抗菌肽AD基因的改造及在毕赤酵母中的表达
东北林蛙皮肤抗菌肽基因在毕赤酵母中表达条件的优化

( C o l l e g e o fWi l a l  ̄ f e R e s o u r c e s ,N o r t h e a s t F o r e s t r y U n i v e r s i t y ,H a r b i n ,1 5 0 0 4 0 ,C h i n a)
1 S T g e n e o f Ra n a d y b o w s k i i s k i n a n t i mi c r o b i l a p e p t i d e s .T h e d y b o ws k i n 一1 S T g e n e wa s s y n t h e s i z e d b y P C R a n d c l o n e d i n t o Pi c h a p a s t o r i s e x p r e s s i o n v e c t o r p P I C 9 K t o g e n e r a t e i n d u c i b l e e x p r e s s i o n v e c t o r p P I C 9 K —d y b o ws k i n一1 S T .T he l i n e a i f z e d p l a s mi d p P I C 9 K —d y b o ws k i n一1 S T w a s t r a n s f o r me d i n t o P i c h i a p a s t o r i s y e a s t G S 1 1 5 .E x p r e s s i o n Wa s i n d u c e d b y BMMY c u l - t u r e me d i u m. Us i n g t h e s i n g l e v a ia r b l e me t h o d a n d o ah o g o n a l e x p e r i me n t me t h o d, w e o p t i mi z e d i n d u c i n g c o n d i t i o n s b y c h a n — g i n g t h e t e mp e r a t u r e, t h e i f n a l c o n c e n t r a t i o n o f me t h a n o l a n d t h e i n d u c t i o n t i me .T h e o p t i ma l c o n d i t i o n s W e r e d e t e mi r n e d i n v i t r o b y S DS—P AGE e l e c t r o p h o r e s i s d e t e c t i o n a n d a n t i b a c t e r i l a t e s t s .F o r t h e o p t i ma l i n d u c t i o n c o n d i t i o n s , me ha t no l i f n l a c o n c e n t r a t i o n,i n d u c t i o n t i me,a n d i n d u c t i o n t e mp e r a t u r e we r e 0 . 5 % ,7 2 h a n d 2 8 ' 1 2, r e s p e c t i v e l y . U n d e r t h e s e c o n d i —
重组牛乳铁蛋白肽衍生肽设计及其在毕赤酵母中的表达

重组牛乳铁蛋白肽衍生肽设计及其在毕赤酵母中的表达
吕自力4*,张恩鹏2,郭爱珍2,罗斌4梁鑫4单旭东4庄静4张霞4王亮2
1成都中医药大学医学与生命科学学院,四川成都,10041) 2(江苏大学食品与生物工程学院,江苏镇江,1201)
乳铁蛋白是一种由哺乳动物黏膜上皮细胞分泌
产生的分子质量为80 kDa的糖基化蛋白,在不同物 种间具有高度的同源性⑴。乳铁蛋白也是转铁蛋白
家族的一员,对铁离子具有高度的
,被认为是
人类和哺乳动物防御系统的基本组成之一⑵。乳铁
蛋白肽是乳铁蛋白在酸
下经胃蛋白酶消 g
的一种短肽,具有良好的耐热性、非抗原性、免疫调
24 2019 Vol. 45 No. 15 (Total 387)
促进牛乳铁蛋白肽与细菌细胞膜脂多糖的结合 ,带正
荷的氨基酸残基与带负电的磷脂发生
互作
用,导致细胞膜穿孔,细胞内容物泄漏[9]。不同于传
生素的抑菌 ,牛乳铁蛋白新型抑菌机制有利
其作为抗生素替代药物的研发和应用。由于天然
来源有限且 工艺
达后发现发酵上清液具有较强的抗菌活性°实验研
究为
优 乳铁蛋白肽的抗菌活
基°
1仪器与试剂
GeoaPluscs Xcell™电转仪,美国伯乐公司;
1645050型DNA电泳仪,美国伯乐公司;EppeudorS
5415R型高速离心机,德国EppeudorS f
; SW-
CJ系
工作台,苏
技术有 【司;
QY200; 温
cmH)第8位Ary: 为Try后,其抗菌活性明显提
抗菌肽LL-37在毕赤酵母SMD1168中的高效表达及活性鉴定

21 年 2 月 00
中 国 预 防 兽
医 学 报
V0 . 1 32. o2 N . Fe . 2 0 b 0l
C i e e J u a o r v n i e Ve ei a y M e i ie h n s o r l f e e t trn r d cn n P v
酵母细胞 S MD16 。经 Z oi 抗性 筛选 ,得到高拷 贝转化子 ,P R检 测表 明 L 一 18 ec n C L3 7基 因与毕赤酵母 染色体稳定
结合 在 G P启动 子 调控 下 ,L . A L3 7蛋 白获得 分 泌表 达 ,其 上 清表 达 量 约 为 27 /。表 达 产物 耐 热 性 强 ,在 3 L mg 10℃ 条件 下 3 n内仍保 持 一 定 的 抗 菌 活性 。表 达 产物 对 大肠杆 菌 D 5 、大 肠杆 菌 D 1 猪链 球 菌 的最 小 0 0 mi H 3和
Ab ta t n o d r t mp o e e p e so fa t c o i lp p i e LL 3 n P c i a tr,L 一 7 g n s d s n d a d s r c :I r e o i r v x rs i n o n i r b a e t 一 7 i ih a p s i L 3 e e wa e i e n mi d o g s n h sz d t n l d h at l y c d n f P a t r a e n t e g n e u n e n o i g h ma n ia tra p p i e y t e ie o icu e t e p r a i o o s o .p s i b s d o h e e s q e c s e c dn u n a t ce i l e t i t o s b d L 一7 L 3 .T e mo i e L 一 7 e e h d f d L 3 g n wa co e i t te GAP 【 i s l n d n o h p Z o A v co n  ̄a s r d n o .p so i MD 1 6 b e tr a d n f me i t P o a tr S s 8 y 1
毕赤酵母中高效引导外源蛋白分泌的信号肽的研究

毕赤酵母中高效引导外源蛋白分泌的信号肽的探究毕赤酵母被广泛应用于外源蛋白的表达和分泌,但其分泌机制依旧存在瓶颈。
信号肽作为外源蛋白分泌的关键信号,可以增进蛋白的正确折叠和定位。
本探究合成了多个信号肽并测试了其诱导外源蛋白分泌的效果。
结果显示,其中一个信号肽在毕赤酵母中具有高效的引导外源蛋白分泌的作用,并可提高外源蛋白的表达量。
这一探究为毕赤酵母外源蛋白分泌的机制探究和工业应用提供了新思路。
关键词:毕赤酵母、信号肽、外源蛋白分泌、表达、折叠定位正文:引言毕赤酵母是一种广泛应用于外源蛋白表达和分泌的真菌,其工业用途广泛,包括生产酶、生物肥料、食品添加剂等。
外源蛋白的表达和分泌是毕赤酵母应用的基础,因此其分泌机制的探究具有重要的理论和应用价值。
信号肽被认为是蛋白在细胞内穿过细胞膜从而被分泌到细胞外所必需的关键信号。
近年来,通过信号肽的调控已经在多种真菌中实现了外源蛋白的高效表达和分泌。
因此,寻找高效的信号肽,探究其引导外源蛋白分泌的机制,具有重要的应用前景。
材料和方法合成了11种可能具有对毕赤酵母分泌效果的信号肽,并转化到毕赤酵母中。
以GFP作为模型蛋白,不同信号肽在GFP表达和分泌过程中的诱导效果进行比较。
同时,测定了其中一个信号肽对外源蛋白表达量和分泌量的影响,并通过Western blot分析外源蛋白的分泌效果和分泌途径分析来探究信号肽的作用机制。
结果在11种信号肽中,有一个信号肽(称为SgPEP1)能够显著增加GFP的分泌效率,同时提高了外源蛋白的表达量。
Western blot和分泌途径分析显示,SgPEP1作用于胞内和胞外蛋白的定位和折叠,增进蛋白正确地进入胞外。
谈论本探究中发现的SgPEP1信号肽在毕赤酵母外源蛋白表达和分泌中具有高效的引导作用,这为改善毕赤酵母的表达和分泌效率提供了新思路。
在信号肽的机理探究中,需要进一步探究其在外源蛋白折叠中的作用方向和机制,以便更好地控制蛋白的定向和拓扑。
拟穴青蟹抗菌肽scygonadin在毕赤酵母中的表达及其抗菌活性

