哺乳动物细胞表达系统
cho细胞表达重组蛋白方案

CHO (Chinese Hamster Ovary) 细胞是常用的哺乳动物细胞系统,用于表达重组蛋白的研究和生产。
以下是一般性的CHO 细胞表达重组蛋白的方案:
1. 购买表达载体:选择适合的表达载体,可以是质粒或病毒载体。
载体应包含适当的启动子、选择标记等。
2. 转染CHO 细胞:将表达载体导入CHO 细胞中。
转染方法可以选择经典的化学或电穿孔法,也可以选择使用特定的转染试剂或转染仪器。
3. 选择稳定转染株:在转染后,使用适当的选择剂(如抗生素) 处理细胞,以选择稳定表达重组蛋白的细胞株。
可通过单克隆分离等方法筛选和扩增单一细胞克隆。
4. 细胞培养条件优化:优化培养基配方和细胞培养条件,包括温度、pH 值、培养基组分等,以提高重组蛋白的产量和纯度。
5. 表达蛋白的诱导:使用适当的诱导剂或方法,例如添加诱导剂(如甲酪酸) 到培养基中,以启动重组蛋白的表达。
6. 重组蛋白的纯化和分析:通过细胞破碎和不同的纯化步骤(如亲和层析、离子交换层析、凝胶过滤等)从培养基或细胞提取物中纯化目标重组蛋白,并使用适当的分析方法验证表达的蛋白的纯度和功能。
在每个步骤中,需要根据具体的重组蛋白和研究目的进行优化和调整。
此外,合理的培养细胞和操作操作也至关重要,以确保产量和纯度的理想达到。
这些方案的细节将根据具体的实验目的和需要进行个体化定制。
蛋白体外表达与纯化

蛋白体外表达与纯化随着后基因组时代的到来,蛋白质组成为科学研究的热点。
蛋白质作为生命机体的主要活动的承担者,其体外表达与纯化在研究相应基因的功能上有重要意义。
蛋白体外表达系统按其表达宿主可分为原核表达系统,真核表达系统和哺乳动物细胞表达系统。
一:原核表达系统原核表达系统的宿主菌主要以大肠杆菌为代表,大肠杆菌表达体系是目前应用最广泛的外源基因表达体系,这也是外源基因表达的首选体系。
该表达体系的优点:遗传学和生理学背景清楚;容易培养;外源基因经常可以高效表达及操作简单、周期短、成本低等。
其不足之处是不能进行典型真核细胞所具有的复杂的翻译后修饰;广泛的二硫键的形成及外源蛋白组装成蛋白复合体的能力也受到限制;另外外源基因产物在大肠杆菌中易形成不溶的包涵体;有时由于真核mRNA的结构特性及密码子使用频率与大肠杆菌的差异,而的不到足够的产物。
二:真核表达系统真核表达系统的宿主菌主要以酵母表达系统为代表,酵母基因表达系统的载体通常既能在酵母中进行复制也能在大肠杆菌中进行复制,形成所谓酵母菌――大肠杆菌穿梭载体。
因以大肠制备质粒DNA较方便,通常利用大肠杆菌系统构建酵母载体以简化手续,缩短时间。
作为基因表达系统的宿主应该具备以下条件:安全无毒,不致病;遗传背景较清楚,容易进行遗传操作;容易进行载体DNA的导入;培养条件简单;有良好的蛋白分泌能力;有类似高等真核生物的蛋白翻译后修饰功能。
三:哺乳动物细胞表达系统由于本专业不涉及哺乳动物细胞表达系统的应用,故此不赘述。
表达载体的种类及相应的分离纯化方法作为表达载体必须具备以下特征:稳定的遗传复制、传代能力,无选择压力下能存在于宿主细胞内;具有显性的筛选标记;启动子的转录是可调控的;启动子的转录的mRNA能够在适当的位置终止;具有外源基因插入的多克隆位点。
在原核表达系统中常用的表达载体有:PET-载体系列,用这类载体表达出的外源蛋白在N端或C端或两端均具有his tag。
哺乳动物表达系统原理

哺乳动物表达系统原理引言:哺乳动物表达系统是指哺乳动物通过语言、声音、姿势和化学信号等方式进行交流和传递信息的机制。
这种表达系统在哺乳动物种群中起着至关重要的作用,它有助于个体之间的合作、繁殖和求偶行为的进行。
本文将介绍哺乳动物表达系统的原理及其在不同物种中的表现。
一、语言交流:语言是人类最为重要的交流方式之一,但在其他哺乳动物中并不普遍存在。
然而,一些哺乳动物如海豚、鲸鱼和某些灵长类动物,通过发出特定的声音来传递信息。
这些声音可以包括呼吸声、鸣叫声、鸣叫声等。
它们通过调整声音的频率、音量和时长来传递不同的意义,从而实现交流。
这种语言交流对于维持个体之间的社交关系、警戒和求偶行为至关重要。
二、声音表达:除了语言交流外,声音也是哺乳动物表达系统中的重要组成部分。
许多哺乳动物通过发出特定的声音来传达不同的信息。
例如,大象通过发出嘶吼声、波动声和低音鼻音来进行交流。
这些声音可以传达警告、求偶和社交的信息。
类似地,狗通过吠叫声来表达不同的心情和需求,如警戒、寻求注意和表达快乐。
三、姿势和动作:除了声音外,姿势和动作也是哺乳动物表达系统中的重要组成部分。
例如,狗通过摇尾巴、耳朵的姿势和身体的动作来表达不同的情绪和意图。
摇尾巴可以表示高兴、友好或兴奋,而低垂的耳朵和身体的低姿态则可能表示害怕或不安。
类似地,猫通过抓、踩和搔抓等动作来表达不同的意思,如攻击、抚摸和警告。
四、化学信号:除了声音、姿势和动作,化学信号也是哺乳动物表达系统中的重要组成部分。
许多哺乳动物通过释放具有特定化学成分的信息素来传递信息。
例如,狗通过尿液中的信息素来标记自己的领地和状态,以及吸引异性。
类似地,许多灵长类动物通过分泌具有特定化学成分的汗液来进行社交和求偶行为。
五、不同物种中的表现:不同的哺乳动物在表达系统中的表现也存在差异。
例如,灵长类动物如黑猩猩和猴子可以通过复杂的声音、姿势和面部表情来进行交流。
而大象则以低音的鸣叫声和身体姿势来进行交流。
哺乳动物细胞表达系统
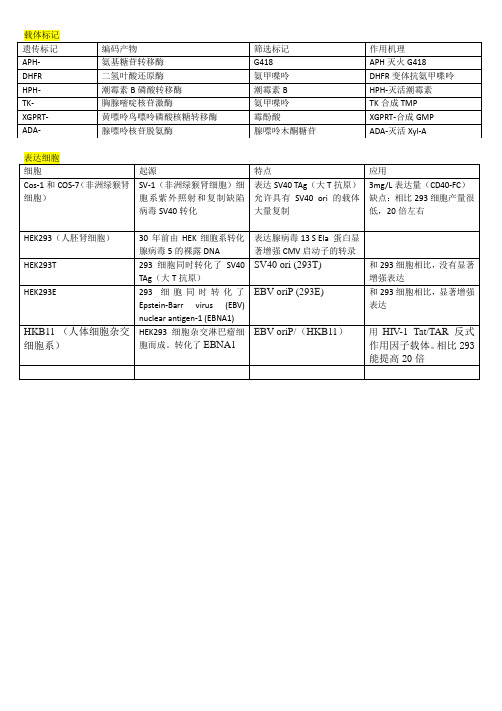
30年前由HEK细胞系转化腺病毒5的裸露DNA
表达腺病毒13 S Ela 蛋白显著增强CMV启动子的转录
HEK293T
293细胞同时转化了SV40 TAg(大T抗原)
SV40 ori (293T)
和293细胞相比,没有显著增强表达
HEK293E
293细胞同时转化了Epstein-Barr virus (EBV) nuclear antigen-1 (EBNA1)
EBV oriP(293E)
和293细胞相比,显著增强表达
HKB11(人体细胞杂交细胞系)
HEK293细胞杂交淋巴瘤细胞而成。转化了EBNA1
EBV oriP/(HKB11)
用HIV-1 Tat/TAR反式作用因子载体。相比293能提高20倍
遗传标记
编码产物
筛选标记
作用机理
APH-
氨基糖苷转移酶
G418
APH灭火G418
DHFR
二氢叶酸还原酶
