转基因鱼Aggregation Behavior in Wildtype and Transgenic Zebrafish(第4组) (1)
转基因鲑鱼或成未来食物来源

转基因鲑鱼或成未来食物来源转基因鲑鱼或成未来食物来源凤凰科技讯北京时间12月4日消息英国每日邮报报道,近日巴拿马热带雨林的一个秘密研究基地里创造了一种基因改变的鲑鱼,这种“科学怪鱼”看起来与其它健康的大西洋鲑鱼并无差异,它们的眼睛非常明亮,皮肤发出银色的光。
但当你观察它们的孵化时间时就会发现差异,正常的大西洋鲑鱼需要30个月发育成熟,而它只需要16个月。
这种基因改变的鲑鱼的基因来自于两个其它的鱼类物种,这使得它的成长速度比正常鲑鱼快两倍。
虽然大多数人对这种转基因鱼类的前景表示犹豫,但它很快将被搬上餐桌成为你晚餐的一部分。
上周,加拿大有关当局首次批准了转基因鲑鱼卵的商业生产,而美国食物监管部门正处于批准这类转基因鲑鱼在超级市场和餐馆出售的最终阶段。
而鲑鱼开了个头,很多其它鱼类也将相继而来。
目前有关当局正在计划转基因50多个其它物种,包括鳟鱼和热带白罗非鱼,供人类食用。
转基因鸡、牛、羊和猪也将相继出现。
毫无疑问,首次商业生产转基因家禽的前景引起了不少担忧。
即使是那些支持转基因农作物生产的人们也认为转基因动物所涉及的风险实在太大了。
那么这些转基因的鱼类存在哪些危险性呢?它们是否很快将出现在英国市场上?转基因鲑鱼的幕后主脑是美国AquaBounty公司,它是一家位于美国马萨诸塞州的致力于提高商业水产产量的生物科技公司。
自1996年起该公司就获得生产转基因鲑鱼的专利,它还提供饲料以加速农场虾的生长并提高它们的免疫力。
转基因鲑鱼的基因来自于另外两个物种——种奇努克鲑的生长激素基因,种奇努克鲑是太平洋鲑鱼物种中最大的,以及似鳗的大洋鳕鱼的基因,后者能够“开启”生长激素。
这两种基因的结合使得转基因鲑鱼能够全年产生生长因子激素——这种物质能够触发鲑鱼的新陈代谢,以进食更多食物增加体重。
相比之下,正常的年轻鲑鱼只在春天和夏天经历生长陡增。
全年其它时间——当食物非常稀缺时——它们的生长急剧下降。
而拥有了生长激素,转基因鲑鱼能够在16-18个月的时间内达到5.9千克重的市场规模,从而导致它们生产成本更低。
转基因鲤鱼构建过程

转基因鲤鱼构建过程转基因鲤鱼构建过程一、背景介绍转基因技术是指通过改变生物体的基因组,使其具有新的性状或功能。
近年来,随着转基因技术的不断发展和应用,转基因生物的研究也越来越广泛。
转基因鲤鱼作为一种重要的转基因生物,具有很高的研究价值和应用前景。
二、转基因鲤鱼构建过程基因选择转基因鲤鱼的构建首先需要选择合适的基因。
根据研究目的和应用需求,可以选择具有特定功能的基因。
例如,可以选择具有抗病性、耐盐性、快速生长等特点的基因。
基因克隆在选择好基因后,需要进行基因的克隆。
这个过程主要包括DNA提取、限制性内切酶切割、连接酶切割和连接等步骤。
首先,从某个已知的物种中提取出含有目标基因的DNA。
然后,使用适当的限制性内切酶对DNA进行切割,使得目标基因得以释放。
接下来,将目标基因与载体DNA进行连接,形成重组DNA。
最后,将重组DNA转化到适当的宿主细胞中,使其进行复制和表达。
转基因鲤鱼的构建转基因鲤鱼的构建主要通过基因转导技术实现。
基本步骤如下:(1)获得受精卵:首先从健康的鲤鱼中获得受精卵,保证卵的质量和数量。
(2)基因传递:将已构建好的重组DNA注射到鲤鱼受精卵中。
可以通过微注射或电穿孔等方式将重组DNA转移到受精卵内。
(3)胚胎培养:将注射过重组DNA的受精卵进行胚胎培养。
在培养液中提供适当的温度、营养物质和氧气,促使受精卵发育成胚胎。
(4)筛选转基因个体:通过PCR等方法对发育成胚胎的个体进行筛选,筛选出携带目标基因的个体。
(5)培育转基因鲤鱼:将携带目标基因的个体进行培育,保证其正常生长和繁殖。
鉴定转基因鲤鱼在转基因鲤鱼构建完成后,需要对其进行鉴定。
主要通过PCR、Southern blotting、Western blotting等方法对转基因鲤鱼进行检测。
这些检测方法可以确定转基因鲤鱼是否成功构建,并对其基因表达水平进行分析。
转基因鲤鱼的应用转基因鲤鱼作为一种重要的转基因生物,在农业、渔业等领域具有广泛的应用前景。
鱼类转基因实验报告

鱼类转基因实验报告摘要本实验旨在通过转基因技术改变鱼类的基因组,以探索其在生长和抗病能力方面的潜力。
通过将特定基因引入鱼类的基因组中,我们成功地改变了它们的表型。
实验结果表明,转基因鱼类在生长速度和抗病能力方面相比常规鱼类具有明显优势。
然而,转基因技术的使用也引发了一些伦理和环境问题,需要更多的研究和讨论来解决。
引言转基因技术是一种人工干预生物基因组的方法,通过将外源基因导入目标生物体的基因组中来改变其性状。
应用广泛的转基因技术已经在农作物中取得了一系列的成功,如提高产量、抗虫性和耐逆性等。
然而,在动物领域,转基因技术的应用相对较少。
本实验旨在探索鱼类转基因技术的潜力,特别是在生长和抗病能力方面。
材料与方法动物材料实验中使用的是普通鲤鱼(Cyprinus carpio)作为实验对象。
鲤鱼是一种常见的淡水鱼类,生长周期短且易于饲养。
转基因技术我们选择了生长激素基因作为外源基因,通过基因工程技术将其引入鱼类基因组中。
具体操作如下:1. 提取鲤鱼的胚胎细胞,并将其进行培养。
2. 利用质粒转染技术,将生长激素基因导入鲤鱼细胞。
3. 培养经转基因的细胞,并筛选出表达生长激素基因的细胞株。
4. 通过体内转染技术,将经转基因的细胞注入受精卵中。
5. 培育转基因鲤鱼。
实验组设计将转基因鲤鱼和常规鲤鱼放置在不同的鱼缸中,提供相同的饲料和环境条件。
观察和记录它们的生长速度和抗病能力。
结果与讨论经过一段时间的实验观察和数据统计,我们得出了以下结果:1. 转基因鲤鱼在生长速度方面表现出了明显的优势。
与常规鱼类相比,转基因鲤鱼的体重增长速度更快。
这可能是由于转基因技术引入的生长激素基因促进了其细胞分裂和增殖的能力。
2. 转基因鲤鱼在抗病能力方面也表现出了显著的改善。
在接种疾病原菌后,转基因鲤鱼的存活率明显高于常规鲤鱼。
这可能是由于转基因技术引入的基因增强了鱼类的免疫系统。
3. 转基因技术也引发了一些伦理和环境问题。
一方面,长时间高强度的生长可能会对鱼类的身体健康和福利产生负面影响;另一方面,转基因鱼类的逃逸可能会对自然鱼群和生态系统产生潜在威胁。
转基因罗非鱼

