LBD1-30基因调控杨树茎干次生生长的功能分析
根际微生态调节对杨树根系活力及土壤有效P转化的效应

大相 关性 ; 毛 白杨 和 北 京 杨根 际 土壤 酸性 磷 酸酶 活性 增 强 , 别 增 加 2 . 1 ~ 3 . 5 和 1 . 9 ~ 使 分 0 9 6 3 33 %
5 . 8 , 根 际 区域 的 土壤 有 效 P具 有 明显 的 富集 效 应 , 集 率 分 别 为 7 4 ~2 . 7 和 1 . 5 ~ 4 9 对 富 .9 5 6 78 %
ssa c n u e t nf r d sg o h u tn sg o t b o s , o tvg r s i a i h s h t s c it n e id c rwih u io m e i n, n t ec ti g r w h, i ma s r o io . o l cd p o p a a ea —
Th o p ig a e t lo e h n e h r n f r a in o v i b e P fo is l b e P mi e a s An h e c u l g n sa s n a c d t e ta so m to fa al l r m n o u l n r l. d t e n a
Po u u × b i i g n i c t n swe e6 7 5 3 6 a d 1 4 一 9 4 h g e h n t o eo h o — p l s ej n e ss u t g r . 1 3 . 4 i 4 n . 9 4 . 6 i h r t a h s ft ec n
杨树摄取氮素的根系-微生物互作机制及靶向调控技术

杨树作为一种重要的绿化和经济林树种,其对氮素的需求量较大。
氮素是植物生长所需的重要营养元素,土壤中氮素浓度直接影响着植物生长和生理特性。
在人工林培育过程中,合理施肥不仅会影响苗木生长,也会影响苗木材性。
因此,研究杨树摄取氮素的根系-微生物互作机制及靶向调控技术具有重要意义。
1. 杨树摄取氮素的根系-微生物互作机制:
杨树根系能够促进土壤微生物的活动和数量的增加,还能改良土壤的肥力。
杨树根系与土壤微生物之间存在密切的互作关系,这种互作关系对杨树摄取氮素具有重要的影响。
研究发现,杨树根系能够通过分泌有机物质、改变土壤pH等方式影响土壤微生物的生存环境和数量,进而影响杨树对氮素的吸收和利用。
2. 靶向调控技术:
为提高杨树对氮素的利用效率,可以采用靶向调控技术,主要包括以下几个方面:
(1)合理施肥:根据杨树的生长需求和土壤条件,制定合适的氮素施肥方案,避免过量施肥导致氮素损失和环境污染。
(2)土壤改良:通过改善土壤结构、增加土壤有机质含量等方法,提高土壤微生物数量和活性,促进杨树对氮素的吸收和利用。
(3)基因工程:通过基因工程技术,研究杨树氮素转运蛋白基因,提高杨树对氮素的吸收和利用能力。
(4)生物菌剂:利用生物菌剂(如固氮菌、硝酸盐还原菌等)可以促进土壤中氮素的转化和利用,提高杨树对氮素的吸收能力。
杨树林带萌蘖更新技术规程征

杨树林带萌Il更新技术规程1范围本标准规定了杨树林带萌弟更新的术语和定义、萌集更新条件、更新前准备、更新模式及关键技术点、更新后抚育管理以及技术档案。
本标准适用于全省杨树林带萌美更新造林。
2规范性引用文件下列文件对于本文件的应用是必不可少的。
凡是注日期的引用文件,仅注日期的版本适用于本文件。
凡是不注日期的引用文件,其最新版本(包括所有的修改单)适用于本文件。
GB13735聚乙烯吹塑农用地面覆盖薄膜GBrr18337.3-2001生态公益林建设技术规程DB32/T2382杨树人工林修枝技术规程3术语和定义下列术语和定义适用于本文件。
3. 1林带更新建立新的林带以代替已衰老林带的措施。
林带树木达自然成熟龄后,将逐渐衰老,稀疏,防护效能也将逐渐降低。
为保持林带防护效益的永续性,建立新一代林带代替衰老的林带。
3.2萌芽更新森林被采伐以后,利用林木伐桩的萌芽力使森林得以恢复的更新方式。
从萌芽条在伐根上发生的部位看,可分为发自伐根断面、伐根中部和根颈三处。
发自伐根断面,由不定芽产生的萌芽条,生长孱弱,常很快死去。
从根颈处发生形成的萌芽条,生活力强,比较稳固,且能很快地在地面接触处形成不定根,进而形成独立的根系。
从伐根中部发生形成的萌芽条,它的生活力介于上述二者之间。
3.3根票更新依靠由根上不定芽长出新的植株形成新林分的更新方式。
4萌篥更新条件3.4 对更新林带的要求(1)待更新林带现有林木保存率应完好,林相相对整齐;(2)原林带密度应适宜,林木生长健壮的成熟林或近熟林;(3)原林带立地条件要相对较好;(4)原林带杨树品种为良种。
3.5 更新季节选择拟要萌麋更新的杨树林带在采伐季节要求应在林木停止生长的冬季到春季树液流动之前(休眠期)进行。
3.6 采伐方式待更新林带应采用锯伐方式。
3.7 伐桩要求待更新林带伐桩高度以不超过5Cm为宜。
5更新前准备更新采伐后,应对林地进行清理,清除采伐过程中出现的枝梗、梢头、杂灌及其剩余物。
基因工程技术在杨树抗逆境方面的研究进展

基因工程技术在杨树抗逆境方面的研究进展摘要:阐述了基因工程技术在杨树抗逆遗传改良方面的应用,总结了转基因杨树抗病虫、抗旱、耐盐以及在环境修复中取得的研究成果,讨论了基因工程技术在杨树抗逆方面存在的问题。
杨树因长速快、产量高、基因组小等特点,具有较高的经济价值和生态效益,同时也是林木遗传改良的模式树种。
尽管对杨树抗逆遗传改良进行了相关研究,但有关外源基因提高杨树抗逆性的机理和应用仍有待进一步深入研究。
挖掘新的抗逆基因、培育具有抗性的杨树新品种仍将是今后的研究重点。
关键词:基因工程;杨树;抗逆收稿日期:2022-10-161转基因杨树抗病虫研究1.1抗病研究1.1.1抗病毒病抗病毒基因工程研究在林业上的应用才刚起步,目前,使用的基因主要有洋李痘病毒的外壳蛋白基因和黄瓜花叶病毒外壳蛋白基因等几种。
Cooper、李玮等分别将花叶病毒外壳蛋白基因和抗菌肽LCI基因导入杨树中,均获得良好抗病效果的转基因植株[4-5]。
1.1.2抗细菌病农杆菌感染杨树后,杨树极易感染冠瘿病。
目前,关于杨树抗冠瘿病的研究主要是采用转基因技术将农杆菌致瘤基因、激素IAA基因、抗菌肽基因(兔防御素NP-Ⅰ基因)导入杨树,获得抗性提高的转基因植株[6]。
1.1.3抗真菌病植物受到病原物侵染后,会使多种病程相关蛋白迅速表达,参与其抗病防卫反应。
在这些病程相关蛋白中,最主要的蛋白之一就是几丁质酶[7],几乎所有的植物器官中均可以发现几丁质酶[8]。
在正常情况下,几丁质酶水平很低,但经诱导因子的诱导,表达量可以迅速增加。
病原真菌、细菌、病毒的侵染、激发子和一些逆境等均可能诱导植物几丁质酶的表达[9]。
近年来,有关对植物几丁质酶的特性、基因结构、分类、分子进化、生物学作用及转几丁质酶基因的研究,已成为植物抗真菌病害的研究热点之一[10-11]。
1991年,Broglie等首次对转几丁质酶基因在植物的抗真菌病方面进行了研究,他们将菜豆几丁质酶基因转入烟草和油菜,并得到表达,有效降低了植株的死苗率,控制了病情的发展[12]。
杨树次生壁纤维素合酶的表达与互作模式分析

杨树次生壁纤维素合酶的表达与互作模式分析魏凯莉1 ,周厚君1,江成1,赵岩秋1,宋学勤1,2* ,卢孟柱1,2【摘要】摘要:[目的]纤维素的合成在木材形成过程中具有重要的作用,纤维素合酶(Cellulose Synthase , CESA)是参与纤维素合成的关键酶。
由于3种CESA形成一个有功能的纤维素合酶复合体,而杨树中包含CESA4、CESA7A. CESA7B. CESA8A和CESA8B5种次生壁CESA ,本文从这5种CESA的表达模式与互作分析入手,探讨了其在次生壁纤维素合成中的工作模式。
[方法]利用RNA-seq与基因芯片数据进行基因表达分析,揭示5种次生壁CESA在根、茎、叶组织的表达模式与次生维管再生过程中的表达变化。
利用启动子驱动的GUS转基因材料GUS染色分析与实时定量PCR揭示5种次生壁CESA在各个组织的表达模式与在激素处理下的响应模式。
利用荧光素酶互补实验揭示CESA7A. CESA7B. CESA8A与CESA8B之间的互作模式。
[结果]基因表达分析表明,杨树5种次生壁CESA基因在杨树成熟茎中高表达,尤其在次生维管组织发育的后期高表达,表明其主要参与木材次生壁纤维素的合成。
