3蛋白转导结构域介导乙型肝炎病毒聚合酶末端蛋白重链可变区抗体体外抑制病毒的复制
3抗体

4、外分泌液中的IgA称分泌型IgA,主要是二聚体。
5、SIgA的分子结构中有J链和SP。SP是粘膜上皮细胞
的多聚Ig受体的胞外区。
(二)生理功能
1、异物入侵的第一道防线。重要的局部免疫 作用。可有效的凝集细菌、中和毒素。
2、SIgA是皮肤和粘膜表面局部抗感染的重要 因素(如免疫屏障、中和病毒及抑制病毒 复制的作用)。
2.免疫球蛋白(immunoglobulin, Ig)
具有抗体活性或化学结构与抗体相似 的球蛋白统称为免疫球蛋白。 Ig = Ab + 化学结构类似物 Ab = Ig Ig ≠ Ab
二、免疫球蛋白的分子结构
(一)基本结构 由二硫键相连的四条对称的多肽链构成 的单体。形成一 “ Y ” 字形结构。 重链 ( heave chain, H ) 轻链 ( light chain, L )
不同Ig分子在超变区结构上各自具有独特的 氨基酸排序和构型特点,也称为Ig分子的独 特型或独特型决定簇。 超变区、互补性决定区和独特型决定簇指的 是Ig分子中的同一结构部位,只是从不同的 角度阐述而已。
⑵ 骨架区(framework region,FR)
Ig分子V区超变区之外的部位,其氨基酸组
IgG3 IgG4
IgG5
(三)生理功能
1、中和毒素和病毒。 2、凝集作用与沉淀作用。 3、激活补体,与补体结合的能力依次是: Ig G3、Ig G1、Ig G2。 Ig G4不固定补体. 4、亲细胞功能。FcR与多种细胞结合,裂 解靶细胞。 FcRI FcRII FcRIII 5、免疫损伤:新生儿溶血、输血反应等。
1.有利于阐明Ig的结构及生物活性 2.有利于生物制品的生产和纯化,避免超敏反应 的发生。
《乙肝两对半》

婴儿期感染 85%~95 % 成年期感染 5%~10% 慢性HBV感染
慢性乙型肝炎
5年发生率12%~25%
肝硬化
5年发生率5%~15%
5年发生率20%~23%
肝细胞癌
肝功能衰竭
肝移植
精选课件
慢性HBV感染的自然病程
免疫耐受 免疫清除 非活动或低复制期 再活动
HBeAg HBV DNA
精选课件
HBV的抗原抗体系统HBeAg和抗HBe1
HBeAg: 是功能蛋白。是临床上表达病毒复制较实
用的血清标志物。
临床意义:在急性HBV感染HBeAg仅存在于感
染的早期,在病变极期之后HBeAg消失,持续存在
者预示趋向慢性。
在AsC,HBeAg是免疫耐受的调节因子,在感染
的较前阶段HBeAg持续存在,是免疫耐受的高感染
精选课件
短链的5'-末端通过长达250-300个核苷酸 的碱基配对而维持分子的环状结构。 DNA多聚酶作用不断延长短链3′端以修 补缺口。缺口可能与HBV的DNA在感染 细胞内的整合有关。
总而言之,Dane颗粒是一个完整的乙肝 病毒颗粒,存在于乙肝病毒携带者或乙 肝病人的血清或肝细胞中,它只能在电 子显微镜下才能被看到,临床上我们检 测乙肝病毒的抗原、抗体以及它的DNA 来判断是否感染了HBV。
HBsAg(-)后数月才可检出。6-12个月后逐步达到高峰,持续多 年后逐步下降。部分人终身不产生抗HBs。
阳性表示:感染HBV后病毒清除形成对HBV免疫力;对疫苗的
免疫应答;或HBIG的被动免疫。
单一低滴度的抗HBs可以是假阳性。国内一期反应夹心法,国外 二期反应夹心法假阳性率较低。
HBV感染时间久,可能表现为HBsAg(-)/抗HBs(+)慢性肝病(隐 匿性乙型肝炎)。抗HBs(+)的肝硬化少数仍由乙型肝炎进展而来; 30%乙型肝炎肝硬化进展的HCC可抗HBs(+)。
生物体内的免疫传感器结构和功能分析

生物体内的免疫传感器结构和功能分析免疫传感器是一种专门用于检测外来物质的生物分子。
它们通常以蛋白质的形式存在,可以识别和结合细菌、病毒、真菌等各种外来微生物及其代谢产物、毒素等,进而触发免疫反应。
在免疫系统中,免疫传感器起着至关重要的作用。
本文将着重分析免疫传感器的结构和功能。
一、免疫传感器的结构免疫传感器可以分为三类:抗体、受体型Toll样受体(TLR)及其配体、NOD 样受体(NLR)及其配体。
它们的结构都含有感受外源性或内源性物质的结构域,分别是抗体的可变区、TLR的TIR(Toll/IL-1接受体亚家族结构域)结构域、NLR 的NACHT结构域。
1. 抗体抗体是一种高度特异性的免疫球蛋白分子,内含有微观世界中最完美的受体结构之一。
它的可变区由四肽链组成,包括两条轻链和两条重链。
每条链都含有可变的N端和连续的C端。
可变区有百万种不同的组合方式,使得抗体识别和结合不同的抗原物质。
抗体的Fc区则是固定免疫巨噬细胞和自然杀伤细胞,促进其发挥巨噬和杀伤作用。
2. TLRTLR通常存在于免疫细胞表面,识别和结合各种病原微生物和它们的产物,如内毒素、DNA、RNA等。
其分子结构包括胞浆侧的TIR结构域和细胞外侧的相应配体结构域。