氨甲喋呤
DHFR变体抗氨甲喋呤
HPH-
潮霉素B磷酸转移酶
潮霉素B
HPH-灭活潮霉素
TK-
胸腺嘧啶核苷激酶
氨甲喋呤
TK合成TMP
XGPRT-
黄嘌呤鸟嘌呤磷酸核糖转移酶
霉酚酸
XGPRT-合成GMP
ADA-
腺嘌呤核苷脱氨酶
腺嘌呤木酮糖苷
ADA-灭活Xyl-A
载体标记
表达细胞
细胞
Hale Waihona Puke 起源特点应用Cos-1和COS-7(非洲绿猴肾细胞)
SV-1(非洲绿猴肾细胞)细胞系紫外照射和复制缺陷病毒SV40转化
表达SV40 TAg(大T抗原)允许具有SV40 ori 的载体大量复制
哺乳动物细胞表达系统原理

哺乳动物细胞表达系统原理
哺乳动物细胞表达系统是一种用于生产重组蛋白和基因治疗的有效工具。
其原理主要基于哺乳动物细胞具有促使蛋白正确折叠和实现复杂修饰的功能,使得表达的蛋白更接近天然状态。
哺乳动物细胞表达系统有两种主要方式:瞬时转染表达和稳定转染表达(稳定细胞系构建)。
瞬时转染表达是指在短时间内表达出一定量的蛋白。
外源基因不整合到宿主染色体上,随着细胞的生长不断丢失,表达小量蛋白的过程。
这种方法简捷,实验周期短。
稳定转染/稳定细胞株筛选则是指将构建好的质粒线性化,在导入到培养好
的哺乳动物细胞内,通过一定的转染方法实现质粒与细胞的融合。
线性化的质粒进入到宿主细胞后,与细胞自身的基因组进行整合,同时随着细胞的生长繁殖而生长,过程中在经过一系列的筛选鉴定,排除未正确融合的重组子,或不能稳定表达下去的重组子,最终筛选出可以稳定表达的细胞株。
以上内容仅供参考,建议查阅哺乳动物细胞表达系统相关书籍获取更全面和准确的信息。
哺乳动物细胞表达系统

据文献报道,在不断提高的选择压力下,dhfr及侧翼序列 能扩增至上千个拷贝,大大增加目的基因的表达水平。
真核表达载体-启动子
外源基因在哺乳动物细胞中的表达与多种因素有关,主要 是启动子和增强子的强弱以及它们之间的搭配。
常用的高等哺乳动物受体细胞
迄今为止,用于医疗用品(药物、抗体、诊断试剂)大规 模生产的高等哺乳动物受体细胞主要还是中国仓鼠卵巢细 胞(CHO),其优势有如下几个方面: 遗传背景清楚,生理代谢稳定 与人的亲缘关系接近,外源蛋白修饰准确 基因转移和载体表达系统完善 耐受剪切力,便于大规模培养 被美国FDA确认为安全的基因工程受体细胞
Pei等用该细胞株表达分泌型的基质金属蛋白酶MMPI3,发现 高表达的阳性细胞克隆可占转染细胞的5%~l0%,其中一个 克隆的表达量可占细胞上清总蛋白的l5%~20%,在细胞单层 贴壁培养情况下表达量达10 mg/L。
对细胞株选择性地进行遗传改造
BHK/vl6细胞株是稳定表达单纯疱疹病毒(HSV)VP16蛋白 的BHK细胞,由于VP16的转录激活作用,载体中的HSV 早期启动子在该工程细胞中有很高的活性。
Clontech公司开发的Tet-off系统中的启动子则由CMV启动 子的核心序列和7个Tet阻遏蛋白结合位点组成。
这些启动子在诱导前后活性可相差4个数量级。
真核表达载体-启动子
在哺乳动物细胞中已发现存在大量在低氧环境中可诱导转录 的基因,如编码红细胞生成素(EPO)、转铁蛋白、血红素加 氧酶-1等的基因,它们都有一个共同的顺式作用元件 (CGTG ),有利于在5’或3’侧翼区的低氧诱导作用因子1(HIF-1)和低氧反应增强子(HRE)结合,激活靶基因的转录, 在低氧浓度下可使重组蛋白大量表达。
哺乳动物细胞表达系统的特点

哺乳动物细胞表达系统的特点
哺乳动物细胞表达系统是一种常用的重组蛋白表达系统,具有以下特点:
1. 表达的蛋白具有正确的翻译后修饰:哺乳动物细胞能够对表达的蛋白进行正确的翻译后修饰,如糖基化、磷酸化、乙酰化等,使表达的蛋白更接近天然蛋白的结构和功能。
2. 蛋白表达量较高:相对于其他表达系统,哺乳动物细胞表达系统能够产生较高水平的重组蛋白。
3. 适用于分泌型蛋白的表达:哺乳动物细胞具有完善的内质网和高尔基体等细胞器,可以将表达的蛋白分泌到细胞外,适用于分泌型蛋白的表达。
4. 产物易于纯化:哺乳动物细胞表达的重组蛋白通常具有较高的纯度,因为它们可以被分泌到细胞外,从而简化了纯化过程。
5. 适合治疗性蛋白的生产:由于哺乳动物细胞表达的蛋白具有与人体自身蛋白相似的结构和功能,因此适合用于生产治疗性蛋白,如单克隆抗体、细胞因子等。
不同蛋白质表达系统的比较

不同蛋白质表达系统的比较蛋白质是细胞内重要的生物大分子,可以发挥许多生命活动的关键作用。
不同的蛋白质表达系统可以用于生产不同类型的蛋白质,比较蛋白质表达系统的优缺点对于选择合适的表达系统具有重要的意义。
本文将介绍现有的几种主要的蛋白质表达系统,并对它们的特点进行比较。
第一种蛋白质表达系统是基于真核细胞的表达系统。
真核细胞是具有细胞核的细胞,其中包括了动物细胞、植物细胞和真菌细胞等。
这种表达系统利用真核细胞的转录和翻译机制来制造目标蛋白质。
在这种系统中,目标基因被插入到真核细胞的基因组中,然后通过RNA剪切和mRNA成熟等机制生成成熟mRNA,从而进行翻译,最终目标蛋白质被产生出来。
真核细胞表达系统的优点包括:能够产生生物活性和功能齐全的蛋白质。
这种系统还适用于产生大量的蛋白质,因此被广泛应用于产生多肽、抗体等药物。
但是,真核细胞表达系统的劣势在于工艺更加复杂,容易出现蛋白质不稳定、失去生物活性的问题。
此外,该工艺需要一定的时间来建立并优化系统。
第二个系统是基于细菌的表达系统。
细菌是单细胞生物,具有非常简单的结构和进化历史,是蛋白质表达方面的主要模型。
在这种表达系统中,表达载体中的目标基因被转化为蛋白质,并通过重组不同的DNA序列来实现该过程。
这种系统的优点在于简单、实时、具有可伸缩性和高效性。
制备蛋白质的成本也相对较低。
然而,这种表达系统的制约因素也很明显。
细菌系统不能表达合成二硫键、表达动力多肽等蛋白质的功能,使其应用领域相对狭窄。
此外,细胞酸碱值和产生蛋白质的环境等因素,都是影响蛋白质表达的关键因素。
第三种蛋白质表达方式是基于哺乳动物细胞的表达系统。
哺乳动物细胞表达系统主要用于大规模制备人用蛋白质。
这种表达系统利用哺乳动物细胞的转录和翻译机制来制造目标蛋白质。
在这种系统中,目标基因经过一系列的转染和筛选过程,被转移到哺乳动物细胞中,利用细胞所提供的设备进行蛋白质合成和修饰。
哺乳动物表达系统具有高表达量、具备更完整的蛋白质修饰等诸多优点。
整理了cho工艺培养中的常见问题

