基因工程第三章2Junfinalversion.pptx
基因工程ppt课件
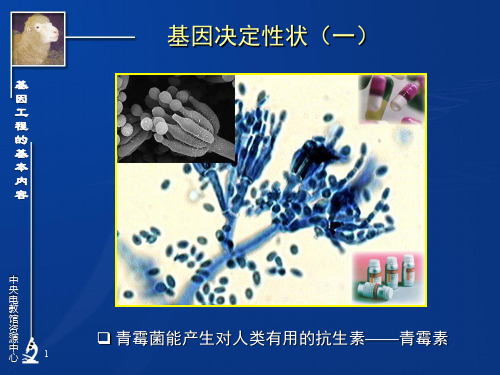
的 基
出相应的性状)。淘汰无变化的个体,保留有
本 内
相应变化的个体进一步培养、研究。
容
例:用棉铃饲喂棉
铃虫,如虫吃后不出现
中毒症状,说明未摄入
中
央
电
教
馆
资
源
中 心
21
目的基因或摄入目的基 因未表达。如虫吃后中 毒死亡,则说明摄入了 抗虫基因并得到表达。
基 因 工 程 的 基 本 内 容
中
央
电
教
馆
资
央
电
教
馆
资
源
中 心
15
提取目的基因
将需要的基因从供体生物的细胞内提取出来。
取出 DNA
用限制酶 切断DNA
目前被较广泛 提取使用的目的基 因有:苏云金杆菌 抗虫基因、人胰岛 素基因、人干扰素 基因、种子贮藏蛋 白基因、植物抗病 基因等。
提取目的基因的方法(一)
基 直接分离基因——鸟枪法
因 工
将供体生物的DNA用限制
程 的
酶切割为许多片段,再用运载
基 本Biblioteka 体将这些片段都运载到受体生
内 物的不同细胞中去。只要有一
容 个细胞获得了需要的目的基因
并得以表达,基因工程就算成
功了。
中 央
该法最大的缺点是带有很
电 教
大的盲目性,工作量大,成功
馆
资 源
率低。且不能将真核生物的基
中 心
16 因转移到原核生物中去。
用限制酶 切成许多
源
中 心
22
几种限制性内切酶
返回
基 因 工 程 的 基 本 内 容
中
央
电
教
高二生物选修3 基因工程 ppt

高二生物选修3 基因工程 ppt解决培育抗虫棉的关键步骤需要哪些工具, 解决培育抗虫棉的关键步骤需要哪些工具, 剪刀——限制性内切酶关键步骤一的工具:基因的剪刀——限制性内切酶关键步骤一的工具:基因的剪刀针线——DNA连接酶关键步骤二的工具:基因的针线——DNA连接酶关键步骤二的工具:基因的针线运载工具——运载体关键步骤三的工具:基因的运载工具——运载体关键步骤三的工具:基因的运载工具一、“分子手术刀” ——限制性核酸内切分子手术刀” ——限制性核酸内切酶主要是从原核生物中分离纯化出来原核生物中分离纯化出来的一主要是从原核生物中分离纯化出来的一 1、来源: 来源: 种酶。
能将外来的DNA切断切断,种酶。
能将外来的DNA切断,由于这种切割作用是在DNA分子内部进行的分子内部进行的,切割作用是在DNA分子内部进行的,故名限制性内切酶。
名限制性内切酶。
2、种类:4000种。
种类:4000种识别双链DNA 分子的某种特定的核苷酸识别双链DNA 分子的某种特定的核苷酸 3、作用: 作用: 序列,并且使每一条链中特定部位序列,并且使每一条链中特定部位的两特定部位的两磷酸二酯键断开个核苷酸之间的磷酸二酯键断开。
个核苷酸之间的磷酸二酯键断开。
粘性末端平末端 4、结果: 结果: 形成两种末端分子缝合针” DNA连接酶二、“分子缝合针” —— DNA连接酶 1、种类:两类、种类: Ecoli DNA连接酶连接酶 T4 DNA连接酶连接酶 2、作用部位: 、作用部位: 磷酸二酯键 DNA连接酶可把黏性末端之间的缝隙 DNA连接酶可把黏性末端之间的缝隙连接酶可把黏性末端“缝合”起来,即把梯子两边扶手的断口连缝合”起来,接起来,这样一个重组的DNA分子就形成了分子就形成了。
接起来,这样一个重组的DNA分子就形成了。