TIR结构域是Toll样受体家族共有的结构域,具有广泛的功能,包括激活适当的信号分子、介导不同的信号通路、调控免疫细胞的生物学作用等。
3. NLRNLR主要包括NACHT结构域、LRR结构域和NOD结构域。
NACHT结构域是NOD样受体家族共同的核苷酸结合区域,包括甲基化NACHT、N-末端NACHT和全长NACHT三种类型。
NACHT结构域在介导免疫细胞活化和调控中都起到了重要的作用。
LRR结构域则位于NACHT结构域和NOD结构域之间,具有受体结构域的特征。
二、免疫传感器的功能免疫传感器的功能是通过检测外来物质和内源性损伤信号,引起免疫反应,维护机体的生命安全。
免疫传感器识别外来物质的方式包括两种:抗原特异性识别和抵抗性通用识别。
基因工程名词解释

名词解释一RNase:RNA水解酶Restriction endonucleasr:限制性核酸内切酶RBS:核糖体结合位点SD sequence:SD序列。
可结合原核生物的核糖体。
Ori:复制起始原点Promptor:启动子Klenow fragment:大肠杆菌pol1的大片段Reverse tranecriptase:反转录酶Transferred DNA:转移DNAMCS(multiple cloning site):多克隆位点IPTG: 异丙基-β-D-硫代半乳糖苷X-gal:5-溴4-氯-3-吲哚-β-D-半乳糖苷GUS:β-葡萄糖苷酸酶X-gluc:5-溴4-氯-3-吲哚-β-D葡萄糖苷酸酯Ampr(ampicillin resistance gene):氨苄青霉素抗性基因Cmr:氯霉素抗性基因Tetr:四环素抗性基因Kanr:卡那霉素抗性基因Ermr:红霉素抗性基因Neor:新霉素抗性基因supF:琥珀突变抑制基因phagemid:噬菌粒plasmid:质粒YAC ( yeast artificial chromosome):酵母人工染色体BAC(Bacterial Artificial Chromosome):细菌人工染色体PAC(P1 artificialchromosome):P1人工染色体TEL: 端粒重复序列CEN:着丝粒ARS: 自主复制序列Cosmid:黏粒PCR(Polymerase Chain Reaction):聚合酶链式反应dNTP:脱氧三磷酸核苷RT-PCR:反转录(reverse transcriptase)PCR或实时定量(real-time)PCR DD(RT)-PCR:差异(反转录)显示PCRTAIL PCR:热不对称相错PCRRACE: cDNA末端的快速扩增RAPD:随机扩增多态性DNAAFLP:扩增片段长度多态性SSH:抑制性扣除杂交FISH:荧光原位杂交Vector:载体Blunt end:平末端Match end/cohesive end:匹配黏端/黏性末端Deoxyribonuclease:脱氧核糖核酸酶TAP:烟草算焦磷酸酶SDS:十二烷基磺酸钠PAGE:聚丙烯酰胺凝胶电泳PFGE:脉冲电场凝胶电泳PEG:聚乙二醇DEPC:焦碳酸二乙酯GFP:绿色荧光蛋白Competent cell:感受态细胞PNA:肽核酸Ptac:乳糖操纵子和色氨酸操纵子的杂合启动子GST(Glutathione S-transferase):谷胱甘肽-S-转移酶DDT:二硫苏糖醇Tag:标记蛋白Polyhis-6:六聚组氨酸肽名词解释二1 同裂酶(isoschizomer)识别相同序列的限制酶称同裂酶同尾酶(isocaudarner)许多不同的限制酶切割DNA产生的末端是相同的,且是对称的,即它们可产生相同的黏性突出末端。
蛋白质体外表达与进化技术

[8]JoobeurTetal.Plant,2004,39(3):283—297[9]FeuilletCetal.ProcNatlAcadSciUSA,2003,100(25):15253—15258[10]TanksleySDetal.TrendsGenet,1995,11(2):63—68[11]SchwartzDCetal.Cell,1984,37:67—75[12]王永军等.遗传学报,2004,31(1):87—90[13]杨立春等.中国农业科学,2004,37(1):65—71[14]MadsenLHetal.MolGenetGenomics,2003,269:150—161文章编号:1000-1336(2005)04-0333-03蛋白质体外表达与进化技术王长松刘友生陈燕平1(第三军医大学西南医院病理学研究所,重庆400038;1西南医院国际合作实验室,重庆400038)摘要:蛋白质体外表达与进化技术包括核糖体展示技术和mRNA展示技术。
与蛋白质体内表达系统相比,体外表达技术可产生较大容量的蛋白质文库(约1012~1013左右),同时在回收编码蛋白质的信息时对文库进行了进化,增加了蛋白质文库的多样性,促进了抗体或配体类蛋白质的亲和成熟能力。
该文介绍了蛋白质体外表达技术在新蛋白质(如抗体或配体等)表达筛选中的应用。