整理了CHO工艺培养中的常见问题一、介绍C H O细胞是重要的哺乳动物细胞表达系统之一,广泛应用于生物制药领域。
在CH O工艺的培养过程中,常常会遇到一些问题,本文将整理常见的问题,并提供解决方案,帮助研究人员更好地进行CH O工艺培养。
二、常见问题与解决方案2.1细胞生长问题2.1.1细胞生长速度过慢细胞生长速度过慢可能是由于以下原因导致的:1.营养物质不足:检查培养基成分,确保细胞所需的营养物质充足;2.麻烦问题:问题描述3.麻烦问题:问题描述4.麻烦问题:问题描述解决方案:1.检查培养基成分,确保细胞所需的营养物质充足;2.麻烦问题:解决方案3.麻烦问题:解决方案4.麻烦问题:解决方案2.1.2细胞过度生长细胞过度生长可能会导致浓度过高和滞后期延长。
为解决这一问题,可以考虑以下措施:1.改变培养基配方,减少细胞生长因子的浓度;2.麻烦问题:问题描述3.麻烦问题:问题描述4.麻烦问题:问题描述解决方案:1.改变培养基配方,减少细胞生长因子的浓度;2.麻烦问题:解决方案3.麻烦问题:解决方案4.麻烦问题:解决方案2.2细胞代谢问题2.2.1酸碱平衡问题细胞培养过程中,酸碱平衡是一个重要的参数。
酸碱平衡问题可能会导致细胞代谢异常,影响细胞生长。
以下是常见的酸碱平衡问题及解决方案:1.培养基p H过高或过低:调整培养基pH,使用缓冲液进行调节;2.麻烦问题:问题描述3.麻烦问题:问题描述4.麻烦问题:问题描述解决方案:1.调整培养基p H,使用缓冲液进行调节;2.麻烦问题:解决方案3.麻烦问题:解决方案4.麻烦问题:解决方案2.2.2能量代谢问题细胞能量代谢异常会影响细胞生长和产物表达。
以下是常见的能量代谢问题及解决方案:1.缺乏能量源:确保培养基中含有合适的能量源,如葡萄糖;2.麻烦问题:问题描述3.麻烦问题:问题描述4.麻烦问题:问题描述解决方案:1.确保培养基中含有合适的能量源,如葡萄糖;2.麻烦问题:解决方案3.麻烦问题:解决方案4.麻烦问题:解决方案三、结论通过对C HO工艺培养的常见问题的整理,可以帮助研究人员更好地解决培养过程中遇到的困难。
蛋白质表达与纯化技术的研究进展

蛋白质表达与纯化技术的研究进展随着生物技术的发展,蛋白质表达与纯化技术也得到了迅速的发展。
蛋白质是生命物质中至关重要的组成部分,为研究生命的机制及开发生物制药提供了重要的基础和前提。
本文将从蛋白质表达及纯化技术的研究进展入手,介绍相关的前沿技术和方法。
一、蛋白质表达技术的研究进展1.1 原核表达系统原核表达系统是一种常用的蛋白质表达技术,它利用细菌的生物学特性,在大规模表达目标蛋白质的同时,具有快速、高效、经济的优势。
近年来,原核表达系统也得到了不断的改良和优化,例如利用基因工程技术将目标蛋白质表达的速度和表达量得到了显著提高,进一步拓宽了其应用范围。
1.2 酵母表达系统酵母表达系统主要利用酵母菌作为载体表达目标蛋白质,具有高表达量、合成质量好、能够进行翻译后修饰等优点。
在酵母表达系统中,利用选择性培养基的筛选方法可以显著提高目标蛋白质表达的效率和纯度。
1.3 昆虫细胞表达系统昆虫细胞表达系统是一种常用的哺乳动物细胞表达系统,利用昆虫细胞(如Sf9、Sf21细胞等)表达目标蛋白质。
这种系统具有易于维护,表达效率高,重组蛋白质具有天然的哺乳动物的修饰等优点。
目前,昆虫细胞表达系统已经被广泛应用于疫苗、生物药物等领域。
1.4 哺乳动物细胞表达系统哺乳动物细胞表达系统是目前最常用的蛋白质表达技术,通过利用哺乳动物细胞表达目标蛋白质并进行不同程度的修饰,可以得到与天然蛋白质相似的重组蛋白质。
此外,该系统还可应用于细胞培养技术、生物药物研发等领域。
二、蛋白质纯化技术的研究进展2.1 柱层析技术柱层析技术作为蛋白质纯化的核心技术,是一种能根据其化学性质和物理性质特征,利用不同的色谱柱实现组分分离的技术。
随着柱层析技术的发展,液相色谱、气相色谱、毛细管电泳等技术的出现,蛋白质的纯化程度得到了进一步提高。
2.2 薄层凝胶电泳技术薄层凝胶电泳技术是一种以物质的分子量为分离基础,利用电泳原理实现生物大分子分离的技术。
哺乳动物细胞表达系统

哺乳动物细胞表达系统按照宿主细胞的类型,可将基因表达系统大致分为原核、酵母、植物、昆虫和哺乳动物细胞表达系统。
与其它系统相比,哺乳动物细胞表达系统的优势在于能够指导蛋白质的正确折叠,提供复杂的N型糖基化和准确的O型糖基化等多种翻译后加工功能,因而表达产物在分子结构、理化特性和生物学功能方面最接近于天然的高等生物蛋白质分子。
从最开始以裸露DNA直接转染哺乳动物细胞至今的30余年间,哺乳动物细胞表达系统不仅已成为多种基因工程药物的生产平台,在新基因的发现、蛋白质的结构和功能研究中亦起了极为重要的作用。
本文主要从表达系统及其两个组成部分一一表达载体和宿主细胞等方面,简要介绍哺乳动物细胞表达系统和相关的研究进展。
研究现状①部分蛋白在哺乳动物细胞中的表达已从实验室研究迈向生产或中试生产阶段。
②已有许多重要的蛋白及糖蛋白利用哺乳动物细胞系统表达和大量制备、生产。
如人组织型血纤蛋白酶原激活因子、凝血因子皿、干扰素、乙肝表面抗原、红血球生成激素、人生长激素、人抗凝血素出,集落刺激因子等。
有些产品已投入临床应用或试用。
③虽然经过多年努力,哺乳动物细胞表达系统的表达水平有大幅度增高,但从整个水平上看仍偏低,一般处在杂交瘤细胞单克隆抗体蛋白产率的下限,即1-30^g/l08细胞/24小时。
有人认为其限速步骤可嚣是在工程细胞中(对于重组蛋白来讲,常是异源的),重组蛋白的分泌效率较低。
1表达载体1. 1表达栽体的类型哺乳动物细胞表达外源重组蛋白可利用质粒转染和病毒载体的感染。
利用质粒转染获得稳定的转染细胞需几周甚至几个月时间,而利用病毒表达系统则可快速感染细胞,在几天内使外源基因整合到病毒载体中,尤其适用于从大量表达产物中检测出目的蛋白。
根据进入宿主细胞的方式,可将表达载体分为病毒载体与质粒载体。
病毒载体是以病毒颗粒的方式,通过病毒包膜蛋白与宿主细胞膜的相互作用使外源基因进入到细胞内。
常用的病毒载体有腺病毒、腺相关病毒、逆转录病毒、semliki森林病毒(sFv)载体等。
哺乳动物细胞表达系统

转录终止序列
真核基因的hnRNA的加工过程需要PolyA信号,在目的基因 3’端加上PolyA,表达水平提高10倍以上。 SV40的早期和晚期PolyA. 牛生长激素基因PolyA. 人工合成PolyA.