泉州 师范学 院学 报
2 1 年 1 月 01 1
各种酶、 N Lgt nKt 自 TKa a公司; D A i i i购 ao a R 胰蛋白胨和酵母提取物购 自 O i x d公司 ; o 丙烯 酰胺 、 N N, 亚
甲基双丙烯酰胺购 自 B I B 公司 ; i u k e E t co i购 自Q G N公 司; 48 酸盐购 自 Qa i l x at n t q cG r i K E G 1硫 华美生物工程 有 限公司; 质粒小量纯化试剂盒 购 自 U gn 公 司; -ee 硅珠 D A提取试剂盒购 自鹭隆公 司; l-oyPci E - N MutC p i a x i h pes nK t 自 Iv rgn rsi i购 o ni oe 公司 ; t 低分子量标准蛋 白质 Makr 自B ̄R D公 司 ; re 购 I A 其他试剂 均为分析纯.
中图分类号 : 8 Q7 文献标识码 : A 文章编号 :0 98 2 (0 10 —0 1I 10 —24 2 1) 60 1一 i
抗菌肽具 有广谱 高效抗 菌活性 , 不易产生抗性 , 在生物体 天然免疫反应 中起着非 常重要 的作用. 抗菌肽分 布广泛 , 在动物、 植物和微生物 中均有发现[ 抗菌肽 被认为是鱼 、 、 1 . 虾 贝等防御系统的主要成分之一. 甲壳动
在Hale Waihona Puke 毕赤酵母中的表达及其抗菌活性 许婉 芳 , 嘉华 陈慧芸 谢 ,
(. 1泉州师范学院 化学与生命科学学 院, 福建 泉州 320 ; 600 310) 605 2厦门大学 , . 海洋与环境学 院 , 近海海洋环境科学 国家重点实验室 , 福建 厦门
摘
要: 采用 P R方法克 隆获得拟 穴青蟹抗菌肽 syo a i 因成熟肽片段 , 其连接到 Pi i atrs C cg n d n基 将 ca s i h p o
酵母双杂交ad自激活验证步骤

酵母双杂交ad自激活验证步骤
酵母双杂交(Yeast Two-Hybrid)是一种常用的蛋白质相互作用研究技术。
下面是酵母双杂交AD自激活验证的步骤:
1. 构建酵母双杂交AD靶蛋白的表达载体:将目标蛋白的编码序列克隆到酵母双杂交AD表达载体中,将其与AD激活域相连,以使目标蛋白能够激活报告基因的表达。
2. 转化AD靶蛋白表达载体到酵母菌株中:通过酵母转化方法将AD靶蛋白表达载体导入酵母菌中,使其能够表达目标蛋白并激活报告基因。
3. 培养转化后的酵母菌株:将转化后的酵母菌株分别培养在选择性培养基上,其中包含AD靶蛋白表达载体所对应的选择性标记物,以筛选出成功转化的酵母菌株。
4. 鉴定AD自激活:通过观察报告基因的表达情况,若转化后的酵母菌株在选择性培养基上形成克隆,表明AD靶蛋白具有自激活能力。
此时需要通过相应的对照实验来确认AD靶蛋白的自激活性质。
需要注意的是,酵母双杂交中AD自激活验证的结果需要慎重解读,因为自激活可能会产生误报。
因此,在进行酵母双杂交实验时,通常需要配对对照实验来排除自激活的影响,以确保结果的准确性。
毕赤酵母表达蛋白步骤

