原核基因表达调控知识梳理
分子生物学第七章原核生物基因表达调控

原核生物基因表达调控的特点
01
原核生物基因表达调控通常由特 定的转录因子、RNA聚合酶以及 其他调控蛋白介导,通过与DNA 的结合或解离来调节基因转录。
02
原核生物基因表达调控具有快速 响应环境变化的特点,能够在短 时间内调整基因表达模式,以适 应外界刺激和压力。
翻译后加工的调控
翻译后加工的调控
在翻译后加工阶段,新合成的蛋白质经过一系列修饰和加工,最终成为具有生物学活性的蛋白质。原 核生物通过控制翻译后加工酶的合成和活性来调控翻译后加工过程。此外,原核生物还可以通过控制 蛋白质的稳定性来影响其功能和表达水平。
总结
翻译后加工是基因表达调控的重要环节,原核生物通过控制翻译后加工酶的合成和活性,以及蛋白质 的稳定性来精细调控基因表达。
翻译延伸的调控
翻译延伸的调控
在翻译延伸阶段,核糖体沿着mRNA移动,将氨基酸组装成蛋白质。原核生物通过控制翻译延伸因子的合成和活 性,以及核糖体的合成和组装来调控翻译延伸。此外,原核生物还可以通过控制mRNA的结构和稳定性来影响翻 译延伸。
总结
翻译延伸是基因表达调控的重要环节,原核生物通过控制翻译延伸因子的合成和活性,以及核糖体的合成和组装, 以及mRNA的结构和稳定性来精细调控基因表达。
翻译起始的调控
原核生物通过控制翻译起始来调控基因表达。在翻译起始阶段, mRNA与核糖体结合,招募翻译所需的起始因子和其他成分。原 核生物通过控制起始因子的合成和活性,以及mRNA与核糖体的 结合来调控翻译起始。
总结
翻译起始是基因表达调控的重要环节,原核生物通过控制翻译起 始因子的合成和活性,以及mRNA与核糖体的结合来精细调控基 因表达。
第六章 原核基因表达调控
第六章 原核基因表达调控模式
-- 乳糖(+)时, 葡萄糖(+)
二.乳糖操纵子调节机制
i基因
有诱导物时
P
O
Lac Z
Lac Y 转录
lac A
阻遏蛋白 阻遏物 蛋白亚基 无活性的 阻遏蛋白
翻译 转录水平低, 没有乳糖酶的合成 β- 半乳糖苷酶 β- 半乳糖苷通透酶 β- 半乳糖苷乙酰转移酶
mRNA
诱导物
第六章 原核基因表达调控模式
一.
概述
根据细菌酶的合成对环境的反应不同,分为:
组成酶 细菌酶 诱导酶
适Байду номын сангаас酶
阻遏酶
第六章 原核基因表达调控模式
3. 原核基因表达的多级调控
四个基本调控点: 基因活化 转录水平上的调控
最有效的调节环节
一.
概述
转录起始--- DNA元件与调控蛋白相互作用调控 mRNA加工成熟水平的调控 转录后水平上的调控 翻译及翻译后水平的调控
调节基因的产物-阻遏蛋白
负控诱导 负控阻遏 正控诱导
调控机制
负转录调控 (为主) 正转录调控
正控阻遏
调节基因的产物-激活蛋白
第六章 原核基因表达调控模式
负调控 Lac O 正调控 Ara O
一.
概述
诱
导 阻遏物 阻遏 诱导物 诱导
失活的阻遏物 失活的活性蛋白 阻遏
活化的激活蛋白 诱导物 诱导
Trp O
(1) 葡萄糖(+), 乳糖(–)时,
- 乳糖(–)时, 无别乳糖存在,阻遏蛋白与操纵子上的 O序列结合, 使操纵子处于关闭状态,三个结构基因 不表达。 - 葡萄糖(+)及cAMP浓度低时,CAP 活性低, 无 cAMP- CAP复合物形成。
原核生物基因表达调控分析