IRES
IRES(internal Ribosome entry site)内部核糖体切入 位点,可从一个mRNA翻译二个ORF。
高等哺乳动物受体细胞的遗传标记
遗传标记 编码产物
筛选药物
作用机理
APH-
氨基糖苷磷酸转移酶 G418
APH灭活G418
DHFR HPHTK-
二氢叶酸还原酶 潮霉素B磷酸转移酶 胸腺嘧啶核苷激酶
氨甲喋呤 潮霉素B 氨甲喋呤
DHFR变体抗氨 甲碟呤
HPH灭活潮霉素 B
TK合成TMP
XGPRTADA-
黄嘌呤过控制 tet 的浓度来控制基因表达的水平。
▪ 应用于研究基因不同水平的表达对细胞,个体发育的影响
有何不同。
作用机制示意图
四环素调控表达系统构成元件
▪ 四环素阻遏子(Tet repressor protein,TetR) ▪ 四环素操纵子(Tet operator DNA Sequence, TetO) ▪ 单纯疱疹病毒Vp16转录活化区(Vp16 activation domain,AD) ▪ 四环素调控反式激活子(Tetracyeline-Controlled
transactivator .tTA)TetR与Vp16组成融合表达蛋白。
哺乳动物细胞表达系统
动物基因工程原理
特点与优势
哺乳动物细胞表达系统的优势在于能够指导蛋白 质的正确折叠,提供复杂的N型糖基化和准确的O型 糖基化等多种翻译后加工功能,因而表达产物在分 子结构、理化特性和生物学功能方面最接近于天然 的高等生物蛋白质分子。
哺乳动物细胞高效表达系统研究进展

第34卷总第89期2013年3月西北民族大学学报(自然科学版)J ournal of N or t hw es t U ni ve r si t y f or N at i o nal i t i es(N at ur al Sci enc e)V oI.34.N o.1M ar ch,2013哺乳动物细胞高效表达系统研究进展柏家林1,2(1.甘肃省动物细胞工程技术研究中心,甘肃兰州730030;2.西北民族大学生命科学与工程学院,甘肃兰州730030)[摘要]哺乳动物细胞高效表达系统是生物制药工业中十分关键的环节,而表达载体和宿主细胞又是哺乳动物细胞表达系统的两个重要组成部分.文章较详细综述了通过目的基因整合位点的优化、增加目的基因拷贝数、提高目的基因的转录和翻译水平等构建哺乳动物细胞高效表达载体的研究进展.同时。
为使宿主细胞更好地适应工业化大规模培养要求,对通过抗凋亡、细胞周期调控、糖基化、细胞增殖控制技术和多基因代谢等细胞代谢工程对其进行改造的研究进展也作了介绍.[关键词]哺乳动物细胞;高效表达载体;细胞代谢工程;生物工程制药[中图分类号】Q811.4[文献标识码]A[文章编号]1009—2102{2013)01—0043—09利用哺乳动物表达系统生产药用蛋白是21世纪生物制药工业的主要发展方向.用于制备基因工程药用蛋白的表达系统有原核和真核表达系统.原核表达系统主要是大肠杆菌表达系统,它具有表达水平高、操作简单、周期短、易于大规模高密度发酵生产、成本低等优点,成为表达产物不需要修饰的小分子蛋白类药物的首选表达系统.而对于糖蛋白和全抗体类生物药物来说,表达产物多肽链的折叠、二硫键的形成、糖基化的有无以及糖基化的类型常常影响表达产物的合成分泌、生物活性、药代动力学行为、体内稳定性以及免疫原性等特性.由于原核细胞缺少内质网和高尔基体等进行糖基化的细胞器,所表达的产物是非糖基化的,且常以包涵体的形式存在.而酵母、昆虫及植物等真核细胞虽能进行糖基化,但糖基化的方式与人类等哺乳动物细胞又不一样,表达产物寡糖链末端多为甘露糖(m ar i nose,M an)、N一羟乙酰神经氨酸(N—gl ycol yneur am i ni c aci d,G l cN A c)和半乳糖(gal act os e,G al),它们均容易被肝细胞、巨噬细胞表面的受体识别而清除….特别是G l cN A c寡糖链,除在人的胚胎期体内存在外,成人一般都不表达,因此对人可能有免疫原性.目的基因在哺乳动物细胞中的表达产物不但能正确组装成多亚基蛋白,而且与天然蛋白的结构、糖基化的类型和方式几乎一致.常用于药用蛋白生产的哺乳动物细胞有3T3、C H O、B H K、H e l a和H epG2,约70%药用蛋白用C H O细胞生产【2J,而且C H O细胞能在10000L以上生物反应器中高密度、无血清悬浮培养【3,4J.经过近30年的努力,已有报遭单个细胞每天表达蛋白量达20Pg的工程细胞株【5】,分批补料式生物反应器悬浮培养细胞密度达2000万细胞/m L 以上,蛋白产量高达10g/L[3,6,7】.但与大肠杆菌相比,哺乳动物细胞的表达水平仍较低、获得高表达工程细胞株所需的时间长、细胞大规模培养的成本高等导致哺乳动物细胞生产蛋白质类药物的成本较高.因此,建立哺乳动物细胞高效表达体系包括高表达载体构建、高表达工程细胞株的获得、生物反应器无[收稿日期】2012—12—20【基金项目]中央高校基本科研业务费专项资金项目(zyz2011074),校企合作项目(h2011—19).[作者简介]柏家林(1966一),男,甘肃天水人,理学博士,博士后,教授,主要从事功能基因组学、动物分子育种及生物技术相关面的研究.一43—血清高密度培养工艺以及目标蛋白的分离纯化工艺,是生物制药工业研究和生产的关键技术.本文就哺乳动物细胞高效表达载体的构建及宿主细胞改造的研究进展作一介绍.1哺乳动物细胞高效表达载体的构建目的基因在哺乳动物细胞中的表达受目的基因所整合染色体区域的状态、目的基因的拷贝数及目的基因的转录、翻译和翻译后加工修饰效率的影响.构建一个高效表达的哺乳动物细胞表达载体,应从表达载体在染色体上整合位点的优化、转录与翻译效率的提高以及目的基因拷贝数的增加等方面综合考虑.1.1整合位点的优化目的基因在C H O细胞染色体上整合位点的状态对于目的基因的表达与否、表达高低以及目的基因在宿主细胞中的稳定性起着决定性作用.只有那些整合位点处于染色体转录活跃区的细胞形成的克隆才可高水平表达目的基因.因此,保证将表达载体整合在C H O细胞染色体上转录活跃位点的细胞克隆挑选出来是提高C H O细胞表达水平的关键步骤之一.1.1.1利用选择基因的弱化表达提高转基因表达水平选择基因(如neo、dhf r)的弱化表达,可使大量整合在低表达位点的细胞由于选择标记基因表达量不足而在选择培养基条件下中毒死亡,只有那些少量整合在转录活跃区的细胞由于表达足够的选择基因产物而存活下来形成克隆.通过在转录水平、翻译水平和构建活性降低的选择基因突变体可实现选择基因的弱化,如将选择基因置于缺失增强子的SV40启动子控制下或内含子中、改变K ozak序列、在选择基因起始密码子A TG前加不同读框的A T G等.B r i an将(1996)选择基因dhf r置于目的基因前一个人工内含子中构建了双顺反子表达载体C M V—D I—t PA,通过筛选表达dhf r基因的克隆,将目的基因的表达量提高了11倍【8].W em er等(1998)在neO的内部插入一个人工内含子,使用这种载体筛选到的阳性克隆经基因扩增后C D20单克隆抗体的表达量高达2r ag/m L[91.1.1.2利用核基质附着区提高转基因表达水平在载体上添加染色体上的某些特定序列如核骨架附着区(scaf f ol d a t t ac hm e nt r egi on,SA R s)或核基质附着区(m at r i x at t achm ent r egi on,M A Rs),可使表达载体整合到宿主细胞染色体后能模拟染色体的高转录活跃区,从而使形成的阳性克隆较均一地高效表达目的基因.K odur i(2001)用反义遗传学方法克隆了表达载体在高表达C H O细胞株染色体上整合位点两侧的侧翼序列H I R PE(hot sp ot f or i nc r eased r ecom bi nant pr ot ei n expr essi on),发现这些序列富含转录活跃区的类A l u序列、基质附着区等元件.