关键词:蛋白质体外表达;核糖体展示;mRNA展示;进化中图分类号:Q51;Q816———————————收稿日期:2005-05-16国家自然科学基金资助项目(No.30271342)作者简介:王长松(1972—),男,博士,主治医师,E-mail:goldenfinger850124@yahoo.com.cn;刘友生(1955—),男,博士,教授,博士生导师,联系作者,E-mail:youshengliu660@hotmail.com从20世纪80年代至今,产生了多种筛选肽或蛋白质文库的新技术,这些技术方法大多以表面展示来命名,如细胞表面展示、质粒展示和酵母双杂交系统等。
hbv各基因区编码产物 -回复

hbv各基因区编码产物-回复HBV(乙型肝炎病毒)是一种致病性病毒,是引起肝炎和肝硬化的主要原因之一。
HBV基因组由四个主要区域编码产物,分别是表面抗原区(S区)、核心蛋白区(C区)、转录调控区(X区)和编码酶及外衣位置区(P区)。
本文将一步一步回答HBV各基因区编码产物的相关内容。
首先,我们来讨论表面抗原区(S区)。
表面抗原区编码产物是HBV外表面的蛋白质,包括三种结构上不同但功能上相似的蛋白质,分别是S、M、L表面抗原。
其中,S抗原是最基本的,它存在于HBV颗粒和血液中,被用于乙肝病毒感染的诊断和治疗。
M表面抗原是通过编码S蛋白前面的预S蛋白质产生的,它在HBV感染早期主要是在内部颗粒中发现,但在血液中也可检测到。
L表面抗原是通过编码S蛋白后面的预S蛋白质产生,它在HBV感染早期主要是在内部颗粒中发现。
接下来是核心蛋白区(C区)。
核心蛋白是HBV颗粒的主要组分之一,也被称为核心抗原。
它在病毒生命周期中起多种重要作用,包括维持病毒颗粒的结构完整性、媒介病毒的复制和颗粒的装配。
核心蛋白的编码产物经过一系列的翻译和修饰过程形成最终的核心抗原蛋白,这些蛋白质能够诱导宿主免疫系统产生免疫应答。
第三个基因区是转录调控区(X区)。
X蛋白是HBV的主要转录调控因子,通过干扰宿主细胞的信号通路和基因表达来保持病毒的生存和复制。
X蛋白的编码产物也具有多种功能,包括抑制宿主细胞的DNA修复、干扰细胞周期和细胞凋亡、调节宿主基因的转录和翻译等。
X蛋白的过度表达和异常调控可能导致乙肝病毒感染的慢性化和癌变。
最后是编码酶及外衣位置区(P区)。
P区编码产物主要包括多种酶,如聚合酶、DNA依赖性的RNA聚合酶、RNA酶和DNA外切酶等。
这些酶在乙肝病毒的复制过程中起着重要作用,如聚合酶负责合成病毒RNA或DNA的互补链,RNA酶则参与RNA的处理和成熟,DNA外切酶参与病毒DNA的修复和整合。
这些酶的功能使得乙肝病毒具有高度的复制活性。
抗病毒药物课件

2.2.1 三氮唑核苷
• ..Ribavirin • 又名利巴韦林 • 1-β-D-呋喃核糖基-1,
2. 本药还有一定骨骼肌和心肌毒性,表现为肌痛 、肌无力、心电图异常,停药可恢复。
3. 其他不良反应有恶心、头痛、发热、疲乏等。 4. 使用本药时应定期查血象和心电图。
齐多夫定Zidovudine, ZDV
药物相互作用
1. 丙磺舒、美沙酮、氟康唑、丙戊 酸钠、苯妥英钠等可增高ZDV血 药浓度;
2. 氨苯砜、氟胞嘧啶、更昔洛韦、 抗癌药物可增强ZDV对骨髓的抑 制,故应尽量避免与其他有骨髓 抑制作用的药物合用。
细胞外
结合
HIV蛋白酶后期修饰 被PIs抑制
脱壳
编码RNA
逆转录
被
RTIs
抑 制
细胞内 双链DNA
整合
病毒蛋白
翻译 病毒mRNA
装配
细胞膜
编码RNA
细胞核
转录
病毒RNA
齐多夫定Zidovudine, ZDV
不良反应
1. 可引起骨髓抑制,表现为白细胞或红细胞减少 、贫血,发生率与用药剂量和疗程有关,多发 生在连续用药6~8周。其骨髓抑制作用可能与 一磷酸ZDV可竞争性抑制细胞胸苷激酶有关。
阿昔洛韦的代谢
O
HN
N
H2N N N O
OH
O HN H2N N
O HN H2N N
N O
NO
OH
E3泛素连接酶三基序25的研究进展▲

E3泛素连接酶三基序25的研究进展▲任伟;王志维【摘要】E3泛素连接酶三基序25(TRIM25)是E3泛素连接酶中三基序蛋白家族的成员之一,在天然免疫、防御病毒感染、调控细胞增殖和癌细胞迁移中起主要作用.研究表明TRIM25也能够结合RNA并调节Lin28a介导的let-7前体尿苷化.TRIM25作为一种新型蛋白在子宫发育、肿瘤发生发展、天然免疫和RNA代谢中发挥重要作用,本文将对其上述作用进行综述.【期刊名称】《广西医学》【年(卷),期】2018(040)021【总页数】4页(P2589-2592)【关键词】三基序25;E3泛素连接酶;子宫发育;肿瘤;天然免疫;RNA代谢;综述【作者】任伟;王志维【作者单位】武汉大学人民医院心血管外科,湖北省武汉市 430060;武汉大学人民医院心血管外科,湖北省武汉市 430060【正文语种】中文【中图分类】R34E3泛素连接酶三基序(tripartite motif,TRIM)家族的成员超过70个,其成员的N末端均包含有1个RING结构域、B盒结构域和卷曲螺旋结构域,此结构有助于确定底物的可变C-末端特异性[1]。