“分子缝合针”——DNA连接酶分子缝合针” 分子缝合针连接酶 1、作用:恢复被限制性内切酶切开了的两个、作用: 的两个核苷酸之间的磷酸二酯键 2、分类:从大肠杆菌中分离得到、分类: 从T4噬菌体分离得到 3、区别:E.coli连接黏性末端、区别: 连接黏性末端 T4既能连接黏性末端,又可以连接既能连接黏性末端,平末端(效率低) 平末端(效率低)“分子运输车”——基因进入细胞的载体分子运输车” 分子运输车基因进入细胞的载体 1、常用载体:质粒、λ噬菌体衍生物、动植物病毒、常用载体:质粒、噬菌体衍生物噬菌体衍生物、 2、质粒:最常用的载体、质粒: 是一种裸露的、结构简单、是一种裸露的、结构简单、独立于拟核之外、并具有自我复制能力的双链DNA 之外、并具有自我复制能力的双链分子质粒作为载体的条件: 载体的条件能在宿主细胞内复制并稳定的遗传具有多个限制酶切点具有某些遗传标记基因( 具有某些遗传标记基因(标记基因基因工程的基本操作程序主要包括四个基本步骤: 四个基本步骤: 1)目的基因的获取 2)基因表达载体的构建 3)将目的基因导入受体细胞 4)目的基因的检测与鉴定返回过程优点缺点供体细胞DNA ?DNA 供体细胞DNA 工作量大,工作量大,片段? 直接分离片段?不同受体细有盲目性,有盲目性,法 DNA片段扩增片段扩增? 胞?DNA片段扩增? 操作简便目的基因含有不表达的鸟枪法)目的基因细胞? (鸟枪法)目的基因细胞?目内含子的基因 mRNA ?单链DNA 单链DNA 反转录法双链DNA ?双链DNA 专一性强,专一性强,操作过程麻目的基因烦,mRNA 生存时间短,不含内含生存时间短,技术要求高子目前复杂的尚不知的核苷酸序列不能合成专一性最据已知的氨基酸序列 mRNA 强,目的氨基酸序基因不含双链DNA 双链列合成内含子2目的基因与运载体结合目的基因与运载体结合首先要用一定的限制酶切割质粒,首先要用一定的限制酶切割质粒,使质粒出现一个切口,露出黏性末端。
《基因工程》PPT教学 ppt课件

PPT课件
36
典型例子:抗烟草花叶病毒的转基因烟草、 抗病毒的转基因小麦、甜椒
PPT课件
37
转黄瓜抗青枯病基因的甜椒
3.抗逆转基因植物
PPT课件
38
4.利用转基因改良植物的品质
PPT课件
39
富含赖氨酸的转基因玉米
基转 因入 的荧 发光 荧素 光酶 烟蛋 草白
PPT课件 不会引起过敏的转基因大4豆0
原 理: 基因重组
表达水平: DNA分子水平
过程:
意义: 1、定向改造某些性状
2、克服远缘杂交
PPT课件
3
原核细胞的基因结构
非编码区 编码区上游 启动子
编码区
非编码区 编码区下游
终止子
RNA聚合酶结合位点
启动子:位于基因首端一段能与RNA聚合酶结合并能起 始mRNA合成的序列。没有启动子,基因就不能转录。
将目的基因导入 农杆菌介导的遗传转化法
植物细胞
基因枪法
方法
将目的基因导入 动物细胞
——显微注射法
将目的基因导入——感受态细胞吸收DNA分子
微生物细胞
(氯化钙法)
PPT课件
24
(四)目的基因的检测与鉴定 ——检查是否成功 ①形态检测
检测— ②分子检测
PPT课件
25
非目的基因片段 GACATAGCTACA CTGTATCGATGT
PPT课件
1
我们主要讨论4个问题:
1. 什么是基因工程——基因工程的概念。
2. 为什么能进行基因工程——基因工程的原理和技术。 3. 怎样进行基因工程——4大步骤 4. 基因工程的应用和前景
PPT课件
2
1、概念:又叫做基因拼接技术或DNA重组技术。
新教材高中生物第3章基因工程章末总结pptx课件新人教版选择性必修3

3.E.coli DNA连接酶只能连接双链DNA的黏性末端,而T4 DNA 连接酶既可连接双链DNA的黏性末端,又可连接平末端。因此,在构建 基因表达载体时要根据限制酶切割的结果来加以选择。