他们用H I R PE构建了pTV l载体,使C TLA4~I g在C H O细胞中的基础表达水平提高了10倍【10|.E m er y (1998)用由p一球蛋白基因的位点控制区(10ci cont r ol r egi on,LC R)的超敏感位点(hyper s ens i t i ve s it es)第2、3和4的核心元件构成了一段序列,它能使鼠红白血病细胞(m ur i ne er yt hr ol e ukem i a,M E L)的克隆形成率提高94倍,且能明显提高a一球蛋白的表达量【11].K i m等(2004)用人J3一球蛋白基因M A R s 构建的载体转染细胞后的表达水平比对照组提高了7倍,并且在每次用叶酸类似物氨甲喋呤(am et hopt er i n,M T X)加压后不用挑选单克隆,大大缩短了获得高表达细胞株的周期[12|.1.1.3利用位点特异性重组提高目的基因表达水平一种方法是通过位点特异性重组将目的基因整合到细胞基因组中的转录活跃区,即先将含有定点重组位点的选择标记基因整合到染色体高表达区,然后将表达目的基因的表达载体和表达重组酶载体共转染上述带有重组位点的细胞系,在重组酶介导下,表达载体通过位点特异性重组定点整合在染色体高表达区,如C r e—k】(P和Fl p—Fr t位点特异性重组系统.K i t o等用带有dhf r扩增基因及融合了Lo】【P 位点的绿色荧光蛋白融合基因的质粒转染C H O—dhf r一细胞株,经过筛选并用M TX加压扩增后得到了可进行基因扩增的定点整合C H O细胞系【131.I nvi t r ogen公司则用Fl p—Fr t位点特异性重组系统建成了多种商品化的定点整合系统,包括293细胞、B H K细胞和正常C H O细胞.但这些细胞的整合位点没有经过系统的优化筛选,而且不能很好地与常用的加压系统(如M TX加压扩增系统等)配合,定点整合一44—的外源基因的表达水平通常都较低.刘志刚等(2004)借助于位点特异性重组和高效筛选,实现了FRT 位点在C H O—dhf r一细胞基因组中转录活跃区的整合,单链抗体一尿激酶融合基因表达量达5t,g/(106细胞24h)¨4|.另一种方法是利用体细胞同源重组即基因打靶技术,实现目的基因定点整合在宿主细胞染色体转录活跃区,但由于体细胞基因打靶比较困难,目前只有少量成功报道【l0I.目前关于真核细胞在染色体水平上调控基因表达的机制还不完全明了,染色体高转录活跃区的鉴定及分离费时费力而且不能保证所获得的就是转录的最活跃区.相比较而言,通过选择基因的弱化,利用位置效应来提高转录效率从而达到目的基因的高表达是一种更为有效的方法.1.2增加目的基因拷贝数单拷贝或低拷贝目的基因,无论表达载体调控元件如何优化、整合的染色体位点多么合适,其外源基因表达量都是有限的.因此,通过增加目的基因拷贝数来获得高表达重组药物的C H O工程细胞株是基因工程药物研究中不可或缺的重要环节.目的基因的扩增常采用目的基因和选择标记基因共扩增的方法,如二氢叶酸还原酶(dhfr)和/或谷氨酰胺合成酶(G S)是常用的扩增基因[15,16].C H O—dhf r扩增系统常采用dhf r基因缺陷的细胞株(C H0一dhf r一),可使目的基因的拷贝数扩增至1000余倍.W em er 等(1998)在nEo基因的内部人工内含子中放置目的基因和dhf r基因偶连的表达单元,通过用M TX对阳性克隆dhf r扩增,C D20单克隆抗体的表达量高达2g/L[9|.与dhf r扩增基因相比,G s扩增基因是一种显性基因扩增选择标志,应用G S基因扩增系统常只需l~2轮蛋氨酸黄酰胺(m et hi oni ne sul phoxi m i ne,M SX)加压筛选即可获得高表达细胞株,扩增效率较高,并且在含有G S基因的细胞株中也可得到有效的基因扩增.在C H O细胞中,G S经1轮扩增,基质金属蛋白酶组织抑制因子(t i s sue i nhi bi t o r of m et al l opr ot ei nas es,TI M P)的表达量能从每106细胞9弘g/d提高至110扯g/dt22|.B ebbi ngt on 等用1株以G S作为扩增基因的鼠骨髓瘤细胞(N So)表达人一鼠嵌合抗体,经l轮扩增后,其表达水平为每106细胞10~15gg/d[17I.1.3转录水平在目的基因拷贝数一定、整合位点固定的情况下,转录作为基因表达的第一步,提高转录效率对一个高效表达载体的构建来说显得尤为重要.启动子及其相应增强子、转录终止信号及多聚腺苷酸加尾信号对转录水平的高低及m R N A的稳定性有很大影响,其中强启动子、强增强子是提高转录水平的关键因素.因此人们希望通过寻找转录起始效率高、适用范围广的启动子、增强子来提高目的基因转录水平的表达效率.目前常用病毒源性和细胞源性的强启动子,如m C M V、hC M V、hE Fl a、人C—f os、鸡胞浆8一肌动蛋白等启动子.Bi等发现在含有人C—l os启动子和绿色荧光蛋白(G FP)的报告基因系统中,C —f os启动子控制下G F P的平均表达量比C M V启动子下的更高[18].研究表明C M V启动子在细胞处于S期、细胞生长迅速时,转录活性最高.但在大规模生产的中、后期,多数细胞生长不旺盛时,可显著影响外源基因的表达水平.与之相比,hE Fl a启动子的转录起始效率更强,并且其转录活性不受细胞周期影响,更适合大规模生产重组蛋白.除寻找强启动子、强增强子之外,用含有不同启动子、增强子的组成元件构建转录效率更高的杂合启动子或杂合增强子也不失为提高转录效率的一个好方法.M a sayuki 等(1997)发现C M V增强子能提高hEF—l a启动子控制下目的基因表达量4~9倍[19].K i m等发现将hE Fl a启动子的第1个内含子置于m C M V启动子与荧光素酶之间能极大地提高目的基因在C H O细胞中的表达,使荧光素酶的表达量提高了8.2倍【20I.xu等(2001)对CM V和B一肌动蛋白的启动子和增强子、内含子及SV40、B G H和m R B G的pol y(A)等不同转录调控元件的多种组合在H eL a、H epG2和E C V304细胞中进行了系统比较后发现,m R B G的pol y(A)比SV40和B G H的pol y(A)的适用范围更广,能更有效地提高目的基因的表达【21J.在转录过程中,转录因子通过D N A结合结构域特异识别并结合目标基因的特异调控序列,通过转录激活结构域调节或将转录作用因子募集至启动子从而启始基因的转录和表达.因此,提高宿主细胞转录因子的表达水平也能增强目的基因的表达.C ocket t(1990)将反式作用于C M V启动子的腺病毒E I A蛋白基因整合到C H O细胞中,通过提高C H O细胞E I A的表达水平,使C M V启动子控制下的一45—TI M P基因表达水平提高了10倍[22|.另有研究表明,EG F和H RG一91(H e r egul i n be t a1)能提高E FI a m R N A的转录量和蛋白质的表达[23|.R eza等(2002)将用作结合D N A的锌指结构和V Pt6蛋白的转录激活结构域融合构建了人工转录激活因子.通过表达人工转录激活因子,将C M V启动子的转录效率提高了2倍多[24|.随着真核细胞在转录水平调控机制的进一步研究,杂合或人工启动子以及人工转录激活因子的构建将是更为有效的提高转录效率的方法.1.4翻译水平除了转录水平的调控外,翻译水平的调控(如m R N A寿命、m R N A的翻译起始效率)和翻译产物加工修饰的效率等也对目的基因的表达产生重要影响.pol y(A)的存在不但能影响m R N A稳定性,而且也能部分起“翻译增强子”的作用,提高m R N A翻译水平.内部核糖体进入位点(i nt er nal r i bosom e ent r y si t e,I R E S)能使同一m R N A中除第1个基因之外的其他基因得到有效表达.翻译增强子可提高翻译效率;通过使用宿主细胞偏爱的密码子来对目的基因的密码子进行优化也可以大幅度提高翻译效率.Jacks on首先从细小R N A病毒中发现了I R ES.I R ES下游的开放阅读框采取不依赖帽子结构的方式,直接起始翻译.同时,I RES有效介导的内部起始翻译要求起始密码子处有一个有利的翻译环境,当第一个A U G周围序列不利于翻译起始时,I R ES可增强下游A U G的翻译起始[25].如在V EG F的5’端非翻译区包含1个有功能的I R ES,当eI F4复合物成为限制因素时,I R ES介导的内部起始翻译与其他m R N A竞争时存在优势,它能在依赖帽子结构翻译受损时提供一种有翻译能力的m R N A[26].