TRIM家族蛋白质在人体内具有多种作用,包括调控天然免疫中的信号转导、防御病毒感染、调控细胞增殖和癌细胞的迁移。
研究表明TRIM25、TRIM28、TRIM56和TRIM71都能结合RNA,形成一个RNA结合的E3泛素连接酶池[2-3]。
E3泛素连接酶能催化结合其泛素部分的靶蛋白,根据泛素链的类型,E3泛素连接酶池可具有不同的功能。
其中研究最多的聚泛素链是K48和K11,其形成的多聚蛋白链可以通过26S蛋白酶体来降解靶蛋白。
同时也有研究表明其他聚泛素链如K63和单泛素,在信号传导、蛋白定位和调节蛋白-蛋白质相互作用中具有重要作用[4]。
而TRIM25可以催化更多的蛋白添加到K48和K63连接的多聚蛋白链上,在天然免疫反应中能够靶向结合支架蛋白14-3-3σ,从而起到降解病毒RNA的作用,同时还可作为针对病毒RNA的下游效应物影响信号转导。
hbv各基因区编码产物

hbv各基因区编码产物一、概述HBv是一种重要的逆转录病毒,也是乙型肝炎病毒的主要类型。
HBv基因组包含一个线形双链DNA分子,分为前基因组和病毒基因组。
HBv基因组编码产物主要包括病毒蛋白、病毒RNA和病毒DNA。
其中,病毒蛋白包括主要抗原蛋白(HBsAg)和病毒复制相关蛋白;病毒RNA包括病毒mRNA和病毒DNA的转录本;病毒DNA则是病毒的遗传物质。
二、各基因区编码产物详解1. 前基因区:前基因区包含病毒复制所必需的启动子和增强子序列,但不编码任何蛋白质。
2. 结构基因区:该区编码HBsAg,包括表面抗原(HBsAg)和HBeAg。
HBsAg 是病毒的主要抗原,可以引发机体免疫应答。
HBeAg与病毒复制和传播有关。
3. 包膜外膜蛋白基因区:该区编码HBcAg,是病毒复制的标志性蛋白,可以刺激机体产生中和抗体。
4. 核心基因区:核心基因区编码病毒DNA聚合酶、HBv复制相关的蛋白质等。
此外,还包括调节病毒复制的转录因子和DNA包装信号序列等。
三、各基因区编码产物的功能(1)HBsAg:引发机体免疫应答,识别并清除病毒感染细胞。
(2)HBeAg:与病毒复制和传播有关,可作为病毒复制活动的指标。
(3)HBcAg:刺激机体产生中和抗体,识别并清除病毒感染细胞,抑制病毒在细胞内的复制。
(4)核心酶:参与病毒DNA的复制和转录,调控病毒的复制过程。
四、各基因区编码产物的表达与调控各基因区编码产物的表达受到多种因素的调控,包括病毒复制状态、宿主免疫反应、环境因素等。
其中,核心基因区的转录和翻译受到病毒RNA的调控,而HBsAg和HBeAg的表达则受到宿主免疫反应的影响。
五、结语HBv各基因区编码产物在病毒的复制、传播和免疫逃逸等方面起着关键作用。
深入理解各基因区编码产物的功能和表达调控机制,对于开发有效的抗病毒药物和治疗策略具有重要意义。
同时,加强乙肝疫苗接种和疾病预防控制工作,提高公众对乙肝防治知识的认识,也是防控乙肝的重要措施。
医学微生物学重点知识总结
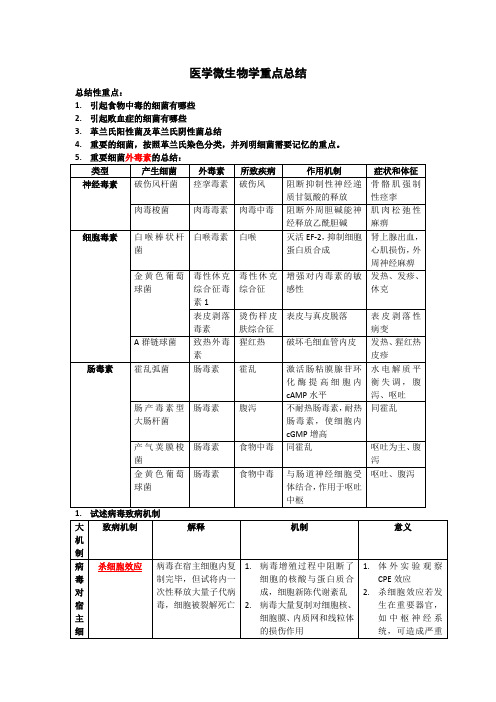
医学微生物学重点总结总结性重点:1.引起食物中毒的细菌有哪些2.引起败血症的细菌有哪些3.革兰氏阳性菌及革兰氏阴性菌总结4.重要的细菌,按照革兰氏染色分类,并列明细菌需要记忆的重点。
6.与口腔相关的厌氧菌:【1】脆弱类【2】放线菌1. 生物的概念:微生物是一类存在于自然界中,体型微小、结构简单、肉眼难见,而需要借助光学乃至电子显微镜放大成千上万倍才能观察到的微小生物。
3.微生物与人类的关系:总结:绝大多数微生物对人类和动物、植物有益,少数引起人类和动物植物病害。
微生物对人类的益处:1.营养作用:产生人类必须的一些营养物质,如VitK2.生物拮抗:人体正常菌群占据人类表皮和黏膜,使得致病菌难以粘附3.免疫作用:正常菌群的存在是维持人类免疫力的基础4.抗衰老作用:乳酸杆菌等在胃肠道的大量存在与长寿有明显对应性。
**科赫法则:1. 特殊病原菌应在同一疾病中查见,而健康人中不存在2. 该特殊致病菌能被分离培养得到纯种3. 该纯培养物接种至易感动物能产生同样疾病4.人工感染的实验动物中能重新分离出该致病菌纯培养。
第一章细菌的形态与结构***细菌的定义:广义:泛指各类原核细胞型微生物,包括细菌、放线菌、支原体、衣原体、立克次体、螺旋体。
狭义:专指其中数量最大、种类最多,具有典型代表性的细菌。
***对细菌的形态观察时机:选择:适宜生长条件下的对数生长期原因:细菌在不利不环境或者衰老时产生形状多形性,亦称衰退型。
1.微生物根据其形态结构组成的差异分为几大类?各有何特点?三类,分别是球菌、杆菌、螺形菌。
补充:【1】肽聚糖组成区别4.L型细菌的特点及医学意义。
【1】由来:当某些因素(如溶菌酶、青霉素等)破坏或抑制了肽聚糖的合成使细菌细胞壁发生部分或完全缺损,在高渗环境下仍可存活而成为细胞壁缺陷型细菌。
