后趋于一极,游离在大核 , 周围, 形状卷曲, 头尾相接, 在大核附近形成微
叶植物中极为少见, 并且在形成花粉粒时, 有的 核质多, 形成巨大小抱子, 有的核质少, 形成很
17
高秆、 中秆和矮秆三种类型( 图版1 3。 经细 , ) 胞学检查证明: 恢复正常的植株 (n 属高秆 2) 类型, 单体植株(n一 11 2 ) l中秆类型, 缺对体植 株 (。 2 属矮秆类型。 2一 ) 其分离比率如表 2 ,
表 2 单 体值株后代分离情况
左右, 茎秆较粗, 穗形似阿勃, 结实率恢复正常,
核( 图版1 80 1 ) ,
表 4 赤霉酸对 单体值株的影响
补} :处 理 有数高穗总数实 粒 穗 种 千 效 (米 厘 ) 小 小穗 穗 小高 子粒 穗株 长 穗不 数 最 满 (重 - t\ 厘 )(米 数 粒 饱度 克 数 )
H3 - -1, - 5, 7叶 期 喷 水 二 次 1 8 3. 6 4 2. 1 7 0. 1 2 9. 0 8 7 6. 2 1 5 6 0.
次 6阿 体 5 勃单 0 8
株 高( 米) 厘 成 熟 期 芒 穗 穗 形 色 形 105 0. 8月 1 0日 长 白 纺 160 0. 8 1 月 9日 无 白 纺 8. 55 8月 1 6日 无 自 纺
换法,可以有目的地将携带所需基因的染色体 由一个品种转移给另一个品种, 使育种目 标更
籽粒饱满度一级, 千粒重斗克左右, 7 有效分葵率
也有所提高。次年不再分离, 株高及其他性状
普通小麦与Elymus rectisetus异附加系的分子细胞遗传学鉴定

Vol 132,N o 112pp 11898-1901 Dec 1,2006作 物 学 报ACT A AG RONOMIC A SI NIC A 第32卷第12期2006年12月 1898~1901页普通小麦与Elymus rectisetus 异附加系的分子细胞遗传学鉴定王长有1 吉万全3,1,2 张改生1 王秋英1 薛秀庄1Ξ(1西北农林科技大学农学院,陕西杨凌712100;2陕西省农业分子生物学重点实验室,陕西杨凌 712100)摘 要:Elymu s rectis etus (Nees in Lehm)A.Love et C onn or 是目前小麦族中发现的唯一的无融合生殖种,为了鉴定和标记从普通小麦与E.rectisetu s BC 2F 2衍生后代中选育的2n =44株系1026A1、1057A 1和1035A2的外源染色体,应用细胞学、基因组原位杂交和RAPD 方法进行了研究。
经细胞学鉴定,3个株系花粉母细胞减数分裂中期Ⅰ(P MC M Ⅰ)染色体构型均为2n =22Ⅱ,与普通小麦Fukuh ok omugi 杂交F 1的P MC M Ⅰ染色体构型均为2n =21Ⅱ+1Ⅰ,两两杂交F 1的PMC M Ⅰ染色体构型均为2n =21Ⅱ+2Ⅰ,表明它们是分别附加了1对互不相同外源染色体的普通小麦2E.rectisetu s 二体异附加系。
标记E.rectisetus 品系1050的基因组DN A 为探针DN A ,对3个异附加系进行原位杂交,分别鉴定出附加的1对 E.rectisetu s 染色体。
应用13个引物对2个亲本和3个异附加系进行RAPD 分析,获得了可分别用于检测1026A 1和1057A 1中所附加的E.rectisetus 染色体遗传物质的分子标记OP B 214900bp 、OPE 209750bp 和OPB 2141000bp 。
关键词:Elymu s rectis etus ;异附加系;普通小麦;基因组原位杂交;RAPD中图分类号:S 512Molecular Cytogenetics Identi f ications on Triticum aestivum 2Elymus rectisetus A lien Addition LinesWANG C hang 2Y ou 1,J I Wan 2Quan 3,1,2ZHANG G ai 2S heng 1,WANG Qiu 2Y ing 1and X UE Xiu 2Z huang 1(1C ollege of Agronom y ,N ort h west S ci 2T ec Univers ity of Agriculture and F ores try ,Y angl i ng 712100,S haanxi ,China ;2K ey Lab oratory for M olecular B iol ogy of Agricultureo f S haanxi P rovince ,Y angling 712100,Shaanxi ,C hina)Abstract:The