Co-repressor
(共阻遏物)
原核生物基因表达调控方式:
负控诱导调节
负控转录调 控系统
调节基因的产物是 阻遏蛋白 (repressor), 阻止了结构基因的 转录。
阻遏蛋白与效应物(诱 导物)结合,使阻遏蛋 白失活,结构基因转录; 阻遏蛋白与效应物(辅阻 遏物)结合,使阻遏蛋白 产生活性,结构基因不转 录。
operon on operon off operon off operon on
Neg.
i- or 不加入I基因产物 I+ or 加入I基因产物
(激活蛋白)
Pos.
●
Repressor binding on O site 阻遏蛋白 阻止转录启动
Expressor binding front p site
安慰诱导物:
如果某种物质能够诱导细菌产生某种酶而本身又不
被分解,这种物质被称为安慰诱导物,如IPTG(异
丙基- β –D-硫代半乳糖苷)。 相反,随环境条件变化而基因表达水平降低的现象 称为阻遏(repression),相应的基因被称为可阻遏的基 因(repressible gene)。 如果某种物质能够阻止细菌产生合成这种物质的酶, 这种物质就是辅阻遏物。(合成代谢)
第一讲 原核生物基因表达 调控
主要内容
一、基因表达调控的基本概念: 二、 基因表达调控的理论与模式;
一、基因表达调控的基本概念:
1、基因表达调控的意义: 原核生物对环境的适应、对营养条件改变适应的 相关应答,都是基因表达的结果;
真核生物的细胞分化, 组织特化 , 个体发育以及 环境对个体表型的影响都是通过基因表达实现的。
组成型突变: lacOc
iC mut. (iC O+P+) constitutive mut. (组成型)
第14章 原核生物基因表达调控

第14章原核生物基因的表达调控重点:操纵子的结构特点和功能;乳糖操纵子的正负调控;色氨酸操纵子的衰减作用。
难点:色氨酸操纵子的衰减作用。
第一节基因调控的基本定律一、基因调控水平二、基因和调控元件三、DNA结合蛋白一、基因调控水平基因表达的调控可以发生在DNA到蛋白质的任意节点上,如基因结构、转录、mRNA 加工、RNA的稳定性、翻译和翻译后修饰。
二、基因和调控元件基因:是指能转录成RNA的DNA序列。
结构基因:编码代谢、生物合成和细胞结构的蛋白质。
调节基因:产物是RNA或蛋白质,控制结构基因的表达。
其产物通常是DNA结合蛋白。
调控元件:不能转录但是能够调控基因表达的DNA序列。
三、DNA结合蛋白调控蛋白通常含有与DNA结合的结构域,一般由60-90个氨基酸组成。
在一个结构域中,只有少数氨基酸与DNA接触。
这些氨基酸(包括天冬氨酸、谷氨酸、甘氨酸、赖氨酸和精氨酸)常与碱基形成氢键,或者与磷酸核糖骨架结合。
根据DNA结合结构域内的模体,可以将DNA结合分成几种类型(图16.2)。
第二节大肠杆菌的乳糖操纵子一、操纵子结构二、正负调控三、乳糖操纵子四、lac突变五、正控制一、操纵子结构原核和真核生物基因调控的主要差异在于功能相关的基因的组成。
细菌的功能相关的基因常常排列在一起,并且由同一启动子控制。
一群一起转录的细菌的结构基因(包括其启动子和控制转录的额外序列)称为操纵子。
二、正负调控转录水平上的调控主要有两种类型:负调控:gene ON 阻遏蛋白 OFF正调控:gene OFF 激活蛋白 ON诱导:活性阻遏蛋白 失活诱导因子+非活性激活蛋白 活性阻遏:失活阻遏蛋白 活性共阻遏蛋白+活性激活蛋白 失活三、乳糖操纵子乳糖操纵子是诱导型操纵子,当诱导物不存在时,阻遏蛋白结合到操纵序列上并阻止转录;当诱导物存在时,阻遏蛋白与诱导物结合后失去活性,转录才得以进行。
四、lac突变为了鉴定乳糖操纵子各个成分的功能,Jacob和Monod做了细菌的接合实验,其中供体菌的F’因子上也带有乳糖操纵子。
原核生物基因表达调控

