有孔虫
有孔虫

•
• •
• •
(三)T-J有孔虫的复苏时期 古生代末期生物大规模绝灭(mass extinction)事件对有孔虫产 生了极大的影响,纺缍虫全部绝灭于P末,内卷虫超科百分之九 十以上的科绝灭,所以T-J是绝灭事件以后的一个复苏时期,其特 点如下: 1、曲杖虫超科和小粟虫超科,新产生了40%的科,自∈纪就 开始出现的瓶虫超科(Lagynacea)和砂盘虫超科 (Ammodiscacea)的发展远比不上曲杖虫和小粟虫超科的发 展。 2、钙质多孔壳的超科开始大量出现,产生了六个超科,而且 有孔虫的分异度开始回升。有重要意义的是新产生的双口虫超 科(Duostominacea)和抱球虫超科(Globigerinacea)。其 中双口虫超科出现于T,而绝灭于中侏罗世,演化快,可视为 标准化石。而且具有演化方面的意义,许多后来的钙质透明壳 都是由他演化而来。抱球虫超科是有孔虫中唯一的浮游类型, 它的出现是有孔虫演化史上的一件大事,有着重要的地质意义。
• • •
(二)C-P有孔虫发展的第一个繁盛时期 C-P是古生代有孔虫最繁盛的时期,出现了许多划分对比地层 的标准化石,主要特征为: 1、钙质微粒壳类型达到极盛,内卷虫超科(Endo-Thyrian) 和纺缍虫超科(Fusulinacea)在这一时期极大的发展,而绝 大多数灭绝于P末期,构成了这一时期的标准化石。 2、曲杖虫超科(Lituolacea)的产生与兴旺,胶结壳的曲杖虫 超科开始出现于C,并立即兴旺超来,这个超科的10个科有一 半始于C、P。 3、小粟虫目(Miliolina)从C纪开始出现。 4、钙质多孔壳类型开始萌芽,如节房虫(Nodosariacea)。
•
•
100 120 140 160 180 160 140 120 100
第二章 有孔虫

144o 72o 216o 横 切 面 0 288o
o
顶 视
0o
壳 侧 视
壳 Байду номын сангаас 视
13
D、曲房虫式,相继生长的两个房室的绕旋平面夹角由120°逐渐增大到180°, 变化的绕旋平面构成“S”形,壳面上可见5个以上的房式,有时可达8-10个房室, 例如曲房虫(sigmoilina)。 5、混合型壳,同一个壳体,早期和后期房室排列形式不同,至少有两种以上的 排列方式;如梯状虫(Climacammina),早期为螺旋双列,后期为单列式;双面 包虫(Dyocilbicides,早期为低螺旋形,后期为螺旋双列。坑壁虫早期平旋,晚 期伸展为扇形。 6、环圈式壳,壳的房室呈同心状排列,壳体呈圆盘状,如环圈虫(Sorites)。
伪足
硅藻
口孔
壳
外 质 内质
现代有孔虫
1
浮游有孔虫活个体
2
有孔虫的生活环境:现代有孔虫绝大多数生活在正常海域,少数生活在泻 湖、沼泽、河口等滨海边缘环境或残留海水等半咸水水域。 有孔虫的生活方式:大多数有孔虫营底栖生活,少数营浮游生活。底栖有孔 虫可以分为小有孔虫和大有孔虫,小有孔虫直径一般为2.5mm以下,大有孔虫 一般大于2.5mm。
幼年配子体 成 年 配 子 体
成 年 裂 殖 体
幼年裂殖体
3
二、有孔虫壳体房室的排列方式
有孔虫其原生质分泌壳质形成壳子,或分泌胶结物粘结其它颗粒形成壳子, 壳子中空,称为房室或壳室。房室顶端有一圆形或其它形状的开口叫口孔。多个 房室组成的壳,其最早形成的,最小的房室叫做初房,以后继续分泌壳质形成第 一房室……最后一个房室叫终室,分隔两个房室的壁称为隔壁,隔壁是壳壁向前 沿伸时向内转折的部分,隔壁与壳壁相交的线称为缝合线。口孔位于终室的顶端 ,其形状变化多样,口孔周围的壁称为前壁。
有孔虫

有孔虫卵形厚壁虫Pachyphloia ovate Lange1)分类位置:原生动物门Protozoa肉足虫纲Sarcodina有孔虫目Foraminiferida轮虫亚目RotaliinaDelage&Herouard节房虫超科nodosariacea Ehrenberg节房虫科Nodosariidae Ehrenberg节房虫亚科Nodosariinae Ehrenberg厚壁虫属Pachyphloia Lange,1925模式种---虱厚壁虫Pachyphloiapediculus Lange,1925卵形厚壁虫Pachyphloiaovate Lange2)特征:钙质透明壳,主要成分为方解石,壳壁微细构造为透明多孔放射状;隔壁分层性为单层隔壁;房室6-9个,排列方式为平旋、单列或绕旋式;口孔末端放射状,少数为缝状或圆形。
壳体小,呈卵圆形。
壳高0.23-0.64mm,壳宽0.11-0.34mm。
初房外径0.03-0.10mm。
3)地层分布:该种的地理分布较广,二叠系各地层单位中均有发现。
(下面在部分地区讨论)隆林阶的有孔虫群Bradyina-Pachyphloia组合带中可能含少量卵形厚壁虫Pachyphloia ovate Lange栖霞阶的下、中、上部均有Pachyphloia ovate Lange 的分布,分别在广西柳州、隆林、靖西,湖南浏阳、礼县,湖北秭归、五峰,贵州石阡等地有发现。
茅口阶的Pachyphloia异常丰富,但其种群与栖霞阶的种群大致相似,所以说茅口阶的有孔虫在一定程度上与栖霞阶的有孔虫有许多共同的特点,茅口阶中的一部分有孔虫是由栖霞阶延续上来的。
吴家坪阶直列式的Pachyphloia等属占优势,分布也最为广泛。
这些有孔虫类,甚至在其他许多海生动物都不能生存的情况下,仍有他们的踪迹,说明它们具有比较强的生命力,有比较环境条件适应性。
长兴阶在华南地区,由于稳定的环境条件,直列式的有孔虫Pachyphloia等属的发展达到高潮。
第六章第二节 1-有孔虫