alien addition lines of T.aestivum can be used to breed the alien substitution lines and the alien translocation lines ,to locate the alien genes on the specific chr om osome ,and to construct the alien chrom osom e DNA library.Elymus rectisetus (Nees in Lehm)A.L o ve et C onnor is the only species known with apom ixes in the tribe of T riticeae.Three 2n =44lines 1026A1,1057A1and 1035A2wer e derived fr om [E.rectisetus ×Fukuhok omugi (T.aestivum)]BC 2F 2progeny.C ytogenetic ,genomic in situ hybridization (G ISH)and RAP D were applied to identify and tag E.rectisetus chr om os omes of three lines.Chrom osom e con figurations at P MC M Ⅰof three lines were 2n =22Ⅱ.C hr om os om e con figurations of hybr id F 1between three li nes and F ukuhokomugi were 2n =21Ⅱ+1Ⅰ.C hr om os om e con figuration of half diallel cross F 1between three lines were 2n =21Ⅱ+2Ⅰ.T he pr obe was total genom ic DNA of E.rectisetu s accession 1050labbe lled with BioNick T M Labe ling S ystem and the block DNA was tota l genom ic DNA of Fukuhokom ug i.G ISH analysis dem onstrated that tw o E.rectisetus chr om os om e of three lines w ere de tected ,respectively.G en om ic DNAs was isolated fr om E.rectisetus accession 1050,Fukuh okomugi and thr ee lines ,and was used for PCR am plification w ith 13R AP D pr imers (OP B 208,OP B 214,OPE 209,OPF 205,OPF 209,OP J 205,OPK 203,OP N 212,OPP 220,OPS 212,OPT 220,OPZ 209and OPZ 211).OP B 214and OPE 209could am plify the E.rectisetus spec ific DNA bands OP B 214900b p and OPE 209750bp i n 1026A1.OP B 214could am plify the E.rectisetu s specific DNA bands OP B 2141000b p in 1057A1.The results of cy togenetic and GI SH analysis showed that 1026A1,1057A1and 1035A2lines were T.aestivum 2E.rectisetus alien disomic addition lines with dif ferent E.rectisetu s chr om osomes.The results o f R AP D am plification showed that OP B 214900bp ,OPE 209750b p and OP B 2141000b p c ould be used as m olecular m arkers to detect E.rectisetus chr omation in alien disom ic addition lines 1026A1and 1057A1,respectively.Ξ基金项目:国家自然科学基金项目(39870529);陕西省自然科学基金项目(2001S M06)。
