发育生物学第四章原肠胚形成课件PPT
发育生物学神经系统发育课件

神经系统的发育和再生: 神经细胞的发生和增殖
细胞分化
细胞迁移
形成神经回路
神经活动 (正常)
修复和再生 (外伤&疾病)
第一节
神经管的发育
➢ 神经胚发育概述 ➢ 神经管的发育 ➢ 神经嵴的衍化 ➢ 小结
一.神经胚发育概述
神经系统的组织发生过程
中枢神经系统 神经系统
周围神经系统
神经嵴 神经胚
神经管 原肠胚外胚层
前脑泡
端脑 端脑泡腔 两个侧脑室
前N孔闭合
间脑前脑泡腔 第三脑室
脑 泡 中脑泡 中脑
Brain vesicle
后脑
菱脑泡 末脑 (后)
菱脑泡腔
背:四叠体 腹:大脑脚 中脑泡腔:中脑 导水管 脑桥、小脑
延髓
第四脑室
脑的发育(解剖学角度)
● 脑发育与脊髓比较
1. 脊髓的神经管壁形成典型的节段性,其结 构基本上一致;而脑部的分化速度则因部 位而不同,其结果在脑部出现了多个脑 泡—成体脑的各部结构;
2. 神经嵴细胞的多能性
表现: ➢周围神经系统的神经节和神经胶质细胞 ➢肾上腺髓质的嗜铬细胞 ➢黑色素细胞 ➢滤泡旁细胞 ➢颈动脉体ⅰ型细胞 ➢间充质细胞→头颈部的部分骨、软骨、肌肉及结缔组织
小结
神经胚形成:
(A) 神经板期 (B) 神经褶期 (C) 神经管期
神经管的形成:受分子调控的过程
神经管与神经嵴:一个过程的两个结果
神经胚的发育-神经板期
神经胚的发育-神经褶期Fra bibliotek神经胚的发育-神经管期
二.神经管的发育
(一)神经管的形成 (二)神经管细胞的增殖 (三)神经管细胞的迁移 (四)神经管的闭合 (五)神经管的分化
形成身体计划:原肠胚是如何形成

形成身体计划:原肠胚是如何形成生命里最重要的时刻,不是出生、结婚或死亡,而是原肠胚的形成。
——路易斯·沃尔伯特在胚胎发育早期,一开始只有简单的细胞分裂,没有发生其他事,之后突然发生的转变让每层细胞都变得各不相同,这也就促生了横跨在满是液体的空间中的双层胚盘。
这样的结构与能够被我们辨识为人的形态还相距甚远。
如果一个理智的人被告知:这对胚盘将变成动物,他很可能会想到水母之类的物种,因为至少水母的伞状体和胚盘具有类似的辐射对称结构:顶端和底部有差异,但是在垂直方向上可以找出无数条对称轴线。
然而,胚胎此刻已经为迅疾变化做好了准备,两天内进行大规模重组。
到时候,几乎每个人都会承认它基本是个婴儿了。
以上就是原肠胚的形成过程。
理解原肠胚形成的方法之一,是首先细致地了解身体通过这个过程会形成的雏形结构,然后再去详细了解原肠胚转化成身体中的各个事件。
这种思路或许是最容易理解的,但是可能会传达出一个错误的信息,导致人们认为这些细胞之前就在某种程度上已经知晓了自己将要形成的身体结构,它们之后便向着这个方向努力。
事实上,发育并不依赖细胞是否掌握了相关的复杂信息,因为即使对我们拥有亿万个细胞的大脑而言,这些信息仍然复杂到无法轻易掌握。
发育靠的是每个细胞对自身周围环境简单、自主的反应。
原肠胚第一个能(在实验动物中)检测到的变化发生在下胚层。
胚盘中部的细胞开始启动新的基因,包括一种合成DNA结合蛋白的特定基因:Hex。
参与触发这些变化的物质和具体的位点尚不为人所知:一种可能是,下胚层所有的细胞都为出现这种变化做好了准备,而大多数细胞都被一种信号蛋白抑制了,这种蛋白质在包围下胚层边缘的支持组织中生成。
而位于胚盘中心的细胞因为远离产生抑制蛋白的地方,所以能不受影响,从而可以启动Hex基因。
这个抑制机制的运作是科研人员的猜想,然而确实有明确证据显示Hex基因启动了表达。
这些产生Hex蛋白的细胞会从它们周围的细胞中挤出一条路,聚集到下胚层外缘的一个点上。
《发育生物学》ppt课件(2024)