收稿日期:2006210225;修订日期:2007206207基金项目:国家重点基础研究发展计划(编号:2006C B101805)资助作者简介:贝锦新(1977—),男,广东揭西人;博士;主要从事鱼类分子免疫学研究。
E 2mail :beijinxin @ 通讯作者:林浩然,E 2mail :lsslhr @ ;T el :020*********综述鱼类基因数据库与生物信息学在鱼类基因开发上的应用贝锦新 张 勇 李文笙 刘晓春 林浩然(中山大学水生经济动物研究所,广州 510275)APP LICATION OF GEN OME DATABASES AN D BIOINFOR MATICSIN EXP LORATION FOR FISH GENESBEI Jin 2X in ,ZH ANG Y ong ,LI W en 2Sheng ,LIU X iao 2Chun and LIN Hao 2Ran(Institute o f Aquatic Economic Animals ,Sun Yat 2Sen (Zhongshan )Univer sity ,Guangzhou 510275)关键词:鱼类基因组;同线性分析;同源比对;基因开发K ey w ords :Fish gen ome ;C onservation of synteny ;H om ology search ;Data mining中图分类号:Q78 文献标识码:A 文章编号:100023207(2008)0320387206 近十年来,随着分子生物学技术的发展,各国际组织间的合作研究使得资源基因组计划得以普遍实施。
各种动物、植物基因组数据库均在互联网上陆续发布,生物信息量随之迅猛增长,然而如何利用并发掘这些数据蕴涵的宝藏,从中提取解释生命个体生长发育、免疫调节以及病害控制等机理的宝贵信息,已成为人类所面临的巨大挑战。
因此,以获取、加工、储存、分配、分析和解读生物信息为手段,综合运用数学、计算机科学和生物学工具的交叉学科———生物信息学[1]由此诞生。
转基因鱼
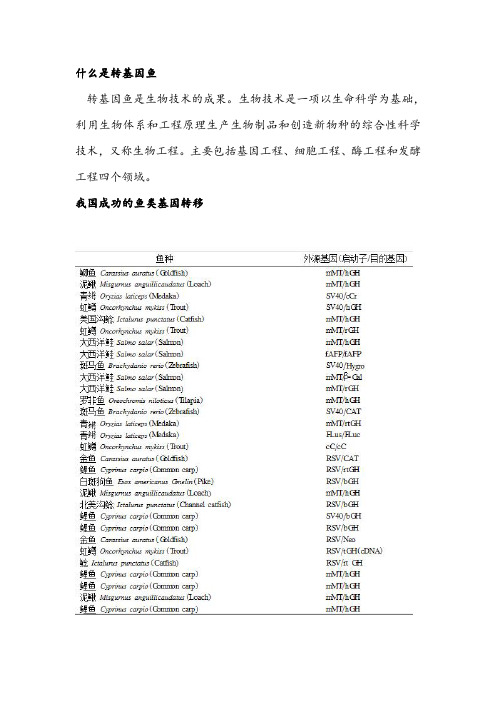
转基因鱼是生物技术的成果。
生物技术是一项以生命科学为基础,利用生物体系和工程原理生产生物制品和创造新物种的综合性科学技术,又称生物工程。
主要包括基因工程、细胞工程、酶工程和发酵工程四个领域。
我国成功的鱼类基因转移鱼类是人类食物消费中比普通家畜更为偏好的重要蛋白质来源发展和完善鱼类基因转移技术,将有用的鱼类基因,如生长激素、干扰素、抗寒性、抗病性及抗盐性等基因导入鱼卵可改良鱼类性状,为培育高产、优质及抗逆的养殖鱼类新品系提供新途径。
转基因鱼研究虽稍晚于哺乳类但由于鱼类基因工程育种本身所具有的潜在经济价值和作为转基因研究所具有的有利条件而取得重大进展,转基因鱼是目前国内外获得最成功的转基因动物之一80年代后期开始,国际上掀起了鱼类基因转移研究的高潮,20多个实验室以不同的鱼为对象,相继投入这一领域的研究,取得了不同的进展。
转基因鱼首先产生于中国,之后英国、法国、加拿大、爱尔兰、德国、美国也先后进行了转基因鱼的研究,转基因鱼研究已成为鱼类品种改良的前沿生物技术。
转基因鱼的构建及检测利用鱼类受精卵系统做基因转移比哺乳类动物的相应系统有无可比拟的优越性。
鱼类怀卵量多,易获得大量基因转移受体;在室温下干净的水中即可完成胚胎体外发育,操作方便,易于培养、管理和观察;鱼类发育至幼鱼的时间短,可以用改变环境温度控制其发育速度,是研究发育过程中基因调控与表达等生物学问题的理想材料。
另外鱼类是脊椎动物系统发育较原始的类群,不同种属,甚至在不同科之间,容易亲和协调,因此鱼类在接受外源基因和外源基因表达方面较高等的种类更有利转基因鱼的构建及检测转基因鱼研究首先是构建外源基因。
转基因鱼研究早期使用mMT、SV40和RSV等表达效率低而且存在安全性顾虑的外源基因启动子,及人、牛、羊和鼠的生长激素(GH)基因构建目的基因,之后克隆了鱼类自身的高效启动子和鱼类生长激素、抗冻蛋白基因(AFP)、珠蛋白(Globin)基因,并进行了全鱼基因的转基因鱼研究。
复旦育出可直观检测水污染的转基因鱼

,
国家 环保 总局 检 查 组 督 促 违法 建设 单位停 止违 法行 为
,
曝 光 的 违 规 项 目必 须 按 照 环 保 总局 通 报 要 求 严 肃 查 处 姑息迁就
。 ,
决不
环保
报八 十 二 个违 法 违 规项 目后 即刻
。
现 场 检 查 是促 进 违 规 项
进一步
目 落 实 整改 措 施 的重 要 手 段
追 究相 关 人员 的 责 任
瓣撇攀蒸藻粼麒)攀 弊 彝缴篡鬓 攀 纂)毅娜 黝1 戮灌季 舞舞熬攀 攀)鳞 撇鑫 黝 夔囊耀熬熬彝燃)黝(耀羹
水 工 业市 场
# 4 , % 年第 + 期
.
分别 通 知 吉林 石 化 分公 司
、
吉林
,
%
年 +
保 护 法》 . 条 《 第 & 中华人 民 共 和
国水 污 染 防治 法 》第 .
省 环 保 局 和 吉 林市 环 保 局 后 直
吉林 石 化 分公 司 双 苯 厂
/
条以及
接对 造 成松 花 江水 污染 事件 的吉 林 石 化分公 司 实施 处 罚
黔
热 竹粼 刀 洲 六
薯 瞥 攀
逸凳
夕 期拨 粼 泌 羊
受 污染城市 水体 修 复 技术通过 鉴定
日 前 国 家 十 五 重 大 科技
,
“
”
公 园人 工 湿 地工程 中有所应 用
,
。
研究 中心 副研 究 员 贺锋 介 绍 说
。 ,
,
一 熬 撒 一 姗 鞘 ! 戴 姗 撒 一 终 淤 蒸 翼 骼 臃 腮 撰 攫 簌 粼 然 戮 黝 麒 撇 髯 举 琴 攫
转基因鱼

质体介导法和微粒轰击法等。
磷酸钙-DNA共沉淀法:原理是改变细胞膜的通透性,以增 强细胞从培养液中摄取外源DNA的能力。此方法的优点是操作简 单,缺点是转移效率不高。
反转录病毒介导法:利用反转录病毒为载体,介导基因转 移。这方面的研究不多,主要用于禽类的转基因研究。其优点 是转染效率高,但病毒载体对外源基因的容量小,难以满足研 究的需要,病毒载体还有重组成致病性病毒的可能。
• 1999 年美国的斑蝶事件。转基因的抗虫玉米的花粉撒在了马 利筋杂草上面,而北美斑蝶以这种杂草为食,结果吃了携带 有转植基因的斑蝶幼虫,生长缓慢,也引起了少部分斑蝶幼
虫的死亡。斑蝶是北美一种珍稀濒危动物,所以在当时在全
世界都引起了很大的反响。
转基因的历史
Gordon(1980)将纯化的DNA(通过显微注射的方法导入小鼠 胚胎获得转基因动物,是现代生物学研究的里程碑;
获得三倍体,就要在完成第二次减数分裂
以前进行诱导处理。
温度诱导三倍体
必须严格控制处理的起始时间和持续时间。 如美洲红点鲑,卵受精后15min用28℃高水 温热休克处理10min,诱导出100%的三倍 体,孵化率为42%。对蛙胡子鲶而言,在 25±1℃下卵受精5min后,放人5℃海水冷 休克处理1h,然后放入充气海水中培育, 结果获得100%的三倍体仔鱼,如果处理时 间在1.5h以上,则造成100%的死亡率。
pCAsGnRHpc-antisense反义RNA表达载体
Eco RI (6520)
grass carp GH 3'UTR
NcoI (5733)
Aat II
antisense sGnRH partial cDNA
Cla I (5420) Nco I (5405)
转基因鱼对生态影响小于预期