图1 有孔虫Myxothera arentlega 的世代交替 (据Loeblich et Tappan ,1978再版) 第二节微体生物1有孔虫(编写人:西南石油大学王占磊)1有孔虫一、概述有孔虫是微小的真核单细胞动物。
其大小一般在1mm 以下,最大可达110mm 。
在分类位置上属原生动物门(Protozoa)肉足虫纲(Sarcodina)中的一个亚纲。
有孔虫是一种微小的单细胞动物,由一团原生质构成。
原生质分化为两层:外质较薄而透明,伸出许多分叉、分枝的丝状伪足起运动和捕食的作用。
外质还分泌钙质或胶结物建造硬壳。
内质颜色较深,细胞核和细胞器包于其中,司消化和生殖等机能。
外质和伪足分泌壳质构成有孔虫壳。
有孔虫有两种生殖方式,无性生殖和有性生殖交替进行,这种现象叫世代交替。
无性生殖为简单的复分裂叫裂配生殖。
成熟的裂殖体分裂产生许多无性胚胎母体,每个无性胚胎发育成一个配子母体,成熟的配子母体产生许多带鞭毛的配子,两个配子接合形成合子为有性生殖。
合子又发育为裂殖体。
裂殖体成熟再进行无性生殖。
这种世代交替现象反映在壳上为无性生殖产生的配子母体世代的壳之初房较大,叫显球型壳;有性生殖产生的裂殖体世代的壳之初房较小,叫微球形壳。
这就是有孔虫壳的双型现象,在鉴定时要特别注意。
现代有孔虫绝大多数是海生生物,极少数属种生活于泻湖,河口等半咸水区域仅假几丁质壳的瓶形虫超科(Lagynacea )的个别属种发现于淡水。
大多数有孔虫营底栖生活,少数在海面浮游。
底栖有孔虫多在浅海底或海藻上缓慢移动。
少数固着生长,从大陆架到4000多米的深洋中都有分布,浮游有孔虫死后,其壳沉落海底甚至在洋底形成抱虫球软泥。
有孔虫化石最早发现于寒武纪、石炭纪、二叠纪为古生代有孔虫的极盛时期,以蜓类和内卷虫类占绝对优势。
中生代初期一度衰退。
到侏罗纪开始再次勃兴,白垩纪又极繁盛,其程度大大超过了晚古生代后期。
第三纪是有孔虫发展史上的全盛时代。
其中很多分子延续到现代。
底栖有孔虫分子多样性及其环境特征研究初探

底栖有孔虫分子多样性及其环境特征研究初探为探索不同海区类型沉积物中底栖有孔虫的分子多样性及与环境因子之间的相互关系,本研究从潮间带、海湾、陆架浅海一直到深海大洋区,共采集了81个站位或季节的共计243个样品。
提取环境样品的总DNA,利用有孔虫特异性引物(s14F3-s17)扩增其18S rRNA基因3’端的一个高变区(大约300 bp),使用Illumina HiSeq 2500测序平台进行双端250 bp测序。
此外,为探索不同DNA提取和染色方法对有孔虫分子鉴定研究的影响,本论文进行了有孔虫单种实验方法学研究。
具体工作包括如下:1.DNA提取和染色方法对有孔虫分子鉴定研究的影响研究以野外采集的钙质瓷质壳类底栖有孔虫五玦虫属Quinqueloculina spp.(Foraminifera,Miliolida)为研究材料,研究经虎红染色后虫体的DNA质量,以此评估虎红染色对有孔虫分子实验的影响。
通过方法学的摸索可知,经1 g/L虎红染色36 hrs的大部分瓷质壳类底栖有孔虫样本可以成功提取到DNA,并且可以用于下游的PCR等分子实验。
2.青岛湾潮间带底栖有孔虫分子多样性月变动及与环境关系研究14个月采集的样品经测序共得到605种底栖有孔虫。
沉积物中底栖有孔虫物种数在冬季和夏季较多,秋季次之,春季最少。
而分子多样性的季节变化整体上呈现夏季高,秋冬季节次之,春季少的规律。
Rotaliida相对丰度在秋冬季节较高,春季其次,夏季较少,Miliolida相对丰度随季节的变化趋势与之相反。
优势属为:Ammonia(卷转虫属),Textularia(串珠虫属),Epistominella(小上口虫属),Operculina(盖形虫属),Bulimina(小泡虫属)。
Epistominella及Operculina相对丰度与水温和泥温均呈显著正相关。
底栖有孔虫的群落多样性在春季和夏季与水温和泥温正相关,与盐度负相关;在秋季与水温、泥温和盐度均呈负相关;而在冬季与水温和泥温呈负相关,与盐度呈正相关。
有孔虫群体行为的生物学意义

有孔虫群体行为的生物学意义有孔虫是一种海洋浮游生物,分布于淡水和海水中。
它们是生物群体行为的杰出代表,能够在形态和行为上形成各种复杂的结构和模式。
有孔虫群体行为的生物学意义一直备受关注,研究人员利用现代技术和方法进行了大量深入的研究,为我们解开了很多谜团。
一、有孔虫群体行为的基础特性有孔虫是一种单细胞微生物,直径一般在0.1~0.5毫米之间。
它们在水中自由游动,在觅食、逃避捕食和繁殖等方面表现出协同行为。
最显著的特征是它们能够形成一系列复杂的群体结构,也可以快速地改变这些结构来适应环境的变化。
通过这种行为,有孔虫在地球生命演化史上扮演了重要的角色。
二、群体行为的涉及原则有孔虫群体行为的涉及原则很多,其中最重要的是基质特异性、化学梯度、机械应力、光线、触角互作和细胞信号等。
通过这些原则的共同作用,有孔虫能够感知周围环境的变化,协调个体行为并调整群体结构,以适应环境中的适应性选择压力。
三、有孔虫群体行为的作用有孔虫群体行为的作用非常多,其中最重要的几个方面如下。
(一)保护和捕食有孔虫能够通过合作来保护群体内的每个个体,例如它们可以形成隐藏或疏散的群体,以逃避捕食压力。
同时,有孔虫还可以利用群体行为来协同捕食微小的有机物或浮游生物,以维持群体的生存。
(二)繁殖和增长有孔虫群体行为还可以促进个体之间的繁殖,从而增强种群的持续增长。
例如当群体处于适宜环境下时,有孔虫可以聚集在一起并释放性激素以调节个体繁殖和产生子代的数量。
这种的群体繁殖模式也使得这些单细胞生物在短时间内就可以达到相当大的数量。
(三)环境调节群体行为也可以帮助有孔虫对环境的变化进行调节和适应。
例如,面对水质变化、氧气含量变化等挑战,有孔虫可以改变群体结构、形成更加紧密的集群,以应对不同的环境应激。
四、总结有孔虫群体行为的生物学意义之丰富,越来越多的研究正在加深人们对这些微生物的认识。
其群体行为机制的深入研究可以为我们探索类似社会生物行为和人类社会行为方式提供新的洞见和思路,而这些还仅仅只是生物学研究的冰山一角。
有孔虫化石分析鉴定方法