Repressor
cAMP
CAP
葡萄糖不存在,乳糖存在,阻遏蛋白失活,cAMP+CAP与CAP位点结合结合,促进基因转录
The Lac Operon: III. 葡萄糖和乳糖都存在
Repressor
RNA Pol.
CAP Bindin
g
Promoter
Operator X
LacZ
Repressor负调节与正调节协调合作
• 阻遏蛋白封闭转录时,CAP不发挥作用 • 如没有CAP加强转录,即使阻遏蛋白从操作基因上解聚仍无转录活性
3)正调控和负调控
正调控(positive control)
在没有调节蛋白质存在时基因是关闭的,加入某种调节蛋白后基因活性就被开启,这样的调控为正转录 调控。
调节基因
操纵基因
结构基因
调节蛋白
mRNA 酶蛋白
负调控(negative control)
在没有调节蛋白质存在时基因是表达的,加入这种调节蛋白质后基因表达活性便被关闭,这样的调 控负转录调控。
2)结构基因和调节基因
➢ 组成基因/管家基因(constitutive gene, housekeeping gene)是指不大受环境变动而持 续表达的一类基因。如DNA聚合酶,RNA聚合酶等代谢过程中十分必需的酶或蛋白质的基因 。 ➢调节基因(regulated gene)指环境的变化容易使其表达水平变动的一类基因。如:不同生 长发育时期表达的一些基因。
• 别乳糖是lac操纵子转录的活性诱导物 • 异丙基硫代半乳糖苷(isopropyl thiogalactoside:IPTG)结构上类似于别乳糖,是乳糖操纵
子非常有效的诱导物。可诱导lac操纵子表达,但不能被β-半乳糖苷酶水解。 • 这种能诱导酶合成,但不能被酶分解的分子称为安慰诱导物(gratuitous inducer)。安慰诱导
第九章:原核生物基因表达调控

抗σ因子与抗抗σ因子
9.1.1.3 双组分调节系统
双 组 分 调 节 系 统 的 组 成
感应激酶 应答调节子
周质结构 域、 胞质结构 域
PhoR和PhoB构成的双组分调节系统
天冬氨酸残基
9.1.2 转录终止阶段的调控
9.1.2.1 弱化子
研究表明色氨酸操纵子两种机制的调控。如果trp操纵子只受 trpR编码的阻遏物调控,那么在缺乏或存在色氨酸时,trpR 突变使trp操纵子表达的酶量应该是相同的。可是,在trpR缺
❖热激蛋白的表达调控主要发生在转录水平上。热激蛋白基 因的启动子被σ32而不是通常的σ70识别。σ32也不能识别σ70启 动子,因为这两种σ因子识别的启动子序列不一样
❖HSP的诱导合成是由于细胞内的σ32合成发 生在翻译水平。 ▪另一方面,在热激条件下σ32的稳定性也增加了。
严谨反应的分子机制
(p)ppGpp与RNA聚合酶β亚基结合,改变了RNA聚合酶对 一系列启动子的亲和力,导致细胞基因表达的整体变化,使细 胞适应新的环境。这些变化包括rRNA和tRNA的合成被抑制, 一系列参与氨基酸合成与运转的基因被激活。
人们在对大肠杆菌relA突变体进行研究时认识到是(p) ppGpp的积累引发了严紧反应。relA突变体即使在氨基酸饥饿
Fur能够感应细胞 内铁的水平。当 细胞内有充足的 铁时,Fur关闭反 义bfr基因,细胞 产生细菌铁蛋白。 在低铁条件下, 反义bfr基因被转 录,产生反义 RNA,阻止细菌 铁蛋白的合成。
分子生物学 第十一章 原核基因表达的调控