基因编辑技术
类器官培养技术
运用CRISPR/Cas9等基因编辑工具,对生 物体的基因组进行精确修饰,研究基因功 能和调控机制。
2024/1/30
通过模拟体内微环境,培养具有类似器官结 构和功能的类器官,用于疾病模拟、药物筛 选和再生医学等领域。
29
未来发展趋势预测
跨学科交叉融合
发育生物学将与遗传学、细胞生物学、生 物医学工程等学科交叉融合,共同推动生
2024/1/30
6
02
细胞分裂、分化与胚胎发育
Chapter
2024/1/30
7
细胞周期与有丝分裂
细胞周期
指连续分裂的细胞从一次分裂完 成时开始,到下一次分裂完成时 为止所经历的全过程,包含DNA 合成前期、DNA合成期、DNA
合成后期和有丝分裂期。
有丝分裂
一种真核细胞分裂产生体细胞的 过程,特点是有纺锤体染色体出 现,子染色体被平均分配到子细
02
配子形成
在减数分裂过程中,染色体只复制一次,而细胞分裂两次。减数分裂的
结果是,成熟生殖细胞中的染色体数目比原始生殖细胞减少一半。
2024/1/30
03
配子的种类
根据染色体的组合不同,可以产生不同种类的配子,增加了后代的遗传
多样性。
9
胚胎发育过程及调控机制
胚胎发育过程
从受精卵开始,经过卵裂、桑葚胚、 囊胚、原肠胚与组织器官形成等阶段 ,最终发育成为完整的胎儿。
2024/1/30
3
课程目标与要求
01
掌握发育生物学的 基本概念、原理和 研究方法
02
了解发育生物学的 最新研究进展和前 沿动态
03
能够运用所学知识 分析和解决发育生 物学领域的实际问 题
原肠胚形成

发育生物学的模式生物
海胆 海胆属无脊椎动物中的棘皮动物门,是一类古
老的后口动物。 线虫 1974年分子生物学家Syndey Brenner开始加强
了对于线虫基因的研究工作,事实证明,线虫是 对于真核细胞生物的发育遗传、细胞生物、神经 生物以及基因结构进行研究的极好实验材料,业 已成为发育生物学的模式动物之一。
1)全裂 卵子彻底分裂成单个细胞。按照第一 代子细胞的大小,全裂又进一步划分为均裂和非均 裂。
2)不全裂 受精卵在卵裂初期发生不完全卵裂 。
全裂包括有下面几种类型: 1)辐射型卵裂:形成8个卵裂球后,以后分裂
均为辐射型排列。如棘皮动物,腔肠动物等 2)螺旋型卵裂:第三次分裂不与赤道板方向
平行,而呈450 角,结果动物极卵裂球位于两植物 极卵裂球之间,此后分裂同此。如螺、蚌,纽虫、 软体动物等。
卵裂是一系列的细胞分裂
受精卵经过一系列迅速分裂活动,形成许多的 卵裂球(卵裂所形成的细胞)的过程。即(卵裂) 活动。在卵裂过程中,与一般细胞分裂活动不同的 是,细胞数量迅速增加,越来越多,但其体积和所 包含的物质不增加。
随着卵裂的进行,分裂的细胞越来越小,总的 体积大致不变。
卵裂的类型和方式
基于卵细胞的空间结构特点和本身所含卵黄的 多少,卵裂分成两种类型:
发育生物学:第四章 原肠胚形成

(二)原肠作用中的主要细胞迁移方式
➢ 外包(epiboly): 以整片细胞(通常为外胚层细 胞)沿胚胎表面扩展,包裹胚体的深层细胞。
➢ 内陷(invagination): 指外层细胞成片地同时 向胚胎内部陷入,在胚胎的局部表面形成内陷。
➢ 内卷(involution): 指胚体表面细胞通过连续细 胞运动向内卷折,去从内铺盖原来的外层细胞。
❖Hyaline layer=inner lamina (fibropellin) + outer lamina (hyalin)
❖内陷时, 只有植物极板细胞分泌硫酸软骨素 蛋白多糖 (chondroitin sulfate proteoglycan, CSPG) 到透明膜内层.