毕赤酵母表达蛋白步骤一、引言毕赤酵母(Pichia pastoris)是一种常用的真菌表达系统,被广泛应用于蛋白质的表达和生物技术研究中。
其优势包括高表达水平、易于培养和操作、能够正确折叠复杂蛋白等。
本文将介绍毕赤酵母表达蛋白的步骤。
二、构建表达载体毕赤酵母表达系统的关键是表达载体的构建。
首先,需要选择适合的表达载体,常用的有pPIC6、pPICZα等。
然后,在载体上选择合适的启动子和信号序列,以确保蛋白质能够被正确表达和分泌。
同时,还需要在表达载体上加入选择标记,如His标签、FLAG标签等,以便后续的蛋白质纯化和检测。
三、转化毕赤酵母将构建好的表达载体转化入毕赤酵母中,使其成为表达宿主。
转化方法包括电击转化、化学转化等。
其中,电击转化是常用的方法,通过电击脉冲使毕赤酵母细胞膜发生破裂,使表达载体进入细胞内。
转化后,将细胞培养在选择性培养基上,筛选出带有表达载体的毕赤酵母克隆。
四、表达蛋白经过转化筛选后,得到含有目标蛋白表达载体的毕赤酵母克隆。
接下来,需要将克隆进行培养,在适当的条件下诱导蛋白的表达。
常用的诱导剂包括甲醇、巯基乙醇等,通过加入适量的诱导剂,可以使目标蛋白得到高效表达。
五、蛋白纯化在蛋白表达后,需要进行蛋白纯化,以获得纯度较高的目标蛋白。
常用的纯化方法包括亲和层析、离子交换层析、凝胶过滤层析等。
在选择纯化方法时,需要根据目标蛋白的性质和需求进行合理选择。
同时,可以利用加入的选择标记,如His标签,通过亲和层析纯化进行快速高效的纯化。
六、蛋白鉴定和功能分析蛋白纯化后,需要进行蛋白的鉴定和功能分析。
常用的鉴定方法包括SDS-PAGE、Western blot等,可以确定蛋白的分子量和纯度。
功能分析则可以通过生物学实验来进行,如酶活测定、结合实验等,以验证目标蛋白的功能。
七、应用和展望毕赤酵母表达系统在生物技术和蛋白质研究领域有着广泛的应用。
通过该系统,可以高效表达各种蛋白,包括抗体、酶和重组蛋白等。
毕赤酵母基因组编辑技术原理
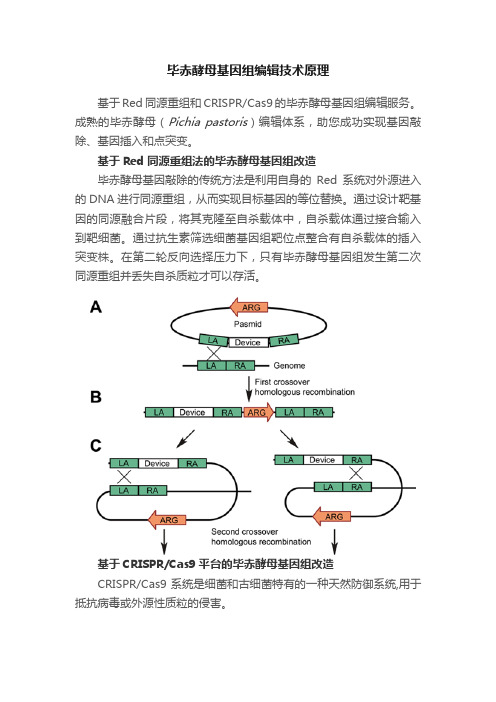
毕赤酵母基因组编辑技术原理基于Red同源重组和CRISPR/Cas9的毕赤酵母基因组编辑服务。
成熟的毕赤酵母(Pichia pastoris)编辑体系,助您成功实现基因敲除、基因插入和点突变。
基于Red同源重组法的毕赤酵母基因组改造毕赤酵母基因敲除的传统方法是利用自身的Red系统对外源进入的DNA进行同源重组,从而实现目标基因的等位替换。
通过设计靶基因的同源融合片段,将其克隆至自杀载体中,自杀载体通过接合输入到靶细菌。
通过抗生素筛选细菌基因组靶位点整合有自杀载体的插入突变株。
在第二轮反向选择压力下,只有毕赤酵母基因组发生第二次同源重组并丢失自杀质粒才可以存活。
基于CRISPR/Cas9平台的毕赤酵母基因组改造CRISPR/Cas9系统是细菌和古细菌特有的一种天然防御系统,用于抵抗病毒或外源性质粒的侵害。
研究内容•毕赤酵母基因敲除(Pichia pastoris gene knockout)•毕赤酵母基因敲入(Pichia pastoris gene knockin)•毕赤酵母基因点突变(Pichia pastoris gene point mutation)下游应用•抗生素及重要工业用酶•发现新的基因功能•优化代谢通路,提升代谢产物产量,实现工业化生产参考文献:•Selle K, Barrangou R. Harnessing CRISPR–Cas systems for bacterial genome editing[J]. Trends in microbiology, 2015, 23(4): 225-232.•Zerbini, F., Zanella, I., Fraccascia, D., König, E., Irene, C., & Frattini, L. F., et al. (2017). Large scale validation of an efficient CRISPR/Cas-based multi gene editing protocol in Escherichia coli.Microbial Cell Factories, 16(1), 68.。
鱼类抗菌肽基因工程表达系统概述

bo 鉴定 , l t 发现 目的蛋 白与预期蛋 白的分 子量相 吻合 , 并能与特异抗 体 a t i nh i s特异性 结合 , 明大菱 鲆抗 菌 表 肽 hpii 因在酵母菌 中得到 了成功表达 ec n基 d 6。蔡 晶
晶等利用 P R技术扩增 获得的大小 约 20 p的黑鲷抗 C 5b
有表达 率高、 产物可分泌、 背景蛋白少 、 易于纯 化、 可高 密度发 酵等优点 , 合外 源蛋 白的表达 。 目前 鱼类 真 适 核表 达 多 采 用 毕 赤 酵母 表 达 系 统。李 伟 将 大 菱 鲆
( c hh lu ciu) So tam s t ns 的抗 菌肽 hp i n基 因成 熟肽 p mx ec i d 序列经 过部分 改 造后 采用 P R 方法 克 隆到毕 赤 酵母 C 胞 内表 达载体 p A Z 中, 建 了重 组胞 内表 达 载体 G PB 构
菌肽 h pi n A ecd ( S—hp2 与毕赤酵母 ( i i p s r ) i ec ) Pv a at i h os
是机体炎症反 应 的组 成 部分 ,是 宿主 防御 细菌 、 菌 真 等病原微生物入侵的重要分子屏障 2。
鱼类抗菌肽在鱼体受 到损伤和感染时能快 速合成
并迅速扩散来保护鱼体抵抗 侵染。随着高密 度的饲养 和抗生素 的大量 使用 , 致 大量 的抗 、 导 耐药 性 细菌 出
现, 传统抗生素类 物质 的效力 和环境 的代谢 压力增大 。
抗菌肽是一类 内源性的具有广谱抗 菌活性 的天然 生物小分子肽 , 广泛存 在于生 物界 , 并成 为非 特异性免
疫进化过程 中的重要组成 … 。抗 菌肽一 般 由 2 0到 6 0
段, 构建融合表 达 载体 p E G X—fe , hp 转化 至 大肠 杆 菌
大菱鲆抗菌肽基因酵母表达载体的构建及表达

・ 1 6 ・
大 菱 鲆 抗 菌肽 基 因酵 母 表 达 载体 的构 建 及 表 达
李 伟 ( 大学生 长江 命科学 学院, 荆州 442) 湖北 305
陈林( 松
源持利重开实 中水科研 院海产 究 可续用点放验 国产学究 黄水研 )
[ 要] 大 菱 鲆 ( cp tamu m xn sh pii 因 成 熟肽 序 列 经过 部 分 改造 后 采用 P R方 法克 隆 到 毕 摘 将 S o hh l s a iu ) ec n基 d C 赤酵 母 胞 内表 达 载 体 p P B 中 , 建 了重 组胞 内表 达 载 体 p P B th p hs GA Z 构 GA Z — b e 6 i。用 Bl 性 化 重 组 质 nI线 粒后 , 电击 转 化 毕 赤 酵 母 S 1 6 。经 过 Zo i 选 和 P R 扩 增 转 化 子 基 因组 筛 选 , 到 了数 株 基 因 工 程 MD 18 ec n筛 C 得 酵 母 菌 。 通 过 S —P GE和 Wetr lt 定 , 现 目的 蛋 白与 预 期 蛋 白的 分 子 量 相 吻 合 , 能 与特 异 抗 Ds A senbo 鉴 发 并
有研 究表 明 , 鱼类 抗菌 肽在鱼 体受 到 损 伤 和感 染 时 , 菌 肽 快 速 合成 并 迅 速 扩 散来 保 护 鱼 体 抵抗 侵 抗 染L 。随着 我 国海水养 殖业 的迅速 发展 , 5 ] 海水 养殖 病害 日益 严 重 。高 密 度 的饲 养环 境 条 件 和抗 生 素物 质
的持续 大量使 用 , 导致 大量 的抗 、 耐性 细菌 出现 , 学合 成抗 生素 物质 的效力 降低 和环 境 的代谢 压力 增大 。 化
在 目前抗病 、 菌疫苗 短缺 的情况 下 , 找新 的天然 活性 的抗 生物 质来 代替 化学合 成抗 生 素就显 得非 常迫 抗 寻 切 。抗 菌肽具 有 分子量 小且 合成迅 速 、 菌谱 广并 能与其 他抗 生素 协 同作用 、 很多抗 性 菌及 其 突变 株有 抗 对 特效且 不产生 选择 性 、 会导 致宿 主产生 脓毒 症等 优 点 , 不 因而 具 有很 好 的 药用 前 景[ 。因此 , 菌肽 的开 6 ] 抗 发和利 用为渔 业资 源保 护和水产 养殖 病害 防治 提供 了新 的选择 。
抗菌肽类和酶类酵母异源表达的研究