有孔虫化石分析鉴定方法
孔虫化石是具有有效史前价值的古生物化石,为了确定它的属,并获得有关它
的史前历史信息,必须对其进行鉴定。
鉴定孔虫化石的方法有多种,如形态鉴定、卵囊鉴定、分子遗传学鉴定、生物学鉴定等。
1、形态鉴定
形态鉴定技术是最常用的孔虫化石鉴定方法,它利用视觉技术对化石的外形和
内部结构进行检查,以便充分了解孔虫化石结构及其成因,从而确定其属和种属。
它可以分析孔虫结构重要的标志,如形态特征、羽节段数、头圈形态、足圈足袋等。
2、卵囊鉴定
卵囊鉴定是孔虫化石鉴定的另一种重要方法。
卵囊鉴定与形态鉴定类似,但在
形态鉴定的基础上,通过实验室对变形的化石进行系统的检查,比如变形的头圈结构和足圈结构;另外还可以检查它的内部结构,可以分析其身体部位,比如腹部、腔壁、鳃壳等,以辨别其属。
3、分子遗传学鉴定
分子遗传学鉴定基于形态学鉴定的原理,利用实验室进行特定的地面操作和比较,以揭示孔虫的遗传标志和遗传型。
尤其是在以前的形态鉴定和卵囊鉴定中没有发掘出有效标志的情况下,分子遗传学鉴定尤其有用。
4、生物学鉴定
生物学鉴定也是孔虫化石鉴定的重要方法,它以生物学学科为基础,以分类学、系统发育学、数量遗传学等生物学理论为基础,对孔虫类群进行比较和探讨。
生物学鉴定可以更准确地讨论孔虫化石的属种,以及相关种群间的关系。
总之,以上就是检定孔虫化石的几种常用鉴定方法,其中各自都有各自的优势
和缺点,但只有结合使用才能发挥出最大的作用。
在实际工作中,可以在上述方法的基础上,根据实际情况,采取不同的方法,以便获得更准确的鉴定结果,为古生物学的研究和史前文明的研究发掘出更多的有价值的信息。
有孔虫ppt课件

幼年配子体
成
年
配
子
体
这个过程就是无性繁殖世代。配子母体达到成
熟就开始了有性繁殖世代,配子母体可产生大
量带鞭毛的配子,两个配子结合形成合子,合
成
子即可发育成长为新的裂殖体,如此反复循环,
年 裂
不断繁衍。有性世代产生壳的初房较小,称为
殖
微球型壳;无性世代产生的裂殖体壳的初房较
体
大,称为显球型壳。这样,同一个种中由于世
壳圈常包围前生长
的壳圈,根据包裹
的程度不同可将平
旋壳分为露旋壳,
包旋壳和半包旋壳。
两侧互为对称,素描时需要 露旋壳:侧视
壳缘视 侧视
素描侧视和壳缘视
8
3、螺旋式壳(frochospiral test),房室在若干个彼此平行的面上,围绕一 根通过初房中心并与这些平面垂直的假想的轴呈螺旋式排列。螺旋式壳有背 和腹之分。在背侧(旋侧),由于后生长的壳圈仅部分包裹先生长的壳圈, 因此初房和各壳圈均可显露;在腹侧(脐则),由于后生长的壳圈包裹了先 生长的壳圈,所以只能看见终壳圈的各房室,在壳的脐侧中央常出现下凹的 脐。二列式壳和三列式壳都是一种规则的螺旋式壳,二列式壳每个旋圈由两 个房室构成,三列式壳每个旋圈由三个房室构成。腹背不对称,需要素描背视、腹视和壳缘视
双房室有孔虫
直
圆盘形
管 形
球形 初房
螺 锥 形 口孔
之字形
6
(三)多房室壳 多房室壳由两个以上的房室构成,每一个房室都代表着个 体发育的一个阶段,由于房室的形状和排列方式不同,壳体形态变化很大,房 室排列方式可归纳成几种类型:
1、单列式壳(uniserial test)壳房室的生长沿直线或弧形单列排列形成单 列式壳,前者称单列直线形壳,后者称单列弧形壳。有的房室呈弧形沿扇面方 向增长,构成扇形(fan-shaped)壳,但仍属于单列式类型。在房室的生长过 程中,如后生长的房室仅掩盖了前一房室的口面部分,侧称为“不超覆”,如后 生长的房室不仅掩盖了前一个房室的口面,还掩盖了口面周围部分的壳壁,侧 称为超覆。
大海里的“小巨人”——记中国科学院海洋研究所郑守仪与‘有孔虫’