因首先在Lister研究所发现,故称L型细菌。
【2】分类:原生质体:G+菌细胞壁几乎完全缺失,仅剩一层细胞膜原生质球:G-菌因有外膜保护,且胞内渗透压较低,对低渗环境仍有一定抵抗力【2】特性:1.产生因素:青霉素、溶菌酶、胆汁、抗体、补体等2.形态:大小不一,高度多形性,革兰氏染色为阴性3.培养特性:高渗低琼脂培养基,营养要求高,生长缓慢,荷包蛋样菌落4.致病性:引发慢性感染【3】医学意义:1.L型细菌在体内、外均能形成,尤其在使用于细胞壁的药物治疗过程中反复出现,某些L型细菌仍有致病力,可引起尿路感染、骨髓炎、心内膜炎等疾病;2.在进行常规细菌学检查时,L型细菌往往被漏检而造成病原菌感染的漏诊。
scfv名词解释(二)

scfv名词解释(二)SCFV名词解释1. SCFV 介绍•SCFV,全称为单链可变片段 (Single-chain variable fragment),是转基因技术的产物,是源自抗体的小分子蛋白。
•抗体是一种由免疫系统产生的蛋白质,用于识别并结合体内外的特定抗原。
SCFV是抗体分子的重要组成部分,具有保留了抗体结合特异性的功能,同时拥有较小的体积和更好的组织渗透能力。
2. SCFV的结构•SCFV是由可变区域 (Variable region) 的轻链和重链连接而成的单一线性链,是一种单链抗体分子。
•可变区域包括了抗体结合抗原所关键的亚区域,负责与特定抗原结合并实现特异性识别。
•SCFV通常由经过基因工程技术合并的单链抗体模块生成,结构更简单,组装更方便,且具有较小的分子量。
3. SCFV应用领域•SCFV在生物医药领域具有广泛的应用前景,尤其在新药研发、诊断和治疗等方面。
•新药研发:SCFV可以通过对抗原进行高效的定向筛选,用于发现和优化具有高亲和力和特异性的新药分子。
•诊断:SCFV可以用于开发高灵敏度和高特异性的诊断试剂盒,用于检测疾病标记物、病原体和药物残留等。
•治疗:通过将SCFV结合到适当的药物或免疫治疗药物上,可以增强药物的靶向性和疗效,减少副作用。
4. SCFV应用案例Case 1: 新药研发•某生物制药公司正在开发一种治疗乳腺癌的新药。
他们使用基因工程技术构建了一种靶向乳腺癌细胞表面特定抗原的SCFV。
通过体外细胞实验和动物试验,确定这种SCFV具有较高的亲和力和特异性,能够有效地识别并杀死乳腺癌细胞。
这为进一步的临床研究奠定了基础。
Case 2: 诊断试剂开发•一家诊断试剂公司正在研发一款用于早期检测艾滋病的检测试剂盒。
他们利用基因工程技术制备了一种特异性与艾滋病毒抗原结合的SCFV。
实验证实,该SCFV能够高灵敏度地结合艾滋病毒抗原,并产生可观察的荧光信号。
这一发现为艾滋病的早期诊断提供了新的工具。
植物转基因抗性策略研究进展

126--农业经济•专题综述 DOI:10.16498/ki.hnnykx.2016.010.035自首次报道转基因植物表达植物病毒序列并表现抗病性以来,人们尝试了各种不同类型的抗性产生方法。
这些方法主要包括表达不同的病毒序列、非病毒序列、宿主来源的抗性基因及各种宿主防御反应因子等。
笔者主要围绕以上各种成分或序列介导产生的抗性展开综述。
1 病毒蛋白介导的抗性策略1.1 外壳蛋白介导的抗性外壳蛋白(Coat protein ,CP )介导的抗性方法主要通过在转基因植株表达病毒的外壳蛋白基因从而获得抗此种病毒或相关病毒的能力。
1986年Abel 等[1]将烟草花叶病毒(TMV )的外壳蛋白编码序列导入到烟草中,获得了具有TMV 抗性的抗病毒植株。
随后,科学家们先后将黄瓜花叶病毒(CMV )、马铃薯Y 属病毒(PVY )、辣椒重症花叶病毒(PepSMV )等的外壳蛋白基因导入烟草植株后,均得到了抗病毒植株。
外壳蛋白介导的抗性可能是蛋白质水平介导的干扰或RNA 水平的沉默的结果,也可能同时存在蛋白质和RNA 两种作用机制。
此外,当接种高病毒剂量或接种病毒RNA 时,外壳蛋白介导的抗性普遍存在容易被打破的共性[2-4]。
1.2 复制酶基因介导的抗性复制酶(Replicase )介导的抗性方法主要通过表达病毒复制酶通读序列、全长序列、突变序列及缺失序列获得具有抗病毒能力的植株。
1990年Golemboski 等[5]将TMV 的54KD 复制酶基因转入烟草植株,获得了高抗TMV 的转基因抗病毒植株。
研究表明,复制酶基因介导的抗性策略产生的抗性不具广谱性,接种非常高剂量的病毒时表现出强抗性,接种病毒RNA 时也表现出高水平的抗性[6],但是表达缺失型复制酶蛋白编码序列表现出广谱抗性[7]。
关于复制酶基因介导的抗性机制尚无定论。
多数早期的研究认为复制酶基因介导的抗性是由蛋白介导的,稍后的研究却发现存在RNA 介导的过程,也许复制酶基因介导的抗性在蛋白质水平和RNA 水平存在互补或替换的过程[6]。
hbv各基因区编码产物 -回复

hbv各基因区编码产物-回复HBV各基因区编码产物乙型肝炎病毒(HBV)是一种致命的病毒感染,广泛流行于全球范围内。
该病毒具有一个特殊的基因组,它可以编码多个产物。