2. 间质细胞膜上的硫酸蛋白多糖(sulfated proteoglycans)
3. 囊胚腔细胞胞外基质(ECM)中只在原肠作用 期表达的ECM18。
去除硫酸基(sulfate deprivation)对海胆初 级间质细胞运动的影响
初 级 原 肠 内 陷
原肠内陷机制(II)
植物极内陷的双金属片模型。植物极板细胞分泌硫酸软骨素蛋白多 糖(CSPG)到透明层内层中,CSPG吸水膨胀,导致透明层弯曲, 相连的细胞层内陷。
早期原肠内陷
❖ 初级间质细胞在囊胚腔内迁移的过程中,仍然留 在植物极板上的细胞移动填补由初级间质细胞内 移而形成的空隙,植物极板进一步变扁平*。
❖ 之后,植物极板向内弯曲,内陷。当植物极板内 陷深及囊胚腔的1/4~1/2时,内陷突然停止。 所陷入的部分称为原肠(archenteron or primitive gut),而原肠在植物极的开口称为 胚孔(blastopore)。*
初级间质细胞 内移的扫描电 镜照片
发育生物学第四章原肠作用PPT课件

原肠作用概述 第一节 海胆的原肠作用 第二节 两栖类的原肠作用 第三节鸟类和哺乳类的原肠作用
第四节 胚轴的建立
1
标题添加
点击此处输入相 关文本内容
前言
点击此处输入 相关文本内容
标题添加
点击此处输入相 关文本内容
点击此处输入 相关文本内容
2
原肠作用概述
1.概念:原肠作用(gastrulation)是卵裂后期囊胚细胞剧
(4)内移(ingression):胚胎 表面一小部分细胞以单个细 胞向里面迁移的方式移入。 它们可由植物极一极移入, 也可在多处同时向内迁移的 多极移入。
6
(5)分层(delimination):一 个细胞层分离为近似平行的二 或多层细胞的过程。
(6)会聚延伸(convergent extension): 指细胞 间相互插入,使所在组织变窄、变薄,并推动组 织一定方向移动。
原肠作用前的海胆晚期 囊胚,植物极变扁平
9
海胆的胚胎发育 10
一、初级间质细胞的内移 海胚它初胆层们级原相将间肠 交 用质细作 于处胞用 骨形的: 的成线形初一状成级圈伪。间,足 质在细腹胞侧在有中一胚分层支和延海内沿运伪连腔并初合体c成针内伸ab胆初囊动足接预形级形(幼的植来球伸状(脱细胚级(mc,lee早级胚,与,定成间成s虫轴el物源的出伪f离胞腔间p)lysi)rlen期间腔主囊占腹稳质碳。索o极于细和足表,,质i,ncmp。cy原质内动胚据侧固细酸状d板小胞收面进称细最hatiirya肠细表伸腔囊面联胞钙合a中分不缩单入为胞终ym)l 胚胞面出壁胚,系融骨胞央裂断线层囊初e形,。
变窄、变薄,并推动组织朝一定方向移动----向背部中轴线迁移。
4
(1)外包(epiboly):表皮 层做为一个整体扩展,使 胚胎的内层被覆盖.
胚胎发育过程PPT课件
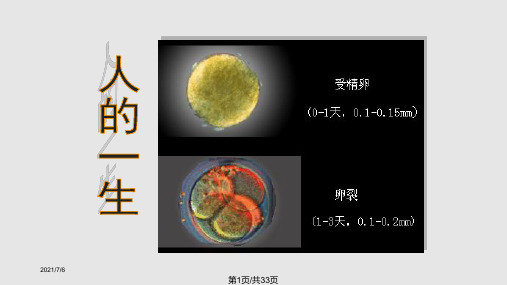
2021/7/6
第11页/共33页
囊胚
内细胞团: 发育各种组织;
滋养层: 发育胎膜、胎盘;
(孵化)过程: 囊胚扩大, 透明带破裂, 胚胎伸展出来。
内细胞团 囊胚腔 滋养层
2021/7/6
第12页/共33页
2021/7/6
原肠胚
外胚层 中胚层
原肠胚期
第13页/共33页
内胚层
2021/7/6
从 囊 胚 到 原 肠 胚 的 模 式 图
2021/7/6
外胚层:外胚表皮神经系
第16页/共33页
内胚层的分化
消化管,消化腺,呼吸道和 肺的上皮组织;
2021/7/6
中耳,甲状旁腺,胸腺和 膀胱等的上皮组织
内胚层:内胚两腺两道皮,
第17页/共33页
中胚层的分化
结缔组织,血管,肌肉组织;
皮肤的真皮,大部分中轴骨 骼及骨骼肌;
泌尿、生殖系统的主要器官, 心脏、体壁的骨骼肌肉血管 和结缔组织,消化和呼吸系 统的肌肉组织,血管和结缔 组织,