本刊记者/鸣莺有孔虫,海洋世界的小生灵。
它,有一个奇特而新颖的名字---大海中的小巨人。
郑守仪,海洋生物学家,中国科学院院士。
研究有孔虫60年,在海洋世界遨游了一生。
如今,这些生长在海洋几千米处,人类肉眼看不到的小生灵,却被郑守义‘请’出海面,与人类见了面。
一一亲手雕琢,按比例放大,上百倍“还原”, 栩栩如生,天然造型。
其中,240余个有孔虫属种原模相继被研究开发出科学教具,科普展品,旅游纪念品,服装服饰,首饰,灯饰,棒棒糖等有孔虫系列产品徐徐游进了人类的生活,进入了人们的视线。
大海里的“小巨人”——记中国科学院海洋研究所郑守仪与‘有孔虫’P ioneer 先锋本栏目冠名:英皇星艺(北京)文化发展有限公司宏观展示微观世界世界第一座“中国有孔虫雕塑园”,2009年,在广东中山三乡镇落成。
它由114座石雕组成,是集科学,艺术,海洋文化为一体的自然景观。
2012年,世界最大的美国史密森博物馆《Smithsonian》杂志,荐举中国有孔虫雕塑园为排列第二的世界10个进化旅游热点之一。
2003年,郑守仪在该领域荣获国际最高奖——美国库什曼“有孔虫”研究杰出人才。
今年6月5日,她从青岛市委书记李群手中接过《青岛市科学技术最高奖》颁奖证书。
有孔虫,从微观海洋世界来到到宏观人类大自然,历尽半个多世纪…显微镜前,青岛39中学生在院士指导下,观察亲自采集的有孔虫样本。
. All Rights Reserved.怀揣对“小巨人”的敬畏和探索时值盛夏,记者驱车驰进了青岛汇泉湾——中国科学院海洋研究所。
海洋生物标本馆,郑守仪研究室,不足20平的房间,俨然一个小型的有孔虫展览馆:模型,展架,标本,及开发的系列‘小巨人’琳琅满目占满了房间。
三面墙壁,环绕展柜,桌面展架,挂,摆,放满了‘有孔虫',凌乱有序。
它们掌心大小,奶白色,咖色,无色,形状似珍珠,葫芦,海螺,辣椒......奇形怪状,漂亮神奇。
有孔虫,是带壳的海洋单细胞动物,因壳内有“口孔”相通而得名。
实习一 有孔虫
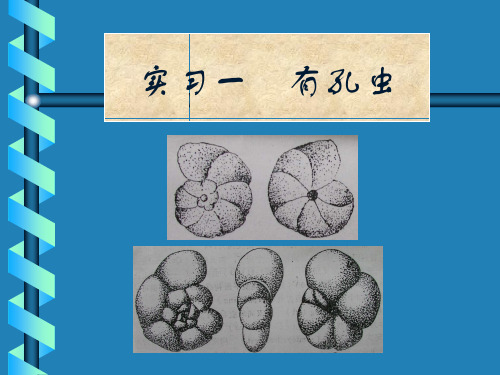
Quinqueloculina(五块虫)
侧视椭圆形,房室按72º角 度绕卷生长,外观只见最 后的5个房室,壳口具棒形 或丁字形齿,J-R 。
瓶虫 Lagena
单室,瓶状, 壳口在末端,圆形, J-R 。
箭头虫 Bolivina
楔形,双列式,壳口 扣眼状,K2-R 。
葡萄虫 Uvigerina
拟抱球虫Globigerinoides :除脐部口外,缝合线上有次壳口,N-R
圆辐虫 Globorotalia
螺旋,双凸或背平背微凹,壳缘具棱 脊,壳口底弧形,于基部,脐封闭, E-R 。
圆球虫 Orbulina
早期抱球虫式,后期被球形的末 室所包围,呈圆球形,壳面有许 多小圆孔,N-R 。
双盖虫 Amphistegina
螺旋壳,壳面光滑,腹面具补充房室,缝 合线强烈弯曲,E-R 。
钙虫 Calcarina
螺旋壳,壳面粗糙,具体6-30个粗刺, K2-R 。
实习一 有孔虫
实习内容
一、观察有孔虫的有关构造
1.壳口构造
2.壳壁微孔及网纹构造
3.多房室壳的排列方式
1)平旋式,房室绕初房在同一平面内旋转排列,壳缘两侧对称, 希望虫(Elphidium)
2)螺旋式 A.平凸形 B.螺旋双列,串球虫(Textularia) 3)绕旋式 A.三玦虫(Triloculina)
三列式,塔形或纺锤形,房室膨 胀,壳口末端,具唇,E-R 。
希望虫 Elphidium
平旋内卷,透镜状, 房 室多,缝合线具一 列壁间 桥和小凹坑, E-R 。
幼体虫 Pullenia
平旋内卷,近球形,末圈4-5室,两侧 对称;壳面光滑; 脐封闭; 壳口低, 新月状. K2-R
温度和光周期对单细胞有孔虫生命周期的影响

温度和光周期对单细胞有孔虫生命周期的影响单细胞有孔虫是一种常见的原生生物,它们生活在水中,甚至可以在湖泊表面形成较大的“水花”。
这种微生物虽然小,但是却有着复杂的生命周期,其生命周期的不同阶段和形态都受到外界的环境因素的影响。
其中,温度和光周期是两个最主要的外界环境因素之一。
本文将从这两个方面探讨温度和光周期对单细胞有孔虫生命周期的影响。
温度对单细胞有孔虫生命周期的影响单细胞有孔虫是一种活跃的微生物,它们的生命活动过程中需要一定的能量支撑。
而温度是影响生物体代谢的重要因素之一,对于单细胞有孔虫而言也不例外。
研究表明,不同的温度下,单细胞有孔虫的生命周期会有所不同。
低温下,单细胞有孔虫生命周期较长,而且其生长速度也会明显减缓。
这是由于低温可以降低单细胞有孔虫代谢的速度,导致其能量供应减少,生命周期变长,生长速度变慢。
而在高温下,单细胞有孔虫生命周期会变短,其生长速度也会加快。
这是由于高温可以提高单细胞有孔虫代谢的速度,促进其能量供应,从而使其生命周期变短,生长速度加快。
需要注意的是,过低或过高的温度都会对单细胞有孔虫产生不良影响。
例如,在过低的温度下,单细胞有孔虫不仅生存状况会受到威胁,甚至还可能死亡。
而在过高的温度下,单细胞有孔虫代谢会过度剧烈,导致其受到严重的伤害或死亡。
因此,维持适宜的温度是促进单细胞有孔虫正常发育和生命活动的重要因素之一。
光周期对单细胞有孔虫生命周期的影响光周期是另一个影响单细胞有孔虫生命周期的因素。
这是由于光周期会影响单细胞有孔虫的代谢,从而进一步影响其生命周期。
不同的光周期对单细胞有孔虫生命周期的影响,主要表现在以下方面:首先是光照强度的影响。
在光照强度不变的情况下,光周期的变化会直接影响单细胞有孔虫的代谢和生命周期。
例如,在短日照条件下,单细胞有孔虫的周期会变短,而在长日照条件下,单细胞有孔虫的周期会变长。
这是由于不同的光周期会影响单细胞有孔虫细胞内的代谢速率,从而使得其生命周期受到影响。
贵州惠水县雅水剖面有孔虫演化研究及意义

贵州惠水县雅水剖面有孔虫演化研究及意义【摘要】本研究旨在探讨贵州惠水县雅水剖面中孔虫的演化历史及其意义。
通过采用多种研究方法,我们获得了丰富的研究结果,揭示了孔虫在不同时期的种类丰富度和数量变化。
研究表明,孔虫在雅水剖面中的分布与环境变化密切相关,反映了古环境演化的过程。
我们讨论了孔虫演化的意义,认为其研究有助于理解古生态环境的演变,为地质学和古生物学领域提供了重要的参考资料。
未来,我们将进一步深入研究孔虫的演化机制,以期更好地揭示其在地质历史中的作用。
总结来看,本研究为贵州古生物学研究提供了重要参考,具有一定的学术价值和意义。
【关键词】孔虫演化研究、贵州惠水县雅水剖面、研究背景、研究意义、研究方法、研究结果、研究讨论、演化意义、意义展望、研究结论、展望未来研究、总结评价。
1. 引言1.1 研究背景:研究背景部分的内容应包括:有孔虫演化研究的历史沿革、贵州惠水县雅水剖面的地质背景和研究现状、近年来关于有孔虫演化的研究进展等。
可以介绍有孔虫在古生物学领域的重要性,以及对地质学、古气候学等领域的重要意义。
需要详细阐述有孔虫在古环境重建、古气候研究、古海洋学等方面的应用,以及对人类社会发展的启示和促进作用。
部分的内容应详尽准确,有条理且具体。
1.2 研究意义本研究对于揭示贵州惠水县雅水剖面中孔虫的演化历史具有重要意义。
孔虫是一类古老的微生物,其在地质历史上具有极高的生物地层学价值。
通过对雅水剖面中孔虫的研究,可以帮助我们更好地理解地球生物演化的过程和规律。
孔虫在古环境和古气候重建中扮演着重要角色,通过对孔虫群落结构和数量变化的分析,可以揭示当地古环境的特征和演化过程。
雅水剖面中孔虫的研究还可以为该地区地质构造和地质历史的探讨提供重要依据。
通过深入研究雅水剖面中孔虫的演化过程,可以为贵州地质史的研究提供新的线索和证据。
本研究的意义不仅在于扩展了我们对孔虫演化历史的认识,同时也为贵州地质史和古生物学领域的研究做出了贡献。
cercozoa分类