在这篇文章中,我们将一步一步地回答关于HBV各基因区编码产物的问题,以帮助读者更好地了解这个病毒。
1. HBV基因组结构HBV的基因组是由一个环状DNA分子组成。
该基因组可以分为四个基因区域,分别是S(表达HBsAg的表面抗原)、C(编码核衣壳蛋白)、P (编码逆转录酶)和X(编码X蛋白)。
2. S基因区编码产物S基因区是HBV基因组中最重要的区域之一,它编码一系列的表面抗原(HBsAg)。
HBsAg是HBV诱发主机产生免疫反应的关键蛋白。
它是病毒颗粒的主要成分,并且可以用来检测HBV感染。
3. C基因区编码产物C基因区编码的产物是核衣壳蛋白(HBcAg)。
HBcAg是HBV中核心颗粒的主要成分,具有基本的抗原性。
它在病毒复制、装配和释放过程中起着重要作用,并且可以作为HBV感染的标志。
4. P基因区编码产物P基因区编码的产物是逆转录酶(RT),它是一个多功能的酶。
逆转录酶能够将病毒的RNA基因组逆转录成DNA,并插入到宿主细胞的基因组中。
这种逆转录过程是HBV感染和复制的关键步骤,因此逆转录酶在HBV感染中具有重要作用。
5. X基因区编码产物X基因区编码的产物是X蛋白。
X蛋白是一个非结构性蛋白,具有多种功能。
它能抑制宿主细胞的基因转录和蛋白合成,从而影响细胞周期和凋亡等过程。
此外,X蛋白还能干扰免疫反应,增加HBV感染的潜在性和病变。
总之,HBV基因组编码的产物在病毒感染的各个阶段起着至关重要的作用。
S蛋白和C蛋白是病毒的主要结构成分,通过它们,我们可以检测HBV感染。
P蛋白和X蛋白则在复制和感染过程中发挥重要的调控作用。
对HBV基因区编码产物的深入了解,有助于我们更好地理解该病毒的感染机制,并有助于开发新的治疗和预防策略。
希望这篇文章能帮助读者更好地了解HBV各基因区编码产物。
新冠病毒的RNA复制机制及抗病毒药物研发进展

新冠病毒的RNA复制机制及抗病毒药物研发进展新冠病毒,也被称为SARS-CoV-2,是一种具有高传染性的病毒,引发了全球范围内的COVID-19疫情。
了解新冠病毒的RNA复制机制以及抗病毒药物的研发进展对于控制疫情和治疗患者至关重要。
新冠病毒是一种正链单股RNA病毒,其基因组长度约为30,000个核苷酸,包含了几个重要的开放阅读框(ORF),编码了多种蛋白质。
其中,蛋白酶切割酶(proteases)、聚合酶(polymerase)以及血清素受体结合蛋白(S protein)等是RNA复制过程中至关重要的因子。
新冠病毒的RNA复制过程可以分为两个主要阶段:转化和复制。
在转化阶段,病毒进入宿主细胞,并释放其基因组RNA。
该RNA然后被翻译成多种蛋白质,这些蛋白质会进一步协助病毒进入复制阶段。
在复制阶段,由病毒聚合酶酶(RdRp)介导的RNA复制过程发生。
病毒RNA的复制是通过RdRp酶将RNA模板复制成新的RNA链实现的。
此过程需要多个辅助蛋白质的参与,其中包括RNA引物依赖性RNA聚合酶(primase)和RNA终止酶(nsp14-Exonuclease)。
了解新冠病毒RNA复制机制的关键过程为抗病毒药物的研发提供了重要的依据。
当下,研究人员正在积极寻找针对病毒复制的抗病毒药物。
一些已有的抗病毒药物,如瑞德西韦和氯喹,已被用于治疗新冠病毒感染的患者。
瑞德西韦是一种广谱抗病毒药物,通过抑制病毒RNA聚合酶来阻断病毒的复制。
临床试验显示,瑞德西韦对新冠病毒感染具有一定的疗效,可以减轻症状并缩短住院时间。
然而,它的疗效仍存在争议,并且需要进一步的研究来确定其在治疗中的作用。
氯喹是一种已有的抗疟药物,也在新冠病毒疫情暴发后被重新关注。
氯喹被认为具有抗病毒作用,可以阻止病毒进入宿主细胞。
然而,氯喹的临床疗效也存在争议,并且一些研究显示其可能对心脏产生不良影响。
除了已有的药物外,研发新的抗病毒药物也是当前的重要研究方向之一。
hbv基因组结构

hbv基因组结构
HBV基因组又称为HBVDNA,结构独特而精密,由不完全的环状双链DNA组成,其长度约为3200个核苷酸。
HBV基因组含有4个部分,分别是包膜蛋白基因(envelopeprotein)、核心蛋白基因(coreprotein)、转录酶基因(polymeraselargerase)和X基因(Xgene)。
1.包膜蛋白基因:该基因编码了S蛋白(表面抗原),是HBV 的主要表面抗原。
它由前S1、前S2和S区三个部分组成,其中前S1和前S2是HBV包膜蛋白的关键结构,前S2蛋白在病毒颗粒装配过程中起重要作用。
2.核心蛋白基因:该基因编码了C蛋白(核心抗原),是HBV 的核心成分。
它由前C区、C区和部分核心启动子区组成。
前C区编码了转氨酶,是HBV的生物合成过程中重要的酶之一。
3.转录酶基因:该基因编码了逆转录酶(RTase)和DNA聚合酶(DNAP),这两种酶是HBV生物合成过程中不可或缺的酶。
转录酶基因结构非常复杂,包括反转录和DNA聚合酶两个区域。