内细胞团内细胞团囊胚腔囊胚腔滋养层滋养层202042714原肠胚期原肠胚原肠胚外胚层中胚层内胚层202042716囊胚期胚泡8细胞期受精卵2细胞期4细胞期16细胞期32细胞期原肠胚期卵裂内胚层外胚层中胚层202042717牙釉质角膜上皮晶状体内耳膜迷路腺垂体口腔鼻腔和肛门的上皮等
2021/7/6
பைடு நூலகம்
第1页/共33页
第21页/共33页
受精后6星期,人形已隐约可见。这时,胚胎
2021/7/6
的心跳每分钟140—150下.是母亲心跳的两倍。
第22页/共33页
2021/7/6
人类胚胎第8周时在子宫内的照片
原肠胚的形成

组织分化、器官、系统形成
胚
幼体
后
温度等外界因素
发 育
成体
生物的个体发育
学习目标: 1、高等动物胚胎发育的大致过程 2、陆生脊椎动物羊膜出现的意义 3、高等动物胚后发育的特点,以及变态发
育的概念 学习重点: 1、胚胎从卵裂、囊胚到原肠胚的发育过程 2、羊膜出现的意义 学习难点:原肠胚的形成
高等动物的个体发育
胚的发育
胚后发育
受精卵
幼体
性成熟个体
一、胚胎发育
(外) (内)ห้องสมุดไป่ตู้(中) (中)
爬行类、鸟类、哺乳类
保证了水环境,具防震 和保护作用,增强了对 陆地环境的适应能力。
1. 麻雀的卵细胞和大象的卵细胞相比哪个大? 2. 卵生动物个体发育易受哪种非生物因素影响?
二、胚后发育
蝌蚪
变态发育
生活在水中 用鳃呼吸
有尾无四肢 心脏一心房
一心室
青蛙
水、陆地 用肺呼吸 有四肢无尾 心脏两心房
受精卵的特点
动物极
颜色深 卵黄少 比重小 朝向上端
植物极
颜色浅 卵黄多 比重大 朝向下端
受精卵进行细胞分裂
原肠胚
再看一遍
受精卵
沿卵轴方向
第一次卵裂
沿卵轴方向
第二次卵裂
偏向动物极
第三次卵裂
①细胞分裂速度差异 ②细胞移动
囊胚腔出现
囊胚
继续进行 植物极细胞內陷
动物极细胞外包
囊胚腔
原肠腔
原肠胚
原肠腔出现是形成原肠胚的标志。
囊胚腔
原肠腔
原肠胚
囊胚腔
原肠腔
原肠胚
囊胚腔
原肠腔
原肠胚
胚胎的早期发育--原肠胚PPT课件

初级 原肠 内陷
囊胚腔 植物极板细胞
植物极板细胞向上推举
透明层
内层 外层
微绒毛
含CSPG的分泌小 分泌到内层的
泡
CSPG吸水膨胀
植物极板内陷的双金属片模型。植物极板细胞分泌硫酸软骨素蛋白 多糖(CSPG)到透明层内层中,CSPG吸水膨胀,导致透明层弯 曲,相连的细胞层内陷。
内胚层建立
初级间质细胞在囊胚腔内迁移的过程中,仍然留在植物极 板上的细胞移动填补由初级间质细胞内移而形成的空隙, 植物极板进一步变扁平。
之后,植物极板向内弯曲,内陷。当植物极板内陷深及囊 胚腔的1/4~1/2时,内陷突然停止。所陷入的部分称为原 肠(archenteron or primitive gut),而原肠在植物极 的开口称为胚孔(blastopore)。
初级间质细胞
植物极板中 央来源于小 分裂球的细 胞不断伸出 和收缩线状 伪足,脱离 表面单层细 胞,进入囊 胚腔,称为 初级间质细 胞。
海胆早期原肠 胚内初级间质 细胞沿囊胚腔 内表面运动, 主动伸出伪足 与囊胚腔壁连 接,占据囊胚 腔预定腹侧面, 并形成稳固联 系。
初级胎的背部,即刚好位于赤 道下方的灰色新月区开始的。
灰色新月区的预定内胚层细胞内陷,形成狭缝状胚孔
(slit-like blastopore),内陷细胞称为瓶状细胞(bottle
cell),它们沿最初的原肠排列 。
边缘区
囊胚腔
爪蟾的晚期囊胚,示边缘区被内胚层细胞覆盖。
从尚未分化到分化为三个胚层和器官原基决定的 关键时期。
原肠胚发育的细胞运动方式
七种细胞运动机制
发育生物学第4章-鸟类盘状卵裂

鸟类原肠作用机制
背腹轴的决定
pH9.5(背) -- - -- - -+ + + + ++ + + pH6.5(腹)
五、鸟类
盘状卵裂。