对启动子驱动的GUS转基因材料观察表明,CESA4、CESA7B. CESA8A和CESA8B 的GUS 信号在杨树茎和叶中较强,但这些次生壁CESA的表达模式存在一定的差异,这种差异在叶脉中表现地尤为明显。
此夕卜,在赤霉素GA3和细胞分裂素6-BA处理下,杨树次生壁CESA的表达量显著上调;在生长素NAA、油菜素内酯BR和乙烯处理下,杨树次生壁CESA的表达量下调,但不同的CESA对激素响应的表达量变化幅度存在差异。
荧光素酶互补实验表明,杨树次生壁CESA7B 和CESA8B、CESA7B 和CESA8A、CESA7A 与CESA8B 之间存在互作,说明它们之间可以形成复合体。
[结论]这些数据显示杨树5种次生壁CESA。
中科院遗传所王国栋白洋合作揭示新进化的植物特异性代谢产物对根系微生物组的调控作用

中科院遗传所王国栋白洋合作揭示新进化的植物特异性代谢产物对根系微生物组的调控作用植物产生超过20万种代谢物(主要是指分子量小于1,000的化合物),这些代谢物在植物的生活史中发挥着重要的生理功能。
由于植物是固着生长,而根系周围有大量的微生物(微生物组),长期以来人们认为植物代谢物在植物与根系微生物互作方面发挥着重要的作用。
但迄今为止,关于植物如何利用自身特异的代谢物来调控根系微生物的组成还鲜有报道。
近日,中国科学院遗传与发育生物学研究所王国栋课题组和白洋课题组合作在Science China Life Sciences发表了题为Recently duplicated sesterterpene (C25) gene clusters in Arabidopsis thaliana modulate root microbiota的研究论文,揭示了植物自身利用新进化的特异性代谢产物对根系微生物组的调控作用。
在前期的研究中,王国栋课题组鉴定了十字花科保守的二半萜生物合成基因簇(GFPPS-TPS-P450)。
同时发现,拟南芥(Arabidopsis thaliana)中新进化获得的两类二半萜化合物的生物合成能力(图1中TPS25的产物为Compound 3和TPS30的产物为Compound 4,结构上两种化合物都具有五环二半萜骨架),而且这两种化合物特异性地在根部积累。
图1. 拟南芥中新进化的二半萜化合物调控根系微生物组。
A和B 表示拟南芥新进化(特有)的两个二半萜生物合成GFPPS-TPS-P450基因簇和对应生成的二半萜(C25)化合物。
C表示两种二半萜化合物对根系微生物组组成的调控既有特异性,也有共性。
通过与白洋课题组合作,比较自然土中生长的野生型(Col-0)和不同的TPS25、TPS30突变体的根系微生物组,发现野生型和TPS25、TPS30突变体形成了显著不同的根系微生物组。
有意思的是,tps25和tps30单突变体根系微生物组的变化具有比较大程度的交叉,暗示植物利用次生代谢物调控根系微生物组成可能存在构效关系,即结构类似的化合物具有类似的调控微生物组成功能。
转基因杨树研究进展

转基因杨树研究进展
抗病基因研究
• 用于杨树抗病转基因研究的有几丁质酶、 防御和抗菌肽等基因
• 转基因植株对转基因株组织提取液对枯草 杆菌、农杆菌和立枯病原菌等的生长有抑 制作用。
转基因杨树研究进展
抗胁迫基因研究
• 近几年来,利用基因工程的方法在研究植 物抗氧化剂,低温,土壤的盐渍化和沙漠 化抗胁迫能力方面取得了重大进展
转基因杨树研究进展
转基因工作中存在的问题及解决方 法
• 转基因植株环境释放后的安全性问题 (1) 打破生态平衡 (2)产生新的过敏原 (3) 基因污染
转基因杨树研究进展• 转基因杨树的应用前景转基因杨树研究进展谢谢大家
转基因杨树研究进展
转基因杨树研究进展
转基因杨树研究进展
转基因杨树研究进展
转基因杨树研究进展
• 方法
转基因杨树研究进展
遗传转化 • (1)间接转化法——农杆菌介导转化法
• (2)直接转化法
•
A、 基因枪转化法
•
B、 电击法
•
C、花粉管通道法
•
D、 PEG介导基因转化法
转基因杨树研究进展
• 自1983年首次获得转基因烟草、马铃薯 • 目前已经获得转基因植株的植物已达数百
转基因杨树研究进展
Bt基因(苏云金芽孢杆菌Bacillus thuringiensis)
B t -t o x i n基因是世界范围内广 泛使用的抗虫基因。是 苏云金杆 菌在芽孢形成过程中构成伴孢 晶 体的最主要 的 蛋白。
Bt蛋白的毒理研究 Bt蛋白本身是无毒的,是一种原 毒素。这种原毒素可以被某些昆 虫体内的酶活化,随后能够结合 在肠道的受体上,造成肠道穿孔。
• 资料表明,昆虫特异性蝎神经毒素AaIT具有较高 的选择性,对哺乳动物和其他动物无害,安全性 好,对许多鳞翅目害虫具有较高的毒性,是十分 理想的抗虫基因源。
农杆菌介导转化和再生的杨树讲解

农杆菌介导法转基因杨树摘要:杨树品种已发展为一种植物转化和再生系统。
叶植,从稳定发芽培养的一个杨树杂交NC - 5339(银白杨标本),被共培养用于农杆菌遗传转化关于一个烟草的看护培养。
致瘤的和无防备的农杆菌株隐藏包含一个双元载体,其中包含两个新霉素磷酸转移酶II(NPT II')和细菌5莽草酸3-磷酸合酶(EPSP)(AROA)嵌合基因融合。
没有开发芽,叶外植体时,双元缴械拉力的根癌农杆菌菌株共培养。
然而,转化的植物,没有野生型的T-DNA获得使用农杆菌株原癌基因的二进制。
NPT II '酶的活性检测,Southern印迹法分析和免疫学检测证实了遗传转化成功细菌EPSP合酶Western印迹。
这是首次报道成功收回转化植株森林树,也是第一个记录的插入和重要农艺性状的外源基因的表达成木本植物物种。
关键词:白杨;转化;农杆菌前言基因工程树种的能力将是特别有用的遗传改良,如大型成熟的植物并长期有性世代倍(Nelson and Haissig 1984; Sederoff and Ledig 1985)。
森林树种的应用重组DNA技术的一个先决条件是发展的基因转移系统。
方法,例如显微注射(Crossway et al.1986)和直接DNA摄入(Paszkowski et al. 1985; Fromm et al. 1986) 已被用于外源基因引入到草本作物物种,但是,最有效的基因转移的方法,利用自然感染冠瘿病的机制造成的有机体,农杆菌(Bevan et al. 1983 ; Fraley et al. 1983 ; Herrera-Estralla, 1983). 。
根癌农杆菌的自然感染周期期间,细菌的T-DNA 整合到宿主植物的染色体,从而导致肿瘤对植物的生产(奇尔顿等人,1980)。
可以删除和替换而不影响根癌农杆菌的T-DNA转移到植物(DeGreve等,1982)的能力,由异源基因的肿瘤诱导基因。
杨树插干与带蔸造林生长及根系发育比较
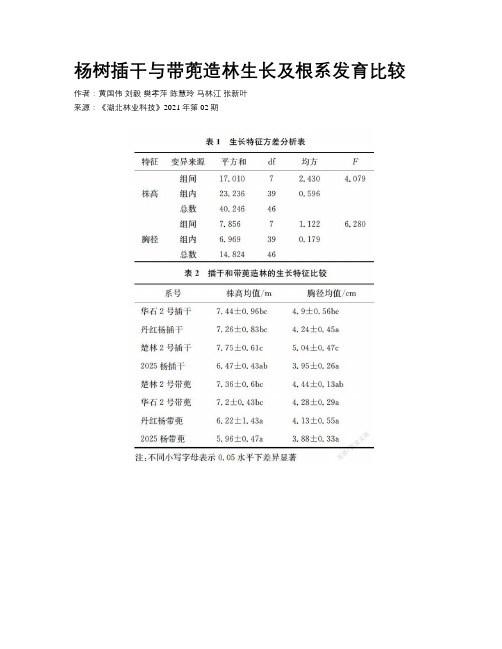
杨树插干与带蔸造林生长及根系发育比较作者:黄国伟刘毅樊孝萍陈慧玲马林江张新叶来源:《湖北林业科技》2021年第02期摘要:以江汉平原4个杨树主栽品种为试验材料,分别进行插干和带兜造林,比较两种造林方式生长和根系发育差异。
结果发现:两种造林方式,生长差异显著,插干造林具有明显的生长优势。
进一步对两种造林方式根系发育进行比较发现,前期根系发育差异不大,随着时间增长,插干造林的生根数量、根重、生根区间长度等特征都显著大于带兜造林。
正是由于插干造林根系发达,分布均匀,吸收根和运输支撑根合理配置,更利于水分、养分的吸收运输,且不易倒伏,最终相比带兜造林表现出显著的生长优势。