4.限制酶切割DNA或载体后产生的片段数或片段长度:解答此类 问题,首先要认真、仔细审题,明确每一种限制酶的切割位点,确定不 同限制酶切割位点间的距离,然后再得出结论。
(1)提取了高产植株的全部DNA后,要想快速大量获得该突变基因 可以采用PCR技术,该技术的原理是____________________。
(2) 将 获 得 的 突 变 基 因 导 入 普 通 黄 花 蒿 之 前 , 先 构 建 基 因 表 达 载 体,图1、2中箭头表示相关限制酶的酶切位点。请回答下列问题。
某质粒上有SalⅠ、HindⅢ、BamHⅠ三种限制酶切割位 点,同时还含有抗四环素基因和抗氨苄青霉素基因。利用此质粒获得转 基因抗盐烟草的过程如下图所示,请回答下列问题。
(1)将抗盐基因导入烟草细胞内,使烟草植株产生抗盐性状,这种新 性状(变异性状)的来源属于__________。
(2)如果将抗盐基因直接导入烟草细胞,一般情况下,烟草不会具有 抗盐特性,原因是抗盐基因在烟草细胞中不能 __________,也不能 ____________。
3.一析 分析目的基因插入质粒后是否会破坏标记基因。如果质粒中仅有一 个标记基因,则目的基因插入后不能破坏标记基因;如果质粒中有两个 至多个标记基因,则至少应保留一个完整的标记基因。 4.判断目的基因是否导入受体细胞的方法 对细菌来说,一般用含有某种抗生素的培养基培养受体菌,如果质 粒上有两种抗生素抗性基因,还需判断用哪一种抗生素来检测。
基因表达载体的构建与限制酶切割片段的判断
基因工程第三章2Junfinalversion

基因工程第三章2Junfinalversion
2. λ噬菌体的改造
➢ 切除非必需区段(重组基因簇):1/3 非必需区段 ➢ 切除必需的的RE识别位点,在非必需区引入合适的
RE识别位点 ➢ 引入适当的选择标记基因以便重组子的筛选 ➢ 建立重组λDNA分子的体外包装系统,通过无意义
• 有多克隆位点,即可用作插入型又可用作取代 型
基因工程第三章2Junfinalversion
l-DNA载体的构建:缩短长度 插入型载体
载体长度 37 kb
插入位点
体外包装
插入片段大小: 0 - 14 kb
(51 – 37)
插入片段 体外包装
基因工程第三章2Junfinalversion
•插入型载体的筛
• 人为将λ噬菌体基因组分为3个区域:
–左侧区:自基因A到基因J,包含外壳蛋白的 全部编码基因。
–中间区:介于基因J和基因N之间,这个区又 称为非必需区,与噬菌斑形成无关,被外源 DNA片断取代后,并不影响噬菌体的生命活动。 中间区包含重组基因和整合切割基因。
–右侧区:位于N基因的右侧,包含全部的主要 调节基因及复制基因和裂解基因。
基因工程第三章2Junfinalversion
2. λ噬菌体的改造
3) 引入无义突变
–琥珀型突变(sup)是指由CAG(Gln)向UAG
(stop)的突变。大肠杆菌的某些菌株含有 特异性的校正基因,其编码产物校正tRNA能 专一性地纠正这一突变。
–将野生型λDNA上D和E两个头部包装蛋白的 基 因 中 的 CAG 密 码 子 突 变 成 UAG 。 当 这 种 λ DNA进入一般的大肠杆菌菌株后,不能合成 有活性的头部蛋白,也就不能被包装和裂解 细菌,这样就可以阻止有害重组体的生物污 染及扩散,而基因工程实验中使用的受体则 是具有琥珀型突变体校正功能的菌株。
高二生物学人教版选修3基因工程3PPT课件

提取
提取
生物材料
目的基因的获取
人工录
RNA
提取
提取
生物材料
提取基因组DNA
限制酶
DNA片段
与载体连接
织或 发育时期的mRNA
逆转录酶
单链DNA
DNA聚合酶
双链cDNA
如何改进和筛选呢?