cercozoa分类
有孔虫门(Foraminifera)属于原生生物界,是一种变形虫状的原生生物,具有网状假足和幼细线状细胞质,能够形成动态的网。
有孔虫门拥有一个或多个室的外壳,大部分个体大小小于一毫米,但部分较大,纪录中最大的样本大小达19厘米。
丝足虫门(Cercozoa)属于原生生物界,是一类具有复杂形态的原生生物,与有孔虫门和放散虫门(Radiolaria)关系密切。
丝足虫门具有多种形态,包括球形、螺旋形、长形等,通常具有丝状或网状的伪足。
因此,有孔虫门和丝足虫门都属于原生生物界,但它们属于不同的分类群。
常见有孔虫中英文对应表_作者_壳质成分

拉丁文名中文名Ammobaculites agglutinans (d'Orbigny)胶结砂杆虫Ammomarginulina tenerissima Shchedrina极娇砂缘虫Ammonia beccarii vars.毕克卷转虫变种Ammonia compressiuscula Brady压扁卷转虫Ammonia convexidorsa Zheng凸背卷转虫Ammonia flevensis (Hofker)显脐卷转虫Ammonia granuloumbilica Zheng粒脐卷转虫Ammonia ketienziensis Ishizaki结缘寺卷转虫Ammonia maruhasii (Kuwano)丸桥卷转虫Ammonia pauciloculata (Phleger and Parker)少室卷转虫Arenoparella asiatica Polski亚洲砂壁虫Astrononion tasmanensis Carter塔斯曼星九字虫Bolivina robusta Brady强壮箭头虫Brizalina seminuda (Cushman)半裸判草虫Brizalina striatula (Cushman)条纹判草虫Buccella frigida Cushman冷水面颊虫Buccella tunicata Ho,Hu and Wang覆盖面颊虫Bulimina aculeata d'Orbigny棘刺小泡虫Bulimina marginata d'Orbigny具缘小泡虫Bulimina sp.小泡虫未定种Buliminella elegantissima d'Orbigny最美微泡虫Cassidulina carinata Silvesri具棱小盔虫Cavarotalia annectens (Parker and Jones)同现孔轮虫Cibicides sp.面包虫未定种Cribrononion frigidum (Cushman)冷水筛九字虫Cribrononion porisuturalis Zheng孔缝筛九字虫Cribrononion subincertum (Asano)亚易变筛九字虫Cribrononion sp.筛九字虫未定种Dentalina communis d'Orbigny普通齿形虫Dentalina decepta (Bagg)隐纹齿形虫Dentalina sp.齿形虫未定种Edentostomina cultrata (Brady)注意无齿虫Eggerella advena (Cushman)异地伊格尔虫Elphidiella kiangsuensis (Ho,Hu and Wang)江苏小希望虫Elphidium advenum (Cushman)异地希望虫Elphidium asiaticum Polski亚洲希望虫Elphidium hispidulum Cushman粗糙希望虫/茸毛希望虫Elphidium jenseni (Cushman)扁肾希望虫Elphidium limpidum Ho,Hu and Wang清晰希望虫Elphidium magellanicum Heron-Allen and Eerland 缝裂希望虫Elphidium simplex Cushman简单希望虫Elphidium subcrispum Nakamura亚波纹希望虫Epistominellla naraensis (Kuwano)奈良小上口虫Esosyrinx curta (Cushman and Ozawa)短小内管虫Fissurina aradasii Seguenza瓜子缝口虫Fissurina crebra(Matthes)常见缝口虫Fissurina cucurbitasema bispinata Ujiie双刺葫芦缝口虫Fissurina laevigata Reuss光滑缝口虫Fissurina lucida (Williamson)明亮缝口虫Fissurina sp.缝口虫未定种Fissurina valida(Matthes)坚强缝口虫Florilus atlanticus (Cushman)大西洋花朵虫Globocassidulina sp.球盔虫未定种Guttulina sp.小滴虫未定种Hopkinsina pacifica Cushman太平洋霍氏虫Lagena clavata Williamson棒形瓶虫Lagena distoma(Seguenza)扭转瓶虫Lagena pliocenica Cushman and Gray上新瓶虫Lagena purlucida (Montagu)透明瓶虫Lagena sp.瓶虫未定种Lagena spicata Cushman and McCulloch尖底瓶虫Lagena substriata Williamson亚线纹瓶虫Lagena wiesneri Parr维氏瓶虫Lenticulina sp.透镜虫未定种Massilina sp.块心虫Miliamina sp.砂粟虫未定种Nonion akitaense Asano秋田九字虫Nonion belriodgense Barbat and Johnson贝尔岭九字虫Nonion boueanum (d'Orbigny)波义九字虫Nonion glabrum Ho,Hu and Wang光滑九字虫Nonionella stella Cushman and Moger星小九字虫Nonionella jacksonensis Cushman杰克逊小九字虫Nonionella sp.小九字虫未定种Pararotalia nipponica (Asano)日本仿轮虫Paratrochammina sp.仿砂轮虫未定种Protelphidium tersum Ho,Hu and Wang洁净先希望虫Protelphidium tuberculatum (d'Orbigny)具瘤先希望虫Psedogyroidina sinensis (Ho,Hu and Wang)中华假圆旋虫Psedononinella sp.假小九字虫未定种Pseudoeponides japonicus Uchio日本假上穹虫Pseudononion minutum Zheng小假九字虫Pseudononionella variabilis Zheng多变假小九字虫Quinqueloculina akneriana rotunda (Gerke)圆短五玦虫Quinqueloculina bellatula Bandy整洁五玦虫Quinqueloculina lamarckiana d'Orbigny拉马克五玦虫Quinqueloculina seminula (Linné)半缺五玦虫Quinqueloculina seminulangulata Mclean半角五玦虫Quinqueloculina sp.五玦虫未定种Quinqueloculina sp.五玦虫未定种Quinqueloculina venusta Karrer悦目五玦虫Quinqueloculina subarenaria Cushman半砂五玦虫Rectobolivina raphana(Parker and Jones)萝卜直箭头虫Rectoelphidiella lepida Ho,Hu and Wang精美直小希望虫Rosalina bradyi (Cushman)布拉德玫瑰虫Schackoinella globosa (Millett)球形小夏克虫Sigmoilina tenuis (Czjzek)窄室曲形虫Spiroloculina bohaiensis Zheng渤海抱环虫Spiroloculina indica Cushman and Todd印度抱环虫Textularia foliacea Heron-Allen and Earland叶状编织虫Trifarina angulosa (Williamson)角状三粉虫Triloculina pentagonalis Wang五角三玦虫Triloculina rectilocula Zheng直室三玦虫Triloculina sp.三玦虫未定种Triloculina trigonula (Lamarck)三角三玦虫Trilouclata tricarinata d'Orbigny三棱三玦虫Trochammina inflata (Montagu)胖砂轮虫Uvigerina canariensis d'Orbigny卡纳利葡萄串胶结质胶结质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质胶结质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质瓷质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质胶结质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质玻璃质瓷质瓷质瓷质瓷质瓷质瓷质瓷质瓷质瓷质玻璃质玻璃质玻璃质玻璃质瓷质瓷质瓷质玻璃质瓷质瓷质瓷质瓷质瓷质胶结质玻璃质。
显微镜下观察有孔虫实验报告