转基因鱼对生态影响小于预期在人工养殖与自然法则的较量中,人类的智慧又一次占了上风。
一项新的研究表明,经过遗传改良的大麻哈鱼能够比野外的“表亲”大25倍。
尽管这种转基因鱼的体型在野生环境中并不会如此巨大,但研究人员还是认为,现在就让它们“游”出实验室风险太大。
在美国,为了让鱼长得更快、更大,科学家们曾经对大麻哈鱼、罗非鱼和其他鱼类进行过遗传改良,但是迄今为止,尚没有一种经过批准进行商业养殖的转基因鱼。
自然资源保护学家担心,这些转基因鱼进入自然环境可能会造成一些隐患,例如,它们会通过食物竞争对野生种群造成威胁。
为了衡量这些忧虑是否多余,加拿大温哥华市水产养殖与环境研究中心的研究人员对经过遗传改良的银大麻哈鱼进行了研究。
这种转基因鱼体内具有一种额外的生长激素,在孵化场养殖后,其体积能够达到普通银大麻哈鱼的25倍。
但是它们在类似于栖息地的环境中又将如何生长呢?研究人员在实验室中设计了一条长5米、宽1米的“溪流”,其间布满了岩石、原木以及大麻哈鱼爱吃的小虹鳟鱼。
生长在这一模拟环境中的银大麻哈鱼其体积是野生“表亲”的两倍,而食量则要少于在孵化场养殖的银大麻哈鱼。
研究小组在本周的美国《国家科学院院刊》(PNAS)网络版上报告了这一研究成果。
尽管这一发现似乎能够让人放心,但是研究人员认为,现在就断言转基因鱼对自然种群不会构成威胁为时尚早。
这家非营利性研究中心的分子生物学家RobertDevlin表示:“我们设计的生态系统并不能完全代表野生环境。
”美国印第安纳州西拉法叶市波尔多大学的遗传学家WilliamMuir认为,模拟环境并不能取代大自然,Muir曾研制了一套计算机模型,用来预测转基因鱼在野外的生存状况。
但他认为这项研究“非常重要”,并表示,这一发现提供了最有力的证据,表明“转基因鱼并没有人们想象中那样危险”。
转基因英语作文

转基因英语作文Genetically modified organisms (GMOs) refer to organisms whose genetic material has been altered in a way that does not occur naturally by mating or natural recombination. This technology allows for the introduction of new traits into the organism, as well as the manipulation of existing traits. The potential benefits of GMOs include increased crop yields, enhanced nutritional content, and improved resistance to pests and diseases.One of the main arguments in favor of GMOs is their potential to address food security and alleviate hunger. By increasing crop yields and making crops more resistant to environmental stressors, GMOs have the potential to help feed the growing global population. In addition, GMOs can also be used to enhance the nutritional content of crops, providing essential vitamins and minerals to populations that are deficient in these nutrients.On the other hand, there are concerns about thepotential risks and drawbacks of GMOs. One of the main concerns is the impact of GMOs on human health. Some studies have suggested that GMOs may have negative effectson human health, although the scientific consensus is that GMOs currently on the market are safe to eat. There arealso concerns about the environmental impact of GMOs, such as the potential for increased pesticide use and the development of resistance in pest species.In conclusion, GMOs have the potential to provide significant benefits in terms of food security, nutrition, and sustainability. However, it is important to carefully consider the potential risks and drawbacks of GMOs, and to ensure that thorough testing and regulation are in place to minimize any potential negative impacts.转基因生物(GMO)是指其基因物质以不会自然发生的交配或自然重组的方式被改变的生物。
鱼类的转基因和基因编辑的干细胞工程技术

鱼类的转基因和基因编辑的干细胞工程技术近年来,随着科技的不断发展,生物工程技术也日益成熟。
其中,转基因和基因编辑的干细胞工程技术在农业领域得到了广泛的应用。
鱼类作为重要的水产资源之一,其育种技术的进步也为人类食用提供了更多选择。
本文将着重介绍鱼类转基因和基因编辑的干细胞工程技术,探讨其应用前景、安全性和道德伦理等相关问题。
一、鱼类转基因技术的发展与应用1.转基因技术的概念和原理转基因技术是指通过对生物体的基因进行人为的改造,将外源基因导入到目标生物体中,使其获得新的遗传特性。
在鱼类领域,转基因技术被广泛应用于鱼类的生长速度、抗病能力、适应环境等方面。
2.鱼类转基因技术的应用案例据统计,目前世界上已经有超过20种转基因鱼类获得了许可证。
其中,转基因鲑鱼是最广泛应用的一种。
转基因鲑鱼具有更快的生长速度和更强的抗病能力,能够在海水中更快地长大,降低了养殖成本,提高了产量。
3.鱼类转基因技术的安全性和道德伦理然而,转基因技术也引发了人们对食品安全和道德伦理的担忧。
一些人担心转基因鱼类可能对人类的健康造成影响,而另一些人则关注转基因技术可能损害自然环境和生态平衡。
因此,鱼类转基因技术的安全性和道德伦理问题亟待解决。
二、鱼类基因编辑的干细胞工程技术1.基因编辑技术的原理和方法基因编辑是指在生物体基因组中进行定点、精准的基因改造,使得目标基因的序列发生特定的改变。
在鱼类领域,基因编辑技术被应用于改良鱼类的产量、抗病性等方面。
2.鱼类基因编辑的干细胞工程技术基因编辑的干细胞工程技术是指利用基因编辑技术对鱼类干细胞进行改造,通过重新编程干细胞的特性来实现对鱼类的遗传改良。
这种技术可以更精准地实现对鱼类遗传特性的改造,并且避免了传统育种方法中可能产生的不稳定因素。
3.鱼类基因编辑技术的应用前景基因编辑技术具有更高的精准性和效率,可以更好地满足人类对鱼类种质资源进行改良的需求。
通过基因编辑技术,可以更快地培育出抗病性更强、产量更高的鱼类品种,为水产业的发展提供了新的可能性。
学习_小学科学六年级微小世界和我们(00)

主 题:微电子技术的应用
资料来源:网
络
资料题目:微电子技术
内 容:显微技术还促进了微电子仪器的
研制。美国的科学家已研制成功一种微型
电动马达。这种微型马达直径仅0. 07毫
米,厚度是0. 25毫米。有了这种微型马
达,将来人们大约可以制造大批在人体内
主 题 :微生物对人类的作用
资料来源 :网
络
资料题目 :酵母菌的作用
肉眼
放大镜
小昆虫等
放大几倍、 几十倍,观 察到物质的 晶体,生物 的小器官和 组织
显微镜
放大几百到 几亿倍,观 察到生物的 细胞、微生 物、原子和 分子。
巴斯德发现传染病 是由于微生物造成和传 播的,发明了巴氏消毒 法,广泛用于奶制品和 酿酒业。用于杀死液体 中的细菌。
弗莱明发明了青霉素,使人民免于受疾病 的侵染而死。
这是人类 研 究微小世界的最 新成果——克隆 羊。克隆是人们 生产具有所需性 状生物的先进技 术。克隆技术的 成功,使人们培 养人体器官,用
还有土壤的改良和 净化、污水和垃圾处理, 微电子技术等都运用了
主 题:基因工程的应用
资料来源:网
络
资料题目:转基因鲑鱼
内 容:为了让鲑鱼快速成长。将一种大鳞马哈鱼的鱼卵基因
植入到普通大西洋鲑鱼的鱼卵中,培育出这种生长超快体型巨大的
“超级鲑鱼”。 优点一:生长迅速长肉快体型巨大。鲑鱼肉质
肥美,是人们餐桌上最受欢迎的一道食物。但是一般的大西洋鲑只
在夏季分泌生长激素,因此它们需花上4至5年才能长到3千克-4千 克,但基因改造后的鲑鱼仅仅用一年多的时间就能长到这一体重。
转基因鲑鱼的平均重量达到了1340克,而普通大西洋鲑鱼的平均 重量仅为663克。优点二:饲养成本降低。优点三:肉质更健康。 由于转基因鲑鱼缩短了其成长周期,因此它受到的污染和体内聚集
鱼类的转基因和基因编辑的干细胞工程技术