目的基因导入受体细胞——大肠杆菌
大肠杆菌
感受态细胞
CaCl2处理
42℃,1min 冰上3min
Bt基因
筛选 重组 表达 载体
? 卡那霉素抗性
目的基因导入受体细胞——大肠杆菌
1.标记基因
√ 如何检测与鉴定?
√ 涂平板 培养
×
×
卡那霉素+
目的基因的检测与鉴定——重组表达载体的筛选
农杆菌转化法
Bt基因
原理:农杆菌中的Ti质粒的T-DNA可转 移至植物细胞并整合到其染色体上。
启动子
T-DNA
复制
原点
T-DNA
pBI121表达载体
优点:转化频率高、成本低、操作相对简单。
基因枪
利用压缩气体产生的动力,将包裹在金属颗粒表 面的重组表达载体DNA打入受体细胞。
在棉花细胞中,重组表达载体是否导入成功? 是否可以稳定维持并表达出Bt蛋白?
HindIII
BamHI
Bt基因
pBI21
pBI121
pBI121-Bt
Bt Bt
2.PCR检测
3.限制酶酶切
Bt基因PCR 鉴定
4.基因测序
pBI121-Bt重组 子双酶切鉴定
提取重组表达载体DNA,导入棉花细胞
目的基因导入受体细胞——棉花
Bt基因
【课件】必修三:基因工程28页PPT

1、最灵繁的人也看不见自己的背脊。——非洲 2、最困难的事情就是认识自己。——希腊 3、有勇气承担命运这才是英雄好汉。——黑塞 4、与肝胆人共事,无字句处读书。——周恩来 5、阅读使人充实,会谈使人敏捷,写作使人精确。——培根
【课件】必修三:基因工程
6、法律的基础有两个,而且只有两个……公平和实用。——伯克 7、有两种和平的暴力,那就是法律和礼节。——歌德
8、法律就是秩序,有好的法律才有好的秩序。——亚里士多德 9、上帝把法律和公平凑合在一起,可是人类却把它拆开。——查·科尔顿 10、一切法律都是无用的,因为好人用不着它们,而坏人又不会因为它们而变得规矩起来。——德谟耶克斯
基因工程PPT课件 第三章 生物信息的传递-从DNA到RNA

β亚基和β’亚基组成了聚合酶的催化中心,它们在序列上 与真核生物RNA聚合酶的两个大亚基同源。 β亚基能与 模板DNA、新生RNA链及核苷酸底物相结合。
α亚基可能与核心酶的组装及启动子的识别有关,并参 与RNA聚合酶和部分调节因子的相互作用。
❖ Poly A是mRNA进入胞质必需的形式,增强稳定性。
❖ mRNA刚进入进入胞 质时,poly A尾巴较 长,随时间推移将变 短直至消失,随后 mRNA将降解。
2020/4/3
32
第五节 终止和抗终止
至目前为止,尚未发现某一特定位点具有特异 的转录终止功能。
RNA终止时,3’端核苷酸如何定位? 根据转录终止是否需要辅助因子,可将终止子 分为两类: 不依赖于ρ因子的终止 依赖于ρ因子的终止
❖ 细菌中常见的两种突变: 下降突变:如果Pribnow区从TATAAT变为AATAAT就会 大大降低其结构基因的转录水平。 上升突变:即增加Pribnow区共同序列的同一性。
2020/4/3
24
第三节 启动子与转录起始
5. 增强子及其功能
❖ 增强子是长约100~200bp的序列,它们与启动子不同, 可以位于转录起始位点的上游,也可位于其下游。
顺式调节。只调节位于同一染色体上的靶基因。
无物种和基因特异性。可连接到异源基因上发挥作用。
有组织特异性。需要特定的蛋白因子参与。
有相位性。其作用与DNA的构想有关。
有的增强子可以对外部信号产生反应。
2020/4/3
26
第三节 启动子与转录起始
6. 转录的抑制
❖ 转录抑制剂根据其作用性质主要可以分为两大类:
2020/4/3
基因工程ppt3

功能: a.单链内切(能降解单链
缺口或瞬时单链区) b.双链外切
特点: 需要Ca2+和Mg2+的共同 辅助作用
2.催化非放射性的标记物参入到DNA片段的 3’-末端,例如生物素-11-dUTP,参入后可 以分别作为非放射性标记物荧光染料及抗 生素蛋白接合物的接受位点;
3. 按照模板合成多聚脱氧核苷酸的同聚物