关键词:杨树;造林方式;生长;根系中图分类号:S722.3;S792.11 文献标识码:A 文章编号:1004-3020(2021)02-0001-05Comparison of Populus simonii Growth and Root Development Between Cuttage Forestation and Afforestation with RootsHuang Guowei(1,2) Liu Yi(3) Fan Xiaoping(4) Chen Huiling(1,2) Ma Linjiang (1,2) Zhang Xinye(1,2)(1.Hubei Academy of Forestry Wuhan 430075;2.Hubei Fast-growing and High-yield Forest Engimeering Technology Research Center Wuhan 430075;3.Hubei Xingdoushan National Nature Reserve Enshi 445000;4 Shishou Research Institute of Poplar for Hubei Academy of Forestry Jingzhou 434400)Abstract: In order to provide technical support for the efficient afforestation of Populus simonii,four main Populus simonii varieties in Jianghan Plain were used as experimental materials to compare the differences of growth and root development between two afforestation methods.The results showed that difference of growth was significant between two afforestation methods,and cuttage forestation had obvious growth advantage.Further comparing the root development of the two afforestation methods,it was found that there was little difference in early root development.As time went on,the rooting number,root weight and length of rooting interval of the cuttage forestation were significantly higher than the afforestation with root.The cuttage forestation had the well-developed root system,uniform distribution,absorption root and transport support root reasonable allocation,it was more conducive to moisture and nutrient absorption,and not easy to lodging,and finally,the cuttage forestation showed significant growth advantages to the afforestation with root.Key words: Populus simonii;afforestation methods;growth;root system楊树Populus simonii作为中国重要的速生用材树种,每年都有的大量的新造林,研究表明造林方式不同会影响杨树的成活率和生长速度[1-4]。
杨树中与次生细胞壁生物合成相关的糖基转移酶研究进展

杨树中与次生细胞壁生物合成相关的糖基转移酶研究进展杨少辉;王洁华;宋英今;胡荣峰【摘要】随着杨树基因组测序计划的完成,加之生物信息学、生物化学、分子生物学和遗传学等研究方法的不断完善,杨树功能基因组学研究近年来被大大推进.目前,有关杨树材性的研究主要集中于MYBs、NACs、bZIP、HB、WRKY、MADS等转录因子的调控机制以及木质素的合成途径.与之相比,有关次生细胞壁中关键碳水化合物结构的合成及代谢仍然不清楚.糖基转移酶参与木本植物次生细胞壁中碳水化合物的合成和架构,直接影响木质部的发育过程.着重介绍了木本模式植物——杨树中参与次生细胞壁生物合成的糖基转移酶基因家族及其生物学功能,并对其在利用基因工程手段改良杨树材性和选育新品种方面进行了展望.%With the completion of genome sequencing of populus trichacarpa,together with the methods of bioinformatics,biochemistry,molecular biology and genetics,the functional genomics of poplar were greatly promoted.So far,the wood-related reseaches in poplar were mainly concentrated on the transcripional factors including MYBs,NACs,bZIP,HB,WRKY,MADS and on the lignin biosynthesis pathway.In comparison,the metabolic pathways of the key structural carbohydrates still remain unclear.Glycosyltransferases are involved in the biosynthesis and remodeling of the carbahydrates in poplar xylem,thus directly affecting xylem developmental process.In this review,an emphasis on the glycosyltransferase gene family and their biological functions involved in secondary cell wall biosynthesis in the woody model system-poplar weere summarized,and the futureperspectives on the modification of wood properties in poplar by genetic engineering and new variety breeding were given.【期刊名称】《生物学杂志》【年(卷),期】2013(030)003【总页数】4页(P68-71)【关键词】杨树;次生细胞壁;糖基转移酶;基因家族【作者】杨少辉;王洁华;宋英今;胡荣峰【作者单位】天津大学环境科学与工程学院,天津300072;天津大学环境科学与工程学院,天津300072;天津大学环境科学与工程学院,天津300072;天津大学环境科学与工程学院,天津300072【正文语种】中文【中图分类】Q555木材对人类生活具有重要的意义,具有显著的经济效益和生态效益。
杨树多基因遗传转化及外源基因表达研究的开题报告

杨树多基因遗传转化及外源基因表达研究的开题报告一、研究背景和意义随着基因工程技术的不断发展和完善,转基因技术在植物领域的应用越来越广泛。
杨树作为重要的经济林木种类,具有快速生长、广泛适应性以及丰富的经济价值等优点。
然而,由于杨树基因组结构较为复杂,传统遗传育种方法难以满足高效、精确的育种需求。
因此,利用基因工程技术为杨树进行基因遗传改良,具有重要的理论和应用价值。
本研究旨在探究杨树基因多样性和转化效率,以及外源基因在杨树中的表达模式,为杨树转基因育种提供理论基础和实验指导,同时也为杨树的遗传资源保护和利用提供支持。
二、研究内容和方法1. 杨树多基因遗传转化通过分析杨树的基因组结构和基因功能,选择多个与抗病、抗逆等相关的基因作为转化目标,利用农杆菌介导的遗传转化技术,将目标基因导入杨树细胞中,并利用筛选方法筛选出转化成功的株系。
2. 转化效率分析通过筛选出的转化株系,分析其转化效率和基因表达水平,并比较不同转化方式、转化载体和转化方法的影响因素,为提高杨树遗传转化效率提供参考。
3. 外源基因在杨树中的表达模式研究通过利用荧光素酶、荧光蛋白等外源基因作为模型基因,观察其在转化株系中的表达模式和对杨树发育及生长的影响,为外源基因在杨树中的应用提供参考。
研究方法包括杨树细胞培养、杨树遗传转化、基因筛选、基因表达分析等。