4.X基因:该基因编码了X蛋白(HBx),是一种多功能蛋白,具有调节转录、复制、细胞生长和凋亡等多种功能。
总之,HBV基因组结构复杂,各部分相互协作,形成了独特的病毒生命循环。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
工生物工程技术服务有限公司完成。 二、方法 1.PCA法选择特异性VH抗体:用PCA法
从酵母展示scFv抗体库中选择出HBV聚合酶 TP区特异性VH抗体[2|,实验证明该抗体在体外 可抑制HBV的复制[】]。
2.重组表达质粒pET28一TAT—VH的构建: PCR扩增与TAT连接的抗体TAT—VH。TAT 核苷酸组成:TATGGCAGGAAGAAGCGGAG— ACAGCGACGAAGA。正义引物为5’-TACC-
400038。China
Corresponding author:LAN Lin,Email:llin6624@hotmail.com
[Abstract]0bjective To study a functional variable fragment of heavy chain(VH)antibody
3.重组蛋白的表达和包涵体的纯化:以 pET28一TAT—VH原核表达载体转化大肠埃希菌 BL21(DE3)LysS,IPTG诱导融合蛋白表达,并利 用融合蛋白携有的(His)。标签,以Ni—NTA. agrose柱进行纯化。用Ppl0脱盐柱除去蛋白洗 脱液中咪唑等小分子杂质。纯化的TAT—VH融 合蛋白经十二烷基硫酸钠一聚丙烯酰胺凝胶电泳 (SD孓PAGE)分析后,采用凝胶扫描的方法测定 蛋白样品的纯度,BCA测定融和蛋白的浓度。
万方数据
24 h),在第1组的2个复孔中加入改良Eagle培 养基(DMEM)20 mL作为阴性对照。第2~6组 (每组2个复孔)均加入TAT—VH融合蛋白至终 浓度为100 mmol/L,分别在15 rain、30 min、1 h、 2 h和4 h,于冰上吸弃培养液,4℃PBS(pH 8.O) 洗涤各孔贴壁细胞10 min,每孔加含20 g/L多聚 甲醛、l g/L Triton X-100的PBS 1 mL,置冰上 30 min。再以4℃PBS洗10 min,加10 g/L的牛 血清白蛋白(BSA),置42℃10 min,4℃PBS洗 10 rain,每孔加鼠抗His抗体(1:1 000)500 ItL, 42℃30 rain后,4℃PBS洗15 min,加异硫氰酸 荧光素(FITC)标记的兔抗鼠IgG(1:1 000), 42℃温育10 min,PBS震洗1 h,以体积分数为 0.5的甘油封片,荧光显微镜下观察并摄片。
【关键词】肝炎病毒,乙型}转导,遗传;免疫球蛋白类,重链}免疫球蛋白可变区;病毒复 制I抗体
Variable fragment of heavy chain antibody against the terminal protein region of hepatitis B virus polymerase introduced by Tat protein transduction domain inhibits the replication of hepatitis B virus in
GAGCTCGGATCCATGATACCGTCCTTCTT(’- GCCTCTGTCGCTGCTTCTCAGGTGCAGCTG卜
CAp3’,反义引物为5'-CTGGACTAGTGGA— TCCTCATGAAGAGACGGTGAC_3 7。用BD In- FusionTM PCR克隆试剂盒作连接反应,将TAT. VH插入原核表达载体pET28a(+),即pET28- TAT—VH,测序鉴定。 in supernatant of HepG2.2.15 celI culture with 5 000 nmol/L TAT-VH was(1.211± 0.132)lg copy/mL,which was significantly lower than control group[(5.325士o.041)lg copy/mL, £=72.91。P<0.05].Meanwhile,the level of intracellular HBV DNA w越(3.521士0.411)lg copy/mL, which was significantly lower than control group[(8.532士0.132)Ig copy/mL.t=28.41,P< 0.os].C.omcausion The HBV replication is inhibited by anti-TP TAT—VH antibodies in vitro,which provides valuable experimemal basis for developing therapy of HBV infection with intracellular antibody.