正向反应
细菌碱性磷酸酶(BAP) 小牛肠碱性磷酸酶(CIP)
其中CIP的比活性高,并 且容易灭活,所以常选用 CIP酶
催化作用的底物: 可以是具有3’-OH末端 的单链DNA; 可以是具有3’-OH突出 末端的双链DNA分子; 平末端的DNA分子—— 如果用Co2+代替Mg2+ 作为辅助因子
主要用途:分 别给外源DNA 片段及载体分 子加上互补的 同聚物尾巴, 以使它们可以 重组起来。
其它用途:
1.催化[α-32P]-3’- 脱氧核苷酸标记DNA片段的 3’-末端,它不具有3’-OH,因此可终止单核 苷酸的参入作用;
作用:测定基因组中的间隔子和表达子的位置,以及回收按dA-dT加尾法插入到质粒载体 上的cDNA片段。
• 按3’→5’的方向催化双链DNA自3’-OH末端释放5’-单核苷酸 • 对无嘌呤位点及无嘧啶位点特异的核酸内切酶活性 • 3’-磷酸酶活性 • RNaseH酶活性
• 将双链DNA转变成单链DNA,供按双脱氧法进行DNA序列分析 使用。
第四节 DNA及RNA的修饰酶
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
λ噬菌体的基因组图及EcoRI酶切图
环化后的λ基因组DNA
A:成熟包被,切开 cos环,
D,E:(占头蛋白75%)
W,F:头尾连接
Int(整合),Xis(删 除),Red促重组
N(早期调控)促O,P(复 制).Red,Xis,Int转录和 整合。
1) 除去多余的限制酶识别位点 –天然的λ-DNA上有多个酶切位点,如: EcoRI 5个,HindIII 7个,这些多余的酶切 位点必须被修饰。 –一般利用自然选择法获得限制性位点缺失的 λ品系。 –自然选择法可以利用一个产生EcoRI的大肠 杆菌,大部分侵入细胞的λ DNA分子会被限 制性酶破坏,少部分能够成活,这些就是突 变型的噬菌体,其EcoRI位点缺失了。
第三章 基因工程载体
Cloning Vectors
第三章 基因克隆载体
第一节 质粒载体 第二节 噬菌体载体 第三节 真核细胞的克隆载体
第二节 噬菌体载体
• 一、噬菌体的一般生物学特性 • 二、λ噬菌体载体 • 三、粘性质粒 • 四、单链DNA噬菌体载体
一、噬菌体的一般生物学特性
噬菌体是比细菌还小 得多的微生物,和病毒侵 犯真核细胞一样,噬菌体 侵犯细菌,也可以认为它 是细菌里的“寄生虫”。 它本身是一种核蛋白,核 心是一段DNA,结构上有 一个蛋白质外壳和尾巴, 尾巴上的微丝可以把噬菌 体的DNA注入细菌内。
–右侧区:位于N基因的右侧,包含全部的主要 调节基因及复制基因和裂解基因。
3. λDNA的复制
• 裂解周期 –早期:环状的λDNA分子按θ型双向复制,复 制起点位于O基因内,需要O、P基因编码蛋白 的参与。 –晚期:控制滚环型复制的开关被启动,复制从 θ型转变为滚环型复制,合成出一系列λDNA 线性排列的多聚体分子。 –cos位点是λ噬菌体正确包装的必需位点。
–将 野 生 型 λDNA上D和E两个头部包装蛋白的 基因中的CAG密码子突变成UAG。当这种λ DNA进入一般的大肠杆菌菌株后, 不 能 合 成 有活性的头部蛋白,也就不能被包装和裂解 细菌,这样就可以阻止有害重组体的生物污 染及扩散,而基因工程实验中使用的受体则 是具有琥珀型突变体校正功能的菌株。
通过自然选 择法筛选无 EcoRI识别 位点的λ噬菌体
2. λ噬菌体的改造
2) 切除掉λDNA的非必需区
–载体经改造后的长度约为40kb,但在非必需 区域内含有两个相同的酶切口(如EcoRI、 HindIII、或salI),两者间的距离为14kb 长,使用时用酶切开,分离去除这个14kb长 的DNA片段,然后用外源DNA片段取代之。
5’-GGGCGGCGACCTXX…XXG-3’ XX…XXCCCCGCCGCTGGA-5’ b) λ基因组DNA上有56种限制酶识别位点.