被cAMP激活 结合位点~22bp I -70 ~ -50
II -50 ~ -40
结合位点序列保守 不同基因受cAMP激活的水平不同
3 CAP的结合对DNA构型的影响
DNA弯曲 弯曲点位于CAP结合位点二重对称的中心 弯曲使CAP能与启动子上的RNA pol 接触
Summary
CA
B A: RNA polymerase B: lac repressor C: CRP-cAMP
Summary of lac operon regulation
Glucose High High Low Low
cAMP Low Low High High
Lactose Absent Present Absent Present
• 加入CAP,转录
• lac UV-5突变, -10区 TATGTT → TATAAT 在无CAP时,转录
• DNA topI 突变,降低起始转录对CAP的依赖
cAMP-CAP复合物的结合,使位点II附近的富含GC 区域双螺旋结构稳定性降低,因而-10区的熔解温度降 低,促进开放型启动子复合物的形成
9 原核生物基因表达的调控
9.1 基因表达概述 9.2 操纵元控制理论 9.3 基因转录的时序调控 9.4 转录后加工的调控 9.5 翻译水平的调控
孙朱乃玉恩贤
9.1 基因表达概述
9.1.1 生物遗传信息
9.1.1.1 C值矛盾 C value paradox
Genome DNA
10%; 结构基因的编码序列
triplet codon 90%; 重复,间隔,调节序列…
基因选择性表达指令 重要的遗传信息
.9.1.1.2 遗传信息的两大类别
生物学原核生物基因表达的调控

第二节
原核生物基因表达的 转录水平调控
Regulation of Prokaryotic Gene Expression at Transcription Level
目录
一、转录调控是以特定的DNA序列和蛋 白质结构为基础
(一)特定的DNA序列是转录起始调控的结构基础
在基因内和基因外都有一些特定的DNA序列,与结 构基因表达调控相关、能够被基因调控蛋白特异性识别 和结合,这些特定的DNA序列称为顺式作用元件(cisacting elements),亦称为顺式调控元件。在原核生物 中主要是启动子、阻遏蛋白结合位点、正调控蛋白结合 位点、增强子等。
transcription
RNA 5'-AGGUCCACG········-3'
启动子及其与转录的关系 ···
目录
(二)阻遏蛋白结合操纵元件对转录起 始进行负调控
阻遏蛋白是一类在转录水平对基因表达产生负 调控作用的蛋白质。阻遏蛋白主要通过抑制开放启 动子复合物的形成而抑制基因的转录。阻遏蛋白与 DNA结合后,RNA聚合酶仍有可能与启动子结合, 但不能形成开放起始复合物,不能启动转录;这种 作用称为阻遏(repression),特定的信号分子与阻 遏蛋白结合,使阻遏蛋白失活,从DNA 上脱落下来, 称为去阻遏,或脱阻遏(derepression)。
usually binds to CAAT box
目录
二、特定蛋白质与DNA结合后控制 转录起始
(一)σ因子和启动子决定转录是否能够起始
-35
-10
+1
5'-TAGTGTATTGACATGATAGAAGCACTCTACTATATTCTCAATAGGTCCACG············-·3·'
分子第五章原核基因表达调控