抗菌肽类和酶类酵母异源表达的研究抗菌肽类和酶类酵母异源表达的研究引言:抗菌肽是一类具有广谱抗菌活性的小分子肽类物质,这些肽在自然界中广泛存在于动植物和一些微生物中。
由于其天然来源、低毒性、高效率等特点,抗菌肽被广泛应用于食品、医药、农业等领域。
然而,抗菌肽的大规模生产受到了许多因素的限制,如成本、稳定性等。
因此,酵母异源表达系统成为抗菌肽及其他功能蛋白高效率制备的理想平台。
本文将介绍抗菌肽类和酶类酵母异源表达的研究现状和挑战。
一、抗菌肽的酵母异源表达1.1 酵母异源表达系统简介酵母是一类简单的真核生物,其异源表达系统具有优异的蛋白质表达性能和机制研究条件。
常用的酵母包括酿酒酵母(酮酸酸酯酵母),毕赤酵母等。
酿酒酵母是一种广泛应用于工业生产中的真菌,具有优秀的产酒性能和设备基础。
因此,酿酒酵母常被选为抗菌肽异源表达的宿主菌株。
1.2 抗菌肽的酵母异源表达策略抗菌肽异源表达的主要策略分为两类:短肽策略和全长策略。
短肽策略通过合成或克隆抗菌肽编码基因的短序列至酵母基因中,将其表达于酵母细胞。
全长策略则采用克隆含有完整抗菌肽基因的表达载体,将其转化至酵母中。
两种策略各有优缺点,在不同情况下可根据需要选择适宜的表达策略。
1.3 抗菌肽异源表达困难与解决抗菌肽的异源表达常常面临以下困难:剧毒性、导入异源抗菌肽基因时的细胞毒性及失活等。
为解决这些问题,研究者采用了许多策略,如信号肽重组、构建多基因拷贝操控系统、优化培养条件等。
这些改进方案显著提高了抗菌肽异源表达的成功率和产量。
二、酶类酵母异源表达的研究2.1 酶类在工业生产中的重要性酶类是一类在化学反应中起催化作用的生物催化剂,具有高效、特异性、低成本等特点。
在日常生活和工业生产中,酶类广泛应用于食品、制药、能源等领域。
因此,酵母异源表达酶类的研究具有重要的应用价值。
2.2 酵母异源表达酶类的策略酵母是一类优质的酶源,其异源表达系统具有高效稳定的表达性能。
常用的酵母异源表达构建策略包括直接表达、表面展示、分泌表达等。
重组猪源抗菌肽Cecropin P1在毕赤酵母中的分泌表达与活性研究

抗 菌肽 ( er pn 是 一类 被 称 为“ 二 防御 体 系 ” C co i) 第 的生 物 活性 小 肽 Ⅲ , 有 广 谱 、 】具 ] 高效 的 抑 菌 活性 , 能攻
ATC 5 2 标 准 株 , C2 9 2 自行保 存 ; ihap s rsX 3 、 P c i a t ‘ 一3 oi
0 5 甲醇 ,. Ag r d H2 . 15 a ,d O。 1 2 试 剂 . 限 制 性 内切 酶 Hi m、 o 、 a 、 v 、 分 子 na Nc S c A r1低 I I I
确 翻译 和 翻译 后加 工 , 而且 能分 泌许 多 蛋 白质产 物 , 使
生 物 素,
内二硫 键 , 有强 碱 性 的 N一 和 强 疏 水 性 的 C 端 , 一 端 _ c 端
酰胺 化 。其 分 子 内含有 两 亲性 一 旋 , 间 是形 成 柔 螺 中
tat 1 3 YNB, ×1 rc, . 4 4 0
生 物素 ,. 甘 油 , 0 05 10
mmo ・ 磷 酸钾缓 冲溶 液 ( H 值 6 0 。 l L p . ) B MMY 培 养 基 : P lp po e 1 Yes x 2 oy e tn , ate ~
收 集 菌体 , 上 清 , 菌 体 重 悬 于 B 弃 将 MMY 培养 基 中, 使 其 oD 。为 1 然后继 续在 3 。 , O℃摇 床 培养 , 每隔 2 4h
向B MMY培 养基 中加 入 甲醇至 终浓 度为 0 5 , 按 . 并
照 2 、6h 4 、 0h 7 、4 h 9 、 2 4h 3 、8h 6 、 2h 8 、 6h 1 0h时 间
游 引 物进行 鉴定 。 1 3 4 阳性 菌 株 的诱 导 表达 . .
毕赤酵母表达系统简介

巴斯德毕赤酵母及启动子1.1 毕赤酵母表达系统简介随着蛋白异源表达的飞速发展,越来越多的表达系统被建立并得到应用。
酵母作为单细胞真核生物,因具有比较完备的基因表达调控机制和对表达产物的加工修饰能力,仍表现出不可比拟的优势。
以甲醇营养型酵母(Methylotrophic yeast)-毕赤酵母为代表的第二代酵母表达系统,是近年来被公认的最有效的外源蛋白表达系统之一,已有多种外源蛋白在该宿主系统中获得了成功表达[1]。
作为生产外源蛋白的重要宿主菌,依靠其各种不同功能的表达载体,已经得到广泛的应用。
表达的蛋白质包括酶、膜蛋白、抗原、抗体和调节蛋白等[2,3]。
毕赤酵母(Pichia pastoris)表达系统是近年来发展迅速、应用广泛的一种真表达系统。
它是甲醇营养型酵母菌,有两个乙醇氧化酶(alcohol oxidase,Aox)码基因AOX1和AOX2,两者序列相似,AOX1基因严格受甲醇诱导和调控。
当甲醇为唯一碳源时,AOX1启动子可被甲醇诱导,启动乙醇氧化酶的表达,从而用甲醇进行代谢[4]。
含AOX1启动子的质粒可用来促进编码外源蛋白的目的因的表达。
随着Invitrogen公司开发的一系列毕赤酵母表达试剂盒的应用,目前用该统已成功表达出了数以千计的来自细菌、真菌、原生动物、植物、无脊椎动物、包括人在内的脊椎动物以及病毒等的具有生物学功能的外源蛋白或蛋白结构[5,6]。
1.1.1 P.Pastoris表达载体及其元件由于毕赤酵母没有稳定的附加质粒,表达载体需与宿主染色体发生同源重组,外源基因表达框架整合于染色体中以实现外源基因的表达整合表达的优点在于保持外源基因稳定性并可产生多拷贝基因。
典型的毕赤氏酵母表达载体含有醇氧化酶基因的调控序列,主要的结构包括:5′AOX1启动子片段、多克隆位点(MCS)、转录终止和polyA形成基因序列(TT)、筛选标记(His4或Zeocin)、3′AOX1基因片段,作为一个能在大肠杆菌中繁殖扩增的穿梭质粒,它还有部分pBR322质粒或COLE1序列。
抗菌肽DCD-1L的随机突变及在毕赤酵母中的表达