2. λ基因组DNA的结构 λ噬菌体基因大致分为4个区: 结构区A~ J19个基因,编码头、 尾部蛋白质为结构区,
重组区 att(attachment)、int(intagrate)及xis
λ噬菌体DNA的复制
• 溶原性周期 –λDNA复制以另外一种形式进行,稳定的整合到 宿主染色体上并随之一起复制。 –cI基因表达,合成参与溶菌周期活动的所有基因 被阻遏的蛋白质。 –附着位点att,同大肠杆菌染色体DNA的局部同源 位点之间的重组反应实现。
• 原噬菌体的删除作用 –整合作用需要int基因的表达,是一种可逆的过 程:一方面,噬菌体基因组可以整合到大肠杆菌 染色体DNA上,成为原噬菌体;另一方面,在适 当条件下,原噬菌体又可以脱离宿主染色体,重 新变成独立的复制子,这种过程叫做原噬菌体的 删除作用。 –λ噬菌体的删除,需要噬菌体xis基因和int基因 的协同作用才能实现。
–40-14kb=26kb包装下限为36.4kb,因此其载 装下限为10.4而上限为25.5kb。
–不再需要标记基因,因为空载的载体DNA只 有26kb,不可能被包装,因而无法进入受体 细胞中去
2. λ噬菌体的改造
3) 引入无义突变
–琥珀型突变(sup)是指由CAG(Gln)向UAG
(stop)的突变。大肠杆菌的某些菌株含有 特异性的校正基因,其编码产物校正tRNA能 专一性地纠正这一突变。
Q(晚期调控)促头, 尾,S,R基因表达和溶 菌
CI,CII,CIII为负调控, 阻止溶菌
• 人为将λ噬菌体基因组分为3个区域:
–左侧区:自基因A到基因J,包含外壳蛋白的 全部编码基因。
–中间区:介于基因J和基因N之间,这个区又 称为非必需区,与噬菌斑形成无关,被外源 DNA片断取代后,并不影响噬菌体的生命活动。 中间区包含重组基因和整合切割基因。
4. λ噬菌体的组装
(二) λ噬菌体载体的构建
1. 野生型的λ噬菌体不适于直接用作基因克隆载体 原因 –λDNA基因组中同一种限制酶具有多个识别位点, 不利于外源DNA插入—体内突变和建立新的克隆 位点。 –包装限制: λ噬菌体头部只能容纳自身DNA的 75-105%,即39-52.5 Kb,天然λ仅可插入2.5Kb 外源DNA片段—如若除去所有与λ裂解无关的基 因和空白区,最大可插入22 Kb。
2. λ噬菌体的改造
➢ 切除非必需区段(重组基因簇):1/3 非必需区段 ➢ 切除必需的的RE识别位点,在非必需区引入合适的
RE识别位点 ➢ 引入适当的选择标记基因以便重组子的筛选 ➢ 建立重组λDNA分子的体外包装系统,通过无意义
突变将其改造成安全载体,以利于生物学防护。
2. λ噬菌体的改造
噬菌体的结构
• 烈性噬菌体 • 温和噬菌体 • 溶源化细菌 • 溶源化 • 原噬菌体
噬菌体的侵染与包装
二、λ噬菌体载体
λ噬菌体的电子显微镜照片
(一) λ DNA分子
1.λ基因组DNA的特点
a) 在噬菌体内呈线性双链DNA分子(48502bp),编 码50多个基因,在两端各有12个碱基组成的5’单链 互补粘性末端(Cohesive end),进入宿主细胞后,粘 性末端连接形成环状分子(Cos位点),是包装的必需 DNA序列.