三、研究预期结果1. 筛选出多个转化成功的杨树株系,并明确其基因型和表型特征。
2. 分析不同转化因素对杨树转化效率的影响,为提高杨树遗传转化效率和精度提供参考。
3. 观察外源基因在杨树中的表达模式和对杨树发育生长的影响,为外源基因在杨树中的应用提供参考。
四、研究意义和应用价值1. 探究杨树基因多样性、遗传转化效率和外源基因表达模式,为杨树遗传育种和遗传资源保护提供理论和实验支持。
2. 认识杨树基因转化效率和外源基因表达模式的影响因素,为提高杨树的抗病、抗逆等性状提供新思路。
3. 探索杨树基因工程育种技术,为提高杨树经济效益和可持续发展提供技术支持和保障。
杨树次生代谢产物的生物合成及其功能研究

杨树次生代谢产物的生物合成及其功能研究随着人类对药物、化妆品等生活必需品需求的不断增长,各类天然产物的开发与利用变得日益重要。
杨树作为一种广泛分布的植物资源,其次生代谢产物具有丰富多样的生化活性,逐渐成为了生物合成及功能研究的热点。
杨树次生代谢产物的简介次生代谢产物指的是在杨树生长过程中,不参与生长和发育过程的化合物。
杨树次生代谢产物可分为苯丙烷类、黄酮类、生物碱类、萜类等多种类型。
其中苯丙烷类次生代谢产物广泛存在于植物体内,玫瑰醇、桂皮醇、莨菪烷等都是苯丙烷类次生代谢产物的典型代表。
杨树次生代谢产物的生物合成途径及调控机制在对杨树的次生代谢产物进行研究时,需要了解它们的生物合成途径及调控机制。
生物合成途径主要包括前体物质的来源、代谢途径及合成酶等。
例如,作为桂皮醇的先导物,苯丙氨酸是桂皮醇的重要前体。
而在苯丙氨酸的生物合成途径中,酚氧化酶和酪氨酸酶是两个非常关键的酶。
此外,调控机制在杨树次生代谢产物的生物合成中也起到了至关重要的作用。
研究表明,一些关键酶的表达水平与次生代谢物的合成水平有密切关系。
例如,在白杨的次生代谢产物中,苯丙氨酸氨联酶和cinnamyl醇可作为瓶颈酶调控桂皮醇生物合成。
针对关键酶的调控机制的研究有助于深入了解杨树次生代谢产物的生物合成机制。
杨树次生代谢产物的功能研究杨树次生代谢产物的生物化学性质决定了它们具备丰富的生物学功能。
例如,黄酮类次生代谢产物具有抗氧化、抗病毒、抗炎等生物活性,玫瑰醇则具有抗真菌、抗肿瘤等生物活性。
除此之外,杨树次生代谢产物的范围还拓展至植物生长和发育调控等方面。
杨树次生代谢产物的生物功能不仅可以被应用于医药、化妆品等领域,还有一定的农业价值。
例如,玫瑰醇和桂皮醇等次生代谢产物的研究发现,它们能够提高植物对环境的适应能力,促进植物生长。
这些研究有助于发掘杨树次生代谢产物在生命科学中更深层次的价值。
结语杨树次生代谢产物的生物合成及功能研究是生命科学领域的一个热门方向,它不仅有助于深入了解杨树次生代谢物的合成机制,还能够为开发新型天然活性物质提供重要参考。
杨树综述

杨树抗性基因工程研究进展林善枝肖基浒张志毅(北京林业大学毛白杨研究所)摘要杨树是世界上广泛栽培的重要造林树种之一,已成为林木基因工程研究的模式植物。
目前在杨树的抗除草剂、抗虫、抗病等生物抗性方面的研究已取得了成功,但在非生物抗性的研究相对较少,该文综述了国内外有关杨树抗性基因工程研究的最新进展,并提出了杨树抗性基因工程育种存在的问题。
关键词杨树抗性基因工程遗传改良杨树是世界上广泛栽培的重要造林树种之一,我国是杨树资源丰富的国家,从新疆到东部沿海,从黑龙江、内蒙古到长江流域均有分布,现已成为世界上杨树人工林面积最大的国家。
杨树因速生丰产、实用性强、分布广、无性繁殖能力强,且基因组较小而成为研究林木生理和利用基因工程方法进行遗传改良的理想模式植物。
但由于杨树具有生长周期长、树体高大等特点,极大地限制了杨树传统育种工作的开展。
也就是说,用常规育种的技术要在短时间内培育出人们所希望的杨树新品种是很困难的,尤其在改良杨树抗性性状方面更加困难。
随着世界人口迅速增长,生态环境破坏的日益加剧,以及地理和气候条件的限制,杨树抗性育种显得更加迫切,已引起国内外林木研究者的普遍关注。
因此查明杨树抗性的生理生化机制及其遗传因素,寻找提高抗性措施,尤其是利用基因工程技术进行杨树抗性育种(包括抗病虫、抗寒冻、抗旱、抗盐碱等),是当代林木分子生物学研究的重要课题之一。
它不仅在基础理论上具有重要意义,在解决生产实际问题上也具有广泛的应用价值。
植物抗性研究已有130多年历史,但林木抗性研究始于20世纪30年代,而真正利用基因工程技术进行杨树抗性育种在20世纪80年代中期才出现。
自从1986年parson等人证实了杨树可以进行遗传转化和外源基因在高等植物细胞中的表达以来,林木基因工程得到了迅速发展,尤其是杨树的基因工程进展最为迅速。
本文对近些年来国内外利用基因工程技术对杨树进行抗性遗传改良研究的现状进行了综述,并对其存在问题及发展前景进行了探讨。
杨树生殖生物学研究进展_张志毅

第22卷 第6期2000年11月北 京 林 业 大 学 学 报JOURNAL OF BEI J IN G FORESTR Y UN IV ERSIT YVol.22,No.6Nov.,20002000207218收稿http :///periodical/bjlydxxb/3“九五”国家攻关课题(962011202204202)和“高等学校青年骨干教师基金”项目资助杨树生殖生物学研究进展3张志毅 于雪松(北京林业大学毛白杨研究所,100083,北京;第一作者男,41岁,博士,教授)摘要 该文介绍了杨树(Populus spp.)生殖生物学的研究概况,包括:①杨树有性生殖过程中雌雄生殖系统形态结构和细胞化学变化及其生物学意义.②雄性不育的机理.③杨树杂交不亲和性的表现和原因及其克服的方法,花粉-柱头识别反应.④杨树生殖工程取得的成就.关键词 杨树,生殖生物学,有性生殖,雄性不育,杂交不亲和性,生殖工程中图分类号 S792.11Zhang Zhiyi ;Yu Xuesong.Advances in reproduction biolo gy of poplars.Journal of Beiji ng Forest ryU niversity (2000)22(6)69~74[Ch ,56ref.]Institute of Popul us tomentosa ,Beijing For.Univ.,100083,P.R.China.In this article ,the advances in reproduction biology of poplars are reviewed in four respects as fol 2lows :①The morphological and cytochemical changes and the biological significance of female and male reproductive systems in sexual reproduction of poplars ;②The mechanism of male sterility in poplars ;③The reasons and descriptions of cross incompatiblity in poplars ,the methods to overcome the cross incompatibility and the stigma pollen recognition reaction ;and ④The progress made in reproductive engineering in poplars.Key words Popul us ,reproduction biology ,sexual reproduction ,male sterility ,cross incompati 2bility ,reproductive engineering 植物生殖生物学是在植物胚胎学的基础上发展起来的.19世纪末,随着人们对植物性别认识的不断深入,逐渐形成了以描述植物生殖过程为主的植物胚胎学.应运而生的植物生殖生物学,不仅仅用植物学的知识和技术手段对植物生殖行为进行研究,并且已经渗透到解剖学、细胞学、生理学、生物化学、遗传学和分子生物学等多学科领域,产生了实验胚胎学、植物生殖生理学和植物生殖遗传学等分支学科.植物生殖生物学与植物育种学有着密切的关系.杨树具有速生等特点,广泛分布于欧、亚和北美洲,一般在北纬30°~72°之间[21].