f. rd cQ ai n et nC n rf a s P oic ,L nh u7 0 3, h a 2 G nu dc, eerh 1P o ut u l s co e t o G nu rv e azo 3 00 C i ; . a s Me i R sac t p i yI e n n a
a dt ep oen e p e so a ea h e e es p r tn . n r ti x r si nc nb c iv d i t u enaa t h nh
Ke o d : n i a tra e td ; yW r s a tb ce il p i e DCD一 L; ie td e o u in; c apa t rs p 1 dr ce v l t o Pihi so i
705 3 0 0)
摘 要 :目的 用毕赤酵母构建抗菌肽DC 1 D.L的随机多肽库 ,并使随机多肽在 培养基中表达 。方法 用定向进化 的方法 在基 因内部造成随机突变,然后将其构建到含有强启动 子G 和Ⅱfc r AP . t 信号肽序列的巴斯德 毕赤酵母载体p P a ao GA Z C@, 线性化后转化到 巴斯德毕赤酵母蛋 白酶缺 陷株S MD16 中,形 成突变库 。利用抗 生素Z o i筛选出阳性克隆 ,并用S — 18 ecn DS P E AG 鉴定表达 蛋白质 。 结果 肽库库容达到7 ×1 f ,在上清中鉴定出了表达蛋 白。结论 成功构建了抗 菌肽DC . 0个eu 8 D-
we ei e td it ePih apa t rse p e so e trp r ns re ot c i so i x r sin v co GAPZa whc o t i dGAP p o o o n —a t rsg a n h C ihc n ane r m tra d f co i n l
抗菌肽的应用及存在的问题

天然抗菌肽由于分子量小, 直接从动植物组织中提取 时 , 分 离 提 纯 存 在 一 定 的 困 难 , 合 成 肽 价 格 昂 贵 。因 此 将 抗 菌肽基因利用基因工程转化进微生物表达系统或导入动 植物体内, 不仅可获得量丰的抗菌肽, 而且为改造物种获 得 优 良 抗 病 品 种 提 供 了 有 效 途 径 。以 下 分 别 从 动 物 和 植 物 两个方面总结抗菌肽在转基因方面的应用。 2.1 转抗菌肽基因动物
1985 年 , 由 广 州 星 群 制 药 厂 、广 东 省 农 科 院 及 华 南 农 业大学合作, 以五龄家蚕熟蚕为材料, 经热激诱导后制成 的干燥粉末, 装入胶囊作口服药物治疗肝炎。经广州市传 染 病 院 ( 52 例 ) 、暨 南 大 学 华 侨 医 院 ( 64 例 ) 、广 州 中 医 院 ( 20 例 ) 及 其 它 临 床 单 位 共 132 例 肝 炎 临 床 试 验 表 明 , 五 龄丸具有改善肝炎病人的蛋白代谢的功能[3] 。
抗菌肽成分为易消化吸收的氨基酸, 可作为畜禽饲料 添加剂取代或部分取代目前饲喂动物所用的抗生素, 减少 抗 生 素 对 动 物 体 的 危 害 。目 前 广 东 省 应 用 抗 菌 肽 基 因 转 化 酵母进行高效表达, 经发酵条件优化, 生产抗菌肽酵母制 剂, 用作畜禽及水产饲料添加剂, 代替抗生素预防及治疗 仔猪白痢和雏鸡白痢有明显效果。
WACHINGER 等 [ 13] 的 研 究 结 果 表 明 , 天 蚕 素 和 蜂 毒 素 可 以 在 亚 毒 性 浓 度 下 抑 制 艾 滋 病 毒 HIV- 1 的 基 因 表 达 。 从 而 减 少 HIV-1 的 增 殖 。 1.4 抗寄生虫作用
DIAZ-ACHIRICA 等 [14] 发 现 一 种 合 成 的 天 蚕 素- 蜂 毒 素 杂 合 体 对 莱 仕 曼 原 鞭 毛 虫 有 损 伤 作 用 。 SHAHABUDDIN[ 15] 研究发现昆虫抗菌肽对感染蚊子的疟原虫的卵囊期和 孢子期有明显的损伤, 而且该抗菌肽浓度对蚊子本身没
抗菌肽Cecropin AD在毕赤酵母中的组成型表达的研究

热和 酸碱稳 定 、 水溶性好 、 免疫 原性 和抗 菌谱广 等 无
特点 … 更 为重要 的是 , 菌肽 对正 常 的真 核细 胞几 , 抗 乎没有 作用 ,仅仅作 用 于原核 细胞 和发生 病 变 的真 核细胞 f 2 ] 。天蚕 素 (erpn 是抗菌 肽 的一种 , 3 ~ ccoi) 由 7 3 9个氨基酸组成 , 相对分子质量约 400k , B m n 0 u 由 o a
A H, S 电击 法转 化 Hci ps r 受 体菌 S 16 , ha at i os MD 8 成 1
功 表达抗 菌肽 C coi A 。 erpn D
1 材料 与方 法 11 材料 .
等 惜古 比天 蚕 H a p r ccoi 中分离 获得 , l 从 yl ha er a蛹 o p 是 目前研 究最 清楚 、效 果最 明显 的抗 菌肽 。天 蚕素 抗 菌肽 有广 谱 的抗 菌 能力 和 抗病 毒 活 性 。此 外 , 天 蚕素 还具有 无残 留毒性 及病 原体难 以产 生抗 药性 等
启 动子 的甲醇诱 导型 表达 , 它存 在诸 多不利 因素 , 如
收稿 日期 :0 9 0 一 5; 回 日期 :0 9 1- 3 2 0 — l0 修 2 0 — 1 1 基金项 目: 北京市重 大科 技计划课题 ( 0 0 0 0 2 0 0 ) D 9 1 14 7 0 0
保 存 ; . lD 5 购 自 TK R Ec i H a o a a a公 司 ; 表 达 载 体
P A Z — 购 于 Iv r e G P aA nio n公 司 。 tg
112 酶 和 试 剂 高 保 真 的 p r et N .. yo s D A聚合 酶 、 b 限 制性 内切酶 、,N TD A连 接酶 、 去磷 酸化 酶等 工具 酶
大黄鱼抗菌肽hepcidin在巴斯德毕赤酵母中的表达及其产物的抑菌活性

中图分类号 : 1 ' 7 3 5
文献标识码 : A
文章编号 : 2 0 9 5 4 9 7 2 ( 2 0 1 3 ) 0 3 - 0 3 8 3 - 0 7
列 . 已经报道, 真鲷( P a g r o s o m u s m a j o r ) 、 鲈 鱼
( L a t e o l a b r a x j a p o n i c u s ) 、 石斑鱼 ( E p i n e p h e l u s d r u m —
异 性 抗 细 菌 免 疫 的认 识 , 也 可 为 大 黄 鱼 病 害 防 治 提 供 新 的思路 . H e p c i d i n是 近年 来新 发现 的一 类具 有 独 特性 质
酸. 将L y e — H e p c 1 8 3 b p的编码 前导肽 和 成熟肽 的前体 肽 ( 2 5 a a 一 8 5 a a ) 基 因片段 克 隆 到毕 赤 酵母 表 达 载体 p P I C Z a A中, 然后 电 击转化 巴斯德 毕 赤 酵母 S MD 1 1 6 8 , 经 高浓度 Z e o c i n筛选获得 多个单克 隆.
第3 2卷
第 3期
应 用 海 洋 学 学 报
J o u r n a l o f Ap p l i e d Oc e a n O g r a p h y
Vo 1 . 3 2. No . 3 Aug ., 2 01 3
2 0 1 3年 8月
大黄鱼抗菌肽 h e p c i d i n在 巴斯 德 毕 赤 酵 母 中 的表 达及 其产 物 的抑 菌 活性
抗菌肽KS26基因的串联及在毕赤酵母中的表达_张元新