此后,Paul ous等(2003)依据IRE S的初级序列和二级结构的保守性以及在离体情况下它们对内部起始翻译要求的条件,将小核糖核酸病毒的IRES分为3类:肠遭病毒(ent erovi r us)和鼻病毒(rhi novi rus)的I R ES,它们在兔网状细胞裂解液(r abbi t r et i cul ar ce l l l ysa t e,R R L)中缺乏特殊的细胞蛋白时不能有效起始翻译;脑心肌炎病毒(EM C V)和口蹄疫病毒(FM D V)的I R ES在不补充R R L时也能有效起始翻译.盐浓度的波动和2A或LB蛋白酶介导的el F4G切割对这类IR ES介导的起始翻译的影响不显著.甲肝病毒的I RE S,它在RR L中高度有效,但在R RL中添加细胞的粗提物时不能刺激它,在eI F4G受2A 或L B蛋白酶切割时受抑制[271.由于第2类IR ES具有对细胞微环境的广泛适应性和内部起始翻译效率高的特点,它已经广泛应用于双顺反子或多顺反子的哺乳动物细胞表达载体的构建.K auf m an等(1991)通过在双顺反子内加入EM C V的内部核糖体进入位点实现了提高目的基因在双顺反子表达载体中的表达效率‘28f.翻译增强子是具有与IR ES不同二级结构的另一类提高翻译起始效率的顺式调控元件.St ei n等(1998)发现在组织缺氧情况下,V E G F m R N A寿命大为提高,V EG F翻译增强的程度可高达40倍,由此发现了具有翻译增强子和IR ES双重功能的V EG F163片断[26].C hr i st i an在纤维细胞原生长因子2 (F G F一2)的3’端非翻译区发现了2个能使C A T的翻译效率提高9倍的独立元件TE l和TE2,它们之间具有累积效应,能与FG F一2m RN A的5’端非翻译区协同作用,增强翻译效率达12倍[29].V i vi nus (2001)在H sp70m R N A的5’端非翻译区发现了一个能作为m R N A通用翻译增强子的元件.H sp70 m R N A的翻译增强子没有I R ES结构,它在C A T编码区上游能提高C A T的表达效率10倍,并且能在依赖于帽子结构的翻译过程中起到增强翻译效率的作用【30】.用宿主细胞偏爱的密码子来对目的基因密码进行优化是另一种提高翻译效率的方法.K i m等用人高表达基因偏爱的密码子系统地设计了人EPO基因,再以酵母偏爱的密码子编码人E PO基因作对照来比较优化,结果人优化E PO基因密码子比非优化人E PO基因密码子的表达效率高2~3倍[31].2宿主细胞的改造随着对细胞代谢途径和调控机理的深入了解,越来越多的研究集中在对细胞本身进行改造的代谢工程来达到优化细胞生长状态,提高产品产量和质量,延长生产周期的目的.目前哺乳动物细胞的代谢工程包括抗凋亡工程、细胞周期调控工程、糖基化工程、细胞增殖控制技术和多基因代谢工程等.2.1抗凋亡工程——46——细胞在大规模培养初期,目的蛋白的表达与细胞的增殖速率呈正相关.但当反应器中细胞的密度达到饱和后,细胞继续增殖会导致养分和氧的大量消耗以及乳酸、氨等有毒代谢产物的大量积累,细胞逐渐凋亡(apopt osi s),重组蛋白表达量逐渐降低.细胞凋亡是一种由遗传基因决定的程序性细胞主动死亡,生物反应器中大多数细胞的死亡都是因细胞凋亡引起的.为防止细胞培养过程中的细胞凋亡,一般可采用如下三种重要措施:①通过培养基和氧的优化供应防止营养和氧的缺乏.②用化学添加剂如抗氧化剂等阻断细胞凋亡过程.③采用抗细胞凋亡基因改造工程细胞.由于细胞凋亡是由一系列蛋白质控制的过程[32I,因此在目的细胞中引入某些特定基因有可能抑制细胞凋亡的发生.抗细胞凋亡的原癌基因bc l一2(B淋巴瘤/自血病蛋白一2)是被研究得最多的细胞凋亡基因,它在细胞凋亡级联反应中处于重要环节.通过遗传工程使哺乳动物细胞表达抗凋亡基因bc l一2,在大多数情况下虽不能防止细胞死亡,但能延长细胞寿命,增加产物产量【33|.Si m ps on等(1997)将bcl 一2稳定转染TB/C3细胞后,细胞活力增强,抗体滴度上升【341.I t oh等(1995)也同样发现转染了bc卜2的2E3细胞,其抗体生产能力比未转染细胞提高了4倍【351.bc l一2家族中的另外一些成员可能也有与bcl一2一样甚至更强的抑制细胞凋亡的作用.如bc l—X L在许多哺乳动物细胞中得到表达后,可对有些诱导因素如营养物缺乏、辐射和糖皮质激素等引起的细胞凋亡有抑制作用[36,37】.另外有实验表明,当共表达bag一1和bcl-2基因时,可产生有相同或协同抗细胞凋亡作用[38|.L e e等(1996)在提高bcl-2表达水平的同时降低凋亡的最终执行者C as pase一3的表达水平,能明显提高细胞的抗凋亡能力[39I.近年来,人们将抗凋亡基因bc卜2应用于动物细胞培养做了大量工作[40|.另外,对bcl—X L或bcl—2的突变体研究表明,抗凋亡基因的出现可对抗营养及血清限制、有毒物质和代谢废物积累、氧消耗、流体力学应激等引起的凋亡,提高细胞活性,延长培养周期,增加最终细胞密度和产物浓度.Fi guer oa等(2004)在C H O细胞内同时表达抗凋亡基因aven和bcl一2家族成员bcl—X L,同时表达比单独表达ave n或bcl—X L具有更强的抗凋亡能力,在无血清培养基中的各重组细胞的活力分别为85%,50%和30%[41].另外,采用反义技术将前凋亡基因c—j H n转染细胞也可获得具有抗凋亡以及增殖受控的细胞系,亦是抗凋亡的一种策略【42|.2.2细胞周期调控工程理想的生产过程必需同时维持细胞活性状态以及产物蛋白的表达,即首先使细胞快速增殖到高密度,在细胞凋亡发生之前,控制细胞增殖速率并诱导其进入一个增殖静止期,即产物形成期,此时细胞将获得的代谢能量从用于细胞增殖转为用于产物分泌,细胞维持在活性相对较低的存活状态.通过控制细胞增殖速率,产物分泌量得以大大提高.随着对细胞周期调节机制研究的深入,研究人员已将细胞周期调控基因应用于规模化培养的细胞增殖控制.Fuss enegger等(1998)在C H O细胞中分别表达了p21、p27和肿瘤抑制基因p53三种细胞周期G1/S抑制蛋白,通过抑制cycl i n—E—C D K2复合物的磷酸化活性,阻止细胞进入s期.其中,p21基因转染后细胞获得了生长抑制.其生长静止可持续几周之久,报告蛋白分泌碱性磷酸酶(secr et ed al kal i ne phospha t ase,SE A P)的单位产量增加10--15倍;p27转染后的细胞生长静止但并不发生凋亡,产物表达量增加15倍.由三种蛋白组成的三顺反子元件转染C H O后,SE A P产量比对照提高30倍[433.2.3糖基化工程蛋白质有两种糖基化方式:N一糖基化和O~糖基化.一般来说,0一糖链对蛋白质的特性影响不大,而N一糖链的不同对产品可产生较大影响.N一糖链通常都有一个五糖核心,即M a n al一6(M an Q1)一3(M an81)--,4G1cN A c81-,4G1cN A c.根据外层链的不同,可分为:高甘露糖型、杂合型与复合型[55].三种因素决定糖链类型不同:①合成肽链的不同.②细胞内糖基化酶的不同.③细胞培养环境的影响.用C H O细胞表达的糖蛋白,其类型与人的尽管近似,但也不尽完全相同.C H O细胞缺少Q一2,6~唾液酸转移酶的功能,因此缺少唾液酸化的糖基.为此,一面人们正尽量寻找能用以生产糖蛋白类药一47—物的人类细胞,如用N a m al w a细胞表达t—pA[44l、pr o—U K[45]和EP O[46|,它们的糖基化和人的自然产品一致.另一方面,人们正打算采用“糖基化工程”(gl ycos yl at i on engi neer i ng),即应用基因工程手段,人为地改变肽链结构、增加某些酶基因以及改进和控制某些培养条件等,以达到正确糖基化的目的.如在t—PA基因中进行点突变,改变了一个氨基酸,使之增加一个糖基化位点,使t—PA在血浆中的清除率比原来减少了10多倍【471.又如M i neh等(1995)将t—PA与Q一2,6一唾液酸转移酶基因共转染C H O 细胞,结果表达的t—PA的糖基化情况与人类的更接近【48】.Lam ot t e等(1999)将丫一干扰素基因与a一2,6一唾液酸转移酶基因共转染C H O细胞,结果了一干扰素中a一2,6一唾液酸化程度比对照提高了68%(不加丁酸钠)和82%(加丁酸钠).此外,控制和改变细胞的培养环境对N一糖基化也可产生一定影响【491.H os oi等(1996)研究表明,通过添加地塞米松、改变培养基糖的成分、使培养温度突然下降等,都可使pr o—U K的糖基化形式从含岩藻糖的2分枝复杂型寡糖链,转变为含岩藻糖的3或4分枝复杂型寡糖链,从而使生产的重组蛋白质糖基化形式与人类的一致[s0].2.4多基因调控代谢工程由于细胞内部调控网络在空间和时间上的复杂性,一个性状并不完全由某一个基因控制.