杨树为雌雄异株,偶有雌雄同株[28,31].目前林业生产上广泛应用的大多是人工培育的杂种无性系[21].杨树主要以无性繁殖为主,对杨树有性生殖研究较少.杨树生殖生物学的研究与农作物等草本植物相比,还处于较落后的水平.从70年代以来,随着生殖生物学理论和实验手段的不断丰富,杨树生殖生物学的研究进展很快,对杨树各派代表树种的整个有性生殖过程有了细致的了解[1~6,11,13,36,43];对杨树受精前后的组织化学变化及其生理意义进行了探究[16~17];从细胞化学和超微结构等方面对杂交不亲和性进行了研究[12,19,20,38,45,53];利用子房、胚和花粉离体培养成功地诱导出多种杨树植株[8~10,22].1 有性生殖过程1.1 花药的结构、小孢子发生与花粉的发育杨树的花药壁分为四层,由表及里依次为:表皮层、纤维层、中层和绒毡层[1,5,13].其中绒毡层对小孢子和花粉的发育有重要意义,主要是因为它分泌胼胝质酶能分解四分体胼胝质壁使小孢子分离,合成识别蛋白释放到花粉外壁,绒毡层的降解产物可作为花粉合成DNA 、RNA 、蛋白质和淀粉的原料.随着小孢子的发生和花粉的发育,腺质绒毡层细胞相应发生一系列变化[5,13].李文钿等(1988)在胡杨(P.euphratica Oliv.)花粉扫描电镜图象中发现,绒毡层与花粉壁之间有细丝相连,推测是孢粉素“桥”,并发现绒毡层的内切向壁的乌氏体,不随绒毡层细胞的退化而消失,而是有些散落在药室中,有些转移到花粉外壁上[11].花粉母细胞的减数分裂为同时型[1,5,6,11,24],四分体小孢子大多呈四面体形,少左右对称形[1,5,11,16].董源(1988)在毛白杨(P.tomentosa Carr.)小孢子中发现细胞自体消化细胞质的活动极为活跃,并有多层的同心圆膜状复合系统[3].刚形成的小孢子体积很快增大,渐渐变为圆形,细胞质增多,并开始液泡化,核增大,从中央移到边上[5].刘玉喜等(1979)在中东杨(P.beroli nensis Dippel.)中发现有多核和多核仁小孢子存在[24].经过短暂的休眠期,休眠期的长短与温度条件有关,毛白杨为2~7d[1].休眠期结束后,小孢子进行有丝分裂,由于纺锤体的不对称,造成分裂不均等,营养细胞明显大于生殖细胞,营养细胞内质体积累大量淀粉粒[6].樊汝汶(1983)和Rougier(1991)分别报道了美洲黑杨(P.deltoi des Marshall)雄性生殖单位(Male G erm Unit,M GU)在发育过程中细胞超微结构的变化[6,46].杨树成熟花粉属于22细胞型[1,5,11,13],球型,无萌发孔,外壁雕纹有皱波状、细网状、小穴状、颗粒状等多种形状[13,27].1.2 大孢子发生和胚囊的发育杨树一般为胚珠倒生,在发育的早期是双珠被,在大孢子母细胞减数分裂期内珠被滞育,并开始解体,最终成为单珠被[4,11,13,36],但大叶杨(sio2 carpa Oliv.)的内珠被不解体,一直为双珠被[18].造孢细胞直接分化为大孢子母细胞[5,36],细胞核大而明显,经减数分裂形成四分体,呈T形或直线形排列,靠珠孔端的三个大孢子退化,合点端的大孢子延伸最终发育成为八核胚囊[2,5,11,13,36],在欧美杨(P.euramericana)和胡杨中发现有时合点端第二个大孢子成为功能孢子[4,11].卵器穿透珠心伸入珠孔,卵细胞和助细胞极性相反[5,13].杨树胚囊的发育属于蓼型[2,5,11,13].1.3 受精过程杨树的花粉落到雌株的柱头上,一般几小时后开始在柱头上萌发,沿狭窄的花柱道进入子房[5,17,36,48,52],在此期间花粉管中的生殖细胞分裂成2个精子[13],花粉管经珠孔进入退化的助细胞,在此处或在卵细胞与次生核之间释放其内含物,进行双受精[2,13,17,36,48].反足细胞是“短命”的,受精后很快就退化[5,36].整个受精作用要消耗大量淀粉粒[17].关于杨树受精过程中精卵结合的详细情况报道不多,对于雌雄性细胞的融合机制,雄性细胞质在受精过程中的作用,卵细胞受精前后生理生化变化等,有待于进一步深入探讨.董源(1984)认为毛白杨的受精作用是介于有丝分裂前型和有丝分裂后型之间的中间型,精子起初整个贴附在卵核的核膜上,处于静止期,随后精子进入卵核中,先是核仁合并,接着核质融合[2].朱彤等(1989)发现大叶杨的受精作用为有丝分裂前型.Russel(1990)对美洲黑杨受精前的精子进行了研究,发现精子在退化的助细胞发生液泡化,细胞质稀释,细胞核染色体浓缩,细胞质减少,核质分离[48].1.4 胚乳和胚的发育精核与次生核融合后,形成大的富含DNA、RNA和蛋白质的初生胚乳核,一般不经过休眠,直接开始初生胚乳核的分裂,比受精卵的分裂早[2,5,13,17,36,56],但在毛白杨中发现,有时受精卵已经形成八细胞原胚,初生胚乳核尚处于休眠状态[2].胚乳在胚的发育过程中被吸收,故成熟种子不含胚乳[5,13,17,36].杨树胚乳发育类型属于核型[2,5,13,17].精卵融合后,进入合子休眠期,核内DNA、RNA 含量上升[17].大叶杨和小叶杨(P.si monii Carr.)的合子休眠期分别为6~8d[17]和6~10d[13].合子在休眠期并不是静止的,而是进行着十分活跃的生理过程,在大叶杨中发现,合子经历了液泡消失、合子皱缩、液泡再现、合子伸长等极性化过程[17].在杨树属各派树种中大叶杨的胚胎学特征十分特殊(见表1),大叶杨胚胎发育类型为茄型,而其他各派均为柳叶菜型[5,13].1.5 有性生殖过程的组织化学植物有性生殖过程中,除了形态和结构发生变化外,各种物质(DNA,RNA,蛋白质,淀粉粒和胼胝质)的含量和分布也发生相应的改变.中国林科院朱彤等(1988)首次报道了杨树小孢子发生过程中的组织化学变化,发现大叶杨小孢子母细胞的营养源位于表皮和纤维层,绒毡层细胞和小孢子母细胞逐渐积累多糖,绒毡层细胞始终含丰富的DNA、RNA和蛋白质,RNA和蛋白质在小孢子发生过程中表现一致性的分布和含量变化[16].在小孢子发生过程中,蛋白质的作用主要是作为功能蛋白和结构蛋白参与代谢,淀粉粒则作为发育中所需的营养来源[11,16].PAS反应表明,胡杨单核花粉晚期细胞质中开始积累淀粉粒,在22细胞花粉的生长过程中,营养细胞内的淀粉粒日渐增多并增大,花粉散出前夕,淀粉粒和其他内含物的积累达到高峰[11].小孢子母细胞在减数分裂前期I开始沉积胼胝质,并逐渐加厚,到四分体后期胼胝质壁解体,释07北 京 林 业 大 学 学 报第22卷 放出小孢子[11].表1 杨树各派代表树种胚胎学观察结果TABL E1The embryological observation of representative species in various poplar sections 派名种名心皮种子珠被胚胎发生类型胚乳胚囊受精所需时间参考文献大叶杨派大叶杨2~3双茄型核型蓼型6~10d17,18青杨派小叶杨2~34~6单柳叶菜型核型蓼型3~6d13,21黑杨派美洲黑杨3~44~32单柳叶菜型核型蓼型2~3d43,48欧洲黑杨2单柳叶菜型核型蓼型31白杨派毛白杨21~2单柳叶菜型核型蓼型40~48h2响叶杨2单柳叶菜型核型蓼型5~7d5美洲山杨2单柳叶菜型核型蓼型43银白杨2~43~4单柳叶菜型核型蓼型31胡杨派胡杨3单柳叶菜型核型蓼型11 大叶杨的大孢子母细胞、四分体及功能大孢子中含不溶性多糖较少,含RNA和蛋白质丰富,成熟胚囊中除反足细胞外充满淀粉粒,围绕在卵核及次生核周围[18],助细胞具多糖性质的丝状器,胚囊的营养来源于子房和胎座细胞内贮存的淀粉粒,卵细胞PAS反应呈阴性.受精后,胚胎发育过程中, DNA、RNA和蛋白质含量和分布发生一系列变化,游离胚乳核中RNA和蛋白质含量较高,不含可溶性多糖及淀粉粒,围绕在卵核及次生核周围的淀粉粒迅速消失,消耗的淀粉一方面水解作为能量的来源,另一方面为合子多糖性质的细胞壁的合成提供原料,合子PAS反应呈阳性[17].Label(1994)发现欧洲黑杨(P.nigra L.)受精前后子房内的ABA含量不变[40].2 雄性不育雄性不育是由于植株不能产生正常的花药、花粉或雄配子,表现形式有:不能形成正常的小孢子发生组织,小孢子发生异常,形成不完善、不能存活的、畸形或败育的花粉,花粉不能成熟或无萌发能力,花药不开裂.花粉发育的每个时期都可能发生败育.