显微镜下观察有孔虫实验报告
实验目的:通过显微镜观察有孔虫的形态结构,了解其特征和生活习性。
实验材料:
1.显微镜
2.盖玻片
3.移液管
4.有孔虫标本
实验步骤:
1.将有孔虫标本放置在盖玻片上。
2.使用移液管将一滴透明液体滴在标本上,以保持标本湿润。
3.将盖玻片放置在显微镜的玻片夹上。
4.逐渐调节显微镜的焦距,直到能够清晰地观察到有孔虫。
5.仔细观察有孔虫的形态结构,包括身体的外形、长度、颜色等特征。
6.注意观察有孔虫的运动方式、摄食方式等生活习性。
实验结果:
通过显微镜观察,我们发现有孔虫身体呈椭圆形,长度约为0.1-0.2毫米,颜色呈透明或淡黄色。
它们具有许多细小的孔洞分布在身体表面,这些孔洞用于呼吸和加速水流进出体内。
有孔虫以游动的方式在水中移动,摄食方式主要通过细胞表面的纤毛将食物颗粒吸
附到口部。
实验结论:
通过显微镜下的观察,我们成功地观察到了有孔虫的形态结构和生活习性。
有孔虫具有特殊的身体结构和运动方式,适应了水生环境中的生活。
这次实验对我们了解有孔虫的生物学特征和生态习性有一定的帮助。
有孔虫实验报告范文

有孔虫实验报告范文实验报告:有孔虫的观察1.引言有孔虫是一种微小的海洋生物,属于原生动物门,具有装甲和伪足,广泛分布于海洋中。
它们的身体结构透明,可以通过显微镜观察和研究。
在本次实验中,我们将通过观察有孔虫的外部结构、移动方式和生长过程,了解它们的基本生物学特征。
2.材料和方法2.1实验材料:-显微镜-有孔虫标本-玻片和盖玻片-盐水和蒸馏水-滴管和显微镜玻璃片2.2实验步骤:-在显微镜玻璃片上滴一滴盐水,将有孔虫标本移入盐水中。
-用显微镜观察有孔虫的外部结构并绘制观察图。
-用显微镜观察有孔虫的移动方式并记录。
-将有孔虫标本移入蒸馏水中,观察其对不同环境的适应情况。
3.结果3.1外部结构通过显微镜观察,我们发现有孔虫的身体呈椭圆形,表面覆盖有装甲。
有孔虫的装甲特征独特,装甲上分布有许多大小不一的孔洞,这也是它们被称为有孔虫的原因。
此外,我们还观察到有孔虫的细长伪足,伪足的运动可以通过这些细长的结构来实现。
3.2移动方式在观察有孔虫的移动方式时,我们发现它们通过伪足的运动来移动。
伪足可以延伸和收缩,通过伪足的延伸和收缩,有孔虫能够在水中游动。
在移动过程中,有孔虫的伪足会不断地伸出和缩回,这种运动方式使它们能够在微小空间中自由移动。
3.3对环境的适应在将有孔虫标本移入蒸馏水中后,我们发现它们对环境的适应能力非常强。
有孔虫在蒸馏水中能够保持良好的生存状态,并继续进行正常的运动。
这表明有孔虫具有良好的耐受力,能够适应不同的环境变化。
4.讨论通过本次实验,我们对有孔虫的外部结构、移动方式和对环境的适应情况有了初步了解。
我们发现有孔虫具有独特的装甲结构和细长的伪足,这些结构使它们能够在海洋中自由游动。
此外,我们的实验结果还表明,有孔虫具有较强的环境适应能力,能够在不同的环境中生存和繁衍。
然而,我们的实验还存在一些限制。
首先,由于有孔虫体积较小,观察过程中可能存在一定的视觉误差;其次,本实验只观察了有孔虫的外部结构和运动方式,对于其生长和繁衍过程还有待深入研究。
有孔虫培育方法