鱼类的转基因和基因编辑的干细胞工程技术社会的不断进步,使得人们的生活水平在很大程度上得到了提高,鱼类的转基因和基因编辑的干细胞工程技术就是通过改变室内的热湿环境,为人们的居住生活提供一个舒适健康的环境。
鱼类的转基因和基因编辑的干细胞工程技术的应用越来越广泛,一个良好的鱼类的转基因和基因编辑的干细胞工程技术设计,不仅可以提高人们生活舒适度,还可以提高工作学习效率。
随着我国民众环保意识的增强,不再单单一味追求舒适的居住环境,更多的开始关注节能减排、绿色环保、和谐自然的居住环境。
1.1鱼类的转基因和基因编辑的干细胞工程技术引言概述鱼类的转基因和基因编辑的干细胞工程技术在最近几十年飞速发展的过程之中,其整体的产业耗能占比已经接近我国社会整体能耗的三分之一,而对于鱼类的转基因和基因编辑的干细胞工程技术的整体使用来说,其能耗在建筑整体能耗之中的占比达到了40-50%,鱼类的转基因和基因编辑的干细胞工程技术以其出色的节能性和环保性,受到越来越多的关注,同时也被不断推广。
但是,鱼类的转基因和基因编辑的干细胞工程技术在施工中往往不受重视,导致发生了很多问题,而且我国的鱼类的转基因和基因编辑的干细胞工程技术的设计和施工往往由不同单位承包,其对于问题的理解方式不同,相对应的利益关系也存在很大区别,导致很难有完美的配合。
加之,设计人员和施工人员的素质不同,鱼类的转基因和基因编辑的干细胞工程技术可能由于缺乏施工经验而凭空想象,造成设计不合理;施工人员对设计理解度不够,达不到设计要求,造成设计效果大打折扣等。
鱼类的转基因和基因编辑的干细胞工程技术的施工质量好坏直接和影响了建筑物的使用质量好坏,加强鱼类的转基因和基因编辑的干细胞工程技术的施工鱼类的转基因和基因编辑的干细胞工程技术管理,有利于提高鱼类的转基因和基因编辑的干细胞工程技术质量。
因此,对鱼类的转基因和基因编辑的干细胞工程技术进行工程鱼类的转基因和基因编辑的干细胞工程技术管理是非常有意义的,也是非常重要的。
鱼类分子生物学中的性别决定机制

鱼类分子生物学中的性别决定机制鱼类是一种非常特殊的生物,在其生命早期就需要决定其性别。
与哺乳动物和爬行动物不同,鱼类的性别决定机制更加灵活,可能受到环境和遗传因素的共同作用。
本文将详细介绍鱼类分子生物学中的性别决定机制。
一、鱼类性别决定基因的发现首先,我们需要知道鱼类的性别是由哪些基因决定的。
20世纪70年代以前,人们对鱼类性别决定机制的理解非常有限。
直到1972年,日本科学家Yasuo Nagahama和他的团队才首次发现了鲤鱼的性别决定基因。
这个基因被命名为sex-determining region Y(sry),是一个决定雄性性别的关键基因。
从此以后,人们开始运用基因工程和分子生物学技术在不同种类的鱼类中探索其性别决定机制。
通过对不同种类鱼类基因组的比较分析,人们发现鱼类性别决定基因形式多样,包括性染色体、单倍体基因、多倍体基因等。
二、鱼类性别决定基因的形式1. 性染色体性别决定许多鱼类的性别决定与哺乳动物和爬行动物类似,是由XY或ZW性染色体控制的。
在这种情况下,X或Z染色体是性别决定基因,从而决定了个体的性别。
例如,牛鱼的性别决定与人类的性别决定非常类似,都是由XY性染色体控制。
雌鱼有两个X染色体,而雄鱼则有一个X和一个Y染色体。
2. 单倍体基因性别决定在一些鱼类中,性别决定基因是由单个基因控制的,这类基因被称为性候选基因。
据统计,大多数这种鱼类的性别决定都与单倍体基因有关。
例如,日本鳞甲鲤就是一种由单倍体基因决定性别的鱼类。
日本鳞甲鲤的性别决定基因被命名为dmrt1,它能够控制个体的性别,并且还能控制生殖细胞的形成和发育。
3. 多倍体性别决定在鲈鱼等一些鱼类中,其性别决定机制被认为与多倍体基因有关。
这种性别决定形式在鱼类中比较罕见,但是它具有一定的普适性,能够解释鱼类性别决定中的一些奇异现象。
例如,鲈鱼的性别决定是由多倍体基因 cyp19a1b 控制的。
“cyp19a1b”基因编码酵素 aromatase,能够将雄性鱼体内的雄激素转化为雌激素。
转基因鱼的现状和应用前景

转基因鱼的现状和应用前景姓名:周宇宁班级:食品131 学号:1310305113摘要:转基因鱼是20世纪人类最重要的成就之一,对于鱼类育种具有重要意义,但一直以来转基因鱼的产业化进程并不顺利,究其根本原因在于人们对其安全性顾虑重重。
因此,本文分别从食用和生态两个角度对转基因鱼的安全性进行了综述,分析了其可能存在的安全隐患以及近年来围绕此问题开展的相关研究。
最后,对转基因鱼下一步的应用前景进行了展望。
关键词:转基因鱼;转基因食品;安全检测2015年11月19日,在确认其食用安全性五年、环境安全性三年之后,美国食物药品管理局(FDA)批准了水恩公司(AquaBounty)的转基因三文鱼品牌“AquAdvantage”上市,从而使之成为首个获批的供食用转基因动物。
这种三文鱼的主要特点是生长迅速,仅需18个月便能长成——而常规三文鱼需要至少三年。
这件事使人们关注起了转基因鱼这个新事物。
1983年,中国科学院水生生物研究所在世界上率先开展鱼类转基因研究,我国也一直走在鱼类转基因的前列。
1.转基因鱼的定义转基因鱼是首先从鱼体中鉴定出与生长、品质、抗病和抗逆等性状相关的基因, 并将此类基因设计加工, 然后转移到拟进行遗传改良的受体鱼染色体内, 从而培育出优良品种[1]。
转基因鱼研究大多基于提高鱼类生长速度和抗病、抗逆能力, 培育新品种, 开展环境监测以及医学研究等目的。
目前已有金鱼、鲫鱼、鲤鱼、泥鳅、鳟鱼、大马哈鱼、鲶鱼、罗非鱼、鲂、鲑鱼等多种淡水鱼和海水鱼用于转基因研究, 多种哺乳类、鸟类的基因亦被成功整合到鱼类的基因组中[2]。
转基因鱼育种不仅突破了物种间的生殖隔离,拓展了遗传资源利用范围, 而且还减少了育种世代、提高了育种效率。
另外, 对转基因鱼的研究可大大提高鱼类的生长速度和抗逆能力, 使之更易应对恶劣的环境条件, 对扩大优质鱼种养殖范围提供了新的途径, 产生相当可观的经济效益。
同时, 利用转基因鱼可快速、直观地评价环境中污染物的内分泌干扰作用, 对于污染物的防治和监控具有重要的应用和理论研究价值。
集宁区第九中学八年级生物下册第八单元降地生活第二章用药和急救同步测试新版新人教版