雄性不育是植物界中一种普遍现象,这方面的研究报道很多,但大多以农作物为研究对象,对树木雄性不育机理的研究不多.樊汝汶等(1992)认为中国鹅掌楸花粉败育是遗传因子造成的,导致小孢子母细胞不能正常启动减数分裂或减数分裂异常.杨树雄性不育机理的研究主要停留在对形态结构的观察,尚未深入到分子遗传水平.目前认为,杨树雄性不育的原因主要是绒毡层发育异常.由于绒毡层是花粉粒发育时的营养来源,它的异常发育将导致绒毡层无法为花粉的发育提供适当的营养,使花粉处于饥饿状态.李文钿等(1982)在小叶杨中同时观察到绒毡层的早期败育和延迟败育,在出现早期败育的绒毡层花粉囊中可以看到败育的花粉母细胞不正常的减数分裂和败育的小孢子四分体;在绒毡层延迟退化的花粉囊中,往往有败育的粘连成片的空壳花粉和畸形的多核大花粉[13].董源(1982)认为毛白杨雄性不育是由于绒毡层过度增生抑制了花粉的发育,绒毡层异常通常发生在小孢子母细胞和四分体时期.败育的四分体无核或核不明显,染色极深[1],并认为低温能诱导雄性不育.康向阳(1996)则认为绒毡层的异常发育只是毛白杨雄性不育的一种表象,其根源在于毛白杨染色体的异源性,导致减数分裂时同源染色体不均衡分配[26].纤维层内壁和侧壁加厚,有利于失水后花粉囊的裂开和散粉,纤维层的加厚异常将导致花药不开裂[13].3 种间杂交不亲和性探索种间杂交不亲和性的原因,是植物生殖生物学中一个很重要的内容,为植物杂交育种工作的深入开展提供了条件.70年代以来,国内外针对杨树种间杂交不亲和性进行了大量的研究[12,19,47,51],揭示了一些种间杂交不亲和性的原因.如花粉萌发率很低[35,37],花粉2柱头不亲和,花粉管不能进入柱头12,32];花柱不亲和,花粉管无法达到子房[32,35];胚囊发育迟缓,花粉管无法进入胚囊[12];杂种胚乳发育不正常导致杂种胚死亡[12]等.不亲和性的表现主要有花粉及花粉管分泌物少,柱头表面和花粉管前端沉积胼胝质[32,41],花粉管在柱头上表现异常,扭曲、盘绕、爬行[19,41,55],柱头细胞提前解体[19],胚乳和胚败育[12,41,55]等.朱彤等(1989)认为柱头细胞的提取解体退化是雌蕊拒绝反应的一部分,表现为细胞质解体,细胞内部液泡化,膜结构逐渐消失以及细胞质电子密度增大.花粉落到柱头上后,花粉2柱头间将发生主动识别反应[38],柱头一旦识别出了不亲和的花粉壁蛋17第6期张志毅等:杨树生殖生物学研究进展 白,通常在乳突细胞内迅速产生胼胝质阻止花粉管的继续侵入[32].Knox(1972)首次在杨树的种间杂交研究中提出了主动识别(positive recognition)的概念,他在美洲黑杨×银白杨的杂交中发现,银白杨花粉与美洲黑杨柱头不亲和是由于绒毡层组织合成的一种特异蛋白,授粉后被释放出来,与柱头上的特异蛋白进行相互识别,并产生一系列反应,从而决定了花粉管能否进入柱头实现受精[37].花粉2柱头相互作用这一识别过程涉及多种酶系的活动,在花粉外壁识别蛋白中常见酯酶、过氧化物酶、酸性磷酸酶和转移酶类等酶系活动,在柱头表膜中有非特异性酯酶、酸性磷酸酶及过氧化物酶等酶系活动[20,30].李文钿等(1990)首次在杨树中报道了柱头表膜A TP酶的活动,发现大叶杨柱头A TP 酶与花粉2柱头相互作用有一定相关性,A TP酶的活性随柱头可授期的到来而增加,幼嫩期和过盛花期的柱头薄膜上A TP酶活性低或没有[20].Villar (1993)发现亲和授粉后(欧洲黑杨×欧洲黑杨)β2半乳糖苷酶的活动上升,而在不亲和杂交组合中(欧洲黑杨×银白杨),β2半乳糖苷酶未表现活性上升,认为β2半乳糖苷酶在花粉管的异养生长中起重要作用,与花粉2柱头相互作用有关.Rougier(1988)在美洲黑杨×银白杨的不亲和杂交中发现,腺苷酸环化酶(CAMPase)的活性比亲和杂交的活性低,认为CAMPase与花粉在柱头上粘附、吸水及萌发有关,活性下降是配子体表型不亲和的细胞化学表现之一[45].G aude(1982)用血细胞凝集技术研究了欧美杨花粉外壁可扩散的细胞粘附分子(CAMS),推测CAMS可能是一种类外源凝集素,与花粉2柱头识别反应有关[34].总之,对这些酶类的研究为探讨不亲和性机理以及寻求克服方法提供了新的思路和途径.杨树杂交中的单向不亲和性较为普遍,对于单向不亲和性的原因尚无一致意见.Villar et al (1987)认为欧洲黑杨×银白杨的单向不亲和性是由于两个种的柱头类型不同造成的,欧洲黑杨具有干柱头的特征,表面有一层薄膜,而银白杨是湿柱头,有丰富的分泌物[53].这与李文钿等(1995)的研究结果不同,李文钿认为杨树柱头都是湿型分泌柱头[15].克服杂交不亲和性的方法很多,如花粉蒙导、用生理活性物质处理雌蕊、重复授粉、受精工程等.在杨树杂交中主要采用蒙导花粉授粉,使柱头不能识别不亲和的花粉,以克服种间杂交不亲和性,获得杂种[33,49,50].美国华盛顿大学的Stettler教授在这方面作了许多工作[49,50,51].4 生殖工程目前,杨树生殖工程的研究主要是以子房、胚珠和花药离体培养为主,已成功地诱导出生长正常的植株.杨树杂交胚胎学的研究表明,杨树一些种间杂交的障碍不是发生在受精之前,而是发生在受精之后,胚在发育过程中败育[12,14,41],因此利用子房和胚珠离体培养可以克服种间杂交胚的败育.迄今为止,利用子房和胚珠培养诱导植株成功的有:小叶杨×美杨,小叶杨×胡杨,小叶杨×大叶杨,欧洲黑杨×大叶杨,大叶杨,银白杨,欧洲黑杨,小黑杨(P.×si monigra Chon2Lin)等[7,29,39,44].李文钿等(1983)对不同发育阶段小叶杨×美杨杂种胚的胚珠进行离体培养,认为离体胚珠的最适接种时间是在大多数杂种胚尚未败育之前,胚龄越大,越容易成活[14].70年代以来,相继成功地利用花药离体培养诱导出多种杨树花粉植株,有:小叶杨×美杨、小叶杨×黑杨、北京杨、中东杨、美杨、美杨×青杨、大青杨、欧洲黑杨、欧洲黑杨×小叶杨、银白杨×小叶杨、美杨×欧洲黑杨、箭杆杨(P.thevesti na Dode)×欧洲黑杨、胡杨等[8~10,22,42],并对花粉植株的倍性和育性进行了研究,杨树花粉植株一般是混倍体和非整倍体,随着树龄的增大,自行调整和加倍到二倍体[22,23].刘玉喜(1986〕发现9~10年生的小叶杨×欧洲黑杨花粉植株的花粉母细胞减数分裂正常,能产生有活力的成熟花粉,授粉后能产生饱满种子[23].一般认为杨树花粉愈伤组织的诱导以单核期接种最为适宜,有利于花粉进行异常分裂和形成多细胞花粉[9].离体培养条件下,雄核发育以均等分裂为主,非均等分裂和多核花粉类型较少,并很快解体[9].5 结 语纵观杨树生殖生物学研究发展情况,它已经形成了初步的理论体系,研究重点从单纯的形态描述转向对结构与功能关系的综合研究.70年代以来,应用电镜和细胞化学等生物新技术,研究探讨了杨树有性生殖过程中某些结构和生理功能的关系,诸如花粉壁和柱头表面的结构成分与花粉2柱头相互识别的关系,绒毡层细胞结构成分与雄性不育的关系等.同时,我们还应看到在杨树生殖生物学的研究中,还有许多急待解决的问题,如雄性不育的机理,受精作用中不亲和性的机制,配子体形成过程中的遗传控制机制和配子体的基因表达等.我们相信,随着对这些问题的深入了解和认识,对杨树有性生殖27北 京 林 业 大 学 学 报第22卷 过程(传粉、受精和胚胎发生等)的完全控制将会成为现实,这必将会使杨树育种工作更紧密地朝着人类所期望的方向发展.参考文献1 董源.毛白杨胚胎学观察I:花药的结构及花粉的发育.北京林学院学报,1982,4(4):80~922 董源.毛白杨胚胎学观察II:胚珠、胚囊的构造、受精作用和胚的发育.北京林学院学报,1984,6(1):83~943 董源.毛白杨小孢子发育的电子显微镜研究初报.北京林业大学学报,1988,10(增刊):109~1114 樊汝汶.黑杨派无性胚珠和胚囊发育的比较.南京林学院学报,1984(3):44~505 樊汝汶.响叶杨种子发育的胚胎学观察.南京林产工业学院学报,1982(3):116~1286 樊汝汶.美洲黑杨无性系I263花粉粒的发育和超微结构.南京林产工业学院学报,1983(3):13~187 李文钿,李江山.杨树杂种胚珠的离体培养.林业科学,1985, 21(4):339~3468 东北林学院树木育种组.杨树单倍体花粉植株的诱导.遗传学报,1977,4(1):49~549 王敬驹.杨树花粉植株的诱导.