最新的代谢工程策略提出在细胞内进行可分别调控表达的多个基因,以期形成一个模拟细胞调控网络的人造调控系统,最简单的是双调控表达技术.Cor nel i a等(2001)通过链阳菌素和四环素双诱导表达体系构建了重组C H O细胞株,能分别表达p27基因和它的反义链.当诱导p27基因表达时实现了细胞生长的停滞,处于G l期细胞增加了40%,而表达p27基因的反义链,则使细胞内自身p27蛋白表达下降,使细胞增殖的速度提高了一倍,促进了细胞的生长[51,52】.2.5细胞增殖控制工程工业化大规模细胞培养中,常采用无血清/无蛋白培养基(Ser um f ree m edi um,SFM/Pr ot ei n f r ee m e di um,PFM)来降低细胞培养和产品纯化的成本.但SFM/PFM缺乏生长刺激因子、粘附因子、扩展因子以及其他细胞生长存活所必需的成分,在培养过程中常表现出细胞活力降低、贴壁性差等现象,细胞增殖能力下降,进而导致分泌目的蛋白的能力下降.在培养基中添加胰岛素和成纤维细胞生长因子可使细胞恢复增殖能力,同时,细胞内的细胞周期调控因子C ycl i n—E的表达也增加.C ycl i n—E能使细胞周期的G1期延长,s期缩短,提示人们可通过表达C ye l i n—E的方法来增加细胞的增殖能力.人们还通过将生长刺激因子、粘附因子基因导入C H O细胞中,让C H O细胞本身提供其自身生长所需要的成分,使其能在S FM/P FM中良好生长.G andor等(1999)将介导C H O细胞在SF M中贴壁和扩展的玻表粘连蛋白(vi t r onec t i n)基因置于M M T V启动子控制下导入Cl I O细胞,使其获得了在PFM中j!占壁生长和扩展的能力[53].另外,同时表达胰岛素样生长因子(I G F一1)和转铁蛋白(t r ansf er r i n)的所谓“超级C H O细胞”在PFM上也能生长良好【54J.随着对细胞周期、细胞凋亡、信号传导,以及细胞周期的调控机制等各方面机理认识的不断深入,可以通过载体的系统优化,将编码细胞生长刺激因子、黏附因子、扩展因子、抗凋亡因子、转录与翻译的反式作用因子以及其他细胞生长存活所必需成分的基因和顺式表达调控元件转入宿主细胞,以提高其表达.与此同时把不利于目的基因表达的基因从宿主细胞中敲除或下调其在宿主细胞中的表达,从而把细胞改造为能在SFM/PFM中培养、培养前期细胞增殖快、当细胞密度达到理想值时细胞长时间不增殖、抗凋亡能力强、高表达目的基因的宿主细胞,最终大幅度降低哺乳动物细胞生产蛋白质类药物的成本.参考文献:[1]涂宣林,宋后燕.寡糖链的研究进展【J].药物生物技术,1998,5:55—59.[2]Ja yapal K P,W l as ehi n K F,H u W S,Y a p H G S.R ec om bi nant prot ei n t he rape ut i c s f r om C H O c el l s一20yea r s and cou nt i ng [J J.C he m Eng Pi ng,2007,103:40—47.[3]Li F,N at araj an V,Shen A,R o ber t K,A m anuU ah A.C e i l cul t ur e proc es se s f or m onocl onal ant i bo dy pr oduct i on[J].Lande-———48..——。
原核,昆虫,哺乳动物表达系统 对比

表达系统是生物体进行交流和传递信息的重要工具,不同类裙的生物体在表达系统上有着独特的特点和功能。
本文将就原核、昆虫和哺乳动物的表达系统进行对比分析。
一、原核1. 原核生物是一类较为简单的生物体,其表达系统主要包括RNA转录和翻译,以及一些原核生物特有的机制,如转座子、限制酶等。
2. 在原核生物中,基因的表达和调控相对简单,通常是通过DNA的转录产生mRNA,然后再通过翻译产生蛋白质。
原核生物的基因组相对较小,基因的结构也相对简单。
3. 原核生物的表达系统不仅包括基本的基因表达,还包括许多在分子水平上进行信息交流和传递的机制,如质粒介导的DNA转移、RNA 介导的基因沉默等。
二、昆虫1. 昆虫是一类较为复杂的生物裙体,其表达系统包括了多种不同的信号传导和调控机制,如激素系统、化学信号系统等。
2. 在昆虫中,基因的表达和调控已经相对复杂起来,有许多基因调控网络参与其中。
除了mRNA的转录和翻译外,昆虫还具有一些特殊的表达机制,如miRNA介导的基因沉默、垂体激素调控等。
3. 昆虫的表达系统在进化上相对比较保守,但在不同物种和不同环境中,表达系统也会出现一些特殊的适应性和多样性。
三、哺乳动物1. 哺乳动物是一类高度复杂的生物裙体,其表达系统包括了多种不同的信号传导和调控机制,如内分泌系统、神经系统等。
哺乳动物的基因组相对较大,基因的结构也十分复杂。
2. 在哺乳动物中,基因的表达和调控已经相对复杂起来,有许多基因调控网络参与其中。
哺乳动物具有多种不同的表达机制,如DNA甲基化、组蛋白修饰等。
3. 哺乳动物的表达系统在进化上相对比较灵活,不同物种和不同环境中会呈现出不同的表达模式和调控机制。
在哺乳动物中,基因表达的调控和信号传导机制十分复杂,涉及到许多不同的细胞信号通路和调控网络。
原核、昆虫和哺乳动物在表达系统上具有明显的差异。
原核生物表达系统相对简单,主要是基因的转录和翻译,以及一些特殊的表达机制。
昆虫和哺乳动物的表达系统则相对复杂,涉及到多种不同的信号传导和调控机制。
哺乳动物细胞表达系统的应用

哺乳动物细胞表达系统的应用哺乳动物细胞表达系统是在哺乳动物细胞中表达外源蛋白的一种常用方法。
这种系统的应用非常广泛,包括基础科学研究、药物开发、生物制药等多个领域。
以下是哺乳动物细胞表达系统的一些主要应用:
1.基础科学研究:
•蛋白功能研究: 通过在哺乳动物细胞中表达外源蛋白,研究者可以深入了解蛋白质的结构、功能和相互作用。
•信号转导研究: 使用哺乳动物细胞表达系统可以模拟细胞内的信号传导途径,帮助研究人员理解细胞信号调控的机制。
2.药物开发:
•蛋白药物生产: 许多生物制药品,如重组蛋白、抗体和疫苗,需要在哺乳动物细胞中表达并进行生产。
•药物靶点验证: 通过在哺乳动物细胞中表达目标蛋白,可以进行新药靶点的验证和筛选。
3.疾病研究:
•疾病模型: 利用哺乳动物细胞表达系统,可以构建疾病相关蛋白的细胞模型,用于研究疾病的发病机制和筛选潜在治疗方法。
•蛋白标记: 将蛋白质标记为荧光蛋白或其他标记物,用于追踪细胞内的特定蛋白。
4.细胞工程:
•基因治疗: 利用哺乳动物细胞表达系统可以生产基因治疗药物,用于治疗一些遗传性疾病。
•细胞药物输送: 将药物载体或纳米颗粒表达在哺乳动物细胞中,用于细胞药物输送研究。
5.病毒研究:
•病毒复制机制: 使用哺乳动物细胞表达系统可以研究病毒的生命周期、复制机制和与宿主细胞的相互作用。
•疫苗研发: 通过表达病毒蛋白,可以进行病毒样粒子 VLP)疫苗的研发。
这些应用表明,哺乳动物细胞表达系统在科学研究和药物开发中具有重要作用,能够提供生产复杂蛋白和进行多方面生物学研究的有效工具。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
动物基因工程原理
特点与优势
哺乳动物细胞表达系统的优势在于能够指导蛋白 质的正确折叠,提供复杂的N型糖基化和准确的O型 糖基化等多种翻译后加工功能,因而表达产物在分 子结构、理化特性和生物学功能方面最接近于天然 的高等生物蛋白质分子。
应用领域
哺乳动物细胞表达系统已成为多种基因工程药物 的生产平台。
已用该细胞株获得了包括人组织纤溶酶原激活剂、生长因 子等在内的多种蛋白的高效稳定表达。
对细胞株选择性地进行遗传改造
基于腺病毒EIA蛋白可反式激活CMV启动子,Cockett等建 立了稳定表达EIA的CHO细胞株,在该细胞中重组前胶原 酶的产量与CHO细胞相比增加了l2倍;该细胞在多种抗体 的表达中亦取得满意的结果。
哺乳动物细胞表达系统在新基因的发现、蛋白质 的结构和功能研究中亦起了极为重要的作用。
研究现状
部分蛋白在哺乳动物细胞中的表达已从实验室研究迈向生 产或中试生产阶段。