两栖动物背唇和胚孔出现是原肠 发育的标志,而爬行动物、鸟类 和哺乳动物的原肠形成是以原条 的出现为标志的。
鸡上、下胚层发育命运关系图
上胚层 下胚层
胚胎的三个胚层
大量的胚胎外膜 卵黄囊
部分的胚胎外膜 基 柄
尿囊
鸡
胚
盘
上
下
胚
层
卵黄囊
的
形
成
与
原
条
的
定
位
原
鸡原条发育过程中的细胞迁移运动
条
(背唇) (胚孔)
25 mV
前后轴的决定
当卵子从输卵管排出 时,以10~15 r/h的速 度旋转。在卵子旋转 时,将来形成细胞层 的细胞质总是朝向下 方,其上方却受到卵 黄的挤压。
将来形成细胞层的细胞质并不是位于卵黄正上方,而是 稍稍偏向ห้องสมุดไป่ตู้侧。胚盘最突出的部分将形成囊胚尾端,即 原肠作用的起始处。
因此,背腹轴和前后轴在卵子 从输卵管排出时就已决定。
在鸟类,暗区细胞上表面与卵黄 膜下表面紧密相连。暗区细胞能 沿卵黄膜下表面扩展。
纤连蛋白附着位点序列Arg-Gly-Asp
六、哺乳动物
人 胚
人 上、下胚层发育命运关系图
胎
发育生物学第四章原肠胚形成课件PPT

海胆正常的胚胎发育
海胆的长 腕幼虫
第二节 果蝇 的原肠作用
原肠作用开始于 腹部预置中胚层 的内陷。
羊浆膜(amnioserosa)的 作用可能是分泌信号分 子,指导胚带(germ band)的收缩,它最后 将退化
第三节
鱼的原肠作用
斑马鱼中囊胚(经历了10次卵裂)的三 类细胞:
1.卵黄多核层(yolk syncytial layer, YSL):胚盘的植物极边缘细胞裂 解,其核和质与卵黄细胞融合在一起而构 成的一层细胞核层。在胚盘下包中,部分 YSL细胞核移向胚盘下成为internal YSL,它们可能起提供营养的作用;边缘 处的为external YSL,它们可能起驱动下 包的作用。
鸡胚的原肠作用
下胚层的形成
鸟类胚胎3个胚层皆起 源于上胚外成分
鸡胚原条(primitive streak)的形成:由后部边缘区的上胚层细胞加厚而成,它的出现 确定了胚胎的A-P轴线。原条内会形成一个凹陷,叫原沟,是上胚层细胞进入囊胚腔的门 户。原条的头部末端是一个加厚层,叫Hansen`s node(亨氏结), 是一个诱导中心。
2. Enveloping layer (EVL):位于 胚盘最外层已表皮化的细胞,发育后期会 脱落。 3. Deep Cells:介于YSL和EVL之 间的细胞,它们将发育为胚胎本体。晚期 囊胚的深层细胞的命运已经建立。
鱼类的盘状偏裂
斑马鱼受精卵的前5次卵裂均为经线裂,产生的32个 细胞为单层分布于卵黄上。其后的分裂方向不规则。囊 胚期开始于128细胞期,属盘状囊胚。
次级间充质 次级间充质 伪足 胚孔 初级间充质 中、内胚层内陷 原肠 肛门 肠
原肠胚
口
原肠延伸机制
原肠顶端形成次级间质细胞,原肠延伸 是以次级间质细胞提供的张力为动力的 。次级间质细胞伸出线状伪足,穿过囊 胚腔液,直达囊胚腔壁内表面。 动物极半球存在着次级间质细胞附着的 靶位。只有当线状伪足接触到特定靶位 的时候,才不会缩回。这些特定的靶位 可能位于将来形成口的区域。
原肠作用及早期发育ppt课件

--
25
至少有三种蛋白可能在间质细胞的迁移过程中起 重要的作用,包括囊胚腔壁基质片层组分纤连蛋 白(fibronectin)、间质细胞膜上的硫酸蛋白多 糖(sulfated proteoglycans)和囊胚腔细胞胞 外基质中只在原肠作用期表达的ECM18。
去除硫酸基(sulfate deprivation)对海胆初级间质细胞运动的
--
26
二、早期原肠内陷
初级间质细胞在囊胚腔内迁移的过程中,仍然留在 植物极板上的细胞移动填补由初级间质细胞内移而 形成的空隙,植物极板进一步变扁平。
之后,植物极板向内弯曲,内陷。当植物极板内陷
深及囊胚腔的1/4~1/2时,内陷突然停止。所陷入
的部分称为原肠(archenteron or primitive