普通小麦品种Alondra's遗传转化体系的建立

・
技术方法 ・
普通 小麦 品种Aln r。遗传转 化体 系 的建 立 o d aS
李 明浩’ ,陈炜’ 邢莉 萍’ 一 , ,肖进’ 王海燕’ 曹爱 忠’ 王秀娥 , , ,
’南京农业 大学作物 遗传 与种 质创新 国家重 点实验室 .南京 2 0 9 10 5 中国科 学院合肥物 质科学研 究 院。合肥 2 0 3 3 01
株,转化率 为05 .%。Aln r。遗传转化体系 的建立丰 富了小麦遗传转化 的基 因型,为小麦 品种 的转基 因改 良和在不 同背景 o das
下研究基 因的功 能奠定了 良好的基础 。 关键词 愈伤 诱导,幼胚,高效再生,小麦
李 明浩,陈炜 ,邢莉萍 ,肖进 ,王海燕 ,曹爱忠 ,王秀娥 (0 0 .普通小麦 品种Ao da 遗 传转化体 系的建立 .植物学报 2 1) ln r’ s
等, 0 0 2 0 ;王永勤等, 0 2。 以小麦幼胚、幼穗作为 20 )
外植 体进行组织培养 ,不仅愈伤组织诱导率较 高,而
且分化成苗 能力 也较强( 乔亚科等 , 0 2 宋 国琦等, 20;
2 0 ;覃建兵等 , 0 5 。 03 2 0 ) 小麦幼胚培 养植株 再生过程 往往 表现 出明显的基因型差异( 林毅等, 0 3 。基因 2 0) 型、 培养基及培养基和 基因型的相互作用对 小麦愈伤 组 织 的 形 成及 植 株 再 生 具 有 重 要 影 响 ( 碧 华 等 , 伍
20 : 0 1 余桂 荣等 , 0 3 。 同的培养基与激素配 比对 2 0 )不 小 麦幼胚愈伤 组织形成及 再生也有着 很大 的影 响( 安 海龙等, 0 2 。目前 的转基 因小麦一般仅局 限于扬麦 2 0)
18 5 、B b i 等 品种。因此建立适于 不同基因型小 o wht e
科学家利用基因组编辑在小麦中产生父本单倍体

VOL.45,No.11 November.2020
究员曹乃亮团队与湖南杂交水稻研究中心联合研制出荧光检测分选仪,可对“杂交水稻之父”袁隆平研发的第三代杂交水稻繁殖系种子进行高精度、大通量分选,助力我国第三代杂交水稻推广应用。
2020年11月2日,袁隆平领衔的第三代杂交水稻取得了双季亩产1530.76公斤的成绩,为确保我国粮食安全作出新的更大的贡献。
但推广背后有个难题。
在第三代杂交水稻育种过程中,科研人员需要将繁殖系种子中遗传工程核不育系种子和工程保持系种子精准区分,才能进行杂交制
一粒一粒地区分,一该团队与湖南杂交水稻研究中心共同研制出第三代杂交水稻繁殖系种子荧光检测分选仪,每小时可对20公斤种子精准分选,解决了制约第三代杂交水稻推广应用的瓶颈。
该成果于11月18日通过了由湖南省农学会组织邀请的专家组现场验收。
中国工程院院士、华南农业大学教授罗锡文等9名专家组成员称,该技术成果为第三代杂交水稻的应用奠定了坚实的基础,其成果水平达到同类研究国际领先水平,建议加快推动产业化进程。
( 来源:新华网 2020-11-24 http://www.xinhuanet.
com/fortune/2020-11/24/c_1126779659.htm )
Copyright©博看网 . All Rights Reserved.。
普通小麦–大赖草易位系T7BS

作物学报 ACTA AGRONOMICA SINICA 2013, 39(2): 191−197 /zwxb/ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@本研究由国家自然科学基金项目(30871519), 国家重点基础研究发展计划(973计划)项目(2009CB118304)和江苏省优势学科建设项目资助。
*通讯作者(Corresponding author): 陈佩度, E-mail: pdchen@第一作者联系方式: E-mail: chengqicui_1986@Received(收稿日期): 2012-07-08; Accepted(接受日期): 2012-10-05; Published online(网络出版日期): 2012-12-11. URL: /kcms/detail/11.1809.S.20121211.1614.008.htmlDOI: 10.3724/SP.J.1006.2013.00191普通小麦–大赖草易位系T7BS·7Lr#1S 和T2AS·2AL-7Lr#1S 的分子细胞遗传学鉴定崔承齐1 王林生1,2 陈佩度1,*1南京农业大学 / 作物遗传改良与种质创新国家重点实验室 / 细胞遗传研究所, 江苏南京 210095; 2河南科技大学农学院, 河南 洛阳 471003 摘 要: 大赖草7Lr#1S 染色体上携带赤霉病抗性基因, 将其导入普通小麦有助于增加赤霉病抗源多样性和选育抗赤霉病品种。