已有许多重要的蛋白及糖蛋白利用哺乳动物细胞系统表达 和大量制备、生产。如人组织型血纤蛋白酶原激活因子、 凝血因子Ⅷ、干扰素、乙肝表面抗原、红血球生成激素、 人生长激素、人抗凝血素Ⅲ,集落刺激因子等。
启动子需包含两个识别序列:mRNA转录起始点和TATA 盒。TATA盒位于转录起始位点上游25--30bp处,是引导 RNA聚合酶在正确起始位点转录所必需的序列,即保证转 录的精确起始。其他上游启动子元件常位于TATA盒上游 100~200bp之间,其功能是调节转录的起始频率和提高转 录效率。
真核表达载体-启动子
高等哺乳动物受体细胞的条件
以高效表达外源基因为目标的高等哺乳动物受体细胞应具 备以下条件: 细胞系特征:丧失细胞接触抑制和锚地依赖特征,便于大 规模培养。 遗传稳定性:外源基因多次传代后不至于丢失,易于长期 保存。 合适的标记:便于转化株的筛选和维持。 生长快且齐:分裂周期短,生长均一,便于控制。 安全性能好:不合成分泌致病物质,不致癌。
真核表达载体-启动子
另一类杂合启动子是由活性很低的启动子与数个转录激活 因子结合位点串联而成,这些位点与转录激活因子的结合 受细胞外小分子药物的调控;这类启动子主要用作基因的 诱导表达。
如Invitrogen公司的ecdysone(脱皮激素)诱导表达系统, 其负责目的基因转录的启动子是由热休克蛋白启动子核心 序列(minimal promotor)和5个E/GRE元件组成;
高等哺乳动物受体细胞的遗传标记
遗传标记 编码产物
筛选药物
作用机理
APH-
氨基糖苷磷酸转移酶 G418
APH灭活G418
DHFR HPHTK-
二氢叶酸还原酶 潮霉素B磷酸转移酶 胸腺嘧啶核苷激酶
氨甲喋呤 潮霉素B 氨甲喋呤
DHFR变体抗氨 甲碟呤
HPH灭活潮霉素 B
TK合成TMP
XGPRTADA-
黄嘌呤鸟嘌呤磷酸核糖 霉酚酸 转移酶
因而可通过控制 tet 的浓度来控制基因表达的水平。
▪ 应用于研究基因不同水平的表达对细胞,个体发育的影响
有何不同。
作用机制示意图
四环素调控表达系统构成元件
▪ 四环素阻遏子(Tet repressor protein,TetR) ▪ 四环素操纵子(Tet operator DNA Sequence, TetO) ▪ 单纯疱疹病毒Vp16转录活化区(Vp16 activation domain,AD) ▪ 四环素调控反式激活子(Tetracyeline-Controlled
对细胞株选择性地进行遗传改造
对细胞其它特性的遗传改造,包括细胞生长周期 的调控、细胞的抗凋亡能力、细胞贴壁能力、蛋 白糖基化的模式等方面,亦有相关报道,其实际 应用价值有待得到更多实验数据的支持。
真核表达载体
哺乳动物细胞表达载体的必要元件包括:一个高活性的启 动子、转录终止序列和一个有效的mRNA翻译信号。
IRES位于小RNA病毒RNA5’非编码区, 约为450bp。 IRES可与它连接的ORF同时翻译,构建双顺反子和 多顺反子真核表达载体。
分泌表达载体
可调控的哺乳动物细胞表达系统
四环素(tet)调控表达系统
▪ 加入或撤除四环素和四环素衍生物来调控基因表达。 ▪ 外源基因表达的水平与所加的 Tc 浓度在一定范围内相关,
启动子和增强子受细胞类型的限制,在不同的细 胞系中有很大不同,因此需根据宿主细胞的类型 选择不同的人巨细胞病毒早期启动 子(CMV-IE)、人延伸因子1-亚基启动子和Rous肉 瘤长末端重复序列;Invitrogen公司开发的pcDNA、 pEF和pRL三种系列载体即分别是以这三种启动子 驱动目的基因的表达。
哺乳动物细胞表达系统的表达水平从整个水平上看仍偏低, 一般处在杂交瘤细胞单克隆抗体蛋白产率的下限,即130μg/l08细胞/24小时。其限速步骤可能是在工程细胞中重 组蛋白的分泌效率较低。
一、稳定表达细胞系的筛选
构建目的基因的表达载体。 确定受体细胞。 确定筛选方法(抗生素的使用浓度)。 转染后48h,传代铺板 (1 : 100) ,加筛选药物。 3-4d换液1次,直至存活细胞长出岛状细胞簇,约2周。 克隆耐药细胞,检测目基因的存在与表达。
据报道红细胞生成素启动子在1%氧浓度比在21%氧浓度下活 性增强100倍。这又是一种新的提高表达量的作用机制。
真核表达载体-增强子
常用的增强子有Rous肉瘤病毒基因长末端重复序列和人巨 细胞病毒增强子。
James等利用糖皮质类固醇诱导的鼠乳房瘤病毒(MMTV)启 动子和鼠乳房瘤病毒长末端重复序列(MMTV-LTR),在 CHO细胞中表达分泌的碱性磷酸酶,产量为利用常规的 SV40和 CMV启动子的10倍,超过0.4mg/ml。
XGPRT和成 GMP
腺嘌呤核苷脱氨酶
腺嘌呤木酮糖苷 ADA-灭活Xyl-A
宿主细胞
常用的非淋巴细胞类有中国仓鼠卵巢(CHO)细胞、小仓鼠 肾(BHK)细胞、COS细胞、小鼠NSO胸腺瘤细胞和小鼠骨 髓瘤SP2/0细胞等。
不同宿主细胞表达的重组蛋白其稳定性和蛋白糖基化类型 不同,需根据要表达的目的蛋白选择最佳的宿主细胞。
常用的高等哺乳动物受体细胞
迄今为止,用于医疗用品(药物、抗体、诊断试剂)大规 模生产的高等哺乳动物受体细胞主要还是中国仓鼠卵巢细 胞(CHO),其优势有如下几个方面: 遗传背景清楚,生理代谢稳定 与人的亲缘关系接近,外源蛋白修饰准确 基因转移和载体表达系统完善 耐受剪切力,便于大规模培养 被美国FDA确认为安全的基因工程受体细胞
宿主细胞
猴肾细胞(COS)是进行外源基因瞬时表达时用途最广的 宿主,其重组载件易于组建,便于使用,而且对插入DNA 的量或者采用基因组DNA序列的情况都没有什么限制,便 于通过检测表达情况来确证cDNA的阳性克隆,也利于快 速分析引入克隆化cDNA序列中的突变。
宿主细胞
近几年也陆续发现了几种新的有较大应用价值的细胞株:如 来源于MadIin-Darby犬肾的高分化内皮细胞株(MDCK)。
内含子
某些内含子有重要的基因功能调控序列,目的基因以基因 组形式克隆入载体能得到高表达。
为增加转录产物的稳定性,表达载体中一般含有天然的或 人工合成的内含子序列。
mRNA前体的剪切能够促进mRNA 从胞核向胞质的运输。 外显子和内含子接头 每个外显子和内含子接头区都有一
段高度保守的一致序列(consensus sequence),即内含子 5’末端大多数是GT开始,3’末端大多数是AG结束,称为 GT-AG法则,是普遍存在于真核基因中RNA的识别信号。
原核DNA序列,包括能在大肠杆菌中自身复制的复制子,便 于筛选含重组细菌的抗生素抗性基因,以及便于目的基因 插入的限制性酶切位点。
可视实验需要加入标志基因、内含子、内部核糖体进入位 点等。
真核表达载体
常用的标记基因有胸腺激酶(tk)基因、二氢叶酸还原酶 (dhfr)基因、新霉素(neo)抗性基因、氯霉素乙酰基转移酶 (cat)基因等
Pei等用该细胞株表达分泌型的基质金属蛋白酶MMPI3,发现 高表达的阳性细胞克隆可占转染细胞的5%~l0%,其中一个 克隆的表达量可占细胞上清总蛋白的l5%~20%,在细胞单层 贴壁培养情况下表达量达10 mg/L。
对细胞株选择性地进行遗传改造
BHK/vl6细胞株是稳定表达单纯疱疹病毒(HSV)VP16蛋白 的BHK细胞,由于VP16的转录激活作用,载体中的HSV 早期启动子在该工程细胞中有很高的活性。
真核表达载体-启动子
近年来又发现了一些新的强启动子,如:人 leukosialin基因启动子和鼠3-磷酸甘油激酶l (PGKI) 基因启动子,活性与CMV-IE相当。
人泛素蛋白(ubiquitin)C基因启动子不仅具有较高 的活性,而且比CMV-IE、PGK1等启动子有更广 泛的宿主细胞范围,几乎在转基因小鼠的所有组 织细胞中都具有较高活性。
真核表达载体-启动子
构建杂合的启动子是获得新启动子的一个重要途径。
比如由人ubiquitin C启动区序列与CMV增强子组成 的杂合启动子、由SV40早期启动子和人I型T淋巴 细胞病毒LTR中的增强子序列(R-U5片段)组成的 SR-α启动子的活性均与CMV-IE相当;
由鸡β-肌动蛋白启动子和CMV增强子序列构成的 杂合启动子不仅活性比CMV—IE高,而且具有更 为广谱的宿主细胞范围。Novagen公司的pBacMam 表达系统即是以该杂合启动子驱动目的基因的转录。
二、哺乳动物细胞表达系统
(一)宿主细胞 (二)表达载体的构建 (三)表达细胞株的基因共圹增和筛选方法 (四)可调控的哺乳动物细胞表达系统
高等哺乳动物受体细胞
高等哺乳动物细胞的生长特性 高等哺乳动物受体细胞的条件 高等哺乳动物受体细胞的遗传标记 常用的高等哺乳动物受体细胞