胞在植物极区域形成一细胞环。
--
31
三、晚期原肠内陷
早期原肠内陷完成之后,经过短暂停歇, 原肠大幅度拉长, 短粗的原肠变成又细又 长的管状结构。在此期间没有新细胞形成, 原肠的拉长过程是通过细胞重排实现的, 原肠周长内细胞数目大为减少。
--
32
海胆胚胎原肠伸长时期的细胞重排。
--
33
原肠顶端形成次级间质细胞,原肠延伸是以次级间质细胞提 供的张力为动力的。次级间质细胞伸出线状伪足,穿过囊胚 腔液,直达囊胚腔壁内表面。
相对运动形成了一个在精子进入点对面的新月形的灰色区域,称为灰色新月
--
55
灰色新月区的预定内胚层细胞内陷,形成狭 缝状胚孔(slit-like blastopore),内陷 细胞称为瓶状细胞(bottle cell),它们 沿最初的原肠排列 。
胚孔 处形 成瓶 状细 胞
--
56
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
去除硫酸基(sulfate deprivation)对海胆初 级间质细胞运动的影响
初 级 原 肠 内 陷
原肠内陷机制(II)
植物极内陷的双金属片模型。植物极板细胞分泌硫酸软骨素蛋白多 糖(CSPG)到透明层内层中,CSPG吸水膨胀,导致透明层弯曲, 相连的细胞层内陷。
Hyaline layer=inner lamina (fibropellin) + outer lamina (hyalin)
六 五 四 三 二 一 哺 乳 类 原 肠 作 用 鸟 类 原 肠 作 用
两 栖 类 原 肠 作 用
鱼 类 原 肠 作 用
海 胆 原 肠 作 用
原 肠 作 用 概 述
主 要 内 容
原肠作用 Gastrulation 概述
原肠作用是指囊胚细胞有规则的移动,使细胞重新排列,用 来形成内胚层和中胚层器官的细胞迁入胚胎内部,而要形成外胚 层的细胞铺展在胚胎表面。原肠作用期的胚胎叫原肠胚(gastrula).
早期原肠内陷
初级间质细胞在囊胚腔内迁移的过程中,仍然留 在植物极板上的细胞移动填补由初级间质细胞内 移而形成的空隙,植物极板进一步变扁平*。
之后,植物极板向内弯曲,内陷。当植物极板内 陷深及囊胚腔的1/4~1/2时,内陷突然停止。 所陷入的部分称为原肠(archenteron or primitive gut),而原肠在植物极的开口称为 胚孔(blastopore)。*
初级间质细胞 内移的扫描电 镜照片
内移至囊胚腔的初级间质细胞被大量胞外基质包绕。原肠作用过 程中沿动植物极轴分布的纤维决定了初级间质细胞的迁移方向。
囊胚腔胞外基质纤维 与动植物极轴平行排 列,最终与初级间质 细胞发生联系。
至少有三种蛋白可能在基质片层组分纤连蛋白( fibronectin) 2. 间质细胞膜上的硫酸蛋白多糖(sulfated proteoglycans) 3. 囊胚腔细胞胞外基质(ECM)中只在原肠作用 期表达的ECM18。
初级间质细胞形成的索状合胞体(将来形成骨针)的扫描电镜照片
腹外侧细胞簇
初级间质细胞在 Lytechinus variegatus早期幼虫 内的定位。
骨针 背链 腹链
原肠内陷机制(I)
细胞粘连作用的变化导致初级间质细胞内移。 初级间质细胞和透明 层以及相邻裂球之间的亲和力减少,而与基质片层之间的亲和力增 加。相比之下,非间质细胞和透明层以及相邻裂球之间的亲和力维 持不变。
纤丝收缩使细胞变为契形, 加之相邻表层细胞运动产生 的推动力,加速植物极板内 内陷时, 只有植物极板细胞分泌硫酸软骨素 陷,形成原肠(archenteron 蛋白多糖 (chondroitin sulfate proteoglycan, or primitive gut) CSPG) 到透明膜内层. CSPG 吸水能力很强. CSPG吸水使透明层 内层膨胀,而外层维持不变.