第二章用药与急救一、单选题(共15题;共30分)1.下列有关安全用药的正确叙述是()A. 按照自己经验用药B. 非处方药简称为OTCC. 处方药可自行购买,按照使用说明服用D. 阅读药品使用说明,只需了解药品的主要成分和生产日期2.下列症状为毛细血管出血的是( )A. 血液暗红色,流速慢B. 血液红色,从伤口渗出C. 血液鲜红,喷涌而出D. 以上都不对3.关于用药安全与人体健康的叙述,错误的是()A. 非处方药也应按照规定的方法或在医生的指导下服用B. 处方药不能随意购买但可以根据个人的习惯服药C. 健康是指身体和心理等各个方面的良好状态D. 拒绝毒品,远离烟酒是健康向上的生活态度4.关于安全用药的有关知识,下列说法错误的是:A. 处方药需要在医生监控或指导下才能使用B. 非处方药不需要医生的处方就可以购买C. 患者可以按规定的方法服用非处方药D. 长期服用非处方药,不需要医生指导5.判断是否动脉出血,要看()A. 出血时,血液呈暗红色,连续不断地从伤口流出B. 出血时,血液呈红色,从伤口渗出或像水珠一样流出C. 出血时,血液呈鲜红色,从伤口喷出或随心跳一股一股地涌出D. 前三种情况都有可能发生6.医护人员给危重病人输氧,下列相关叙述正确的是()A. 输的是纯氧B. 含有一定的二氧化碳 C. 含有78%的氮气 D. 有一定的一氧化碳7.医用纱布在家庭药箱中属于()A. 医用器具B. 外用药 C. 内服药 D. 特殊药8.“良药苦口利于病”正确用药对于治疗疾病非常重要,下列做法不正确的是()A. 处方药可到医药超市随便购买 B. 非处方药可自己到医药超市购买C. 用药前要仔细阅读药品说明书 D. “是药三分毒”,药物要合理使用9.小红在体育课上不慎受伤,鲜红的血液从伤口喷射而出,在一旁的你可以采用怎样的急救措施呢?()A. 不需处理,让伤口自动止血 B. 用止血带在远心端包扎止血C. 用止血带在近心端包扎止血 D. 清洗伤口,贴上创口贴即可10.下列关于“人工胸外心脏挤压”的操作要领,错误的一项是()A. 挤压时,救护者要站在病人的左侧B. 救护者双手叠放在一起,有节奏地、带有冲击性地用力向下按C. 应按压病人的胸骨下端,使其下陷3~4厘米,然后放松D. 每做一次人工呼吸,就做一次心脏挤压,反复地进行11.了解必要的用药知识和急救技能,对于保障身体健康,挽救生命具有重要意义,下列有关说法正确的是()A. 处方药适用于自我诊断,自我治疗的小伤小病B. 动脉出血,要压迫伤口远心端C. 心肺复苏,先做30次心脏按压,再做2次人工呼吸D. 感冒药是家庭常备药物,过期一两周仍可服用12.下列关于青霉素的叙述,正确的是()A. 青霉素是处分药,必须由医生开处方,在医生的指导下使用B. 随便吃青霉素一定引起人的过敏反应C. 青霉素是广谱药物,什么病都治D. 青霉素是非处分药,可以自行购买13.溺水是夏季青少年容易出现的意外事故.当溺水者被救后,在急救车到达前,若心跳、呼吸停止,必须立即实施心肺复苏.心肺复苏技术的顺序应该是()A. ①③④②B. ③①④②C. ②③①④D. ③②①④14.(2014•玉林)李岚同学一家人准备到北京旅游,由于他的奶奶有心脏病、妈妈晕车,爸爸有胃病,为了能够照顾好家人,他的旅行小药箱里备用的常用药必须要有()A. 乘晕宁、珍视明、咽喉片 B. 止痛素、止泻灵、感康C. 乘晕宁、胃安、白加黑 D. 硝酸甘油、乘晕宁、胃长宁15.(2015•上海)下列有关医药常识的叙述中,错误的是()A. 非处方药不需要医生处方即可购买,按所附药品说明书服用B. 若木刺刺伤较小较浅,可用消毒的镊子将其拔出,伤口涂抹抗生素软膏C. 医生给高烧病人的额头上敷冷毛巾或冰袋来降温D. 患感冒时,可找出家中储备的感冒药,连同抗生素一起服用二、填空题(共4题;共5分)16.生活中难免会出现一些意外,当外伤出血时,发现血液鲜红,流速较快,此时应立即包扎伤口的心端,再送医院作进一步治疗.17.下列是家庭药箱中常放置的物品:纱布碘酒、云南白药、黄连素、眼药水、酒精棉球等,其中属于外用药的是________ ,属于内服药的是________ .18.OTC是________的简称,适于消费者自我诊断和治疗。
转基因的英语作文

转基因的英语作文In the contemporary era of rapid technological advancements, the field of agriculture has not been left behind. One of the most significant breakthroughs in this domain is the development and use of genetically modified organisms (GMOs). These organisms, which have been altered at the DNA level to exhibit certain desirable traits, have sparked a myriad of debates and discussions worldwide.Advantages of GMOs1. Increased Crop Yields: Genetic modification has enabledthe production of crops that are resistant to pests, diseases, and harsh environmental conditions. This has led to a significant increase in crop yields, ensuring food securityfor a growing global population.2. Enhanced Nutritional Value: Scientists have been able to enhance the nutritional content of certain crops through genetic engineering. For example, the development of Golden Rice, which is fortified with Vitamin A, has the potential to combat vitamin deficiencies in developing countries.3. Reduced Use of Chemicals: GMOs that are resistant to pests reduce the need for chemical pesticides. This not only lowers the cost of production but also has positive environmental implications by reducing chemical runoff into water systems.Challenges and Concerns1. Ecological Impact: There are concerns that GMOs could have unintended ecological consequences. For instance, the cross-pollination of GMO crops with wild relatives could lead tothe spread of modified genes into the wild.2. Health Implications: Some critics argue that the long-term health effects of consuming GMOs are not fully understood. There is a call for more extensive research to ensure that these foods are safe for human consumption.3. Ethical Considerations: The ethical implications of manipulating the genetic makeup of organisms are also a point of contention. Some argue that it is not our place to play'God' by altering the natural order of life.Regulation and Public PerceptionThe regulation of GMOs varies greatly from country to country. While some nations have embraced this technology, others have imposed strict regulations or outright bans. Publicperception plays a crucial role in shaping these policies,and it is essential for scientists, policymakers, and the public to engage in informed dialogue to address concerns and misconceptions.ConclusionThe use of genetically modified organisms in agriculture presents both opportunities and challenges. While GMOs havethe potential to address pressing global issues such as food security and malnutrition, it is imperative that we proceed with caution, ensuring that the technology is used responsibly and ethically. As we move forward, it is vital to balance the benefits with the potential risks, and to engage in ongoing research and public education to make informed decisions about the role of GMOs in our food system.。
转基因鱼对生态影响小于预期