利用染色体C-分带、荧光原位杂交和分子标记技术从普通小麦–大赖草7Lr#1单体异附加系的花粉辐射后代中, 选育出易位系T7BS·7Lr#1S (NAU639)和T2AS·2AL-7Lr#1S (NAU640)。
经连续3年大田、温室赤霉病接种鉴定, 这2个易位系的赤霉病抗性均显著高于感病亲本中国春、感病对照绵阳8545和石麦4185, 为赤霉病抗病育种提供了新的种质资源。
小麦单体与缺体的研究和利用

过单体分析,鉴定出恢复系的各个恢复基因所 在的染色体,对进一步了解雄性不育与育性恢 复机制,解决育性恢复问题,提供了线索和依
F 单体 X京红 1 : 号 注意选择遗传性接 近京红 1 号的单体
4
21 1 1
F 单体 0 ‘
2 1-1 (-2 京 0 1-1 1 1 {
( 四)远缘杂交方面的利用 最近美国 Sa es r报道了“ 小麦染色体配对遗 传控制研究”确定了中国春配对抑制基因在染 ,
幼穗固定方法:卡诺氏液(5 9 另酒精: 冰醋 酸=31中, :) 固定 2 小时后, 4 换人 7 的酒精 0 中在冰箱内保存, 以待镜检。醋酸洋红( 加一点 醋酸铁矾苏木精) 染色, 压片法观察。 压片方法:根据小麦幼穗发育规律,找到 合适的花药,加一小滴染色剂,用镊子尖挤压 多次, 挤出花粉母细胞, 弃去花药壁和杂质, 显 微镜下初查,如果正是花粉母细胞减数分裂中 期I 终变期、 或后期 I 立即加上盖片, 时, 并增 添一点染色剂, 在酒精灯上稍加热烘烤, 烧掉胞 质, 压片后可看到深红色的染色体。 ( 二)单、 缺体正确可靠性的鉴定 由于单价体的不稳定特性,利用中国春具 顶端着丝点单体品系, 鉴定单、 缺体的正确可靠 性是非常必要的。 具体方法为:() 1受查单体
红1 号单体品系)
20 1 1
色体, () 3 (, 3 S , 和4 B , S f, (, L D l A B ) ) D上; 配对促进基因在染色体 5 ( , 5 () 5 D S, S, L ) B A
( ) 3 ( ) 2 () D , , ( , L , L , S 、3 ( a 3 L a B A L ) A ) 某品种的某一优良特性受某一基因 控制后, 用 上。去掉配对主要抑制基因 5, B 提高配对促进 带有这个基因的染色体去替代另一个品种相应 基因 5 D的剂量, 为提高种间、 属间杂交配对率 的染色体, 从而使某品种的优良特性转移上去。 提供了理论依据。 此外, 还有人利用中国春单 体品系与黑麦、 冰草、 山羊草、鹅冠草等杂交, 京红 1 5 号 D单体X 某品种( 高蛋白) 杏 也都取得一定成效。 京红 1 5 单体 XF (0 1 D某) 号 D , 1+5 2 奋 总之, 小麦单、 缺体的研究已经广泛引起了 京红 1 5 单体 )F 号 D (x 育种工作者的注意和兴趣, 利用单、 缺体进行染 y
普通遗传学实验二-小麦单体

2.
3. 4. 5. 6.
染色约5分钟,加盖玻片。
用吸水纸包住载玻片,用大拇指垂直向下压片。 显微镜下观察,中期Ⅰ-四分体时期的细胞学特征 并绘图。
六、实验结果
1.
2.
3.
要求观察小麦单体花粉母细胞减数分裂中期 Ⅰ-四分体时期的细胞学特征。 绘出所观察到单体减数分裂各个时期的细胞 图(至少4个时期),并注明单价体的行为 特征(如落后染色体或微核)。 统计单体后代单体与双体的比例
普通遗传学实验二
小麦单体细胞学制片与观察
一、实验目的
1. 2.
了解单体减数分裂中染色体的行为特征。 掌握单体材料的细胞学鉴定方法。
二、实验原理
单体植株的减数分裂染色体行为与整倍体有所不同,由 于单体中有一条染色体缺少同源染色体与之配对,这条 染色体一般以单价体的形式存在。 单价体具有减速运动的特点,即单价体的行为落后于二 价体,形成落后染色体或微核。 有些单价体在第一次减数分裂时发生分裂或产生错分裂 (横裂)。错分裂程度依品种不同而异的类型,有端体 型和等臂型两种。 第一次分裂时未分裂的单价体到第二次分裂时可正常纵 裂,已分裂的单价体到第二次分裂时会发生再次等分裂 和错分裂,并有减速特点。
3.
小麦挑旗期
小麦的小花
单体幼穗
双体幼穗
小麦单体与双体的穗型比较
四、实验用品
1. 2.
试剂:卡宝品红染色剂 仪器及器具:显微镜、镊子、解剖针、载 玻片、盖玻片等
五、实验方法与步骤
1.