次级间充质 次级间充质 伪足 胚孔 初级间充质 中、内胚层内陷 原肠 肛门 肠
原肠胚
口
海胆原肠作用过程:初级间质细胞在中胚层和内胚层相 交处形成一圈,在腹侧有一分支延 伸,它们将用于骨的形成。
随着植物极板的内陷,初级间质细胞在植物极区域形 成一细胞环。
海胆早期原肠胚 内初级间质细胞 沿囊胚腔内表面 运动,主动伸出 伪足与囊胚腔壁 连接,占据囊胚 腔预定腹侧面, 并形成稳固联系。 初级间质细胞融 合形成索状合胞 体(syncytial cable),最终形 成幼虫碳酸钙骨 针的轴。
表皮细胞和间质细胞的概念
表皮细胞(epithelial cells):细胞与细胞间紧密连接成管 状或片层状结构, 局部或整个结构一起运动。 间质细胞(mesenchymal cells):细胞与细胞间松散相连,每 个细胞为一个行动单元。
第 一 节 海 胆 的 原 肠 作 用
海胆的早期卵裂
海胆胚胎第四次卵裂时形成的小分裂球( micromere)
初级间质细胞 的内移
植物极板中央来 源于小分裂球的 细胞不断伸出和 收缩线状伪足 (filopdia),脱 离表面单层细胞 进入囊胚腔,称 为初级间质细胞 (primary mesenchyme cell)。
初级间 质细胞 的线状 伪足*
海胆原肠作用过程:原植物极中央细胞内陷进入囊胚腔,表皮细胞 转变成为初级间质细胞,然后中、内胚层表皮细胞内陷和扩展,其 前端表皮细胞转化为次级间质细胞。两种间质细胞都将长出伪足, 后者起定向和驱动细胞移动的作用。
(二)原肠作用中的主要细胞迁移方式
外包(epiboly): 以整片细胞(通常为外胚层细 胞)沿胚胎表面扩展,包裹胚体的深层细胞。
内陷(invagination): 指外层细胞成片地同时向 胚胎内部陷入,在胚胎的局部表面形成内陷。 内卷(involution): 指胚体表面细胞通过连续细 胞运动向内卷折,去从内铺盖原来的外层细胞。
海胆的囊胚 : (A)细胞持续的分裂形成一个囊胚; (B) 快速分裂的末期,圆形的胚胎细胞形成一层上皮,受精膜依 然可见; (C)海胆胚胎的早期囊胚,植物极板变厚,动物极细胞分泌孵化 酶,使胚胎从受精膜中孵化出来。
原肠作用前海 胆囊胚的结构
海胆的晚期囊胚 由1000个左右单 层细胞构成,植 物极变扁平。
(一) 原肠作用的概念和主要特点
原肠作用是细胞和细胞层高度协调一致的运动 .
通过原肠作用,胚胎首先建立起三个胚层,即外胚层、中胚层和内 胚层; 其次为重新占有新位置的胚胎细胞之间的相互作用奠定了基础。因 此,原肠作用是从尚未分化,到分化为三个胚层和器官原基决定的 关键时期。 原肠胚一部分细胞的移动必然和同时发生的其他细胞的运动紧密配 合,因此原肠作用过程中的细胞运动涉及整个胚胎。
内移(ingression): 指表层的单个细胞迁入胚胎 的内部。
分层(delamination):指一个细胞层分成两层或 多或少平行的细胞层。 会聚伸展(convergent extension): 指细胞间 相互插入,使所在组织变窄、变薄,并推动组织 一定方向移动。
会聚伸展Convergent extension