自20世纪80年代以来,科学家们开始 研究转基因鱼,并取得了一系列重要 成果。
转基因鱼的研究目的
通过转基因技术,可以改良鱼的品种 ,提高其生长速度、抗病能力等,为 水产养殖业带来更大的经济效益。
生态影响的研究意义
生态安全问题
随着转基因技术的广泛应用,生 态安全问题逐渐成为人们关注的 焦点。转基因鱼对生态的影响是 生态安全领域的重要研究内容之
一。
保障生态环境健康
研究转基因鱼对生态的影响,有 助于了解转基因生物在生态系统 中的行为和影响,为生态环境保
护提供科学依据。
促进可持续发展
通过研究转基因鱼对生态的影响 ,可以评估转基因技术在渔业养 殖中的可持续性,为水产养殖业
的可持续发展提供支持。
02
转基因鱼技术概述
转基因技术原理
转基因技术定义
转基因鱼对生态影响小于预 期
汇报人: 2023-12-14
目录
• 引言 • 转基因鱼技术概述 • 转基因鱼对生态影响分析 • 实验研究与结果展示 • 结论与展望
01
引言转基因ຫໍສະໝຸດ 的研究背景转基因技术发展迅速
转基因鱼的研究历程
随着生物技术的不断发展,转基因技 术已经成为现代生物科学研究的重要 手段。
转基因技术是一种通过改变生物 体的遗传物质,从而改变其性状 的技术。
转基因技术原理
转基因技术的基本原理是利用重 组DNA技术,将外源基因导入受 体细胞或受体生物,使其表达特 定的性状或功能。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Aggregation Behavior in Wildtype and Transgenic Zebrafish Jennifer L.Snekser*,Scott P.McRobert*,Caleb E.Murphy &Ethan D.Clotfelter*Department of Biology,Saint Joseph’s University,Philadelphia,PA,USADepartment of Biology,Amherst College,Amherst,MA,USAIntroductionZebrafish(Danio rerio)have been genetically engin-eered to express greenfluorescent protein(GFP)and redfluorescent protein(RFP)derived from the cnid-arians Aequorea victoria and Discosoma sp.,respectively (Wan et al.2002;Gong et al.2003).To create these transgenicfish,DNA constructs were made via inser-tion of a fast-muscle specific promoter gene into red and GFP genes,which were then microinjected into one-or two-cell zebrafish embryos.Fluorescent pro-teins are expressed in the skeletal muscles and are visible under normal light conditions.Designed for use in developmental and toxicological studies,these fish have been marketed by the aquarium industry under the trademark GloFish TM.To date,only the RFP zebrafish are commercially available.They are thefirst transgenicfish sold to the general public (currently restricted to the U.S.).While zebrafish have been an important biomedi-cal model system for years(Haffter et al.1996;Lele &Krone1996;Heindel2000),our understanding of their social behavior is relatively recent(Kitevski& Pyron2003;Mann et al.2003;Pyron2003;Turnell et al.2003;Peichel2004).However,it is known that zebrafish,like many teleostfishes,form social groups termed‘shoals’(Pitcher1986).Shoaling behavior confers many advantages upon individualfish,inclu-ding reduction of predation risk through a phenom-enon known as the‘confusion effect’(Krause& Ruxton2002).According to the confusion effect model,a predator faced with a group of similarfish has difficulty targeting an individual to attack(Land-eau&Terborgh1986).This phenomenon is directly related to the‘oddity effect,’which predicts that physically distinct(or‘odd’)individuals within a shoal are more likely to be targeted by predators. Therefore,an individual’s ability to recognize and associate with similarfish should provide a selective advantage.Previous studies have shown thatfish prefer to shoal with phenotypically similar individu-als(reviewed in Krause et al.2000).CorrespondenceScott P.McRobert,Department of Biology, Saint Joseph’s University,Philadelphia,PA 19131,USA.E-mail:smcrober@Received:March19,2005Initial acceptance:March29,2005Final acceptance:June2,2005(S.Forbes)AbstractRecent advances in the development and availability of genetically modified animals enable researchers to examine the effects of pheno-typic characters on social behavior.Infish,shoaling behavior is known to be influenced by characteristics such as body coloration,striping pat-tern,body shape,and size.GloFish TM are genetically engineered zebra-fish(Danio rerio)that express redfluorescent protein(RFP),resulting in on overall red coloration under the dark longitudinal stripes.The Glo-Fish pattern is distinct from the light body coloration underlying the dark longitudinal stripes seen in wildtype zebrafish.We presented wild-type and transgenic RFP zebrafish with same-sex shoals of both strains offish in dichotomous choice tests.No preference for either of the shoals was shown,however,both strains showed significant preferences for swimming near shoals vs.swimming near an empty tank compart-ment.When presented with opposite-sex individuals of both strains,no preference was shown by either sex of either strain.Thus,the red body coloration of transgenic zebrafish does not appear to affect choice of social partner,in either a shoaling or a potentially reproductive context.EthologyEthology112(2006)181–187ª2006The AuthorsJournal compilationª2006Blackwell Verlag,Berlin181Shoaling behavior can also provide increased opportunity for mating(Krause&Ruxton2002).By choosing shoal-mates of similar phenotype,indivi-duals naturally form conspecific social groups.This brings males and females together and provides them with an opportunity to preview future mating partners.In zebrafish,a variety of phenotypic cues have been examined to determine the influence on social associations such as body size(Pyron2003),fin length(Kitevski&Pyron2003),group size(Ruhl& McRobert2005),and stripe pattern(McCann et al. 1971;Turnell et al.2003).The effect of overall body coloration has yet to be established.Many other cyp-rinidfishes show a preference for a reddish or ‘brassy’nuptial coloration(Winfield&Nelson1991; Braithwaite&Barber2000),like that of the RFP transgenic zebrafish.These observations lead us to the formulation of our two hypotheses.First,when given the choice between a shoal of wildtype zebrafish and a shoal of RFP transgenic zebrafish that are of the same gender, both strains would choose shoals of their own strain, as predicted by the‘oddity effect.’Second,when placed in a potentially reproductive context,wild-typefish would prefer the redness of the transgenic RFPfish.MethodsAnimal SubjectsWildtype and transgenic zebrafish were obtained from commercial suppliers(World Wide Aquarium, Philadelphia,PA and5-D Tropical in Florida).Fish used for stimulus shoals were housed separately from testfish,in40-or80-l holding tanks.Males and females were housed separately.All tanks were covered on three sides with opaque white paper in order to visually isolate the holding tanks.All tanks were maintained underfluorescent lighting on a 12L:12D cycle,at23–25°C.Fish were fedflake food to satiation daily.Test TanksFor shoaling experiments(Shoaling I–III),test tanks were constructed by dividing aquaria (74·31·31cm)into three compartments(see McRobert&Bradner1998).Each end compartment (18·31·31cm)was separated from the central compartment(38·31·31cm)by clear glass parti-tions sealed with silicone caulk.The central compart-ment was further divided into left and right sides by opaque Plexiglas partitions(25·18cm).These par-titions allowed testfish to swim to either side of the central compartment but prevented a testfish on one side from seeing the stimulusfish on the oppos-ite side.Each of the end compartments contained a heater and boxfilter.Mate choice experiments,in which a testfish was given the opportunity to choose between wildtype and transgenic individuals of the opposite sex were performed in two stages.Mate choice I tests were performed in a7-l polycarbonate tank.Stimulusfish were held in1-l polycarbonate containers placed at each end of the test tank.A line was drawn on the glass to separate the tank into halves.Mate choice II tests,which were more conservative,were per-formed in a modified aquarium(50·25·31cm) divided into three compartments by the addition of glass panels,secured with silicon caulk.Each end compartment(which held the stimulusfish)meas-ured14·25·31cm.Lines drawn on central com-partment glass were used to identify two‘choice zones,’ 6.5cm(approximately two body lengths) from each end compartment.Behavioral ObservationsAll observations were performed between09:00and 17:00hours from Jan.2004to Feb.2005.Prior to each test,a single testfish was placed into the central compartment and allowed1h to acclimate.Testfish were chosen at random from a holding tank and no fish was used more than once for a given experi-ment.Stimulus shoals were also placed into the end compartments of the test tanks prior to testing.Dur-ing the1h of acclimation to the test tank,allfish were able to swim freely in their chambers.For all three shoaling experiments,an equal number of tests were run withfish in the left and right end compart-ments to eliminate the risk of side biases.Each test-ing session consisted of a600-s observation period during which the time(in seconds)that