第31卷第3期吉林化工学院学报Vol.31No.32014年3月JOURNAL OF JILIN INSTITUTE OF CHEMICAL TECHNOLOGYMar.2014收稿日期:2014-01-09作者简介:张元新(1978-),男,辽宁建昌人,吉林化工学院讲师,博士,主要从事天然抗肿瘤药物作用机制方面的研究.*通信作者:葛雅琨,E-mail :yakunge@126.com文章编号:1007-2853(2014)03-0014-04抗菌肽KS26基因的串联及在毕赤酵母中的表达张元新,葛雅琨*(吉林化工学院化工与生物技术学院,吉林吉林132022)摘要:构建小肽基因的串联体,对其在真核系统中表达、纯化和性质研究具有重要的意义.本研究利用SOE-PCR的方法获取小分子多肽的全基因,利用同尾酶将基因串联,并将其插入到PICZ α真核表达载体中诱导表达.经酶切和DNA 序列分析表明,该串联体基因与所设计的基因序列完全一致;SDS-PAGE 的分析结果显示,该小肽基因在毕赤酵母中有较高水平的表达.关键词:抗菌肽;分子重组;串联;毕赤酵母;真核表达中图分类号:Q 812文献标志码:A自青霉素诞生以来,抗生素一直是治疗人类病原微生物感染疾病的重要武器;但是,随着抗生素的滥用及致病菌的多药耐药性的产生,抗生素逐渐失去往日的作用效果,所以寻找新型的抗菌资源已成为备受关注的焦点[1].阳离子抗菌肽是由12-50个氨基酸组成的阳离子型的两亲性分子.目前,已作为一种新型抗生素被广泛研究,它们存在于动物、植物、微生物中,是细菌、植物、低等动物的主要防御机制[2].在脊椎动物中,也能够运用特殊的免疫方式,作为一线防御物质,在病原微生物侵入早期抑制其生长[3].因此,阳离子抗菌肽因其具有的阳离子电荷和两亲构象使其具有良好的抗菌活性,备受科研工作者的重视[4].利用固相态化学合成小片段阳离子小分子多肽成本太高[5].因此,利用毕赤酵母系统进行体外重组表达的方法能够有效降低成本[6].该系统具有很多优点[7]:(1)利用受甲醇诱导的醇氧化酶(alcohol oxidase Ⅰ,AOX1)启动子,可严格控制外源基因的表达;(2)毕赤酵母生长快速,培养简单,适合高密度培养,发酵后每升培养液细胞湿重可达450克,有利于提高目的蛋白产量;(3)毕赤酵母作为一种真核细胞生物,可以进行蛋白翻译后的加工,以使外源蛋白蛋白得到正确的折叠盒修饰;(4)该系统已有20多年的研究开发历史,有多重受体菌和表达载体可供选择,可以进行胞内表达和分泌表达.因此,利用毕赤酵母真核表达系统进行小肽的重组表达,将有效降低生产成本,保证小肽的正确构象,提升其活性.本研究利用SOE-PCR方法快速、高效的获得了小分子多肽全基因,利用同尾酶得到基因串联体,并将其构建到毕赤酵母表在载体pPICZ αA 上,成功诱导出小肽的蛋白产物,为小分子多肽的真核表达研究奠定实验基础.1材料与方法1.1材料1.1.1质粒和菌种pPICZ αA 真核表达质粒、DH5α、GS115感受态细胞为吉林化工学院生物技术研究室保存.1.1.2主要试剂与仪器质粒小量提取试剂盒(BioTeke ),琼脂糖凝胶DNA 回收试剂盒(BioTeke ),限制性内切酶XhoI 、XbaI 、SalI 、Sac Ⅰ(Takara ),低分子量蛋白Marker (BioTeke ),T4DNA 连接酶(Takara ),Taq DNA polymerase (Fermentas ),Zeocin (Invivogen ),Tricine (Sigma ).PCR仪(Bio-Rad ),电子分析天平(Sartorius ),电泳仪和电泳转移槽(北京市六一仪器厂),紫外可见透射反射仪(上海精科实业有限公司),电热恒温培养箱(上海精宏实验设备有限公司),振荡培养箱(哈尔滨市东联电子技术开发有限公司),凝胶成像系统(Bio-Rad ).1.2方法1.2.1KS26基因的设计以毕赤酵母(Pichia pastoris )的偏爱密码子、多肽氨基酸序列为基础,设计基因全长78bp.SOE-PCR的引物均用Primer5辅助设计,克隆引物加入酶切位点XhoI、XbaI,在上下游引物的3,末端设计互补重叠序列,在反应体系中使两引物互为模板进行PCR.1.2.2KS26全基因的获取通过SOE-PCR的方法获取KS26全基因(78bp).PCR程序:94ħ预变性5min;94ħ变性30s,60ħ退火30s,72ħ延伸1min,30个循环后72ħ10min结束反应.反应体系:10ˑPCRbuffer5μl、MgCl23μl、dNTP1μl、引物F1μl、引物R1μl、Taq1μl、加ddH2O至总体积50μl.1.2.3KS26基因的串联以KS26基因为基础,Primer5辅助设计,分别在上游引物和下游引物加上同尾酶Xho I、Sal I,引物见表1.酶切连接反应的缓冲液为:Tris-HCl(Ph7.6)660mmol/L、MgCl266mmol/L、ATP 1mmol/L、DTT100mmol/L、NaCl1mol/L.反应体系:10ˑbuffer2μl,新鲜的PCR产物12μl,Xho I2μl,Sal I2μl,T4DNA Ligase2μl,总体积为20μl.酶切连接反应条件:37ħ酶切1h;22ħ连接1h,10个循环后结束反应.表1KS26基因串联引物*名称序列Forward Primer TCGACTCGAGATGAAGAAGGTTReverse Primer ACGCGTCGACGTCGACTCCATT *划线部分为酶切位点1.2.4重组载体PICZα-KS26的构建KS26基因的PCR产物和载体pPICZαA均用XhoI、XbaI双酶切,将胶回收后的双联体KS26 PCR产物与载体pPICZαA在T4DNA连接酶作用下于16ħ连接过夜.连接产物转化感受态E.coli DH5α,转化平板(含zeocin)37ħ过夜培养.挑取单一菌落于10μl无菌水中充分悬浮菌体,100ħ裂解5min,取1μl作为模板进行PCR验证.将验证阳性的单克隆接种于10 15mL LB 培养基(zeocin,终浓度为100μg/mL),37ħ,200rpm培养过夜,质粒小提后,进行酶切鉴定,鉴定正确的质粒交由上海生工生物工程有限公司测序部进行DNA测序.1.2.5pPICZα-KS26转化P.pastoris GS115(his-mut+)转化子的筛选和鉴定感受态P.pastoris GS115(his-mut+)(80μL)与SacⅠ线性化的pPICZα-小分子多肽(5μg)混合,转移至预冷的0.2cm电转杯中,置冰上5min,1.5kV、25μF、200Ω电击5ms,立即加入1mL预冷的1mol/l山梨醇,取出200μL涂布于MD板上,30ħ培养2d,通过比较转化子在MM 和MD板上的生长速度来筛选Mut+转化子.采用PCR方法分析P.pastoris转化子,挑取平板上的菌体,加入5μL无菌水中,煮沸5min,加入重新设计的鉴定引物F-tcgactcgagatgaagaag-gtttcc和R-ctgacttctagagtcgactccattg,反应体系同上,进行PCR反应.1.2.6重组子PICZα-KS26在P.pastoris菌在摇瓶中的诱导表达挑取重组P.pastoris,接种到BMGY中,30ħ、230rpm/min振荡培养约20h至OD600达到2 6;室温3000rmp/min离心5min,用BMMY (含1%Casaminoacids)重悬至OD600约为1.0,30ħ、250rpm/min震荡培养,进行甲醇诱导表达,每隔24hr取样,同时补加100%甲醇至终浓度为0.5%.1.2.7Tricine-SDS-PAGE取5μl表达上清液进行Tricine-SDS-PAGE,凝胶采用硝酸银染色,实验步骤参照Hermann的方法.2结果2.1KS26全基因的获取及串联体的构建为了能使KS26在毕赤酵母系统中高效表达,我们按照毕赤酵母基因翻译的偏爱密码子设计合成F、R两条互补引物,并使两引物3’段的11个碱基互补,见表2.