植物学报,1975,17(1):56~59 10 黑龙江省林业科学院林业研究所.花药离体培养诱导杨树单倍体植株.遗传学报,1976,3(2):145~14911 李文钿,朱彤.胡杨花粉和胚囊的发育.林业科学研究,1988,1(2):132~13712 李文钿,朱彤.胡杨与小叶杨远缘杂交不亲和性的障碍.林业科学,1986,22(1):1~913 李文钿.小叶杨种子发育的胚胎学观察.林业科学,1982,18(2):113~11914 李文钿.小叶杨与美杨种间杂交的胚胎发育和杂种胚珠的离体培养.植物学报,1983,25(5):409~41715 李文钿.杨树的分泌柱头:发育、细胞化学、超微结构及其与组间杂交不亲和性的关系.植物学报,1995,37(7):514~52116 朱彤,李文钿.大叶杨小孢子发生的组织化学观察.林业科学研究,1988,1(3):289~29317 朱彤,李文钿.大叶杨的受精作用和胚胎发育.植物学报,1989, 31(5):355~36018 朱彤,李文钿.大叶杨胚囊及胚珠的形成和发育.武汉植物学研究,1989,7(1):13~2019 朱彤,李文钿.大叶杨组组间杂交花粉柱头相互作用的超微结构研究.植物学报,1988,30(1):14~2020 朱彤,李文钿.大叶杨柱头细胞ATP酶的超微结构定位.植物学报,1990,32(2):91~9621 徐纬英主编.杨树.哈尔滨:黑龙江人民出版社,198822 朱湘渝.杨树花粉植株的诱导.林业科学,1980,16(3):190~19723 刘玉喜.杨树花粉植株的育性研究.东北林业大学学报,1986, 14(1):20~2524 刘玉喜,陆志华.中东杨小孢子的发生.东北林学院学报, 1979,(2):1~425 张 方.杨树花药培养中小孢子发育的细胞学观察.东北林学院学报,1979,(2):5~1426 康向阳.毛白杨细胞遗传与三倍体选育的研究:[学位论文].北京:北京林业大学图书馆,199627 张绮纹.杨树各派代表树种花粉粒表面微观结构研究.林业科学,1988,24(1):76~7828 张志毅,黄智慧,张东方,等.毛白杨标本园无性系开花结实的研究.北京林业大学学报,1992,14(增刊):43~5129 吴克贤,徐妙珍.从杨树为授粉子房培养出母系单倍体植株.科学通报,1983,15:960~96330 Ashford A E and Knox R B.Characteristics of pollen diffusates and pollen2wall cytochemistry in poplars.J Cell Sci,1980,44:1~18 31 Boes T K and Strauss S H.Floral phenology and morphology of black cottonwood,Popul us t richocarpa(Salicaceae).Am J Bot, 1994,81(5):562~56732 G aget M.Pollen2pistil interaction in interspecific cross of Popul us (Section Aigeiros and Leuce):pollen adhesion,hydration and cal2 lose responses.J Cell Sci,1984,72:173~18433 G aget M.The mentor pollen phenomenon poplars:a new concept.Theor Appl G enet,1989,78(1):129~13534 G aude T.Are lectin2like compounds involved in stigma2pollen ad2 hesion and/or recognition in Popul us and B rassica?In Pollen:bi2 ology and implications for plant breeding.Mulcahy D L and Otta2 viano E〔ed.〕,1982,265~272,Italy35 Guries R P and Stettler R F.Pre2fertilization barriers to hybridiza2 tion in the poplars.Silvae G enetica,1976,25(2):38~4436 Khurana D K.Ontogeny of catkin2drop and embryology of Popu2 l us ciliata and its crosses with P.deltoi ds.In Improvement of Forest Biomass,Khosla P K ed.,India,198037 Knox R B.Role of polen2wall proteins as recognition substances in interspecific incompatibility in poplars.Nature,1972,237:381~38338 Knox R B.Interspecific hybridization of poplars using recognition pollen.Silvae G enetica,1972,21:65~6939 K ouider M.A method to culture immature embryos of Popul us deltoi des in vitro.Can J For Res,1984,14:956~95840 Label P.EL ISA quantitation and GC2MS identification of abscisic acid in stigma,ovary and pedicel of pollinated poplar flowers(Pop2 ul us nigra L.).Tree Physiology,1994,14(5):521~53041 Li W D.The behavior of pollen tubes on the stigma in the intersec2 tional crosses in Popul us and its relation to seed2setting.Chin J Bot,1991,3:102~10942 Mofidabadi A.Callus induction and haploid plant regeneration from anther culture of two poplar.Silvae G enetica,1995,44(2~3): 141~14543 Nagaraj M.Floral Morphology of Popul us deltoi des and P.t rem uloi des.Bot G az,1952,114:222~24344 Raquin C and Troussard L.In2ovary embryo culture as a tool for poplar hybridization.Can J Bot,1993,71(10):1271~127545 Rougier M.Localization of adenylate cyclase activity in Popul us: its relation to pollen2pistil recognition and incompatibility.Sex P Reprod,1988,1(3):140~14946 Rougier M.Male gametophyte development and formation of the male germ unit in Popul us deltoi des following compatible pollina2 tion.Protoplasma,1991,162(2~3):140~15047 Rougier M.Interspecific incompatibility in Popul us:inhibition of tube growth and behaviour of the male germ unit in P.deltoi des×37第6期张志毅等:杨树生殖生物学研究进展 