【Key words]Hepatitis B virus}Transduetion,genetic Immunoglobulins,heavy chain} lmmunoglobulin variable region t Virus replication Antibodies
由于HBV系细胞内复制,传统体液免疫中的 抗体难以发挥作用。近年来,细胞内抗体的研究 给乙型肝炎的抗病毒治疗带来了希望。HBV聚 合酶末端蛋白(TP)区由177个氨基酸组成,有2 个抗原决定簇。在我们以前的研究中用蛋白片段 互补法(PCA)选择出具有特定功能的重链可变区 (VH)抗体,特异性VH抗体基因定向亚克隆入真 核表达载体pZeoSV2(+),构建pZeoSV2(+)一 VH重组质粒,用脂质体转染技术,将其导入 HepG2.2.15细胞。表明该抗体可抑制HBV的复 制[1]。传统的细胞内抗体技术存在两个问题:操 作复杂,动物实验更困难;如应用于人体可能存在 安全隐患。蛋白转导结构域的研究可能解决这个 问题。我们将蛋白转导结构域TAT与特异性 VH抗体基因连接,定向亚克隆入原核表达载体 pET28a(+),经异丙基一pn硫代半乳糖苷 (IPTG)诱导,在大肠埃希菌表达,纯化TAT.VH 融合蛋白,并将其直接导入HepG2.2.15细胞,研 究特异性TAT—VH抗体在体外对HBV复制的影 响,以期为慢性乙型肝炎的治疗提供新的手段。
against the terminal protein(TP)region of hepatitis B virus(HBV)polymerase introduced by human
immunodeficiency virus Tat protein transduction domain(TAT)and the inhibitive activity of TAT—VH
DNA level in HepG2.2.15 cell culture was measured using quantitative polymerase chain reaction (PCR).The data were analyzed by one-factor analysis of variance and t test.Results TAT.VH fusion protein was successfully expressed and purified.It was confirmed by IFA and MTT assay that TAT—VH was introduced into HepG2.2.15 cells and the eell growth was not affected.The level of
4.HepG2.2.15细胞的培养:细胞在含体积分 数为0.1的胎牛血清、含青霉素10万U/L、链霉素 10万U/L的RPMI 1640培养基,37℃,体积分数 为0.05的C02中用24孔培养板培养。每3天换 液1次,6 d传代。
5.TAT-VH跨膜转导效率的鉴定:将 HepG2.2.15细胞以2×108/L的浓度接种于内置 爬片的24孔细胞培养板。待细胞贴壁后(约
材料与方法
一、材料 BD In-FusionTM PCR克隆试剂盒购自美国 BD生物科学克隆技术公司,大肠埃希菌BL21 (DE3)LysS购自美国Promega公司,原核表达载 体pET28a(+)购自美国Novagen公司,HBV PCR荧光定量检测试剂盒由上海克隆生物高技术 有限公司生产,二辛可宁酸法(BCA)蛋白定量试 剂盒购自江苏海门市碧云天生物技术研究所,Ni- NTA—agrose购自美国基因公司,PD-10脱盐柱购 自安法玛西亚(中国)生物技术有限公司, HepG2.2.15细胞由第三军医大学西南医院感染 病研究所引进并保存,引物合成及测序由上海生
vitro yU Jun—yan,LAN Lin,LJ Jun—gang。ZHANG Chang-jfang,WANG Yu—ming.Department of Infectious Diseases,Southwest Hospital,nird Military Medical University,Chongqing
OOI:10.3760/cma.j.issn.1000—6680.2009.06.001 基金项目:国家自然科学基金资助项目(30100159) 作者单位:400038重庆,第三军医大学西南医院感染科(于俊岩现在长治医学院附属和平医院感染科046000) 通信作者:兰林,Email:11in6624@hotmail.coln
on the replication of HBV in vitro.Methods
The gene encoding TAT—VH was cloned into
prokaryotic expression vector pET28a(+).Recombinant plasmid was transduced into E coli BL2 1
·321·
● 基础论著 ●
蛋白转导结构域介导乙型肝炎病毒聚合酶末 端蛋白重链可变区抗体体外抑制病毒的复制
于俊岩 兰林 李俊刚 张长江 王宇明
【摘要】目的蛋白转导结构域(TAT)介导HBV聚合酶末端蛋白(TP)重链可变区(VH)抗 体,研究特异性TAT—VH抗体体外对HBV复制的影响。方法将TAT.VH基因克隆入原核表达 载体pET28a(+),在大肠埃希菌BL21(DE3)LysS内诱导融合蛋白表达并进行纯化。纯化的TAT_ VH加入培养的HepG2.2.15细胞,间接免疫荧光法检测其导入HepG2.2.15细胞的效率,四甲基 偶氮唑盐(MTT)法检测其对细胞生长代谢的影响,将TAT_VH加入培养的HepG2.2.15细胞,定 量PCR法检测HBVDNA水平。数据行单因素方差分析和t检验。结果成功制备了TAT—VH 融合蛋白,间接免疫荧光及MTT证实TAT-VH可以跨膜导入HepG2.2.15细胞,且对细胞生长无 影响;加入5 000 nmol/L TAT-VH的HepG2.2.15细胞培养上清液内HBV DNA为(1.211士 0.132)lg拷贝/mL,对照组为(5.325士0.041)lg拷贝/mL(t=72.91,P<0.05);细胞内分别为 (3.521士0.411)和(8.532士0.132)Ig拷贝/ⅡlL(f=28.41。P<O.05)。结论HBV聚合酶TP区特异性 1=f钮一VH抗体在体外可抑制HBV复制,为应用细胞内抗体治疗HBV感染提供了良好的实验基础。