根据重叠PCR的原理,获取了78bp大小的PCR产物,见图1A.表2KS26的氨基酸、基因和引物序列*类型序列KS26氨基酸序列KKVSFKVKFKSAWAKTVHTAKLASIS KS26基因aagaaggtttccttcaaggttaagttcaagtccgcttgggctaagactgttcatactgctaagctggcttccatttcc 引物F tcgactcgagatgaagaaggtttccttcaaggttaagttcaagtccgcttgggctaagactgttRctgacttctagagtcgactccattggaaatggaagccagcttagcagtatgaacagtcttag*单线划线部分为酶切位点;双线划线部分为两引物的互补序列.51第3期张元新,等:抗菌肽KS26基因的串联及在毕赤酵母中的表达根据KS26全基因序列,我们重新设计了具有同尾酶Xho I 、Sal I 的上游和下游引物,并以SOE-PCR产物为模板,通过在PCR中重复完成酶切-连接的程序,成功获得了具有Xho I 、Sal I 酶切位点的二联体、三联体产物,见图1B.图1SOE-PCR方法获取的KS26全基因及其串联体2.2重组载体pPICZ α-KS26的构建将KS26基因进行胶回收,其产物和载体pPICZ α均用XhoI 、XbaI 双酶切,用T4DNA Ligase 连接,转化感受态E.coli DH5α菌,筛选阳性克隆.重组质粒经双酶切鉴定并进行DNA 测序,结果表明目的基因正确插入,DNA 自动测序仪测序,序列如下:AAGAAGGTTTCCTTCAAGGTTA-AGTTCAAGTCCGCTTGGGCTAAGACTGTTCATA CTGCTAAGCTGGCTCCTATTTCC.2.3筛选、鉴定Mut +转化子为了获得更高的转化效率和高拷贝,采用电转化法将Sac Ⅰ线性化的重组表达质粒pPICZ α-KS26转化P.pastoris GS115(his-mut +),通过MM 和MD 板鉴定表型,大约80%的为Mut +;PCR扩增结果显示,阳性重组子出现了100bp 左右的插入目的片段,与预期相符,见图2.图2重组子的转化及PCR鉴定结果(DNA 标准同图1)2.4Tricine-SDS-PAGE抗菌肽KS26的分子量只有2.9kD ,在常规SDS-PAGE 系统中难以得到理想的分辨率,我们使用Tricine-SDS-PAGE (16%分离胶,10%浓缩胶,4%浓缩胶,使重组小分子多肽得到较好的分离.Tricine-SDS-PAGE 结果显示:甲醇诱导表达上清在2.9kD 处出现一条带,见图3.M.蛋白分子量标准.C.对照.1.重组蛋白KS26图3重组蛋白pPICZ α-KS26的Tricine-SDS-PAGE 电泳图3结论抗菌肽不仅具有杀菌还具有抗病毒和抗肿瘤细胞而不破坏人体正常细胞的作用,因此在农业、畜牧业、医药及食品工业中显示出越来越多的应用前景.阳离子抗菌肽水溶性好,且等电点在8.9 10.7之间,具有较强的阳离子特征,同时具有两亲α-螺旋和折β-叠片结构.阳离子抗菌肽具有61吉林化工学院学报2014年广谱的抗菌活性,来自昆虫、猪、蛙、人的阳离子抗菌肽既有抗革兰氏阳性和革兰氏阴性菌的作用,又有抗真菌、抗病毒、抗肿瘤的作用[7-9].通常研制和应用的抗菌肽均是从昆虫体液等天然物质中提取,来源途径广泛,但含量极低,提取步骤繁杂,很难获得大量高纯度的抗菌肽.目前阳离子抗菌肽的合成方法有化学合成和基因表达两种.化学合成得到的样品数量有限,而且差错率和副反应随着氨基酸数量的增多而增大,一般超过50个氨基酸的小肽很难合成.但阳离子抗菌肽结构简单、氨基酸残疾未被修饰,基因工程表达法是大量生产抗菌肽的理想方法[10].本研究利用SOE-PCR获取抗菌肽KS26的全基因及其串联体并构建了真核毕赤酵母表达系统,成功构建到进行了诱导表达,为快速、高效的进行小肽重组表达载体的构建、表达进行了有益的探索,有效地解决了小片段氨基酸合成成本过高的问题,将为短肽的规模化生产提供有效的实验依据.参考文献:[1]Bonomo RA.Multiple antibiotic-resistiant bacteria inlong-term care facilities :an emerging problem in the practice of in factious diseases [J ].Clin.Infect.Dis.2000,31(6):1414-1422.[2]Steiner H ,Hultmark D ,Engstrom A ,et al.Sequence and specificity of two antibacterial proteins involved ininsect immunity [J ].Nature 1981,292:246-248.[3]Hancock RE ,Chapple DS.Peptide antibiotics [J ].Antimicrob.Agents Chemother.1999,43:1317-1323.[4]Gottlieb C T ,Thomsen L E ,Ingmer H ,et al.Antimicrobialpeptides effectively kill a broad spectrum of Listeria monocytogenes and Staphylococcus aureus strains independently of origin ,sub-type ,or virulence factor expression [J ].BMC Microbiol.2008,8(1):205-214.[5]Hong In-Pyo ,Lee SJ ,Kim YS ,et al.Recombinantexpression of human cathelicidin (hCAP18/LL-37)in Pichia pastoris [J ].Biotechnol Lett.2007,29:73-78.[6]Jin FL ,Xu XX ,Yu XQ ,et al.Expression and charac-terization of antimicrobial peptide CecropinAD in the methylotrophic yeast Pichia pastoris [J ].Process Biochem.2009,44:11-16.[7]齐连全,陈薇,来大志,等.毕赤酵母表达系统的研究进展[J ].中国生物工程杂志2002,22(6):45-47.[8]Bellm L ,Lehrer RI ,Ganz T.Protegrins :new antibioticsof mammalian origin [J ].Expert Opin.Inv.Drug.2000,9(8):1731-1742.[9]Andra J ,Berninghausen O ,Leippe M.Cecropins ,antibacterial peptites from insects and mammals ,are potently fungicidal against candida albicans [J ].Med.Mcirobiol.Immunol.2001,189(3):169-173.