the testfish spent on each side of the central compartment(for the three shoaling experiments and mate choice experiment I)or within the two-body-length‘choice’zones in mate choice experiment II was recorded.In shoaling experiment I,three wildtype zebrafish were placed into one of the end compartments, while the other end compartment was empty.In shoaling experiment II,three transgenic zebrafish were placed into one of the end compartments, while the other was empty.In shoaling experiment III,three wildtype zebrafish were placed into oneAggregation Behavior in Wildtype and Transgenic Zebrafish J.L.Snekser,S.P.McRobert,C.E.Murphy&E.D.ClotfelterEthology112(2006)181–187ª2006The Authors 182Journal compilationª2006Blackwell Verlag,Berlinend compartment and three transgenic zebrafish were placed into the other end compartment.Both wildtype and transgenicfish were used as testfish (in the central compartment)in each experiment. The end compartments always containedfish of the same gender as the focal testfish.For the mate choice experiments,testfish were given the choice between a single wildtype zebrafish and a single transgenic zebrafish of the opposite sex. Mate choice experiment I was performed with wild-type testfish only,mate choice II examined the pref-erences of both wildtype and transgenic testfish. Statistical AnalysisFor each shoaling experiment one-sample t-tests, compared with a null expectation(0.5total time), were used to compare time spent on each side of the tank.For the mate choice experiments,paired sam-ple t-tests were used to compare time spent in each part of the test tank.ResultsShoaling ExperimentsIn shoaling experiments I and II,in which testfish were given the choice between a shoal of three ze-brafish(either transgenic or wildtype)and an empty compartment,the testfish always showed a signifi-cant preference for the compartment containing the shoal offish(responses to a wildtype shoal and a transgenic shoal,respectively:n=20,wildtype female testfish:t=5.45,p<0.001and t=8.39, p<0.001;wildtype male testfish:t=3.56,p<0.01 and t=3.26,p<0.01;transgenic female testfish: t=4.31,p<0.001and t=3.62,p<0.01;transgenic male testfish:t=6.28,p<0.001and t=5.08, p<0.001).However,no significant preferences were shown in shoaling experiment III,when testfish were given a choice between a shoal of three wild-typefish and a shoal of three transgenicfish (n=20,wildtype female testfish:t=0.21,p=0.84; wildtype male testfish:t=0.48,p=0.64;transgenic female testfish:t=0.18,p=0.86;transgenic male testfish:t=0.042,p=0.97,Fig.1).Mate Choice ExperimentsIn mate choice experiment I,wildtype male zebra-fish spent significantly more time swimming near a transgenic female than near a wildtype female (n=35,t= 2.54,p<0.05).Similarly,wildtype females spent significantly more time swimming near a transgenic male,than a wildtype male (n=35,t=2.40,p<0.05;Fig.2).However,in the more conservative mate choice experiment II,wildtype males and females showed no significant preference for either wildtype or trans-genicfish of the opposite sex(n=20,wildtype female testfish:t=0.65,p=0.52;wildtype male testfish:t=2.02,p=0.058).Similarly,transgenic male and female testfish showed no significant pref-erence for either wildtype or transgenicfish of the opposite sex(n=20,transgenic female testfish: t=0.68,p=0.51;transgenic male testfish:t=1.47, p=0.16,Fig.3).DiscussionThe development of a red transgenicfish creates an opportunity for studying the importance of body col-oration in both social and reproductive contexts. This question is important asfish are known to dis-criminate between potential shoal-mates on the basis of phenotype,including body coloration.In mollies (Poecilia latipinna),for example,black and white color morphs choose shoal-mates of similar color-ation(McRobert&Bradner1998).Interestingly,the results presented here show that the red coloration found in transgenic GloFish TM does not influence choice of same-sex shoal-mates.Yet,in some cir-cumstances,body coloration does influence prefer-ence for individual,opposite-sexfish.By shoaling withfish with a similar phenotype,it is thought thatfish reduce the likelihood that they will be preyed upon(Landeau&Terborgh1986). Thus,choosing shoal-mates wisely is evolutionarily beneficial.Many studies have shown thatfish will tend to shoal withfish that are alike in appearance, based not only on body coloration(McRobert& Bradner1998),but also on body shape(Hauser et al. 1998),body size(Pritchard et al.2001;Pyron2003), parasite load(Krause&Godin1996),and body pattern(McCann et al.1971;Turnell et al.2003; Engeszer et al.2004).Through the use of transgenic zebrafish,this study attempts to hold other pheno-typic characters constant and focus solely on the role of coloration.In shoaling experiment III,in which testfish were given the choice between a shoal of similarly coloredfish and dissimilarly coloredfish, no preference was shown for either shoal.The lack of preference for,or avoidance of,the RFP trans-genic zebrafish may indicate that overall body color-ation may not play an important role in shoal-mate preference in this species.J.L.Snekser,S.P.McRobert,C.E.Murphy&E.D.Clotfelter Aggregation Behavior in Wildtype and Transgenic Zebrafish Ethology112(2006)181–187ª2006The AuthorsJournal compilationª2006Blackwell Verlag,Berlin183Despite the fact that the red body coloration ofGloFish TM does not influence shoaling choices,itseems apparent that zebrafish can detect the colorred.Zebrafish are tetrachromatic,with four types of cones,in addition to rods,that absorb light at red,green,blue,and UV wavelengths (Robinson etal.Fig.1:Mean association time (±SE)of wild-type females and males (a)and transgenicfemales and males (b)when given a choice inthree shoaling preference experiments:(1)three RFP transgenic zebrafish (gray)and anempty chamber (white);(2)three wildtypezebrafish (black)and an empty chamber(white);and (3)between three RFP transgeniczebrafish (gray)and three wildtype fish (black).**p <0.01;***p <0.001Fig.2:Mean association time (±SE)of wild-type and RFP transgenic females and maleswhen given a choice in an experiment thatcontained one RFP transgenic (gray)or onewildtype fish (black)of the opposite sex in adichotomous mate choice test.*p <0.05Aggregation Behavior in Wildtype and Transgenic Zebrafish J.L.Snekser,S.P.McRobert,C.E.Murphy &E.D.ClotfelterEthology 112(2006)181–187ª2006The Authors184Journal compilation ª2006Blackwell Verlag,Berlin1993).In fact,zebrafish actually have two distinctred opsin photopigments:one maximally sensitive at558nm and the other at 548nm (Takechi &Kawa-mura 2005).In behavioral screens,zebrafish haveexhibited the optokinetic response,displayingsmooth pursuit and saccade eye movements inresponse to illuminated rotating stripes under bothwhite and red lights (Brockerhoff et al.1997).TheRFP of the transgenic zebrafish has an emission spec-trum of 550–675nm,with emission peaks of 558and 583nm (Matz et al.1999).It is thereforeassumed that the zebrafish can detect the red color-ation of the transgenic fish.To further demonstrate that the zebrafish inthese studies were able to see red,wildtype zebra-fish females were presented with a choice betweenthree female wildtype zebrafish and an emptychamber,with red light (600–620nm)as the onlylight source.In these assays the test fish spent sig-nificantly more time near the shoal of fish(n =10,t =4.333,p <0.01).If the zebrafish wereunable to see in this limited red light condition,then no preference would have been expected.The strong preference for the shoal of fish showsthat wildtype zebrafish are capable of perceivingthe color red.To determine whether ultraviolet cues affectshoaling decisions,wildtype test fish were given thechoice between a shoal of three wildtype fish andthree transgenic fish under ultraviolet lights.Inthese tests,as in tests under fluorescent lighting,nosignificant choice was made by the test fish (n =20,transgenic female test fish:t =0.950,p =0.354).These results indicate that ultraviolet cues have noeffect on shoaling preferences of zebrafish.In a dichotomous choice shoaling study done with modified photographs of zebrafish,wildtype fish showed a preference for striped fish over fish lacking stripes,even if the stripe pattern was altered so that the normal light and dark stripes were reversed (McCann et al.1971).Turnell et al.