有孔虫培育方法有孔虫(Foraminifera)是一类生活在海洋和淡水环境中的微型单细胞生物。
它们的身体被一个有孔的壳所保护,这个壳通常具有复杂的纹饰和结构。
有孔虫在地质学、生物学和环境科学等领域具有重要的研究价值。
为了进行有孔虫的研究,科学家们需要培育这些微生物,以便进行实验和观察。
下面将介绍几种常见的有孔虫培育方法。
第一种方法是使用海水培养基。
有孔虫是海洋生物,因此海水是它们生长和繁殖所必需的环境。
科学家们可以收集海水并加入适当的营养物质,如有机碳源和氮源,制备海水培养基。
然后,将有孔虫样本放入培养基中,并提供适当的温度、pH值和光照条件。
有孔虫会在培养基中生长和繁殖,科学家们可以观察它们的行为和形态变化。
第二种方法是使用人工合成培养基。
有时候,科学家们会根据有孔虫的营养需求和生长条件,自行合成培养基。
他们会根据已有的研究和实验数据,确定培养基的成分和浓度。
通过调整培养基的pH 值、温度和光照条件,科学家们可以模拟特定的环境,从而促进有孔虫的生长和繁殖。
第三种方法是共培养法。
有些有孔虫是共生生物,它们与其他生物(如藻类或细菌)之间存在共生关系。
科学家们可以将这些共生物种一起培养,以促进有孔虫的生长和繁殖。
共培养法可以更好地模拟自然环境中的生态系统,提供更逼真的研究结果。
值得注意的是,有孔虫是非常微小的生物,其培养需要一定的技术和设备支持。
科学家们通常使用显微镜和特殊培养器具来观察和操作有孔虫。
他们还需要严格控制培养条件,如温度、光照强度和营养物质浓度,以确保有孔虫能够正常生长和繁殖。
有孔虫的培育方法还在不断发展和改进中。
随着科学技术的进步,科学家们可以更好地了解有孔虫的生态特征和生理过程。
这将有助于揭示有孔虫在地球历史和环境变化中的作用,以及它们对气候变化和海洋生态系统的响应。
有孔虫培育方法的研究和应用,将为我们认识自然界的微观生物提供更多的线索和信息,推动科学的进步和发展。
有孔虫演讲稿