取幼小花蕾置载玻片上,用解剖针剥取长大约 1mm的花药2-3枚。
加1滴卡宝品红染色剂后,用镊子充分挤压花药, 使其内的花粉母细胞释放出来。 将大片的残渣去掉后。
普通小麦的端着丝点染色体(续完)

普通小麦的端着丝点染色体(续完)的报告,600字
小麦是一种非常重要的农作物,而用染色体端着丝点可以有效地检测其遗传基因。
本报告来自对一种普通小麦采用染色体端着丝点技术进行检测的研究结果。
该样本小麦属于雅怀特 8 与欧洲亚洲高等植物中的细粒面瘤小麦种类的结果。
为了进一步检测,小麦植株使用五种外源
DNA探针试剂向根部及茎部基因组进行染色体端着丝点检测。
染色体端着丝点技术的本质是用DNA探针试剂和植物减数分
裂中的基因组染色体进行匹配,在染色体的端着丝点上观察到基因组物质变化,从而确定其遗传基因,进行遗传基因分析。
结果显示,普通小麦中的染色体有三条,分别是第一条,第三条和第五条染色体,在染色体端着丝点检测中,有四个主要的基因类别,即噻唑类、抗虫类、抗病类和光合能力类。
在噻唑类基因中,第一条染色体的染色体端着丝点检测结果显示具有抗旱和耐高温能力;第三条染色体的染色体端着丝点检测结果显示具有抗病毒性和耐寒冷能力;第五条染色体的染色体端着丝点检测结果显示具有抗水分丢失和抗肥力能力。
此外,在抗虫、抗病和光合能力的基因类别中,普通小麦的染色体端着丝点检测结果显示普遍存在一定水平的较低抗性,但有一定的改进空间。
由此可见,通过染色体端着丝点技术对普通小麦的遗传基因进行检测,为普通小麦的遗传育种提供了可靠的数据,有助于提高小麦作物的生产效率和质量。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
五、实验方法与步骤
1.
取幼小花蕾置载玻片上,用解剖针剥取长大约 1mm的花药2-3枚。
加1滴卡宝品红染色剂后,用镊子充分挤压花药, 使其内的花粉母细胞释放出来。 将大片的残渣去掉后,加盖玻片。
2.
3. 4. 5. 6.
染色约5分钟。
用吸水纸包住载玻片,用大拇指垂直向下压片。 显微镜下观察,中期Ⅰ-四分体时期的细胞学特征 并绘图。
2.
中国春小麦5A单体是我们教研室已故的余 毓君教授在科研中发现的,5A单体的穗型 与双体有明显的差异,这样使我们很容易 从田间通过穗型的差异直接获得单体材料 (而中国春小麦的其它20个单体材料在穗 型上与双体没有差异)。
单体幼穗
双体幼穗
小麦单体与双体的穗型比较
四、实验用品
1. 2.
试剂:卡宝品红染色剂 仪器及器具:显微镜、镊子、解剖针、载 玻片、盖玻片等
六、实验结果
1.
2.
要求观察小麦单体花粉母细胞减数分裂中期 Ⅰ-四分体时期的细胞学特征。 绘出所观察到单体减数分裂各个时期的细胞 图(至少4个时期),并注明单价体的行为 特征(如落后染色体或微核)。
错分裂的类型
端体型
Hale Waihona Puke 等臂型中期Ⅰ
后期Ⅰ
末期Ⅰ
中期Ⅱ
后期Ⅱ
末期Ⅱ
四分体
提示
本实验是通过花粉母细胞减数分裂中染色 体的行为特征鉴定单体 但我们应该知道,也可以通过采用体细胞 染色体数目计数的方法,鉴定单体材料。
三、实验材料
1.
实验材料:中国春小麦5A单体(2n=42-1) 的幼穗
普通遗传学实验二
小麦单体细胞学制片与观察
一、实验目的
1. 2.
了解单体减数分裂中染色体的行为特征。 掌握单体材料的细胞学鉴定方法。
二、实验原理
单体植株的减数分裂染色体行为与整倍体有所不同,由 于单体中有一条染色体缺少同源染色体与之配对,这条 染色体一般以单价体的形式存在。 单价体具有减速运动的特点,即单价体的行为落后于二 价体,形成落后染色体或微核。 有些单价体在第一次减数分裂时发生分裂或产生错分裂 (横裂)。错分裂程度依品种不同而异的类型,有端体 型和等臂型两种。 第一次分裂时未分裂的单价体到第二次分裂时可正常纵 裂,已分裂的单价体到第二次分裂时会发生再次等分裂 和错分裂,并有减速特点。