P.alba.Protoplasma,1992,168(3~4):107~11248 Russel S anizationof the early post2fertilization megagame2 tophyte of Popul us deltoi des:ultrastructure and implication for male cytoplasmic transmission.Protoplasma,1990,155:153~16549 Stettler R F.Irradiated Mentor Pollen:its use in remote hybridiza2 tion of black cottonwood.Nature,1968,219:746~74750 Stettler R F and Guries R P.The mentor pollen phenomenon in black cottonwood.Can J Bot,1976,54:820~83051 Stettler R F.Interspecific crossability studies in Poplars.Theor Appl G enet,1980,58(6):273~28252 Villar M.The route of the pollen tube from stigma to ovule in Popul us nigra:A new look.Ann Sci For,1987,44(2):259~26453 Villar M.Incompatibility in Popul us:Structural and cytochemical characteristics of the receptive stigmas of P.alba and P.nigra.J Cell Sci,1987,87:483~49054 Villar M.Pollen2pistil interaction in Popul us:β2galactosidase ac2 tivity associated with pollen2tube growth in the crosses P.nigra×P.nigra and P.nigra×P.alba.Sex P Reprod,1993,6(4): 249~25655 Von Melchior G H.Interspecific cross sterility within the poplar section.Silvae G enetica,1968,17(2~3):88~9256 Winton L L.Fertilization in forced quaking aspen and cottonwood.Silvae G enetica,1968,17(1):20~21(责任编辑 胡 涌)敬 告 读 者20多年来,在各位作者的热情支持下,《北京林业大学学报》的稿源日趋丰富,学术质量稳步提高.对此,我们向全体给予我们厚爱的作者表示衷心的感谢.由于《北京林业大学学报》是以刊登学术论文和研究报告为主的国家核心期刊,每期在“综合述评”栏目里只能刊登2~3篇文章,全年只能刊登12~15篇文章.要求作者要有一定的科学研究经历,能对学科发展进行权威、科学、客观的总结、分析评论和预测.主要的征稿对象是全国有关院校和科研单位的学科带头人、骨干教师和科研人员、博士后及少部分有独到见解的研究生.从2001年起,《北京林业大学学报》原则上不再发表以博士生和硕士生的“文献综述课”作业或开题报告内容为主的“综合述评”稿件.特此通告《北京林业大学学报》编辑部2000111110 47北 京 林 业 大 学 学 报第22卷 。
从初生生长到次生生长:植物维管形成层发育的调控机制

从初生生长到次生生长:植物维管形成层发育的调控机制王国栋;宋银
【期刊名称】《中学生物教学》
【年(卷),期】2011()11
【摘要】从初生生长到次生生长,是高等植物重要的生长发育过程,在这一生长发育过程中受到很多遗传与内源因子的调控,特别是一些转录因子,受体激酶、类受体蛋白及多肽激素等参与了这一过程的调控。
本文着重介绍了近年来受体激酶、类受体蛋白及多肽激素对植物维管形成层发育调控机制的认识,在此基础上提出了以后的工作应侧重于形成层中受体激酶、类受体蛋白和多肽激素的研究,并尝试阐明某些已知或未知的CLE多肽和(或)受体激酶及类受体蛋白是如何参与调控形成层细胞的增殖和分化并进一步如何影响维管系统的发育。
【总页数】3页(P4-6)
【关键词】维管形成层;类受体蛋白;受体激酶;CLE多肽;调控机制
【作者】王国栋;宋银
【作者单位】陕西师范大学生命科学学院
【正文语种】中文
【中图分类】G78
【相关文献】
1.研究拟南芥生长发育调控以及抗病机制为农作物改良提供理论依据——浙江大学生命科学学院植物生物学研究所徐娟副教授 [J], 刘强
2.[生物化学]上海生科院揭示环境信号协同调控植物生长发育机制 [J],
3.茎部形成层赤霉素在植物生长发育中的调控作用 [J], 杨钧贺;刘畅;钮世辉;李伟
4.GRF转录因子对植物生长发育及胁迫响应调控的分子机制 [J], 马超;原佳乐;张苏;贾琦石;冯雅岚
5.我国科学家发现光调控植物发育新机制阳光对植物生长速度和形态影响最大 [J],因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
LBD1-30基因调控杨树茎干次生生长的功能分析次生生长是多年生树木木材形成的生物学基础,在此过程中形成层(vascular cambium)干细胞(stem cell)经过连续的细胞分裂和分化、细胞扩张、次生细胞壁合成以及细胞程序化死亡等过程,向外产生次生韧皮部(secondary phloem),向内产生次生木质部(secondary xylem)。
目前对次生生长的研究大多集中于木质部表达量高于韧皮部的基因,对于韧皮部表达量高于木质部的基因的研究较少,本研究将以后者作为出发点,通过转基因工程来研究基因对次生生长的调控。
通过对前期的RNA-seq结果进行分析数据,我们分析了杨树茎段木质部和韧皮部的全基因表达水平,发现了LBD基因家族中在植物次生维管组织中显著差异表达的基因,并重点分析在韧皮部表达量高于木质部的基因。
本研究以84K杨茎段为材料,克隆了杨树LBD1-30基因(Potri.008G043900),构建了35S:LBD1-30的过表达载体,利用农杆菌介导的转化体系,将LBD1-30基因转入84K杨,并通过表型分析和切片分析等初步验证了LBD1-30对杨树次生维管组织发育的影响,证明其对杨树次生生长具有显著的调控作用。
该研究为今后培育速生高产的杨树新品种提供了重要的现实意义和理论价值。
主要研究结果如下:(1)LBD1-30基因的克隆及序列分析。
从毛果杨基因数据库中查询出LBD1-30基因的CDS序列,其包含的CDS序列总长度为666bp,编码氨基酸数量为221个。
设计引物,并用84K杨茎段为材料提取总RNA,利用RT-PCR技术克隆了LBD1-30基因。
比对分析测序结果,84K杨中LBD1-30的编码序列与毛果杨参考序列相比有多个单碱基突变(SNP),但是蛋白序列有着96.4%的相似度。
(2)构建了
35S:LBD1-30的过表达载体,并成功地转入了84K杨中。
通过农杆菌介导的转化方法转化84K杨。
将愈伤组织产生的生根转基因苗切头转入新的含有30mg/L Kana的生根培养基上再进行筛选,选择生根的转基因苗提取DNA进行PCR鉴定,最后获得转基因的阳性植株。
(3)转基因苗中LBD1-30基因的表达水平定量分析和表型分析。
转基因苗中的LBD1-30基因的表达量远远高于野生型,并且不同株系之间的表达量存在明显差异。
转基因植株在表型上可分为高粗型和矮粗型,且基因表达量越高表型越倾向于矮粗型,但这种转基因株系生根慢甚至不生根,较难准备材料。
高粗型株系基因表达量略低于矮粗型,但生根及生长和野生型无异,后期生长量大。
(4)LBD1-30基因对次生生长的调控。
与野生型相比,LBD1-30过表达转基因苗中次生韧皮纤维木质化增加,而次生木质部木质化则被抑制,甚至出现明显的不规则木质化。
表明LBD1-30基因在次生生长过程中,对形成层干细胞的分裂增生和次生木质部/次生韧皮部的分化过程可能发挥重要的调控作用。