有孔虫演讲稿
亲爱的老师、同学们:
大家好!
我今天微大家演讲的题目是《有孔虫》。
浮游有孔虫是一群古老的生物,但是很多人对它们知之甚少。
黄舒雯同学向我们介绍了这样一群从侏罗纪时代就存在的生物:它们很小却分布广泛、它们生命力顽强,对水温十分敏感,所形成的化石对海洋环境研究具有重要意义。
从世界迈入近代史以来,工业排放量不断增加,近些年来,“气候变暖”一词也频繁被提及。
然而每到冬天,在寒风中冻得瑟瑟发抖的同学们,是否常常想:现在说是气候变暖了,还是这么冷,那么以前的气候是怎样的呢?
是啊,以前的天气有多冷呢?我们要借助一些化石、遗迹来推断。
可是几千年文明发展,无数刀耕火种、攻城掠地,哪片土地上能找到足够古老的、能反映气候变化的东西呢?还是有办法的,就从海里找嘛。
海洋,人类近代才开始利用的蓝色宝藏,不负众望,确实藏有远古气候的密码——有孔虫。
我的演讲到此结束,谢谢大家。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Foraminifera
微体古生物法:
• 微体古生物法是古海洋学最主要的研究手段。有孔虫、放射虫、 硅藻、颗石藻等微体或超微体生物的生活,主要受海水深度、 温度、盐度、浊度、营养盐以及水体运动等各种物理化学条件 的控制。这些要素变化的信息便记录在生物个体、生物组合、 分异度等特征上,因此海洋生物是海洋环境的灵敏标志。
我国东海有孔虫组合、丰度等与深度的关系
19
•
3、盐度 盐度是控制有孔虫组和类群的另一个重要因素,对浮游有孔 虫的影响尤为明显。按照有孔虫对盐度的适应范围可分为广盐分子和 窄盐分子。盐度对有孔虫的影响主要表现为下列几个方面:
•
•
1)化石群的分异度:一般情况下,盐度越偏离正常,化石分异度就越 低,常表现为丰度高而分异度低。
60
120 140 160 180 160 140 120 100 80
60 40 18
2、深度 有孔虫组合及类群的分布与水深之间的关系最为明显,可归纳如下: 1、有孔虫的分异度和丰度随水深度的加大而增加;2、浮游有孔虫的数量随着离 岸距离及深度的增加而增加;3、在滨海和陆棚地带,随着水体深度的增加,胶 结壳类型逐渐减少,钙质壳类型逐渐增多。 部 位 水深(m) 河口 <20 内陆棚 <50 中陆棚 50-100 外陆棚 100-150 假棱串珠 虫-珍珠面 包虫组合 n×102n×103 70-90% 陆 坡 150-700 海槽 700-2000 瓶虫类 组合
外质 丝状的伪足
内质
二、有孔虫——生活环境与生活方式
• 有孔虫的生活环境:现代有孔虫绝大多数生活在正常海域,少 数生活在泻湖、沼泽、河口等滨海边缘环境或残留海水等半咸 水水域。
有孔虫的生活方式:大多数有孔虫营底栖生活,少数营浮游生 活。底栖有孔虫可以分为小有孔虫和大有孔虫,小有孔虫直径 一般为2.5mm以下,大有孔虫一般大于2.5mm。
•
2、有孔虫从始新世进入全盛时期,成为重要的造岩生物。以 致大半个欧洲都以“货币虫灰岩”作为“下第三系”的同义名, 同时也出现了峰巢虫石灰岩,小粟虫石灰岩等有孔虫灰岩。
有孔虫 各亚目 及超科 地史分 布图
假几丁质壳 胶结壳 钙质微粒壳 似瓷质壳 钙质透明多 孔放射状或 钙质微孔壳
类
别
Є O S D C P T
2)有孔虫壳体发生畸形,在盐度偏高的不正常海水中有孔虫壳体常发 生畸形。
•
•
3)形成广盐性或特殊的半咸水组合
4)海相与陆相化石共生盐度的变化主要受海陆过渡带的控制。即由于 淡水的注入或障壁物的形成改变水体的盐度。因此,在有孔虫组合中 出现某些陆相化石应是近岸过渡带的指示,这种有孔虫组合很可能会 显示出不正常盐度的环境特征。在陆海过渡地区,尤其在三角洲前缘, 是重要的潜在的生油和储油区。因此,过渡相有孔虫组合的辨认和识 别对于寻找石油具有重要的意义。 4、基底 基底对浮游有孔虫一般不产生影响,但对底栖有孔虫影响较 大。
• • •
五)第三纪 有孔虫发展的第三个繁盛时期 继白垩纪之后,有孔虫在古新世经历了短暂的停滞,从始新世 又开始了第三个繁盛时期,主要特点为: 1、底栖有孔虫中许多具白垩纪特色的科属完全绝灭,大量的 新类型开始出现,如古新世绝灭的圆片虫科(Orbifoididae) 等,新产生的如轮虫科(Rotaliporidae)。
•
•
100 120 140 160 180 160 140 120 100
80
60
40
60
60
太平洋浮游有 孔虫的分布图
北洋 亚北极海 40 迁移带 40
中央
20
北亚热带
20
赤道系中央
热带
0
0
生活群集带的界限 20 南中央 40 南亚热带 40 20
遗核群集带的界限 60
亚南极海
亚南极海 亚南极海
• • •
(二)C-P有孔虫发展的第一个繁盛时期 C-P是古生代有孔虫最繁盛的时期,出现了许多划分对比地层 的标准化石,主要特征为: 1、钙质微粒壳类型达到极盛,内卷虫超科(Endo-Thyrian) 和纺缍虫超科(Fusulinacea)在这一时期极大的发展,而绝 大多数灭绝于P末期,构成了这一时期的标准化石。 2、曲杖虫超科(Lituolacea)的产生与兴旺,胶结壳的曲杖虫 超科开始出现于C,并立即兴旺超来,这个超科的10个科有一 半始于C、P。 3、小粟虫目(Miliolina)从C纪开始出现。 4、钙质多孔壳类型开始萌芽,如节房虫(Nodosariacea)。
•
图片来源: F. Lejzerowicz等,《生物学快报》
三、有孔虫——壳的构造
• 有孔虫其原生质分泌壳质形 成壳子,或分泌胶结物粘结 其它颗粒形成壳子,壳子中 空,称为房室或壳室。 房室顶端有一圆形或其它形 状的开口叫口孔。多个房室 组成的壳,其最早形成的, 最小的房室叫做初房,以后 继续分泌壳质形成第一房 室……最后一个房室叫终室, 分隔两个房室的壁称为隔壁, 隔壁是壳壁向前沿伸时向内 转折的部分,隔壁与壳壁相 交的线称为缝合线。 口孔位于终室的顶端,其形 状变化多样,口孔周围的壁 称为前壁。
•
• •
• •
(三)T-J有孔虫的复苏时期 古生代末期生物大规模绝灭(mass extinction)事件对有孔虫产 生了极大的影响,纺缍虫全部绝灭于P末,内卷虫超科百分之九 十以上的科绝灭,所以T-J是绝灭事件以后的一个复苏时期,其特 点如下: 1、曲杖虫超科和小粟虫超科,新产生了40%的科,自∈纪就 开始出现的瓶虫超科(Lagynacea)和砂盘虫超科 (Ammodiscacea)的发展远比不上曲杖虫和小粟虫超科的发 展。 2、钙质多孔壳的超科开始大量出现,产生了六个超科,而且 有孔虫的分异度开始回升。有重要意义的是新产生的双口虫超 科(Duostominacea)和抱球虫超科(Globigerinacea)。其 中双口虫超科出现于T,而绝灭于中侏罗世,演化快,可视为 标准化石。而且具有演化方面的意义,许多后来的钙质透明壳 都是由他演化而来。抱球虫超科是有孔虫中唯一的浮游类型, 它的出现是有孔虫演化史上的一件大事,有着重要的地质意义。
•
•
四、有孔虫——地史分布
•
• •
有孔虫自∈纪开始出现,一直延续到现在,在其发展演化的过 程中曾几经兴衰,大致可划分为五个重要的发展阶段:
(一)∈-D有孔虫发展的初期阶段 在早古生代有孔虫化石比较贫乏,前寒武纪岩石中缺乏令人信 服的有孔虫化石证据。奥陶纪前的有孔虫都是一些简单的类型, 如单房室、树枝状、管状等。这是有孔虫的发生阶段。S-D是 有孔虫的初期发展阶段,此时胶结壳的类型开始繁盛,并开始 出现了一些钙质微粒壳类型。总的来看,此阶段的有孔虫以伪 几丁质壳和胶结壳为主,类型原始。
•
• •
(二)有孔虫的生态控制因素
有孔虫所生活的海洋环境相当复杂,控制因素相当之多,下面 主要介绍几个重要的控制因素。 1、温度 温度是影响有孔虫的重要因素,不仅制约着其地理 分布,而且对它的形态、生存、繁殖等都有巨大的影响。深度 不同的海底水温不同,底栖有孔虫的类型就不同;纬度不同的 洋面水的温度不同,浮游有孔虫的组合也有差异。 水深100m以上,水温季节性变化大,为变温带,此带主要生 存的是广温性属种,如卷转虫(Ammonia)、希望虫 (Elphidium)、微泡虫(Buliminella)等。 水深在100米以下的恒温带,主要生存的是窄温性属种,如盔形 虫(Cassidulina)、小泡虫(Bulimina)、葡萄虫 (Uvigerina)、深海管虫(Bathysiphon)等。
J K E
N Q
ALLOGROMIINA(奇杆虫亚目) Lagynacea(瓶形虫超科) TEXTULARIINA(串珠虫亚目) Ammodiscacea(砂盘虫超科) Lituolacea(曲杖虫超科) ENDOTHYRINA(内卷虫亚目) Parthuramminacea(拟砂户虫超科) Endothyracea(内卷虫超科) FUSULININA(蜓亚目) Fusulinacea(纺锤虫超科) Verbeekinacea(费伯克蜓超科) MILIOLINA(小粟虫亚目) Miliolacea(小粟虫超科) ROTALIINA(轮虫亚目) Nodosariacea(节房虫超科) Spirillinacea (盘旋虫超科) Robertinacea (罗伯特虫超科) Buliminacea (小泡虫超科) Discorbacea (圆盘虫超科) Duostominacea (双口虫超科) Nonionacea (诺宁虫超科) Cassidulinacea (盔形虫超科) Rotalicea (轮虫超科) Globigerinacea (抱球虫超科) Orbitoidacea (圆片虫超科) Carterinacea (伞形虫超科)
•
•
• •
•
抱球虫软泥 主要由浮游性有孔虫,尤以抱球虫遗骸组成,浮游有孔虫亦含 大量颗石藻片、放射虫、硅藻以及翼足虫等。 分布面积占海洋底总面积的40%以上。钙质成分在水深不到 1000米的地带达90%,4000~5000米的地带为30%左右。主 要分布在太平洋南半部、大西洋和印度洋的热带和亚热带海域。 将来可能将其作为石灰原料开采。
•
•
• • • • • •
(四)白垩纪,有孔虫发展的第二个繁盛时期 白垩纪是有孔虫全面繁盛的起点,所有的超科至此已全部出现, 主要特点为: 1、新超科大量出现,仅钙质多孔壳的就出现12个科。 2、胶结壳的曲杖虫超科达到极盛,新产生了两个新科和七个 亚科。 3、抱球虫超科,该超科共有八个科,其中五个科出现于白垩 纪。 4、大型底栖有孔虫类群和特有浮游类群的出现使白垩纪有孔 虫呈现出独特的面貌,胶结壳的有圆锥虫科(Orbifolinidae), 似瓷质壳的有蜂巢虫科(Alveolinidae),钙质多孔壳的假圆 片虫科(Pseudobifoididae)和货币虫科(Nummulitidae)为 代表。