[10]Hancock RE.Cantiontic peptites :effectors in innateimmunity and novel antimicrobibals [J ].Lancet Infect.Dis.2001,1(3):156-164.The Tandem of Antibacterial Peptide KS26Geneand Expression in Pichia PastorisZHANG Yuan-xin ,GE Ya-kun(College of Chemical and Biological technology ,Jilin Institute of Chemical Technology ,Jilin City 132022,China )Abstract :It is very important to tandem construction ,expression ,purification ,characteristic research of small peptide gene.In this study ,the whole-gene sequence of antibacterial peptide was achieved by means of SOE-PCR,constructed gene tandem through isocaudarner method ,and constructed recombinant eukaryotic expression vector.Enzyme digest and sequencing showed that gene tandem was equal to the sequence of the designed gene.SDS-PAGE analysis indicated that antibacterial peptide gene expressed highly in pichia pastoris.Key words :antibacterial peptide ;molecular recombination ;tandem ;Pichia pastoris ;Eukaryotic expression71第3期张元新,等:抗菌肽KS26基因的串联及在毕赤酵母中的表达。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Amp cl n iil e为 P o g i r me a公 司 产 品 ; e o i 为 I z r cn n
vto e i g n公 司 产 品 ; y tn 、 at xrc 、 i、 r Tr po e Ye s E ta tTr s 丙 烯 酰 胺 、 叉 双 丙 烯 酰 胺 为 Sg 甲 ima公 司 产 品 ; 其 余 常 用 试 剂 为 国产 分 析 纯 试 剂 . 1 2 方 法 .
ቤተ መጻሕፍቲ ባይዱ
限制 性 内 切 酶 、 4 T DNA 连 接 酶 、 g l I T 、 X.a 、 P G
关键 词 :抗 茵肽 ;毕 赤酵母 ; 选 表 中图 分类 号 : 96 Q 6 文献 标识码 : A
抗 菌 肽 具 有 高 效 广 谱 抗 菌 活 性 , 具 有 抗 肿 瘤 并 甚 至 抑 制 病 毒 复 制 作 用 J在 医 学 上 有 较 大 的 应 1 , 用 潜 力 , 此 国 内 外 在 抗 菌 肽 的 基 因 工 程 方 面 开 展 因 了不 步 研 究 工 作 . 关 抗 菌 肽 基 因 在 大 肠 杆 菌 、 酒 有 酿 酵 母 和 昆 虫 杆 状 病 毒 载 体 中 的 表 达 研 究 均 已 有 报
ATG G A A G TTG TTCA A A A AG 3
引 物 AD2: GCCGCCGAATTCTT 5 AGTT.
C TTA G CCA A G G CA G TA G CTTG A G C 3
引 物 3 5 GACTGGT : TCCAATTGACAAG.
C 3
摘
要 : 用 P R技 术改造 抗 茵肽 AD基 因, 其 C末端 改 造为 A h编码 , 采 C 将 s 改造 后 的 抗
茵肽 C co i erpnAD基 因克 隆到 p I Z P C mA栽体 上 , 建 成 酵母 重 组分 泌 型表 达 栽体 , 化 构 转 毕 赤酵母 ( i i p s r ) P c a at i 受体 茵 G 1 5中. 用 zrc h os S1 采 e i o n抗性标 记 筛选 重 组转化 子 , 经摇 瓶发酵, 农缩发 酵 液进行 酸性 聚 丙烯 酰胺凝胶 电泳 分析 和 抑 茵 活性检 测 结 果表 明 , 造 改 后 的 C co i D基 因在 毕赤酵母 中获得 了表 达 , erp nA 表达 产物 经  ̄F c r 号肽 引 导分 泌到 at 信 o 胞 外 , 有较 强的 杀 菌活性 . 具
维普资讯
华 南 理 工 大 学 学 报 (自 然 科 学 版 )
第 3 0卷 第 2期
20 0 2年 2月
J u n fS t o r alo ou h Chi n ve st f Te hn o y na U i r iy o c ol g
E.oi 2 1Pc [ p s r 受 体 菌 G l5 克 隆 载 clK1 ; i a at i D3 h os Sl ; 体 p E T es etr表 达 载 体 p I Z . G M- a vco ; y PC aA 11 2 酶和 试 剂 . Tq a DNA 聚 合 酶 、 NT 、 种 d P各
( trl ce c dt n Naua S in eE i o ) i
V l0 N . O_ O2 3
F bu r 2 0 e ray 02
文章编号 : 0 055 2 0 )20 1—4 1 0 .6 X(0 2 0 .0 30
抗 菌肽 A 基 因的改 造及 在 毕 赤酵 母 中 的表 达 D
其 中 引 物 AD1 引 物 AD 、 2用 于 扩 增 改 造 抗 苗 肽 AD 基 因 , 物 3 于 筛 选 重 组 转 化 子 和 测 序 引 用 ( ) P R 扩 增 以 含 C co i D 基 因 的 2 C erp A n P a . D 质 粒 为 模 板 , 引 物 A 、 物 A 2为 配 A 以 D1 引 D 对 引物 , 9 按 3℃ 、0s5 ℃ 、0s7 ℃ 、0 s进 行 3 3 ,2 6 。2 6 , 5 个 循环 , 行 至最 后 一步 时 , 7 ℃ 延 伸 反 应 1 i, 进 在 2 0m n 确保 P R 产 物 3 为 单 A 核 苷 酸 , 止 反应 . C 端 终 ( ) 因 克隆 3 基 在 TD 4 NA 连接 酶 作 用 下 . 上 述 将
1 2 1 Ce r p n AD 基 因 的 改 造 . . c o i
道 ~ j 目前 主要 存 在 的 问题 是 表 达效 率 较低 , 6, 分
离 纯 化 困 难 , 以 进 行 工 业 化 生 产 . 此 , 们 利 用 难 为 我 甲 醇 营 养 型 酵 母 Pc i p so i 效 表 达 系 统 , iha a tr s高 分 撼 表 达 了具 有 生 物 活 性 的 抗 菌 肽 AD 基 因 .
( )引 物 合 成 1
引 物 AD1 5 :
根 据 抗 菌 肽 基 因 改 造 和 克 隆 的
GCCGCCGAATTCATGAA.
需要 , 计 、 成 引物 序列如下 : 设 合
1 材料 和 方 法
1 1 材 料 .
11 1菌株 和质粒 . E.olDtS ; 菌肽敏 感菌 株 ci t a 抗
黄 亚 东 郑 青 李 校 茔 姚 汝 华 黄 自然
( 暨 南 太 学 医药 生 物 技 术 研 究 开 发 中 心 , 东 广 州 5 0 3 ; 1 广 16 0
2 .华 南 理 工 太学 食 品 与生 物 工 程 学 院 , 东 广 州 5 0 4 ; 广 1 6 0
3 华 南 农 业大 学 蚕 桑 系 ,广东 广 州 5 0 4 ) 162