(2003)recently used video stimuli to demonstrate that the orienta-tion of zebrafish stripe patterns significantly affected associative preferences.These results,along with the results from the current study,indicate that it may be the presence of stripes and not general body col-oration that is the important cue in shoal-mate pref-erence in zebrafish.With respect to mate choice,female zebrafish,as in many vertebrates,are the ‘choosier’sex (Pyron 2003).In some species of fish,such as two-spotted gobies (Gobiusculus flavescens ),three-spined stickle-backs (Gasterosteus aculeatus ),and brook sticklebacks (Culaea inconstans ),males also show a preference for females,usually in correlation with the large size of the female,and also for the female’s coloration (McLennan 1995;Amundsen &Forsgreen 2001).The larger size and noticeable ‘nuptial’coloration of the females in these species are indicators of sexual receptiveness,which would lead to a more successful mating attempt by the soliciting males.In zebrafish,however,females do not gain a distinct coloration associated with sexual behavior.The level of sexual dimorphism in zebrafish is low,with females being slightly more robust and males developing a slightly more yellowish coloration.The red coloration of the transgenic zebrafish only seemed to affect mate choice within certain experi-mental parameters.In the first mate choice experi-ment,wildtype fish showed a preferencefor Fig.3:Mean association time (±SE)of wild-type and RFP transgenic females and maleswhen given a choice in an experiment thatcontained one RFP transgenic (gray)or onewildtype fish (black)of the opposite sex in adichotomous mate choice testJ.L.Snekser,S.P.McRobert,C.E.Murphy &E.D.Clotfelter Aggregation Behavior in Wildtype and Transgenic Zebrafish Ethology 112(2006)181–187ª2006The AuthorsJournal compilation ª2006Blackwell Verlag,Berlin 185transgenicfish of the opposite sex over wildtypefish of the opposite sex.However,in these tests,choice was determined by the testfish swimming in the general vicinity of the stimulusfish.In the more conservative mate choice experiment II,in which choice was determined by the testfish swimming within two body lengths of the stimulusfish for a significant fraction of the observation period,no sig-nificant preference was shown for the transgenic fish.Similarly,the transgenicfish showed no prefer-ence for either strain when tested in the mate choice experiment II.These results appear to be supported by Seah,W.K.,Li, D.,Chen,M.,&Gong, Z.(unpublished data),who reported a lack of advan-tage for transgenic zebrafish in a dichotomous choice test(see Gong et al.2003).As red coloration is not usually apparent in zebra-fish,it will be interesting to examine patterns of mate choice using a stimulus that may invoke a more natural bias,such as the GFP transgenic zebra-fish that will soon be available to the general public. Future studies should also focus on the number and success of actual spawning attempts,rather than simply the preferences indicated in dichotomous tests such as those presented here.These results will give a better indication of what role,if any,body coloration plays in zebrafish mate choice. AcknowledgementsWe thank C.McRobert,M.G.Manning,and N. Ruhl for assistance with trials andfish care.This work was funded by a NSF GK-120139303grant awarded to Saint Joseph’s Biology Department,an internal grant from the Saint Joseph’s University chapter of Sigma Xi,and funding from the Dean of Faculty’s office at Amherst College.Literature CitedAmundsen,T.&Forsgreen,E.2001:Male mate choice selects for coloration infish.Proc.Natl.Acad.Sci.U.S.A.98,13155—13160.Braithwaite,V.A.&Barber,I.2000:Limitations to col-our-based sexual preferences in three-spined stickle-backs(Gasterosteus aculeatus).Behav.Ecol.Sociobiol. 47,413—416.Brockerhoff,S.E.,Hurley,J.B.,Niemi,G.A.&Dowling, J.E.1997:A new form of inherited red-blindness iden-tified in zebrafish.J.Neurosci.17,4236—4242. Engeszer,R.E.,Ryan,M.J.&Parichy,D.M.2004. Learned social preference in zebrafish.Curr.Biol.14, 881—884.Gong,Z.,Wan,H.,Tay,T.L.,Wang,H.,Chen,M.& Yan,T.2003:Development of transgenicfish for orna-mental and bioreactor by strong expression offluores-cent proteins in skeletal muscle.Biochem.Biophys. mun.308,58—63.Haffter,P.,Granato,M.,Brand,M.,Mullins,M.C.,Ham-merschmidt,M.,Kane,D.A.,Odenthal,J.,van Eeden, F.J.M.,Jiang,Y.-J.,Heisenberg,C.-P.,Kelsh,R.N., Furutani-Seiki,M.,Vogelsang,E.,Beuchle,D.,Schach, U.,Fabian,C.&Nu¨sslein-Volhard,C.1996:The identi-fication of genes with unique and essential functionsin the development of the zebrafish,Danio rerio.Devel-opment123,1—36.Hauser,L.,Carvalho,G.R.&Pitcher,T.J.1998:Genetic population structure in the Lake Tanganyika sardines Limnothrissa miodon.J.Fish Biol.53(Suppl.A), 413—429.Heindel,J.2000:Zebrafish:a model organism for bio-medical research.Environ.Health Perspect.108,A419. Kitevski,B.&Pyron,M.2003:Female zebrafish(Danio rerio)do not prefer mutant longfin males.J.Freshw. Ecol.18,501—502.Krause,J.&Godin,J.-G.J.1996:Influence of parasitism on shoal choice in banded killifish(Fundulus diaphanus, Teleostei,Cyprinodontidae).Ethology102,40—49. Krause,J.&Ruxton,G.D.2002:Living in Groups. Oxford Univ.Press,New York.Krause,J.,Butlin,R.K.,Peuhkuri,N.&Pritchard,V.L. 2000:The social organization offish shoals:a test of the predictive power of laboratory experiments for the field.Biol.Rev.75,477—501.Landeau,L.&Terborgh,J.1986:Oddity and the‘confu-sion effect’in predation.Anim.Behav.34, 1372—1380.Lele,Z.&Krone,P.H.1996:The zebrafish as a model system in developmental,toxicological and transgenic research.Biotechnol.Adv.14,57—72.Mann,K.D.,Turnell,E.R.,Atema,J.&Gerlach,G. 2003:Kin recognition in juvenile zebrafish(Danio rerio) based on olfactory cues.Biol.Bull.205,224—225. Matz,M.V.,Fradkov,A.F.,Labas,Y.A.,Savitsky,A.P., Zaraisky,A.G.,Markelov,M.L.&Lukyanov,S.A. 1999:Fluorescent proteins from nonbioluminescent Anthozoa species.Nat.Biotechnol.17,969—973. McCann,L.I.,Kohen,D.J.&Kline,N.J.1971:The effects of body size and body markings on nonpolarized schooling behavior of zebrafish(Brachydanio rerio).J. Psychol.79,71—75.McLennan,D.A.1995:Male mate choice based upon female nuptial coloration in the brook stickleback,Cul-aea inconstans.Anim.Behav.50,213—221. McRobert,S.P.&Bradner,J.1998:The influence of body coloration on shoaling preferences infish.Anim. Behav.56,611—615.Aggregation Behavior in Wildtype and Transgenic Zebrafish J.L.Snekser,S.P.McRobert,C.E.Murphy&E.D.ClotfelterEthology112(2006)181–187ª2006The Authors 186Journal compilationª2006Blackwell Verlag,BerlinPeichel,C.L.2004:Social Behavior:How dofishfind their shoal mate?Curr.Biol.14,R503—R504. Pitcher,T.J.1986:Functions of shoaling behavior in teleosts.In:The Behavior of Teleost Fish(Pitcher,T.J., ed.).Chapman&Hall,London,pp.294—337. Pritchard,V.L.,Lawrence,J.,Butlin,R.K.&Krause,J. 2001:Shoal choice in zebrafish,Danio rerio:the influ-ence of shoal size and activity.Anim.Behav.62, 1085—1088.Pyron,M.2003:Female preferences and male–male interactions in zebrafish(Danio rerio).Can.J.Zool.81, 122—125.Robinson,J.,Schmitt,E.A.,Harosi,F.I.,Reece,R.J.& Dowling,J.E.1993:Zebrafish ultraviolet visual pig-ment:absorption spectrum,sequence,and localization. Proc.Natl.Acad.Sci.U.S.A.90,6009—6012.Ruhl,N.&McRobert,S.P.2005:The effect of sex& shoal size on shoaling behavior in zebrafish(Danio rerio).J.Fish Biol.67,1—9.Takechi,M.&Kawamura,S.2005:Temporal and spatial changes in the expression pattern of multiple red and green subtype opsin genes during zebrafish develop-ment.J.Exp.Biol.208,1337—1345.Turnell,E.R.,Mann,K.D.,Rosenthal,G.G.&Gerlach, G.2003:Mate choice in zebrafish(Danio rerio)ana-lyzed with video-stimulus techniques.Biol.Bull.205, 225—226.Wan,H.,He,J.,Ju,B.,Yan,T.,Lam,T.J.&Gong,Z. 2002:Generation of two-color transgenic zebrafish using the green and redfluorescent protein reporter genes gfp and rfp.Mar.Biotechnol.4,146—154.Winfield,I.J.&Nelson,J.S.(Eds.)1991:Cyprinid Fishes;Systematics,Biology and Exploitation.Chap-man and Hall Ltd,London.J.L.Snekser,S.P.McRobert,C.E.Murphy&E.D.Clotfelter Aggregation Behavior in Wildtype and Transgenic Zebrafish Ethology112(2006)181–187ª2006The AuthorsJournal compilationª2006Blackwell Verlag,Berlin187。