CAP正调控 + + + + 转录
DNA
CAP P O Z Y A
CAP CAP CAP CAP 无葡萄糖,cAMP浓度高时 促进转录
CAP
有葡萄糖,cAMP浓度低时
2、影响因子
(5)cAMP与代谢物激活蛋白 ◇ cAMP的浓度受到葡萄糖代谢的调节。 ◇由Crp基因编码的代谢物激活蛋白(CAP)能与cAMP形成复合物。 ◇ cAMP—CAP复合物是激活lac的重要组成部分,这与阻遏体系无 关,细菌对它的需要是独立的。转录必须有cAMP—CAP复合物结合 在启动子区。
Abstract
◇基因表达调控主要表现在以下两个方面: 1、转录水平上的调控(transcriptional regulation)。 2、转录后水平上的调控(post-transcriptional regulation): mRNA加工成熟水平上的调控、翻译水平上的调控 。
一、基本概念
1、组成蛋白和调节蛋白 ◇组成蛋白:细胞内有许多种蛋白质的数量几乎不受外界环境 的影响,这些蛋白质称为组成蛋白。 ◇调节蛋白:是一类特殊的蛋白质,它们可以影响一种或多种 基因的表达。调节蛋白包括:正调节蛋白和负调节蛋白。前者 是激活蛋白,后者是阻遏蛋白。
激活蛋白
启动子 操纵子
负调控
阻遏蛋白
启动子 操纵子
正调控和负调控
一、基本概念
5、操纵基因和操纵子 ◇操纵基因(operator):也叫操作子,是操纵子中的控制基因,在 操纵子上一般与启动子相邻,通常处于开放状态,使RNA聚合酶 通过并作用于启动子启动转录。 ◇操纵子(operon):由操纵基因以及相邻的若干结构基因所组成 的功能单位,其中结构基因转录受操纵基因控制。
5 第二章 原核生物基因表达调控

2.依赖于Rho因子的终止子
在体外实验中,发现尽管有该类终止子存在,但RNA聚合酶
只在终止子处暂停,转录并不终止。向该反应系统中加入ρ因子,
则可使转录在特定的位点终止。这种终止称为依赖ρ因子的转录
终止。
依赖于Rho因子的终止子不但柄部G•C含量较低,因而暂停
•
枯草杆菌在芽孢形成过程中采取更换σ因子的策略
2. 枯草杆菌噬菌体SP01的感染周期
一组基因的表达导致另一组基因随后表达,称为基因表达的
时序调控或级联调控,这是噬菌体感染周期中的普遍特征。 在枯草杆菌噬菌体SP01的感染周期中,其基因表达的时序调 控是由一系列的σ因子来实现的。 SP01的感染周期经历早中晚三个阶段: i. 刚刚感染时,早期基因被转录。
其一致序列为T80A95T45A60A50T96,又称为Pribnow框.
Pribnow盒是RNA聚合酶的牢固结合位点,又称结合位点,或称
为解链区。
该区域富含AT对,熔点较低, RNA聚合酶的结合使得这个AT丰
富区消耗较低的能量而解旋,此时RNA聚合酶与启动子的复合物便
由封闭复合物(closed-promoter complex) 转变为开放复合物(openpromoter complex) ,转录启动。 (2)-35区 在转录起始点上游-35bp处,有另一个保守序列,称 为-35序列。其一致序列为T82T84G78A65C54A45。该保守序列又称
• 转 录 起 始 主 要 由 DNA 分 子 上 的 启 动 子 (promoter)控制;终止由终止子控制. • DNA链上提供转录终止信号的序列称为终 止子(terminator)。是一个基因的末端或 是一个操纵子的末端的一段特定序列。
第六章 原核生物基因表达调控

图6-7 乳糖操纵子结构模式图
第二节 原核基因表达的调控
乳糖操纵子的上游有一个独立转录的基因lacI,其编码产物LacI 可以结合在乳糖操纵子的操纵基因(lacO)上,即转录控制区,阻抑 下游结构基因的表达。因此,乳糖操纵子是一个负调控系统(图68)。其中,LacI是具有负调控作用的反式作用因子,LacI作用的靶 DNA序列lacO是顺式作用元件。
trpB(UGA处翻译终止) -UGA -GAA-AUC- UGA-UGG-AA A UG-G AAtrpA(AUG处翻译起始)
第二节 原核基因表达的调控
3.稀有密码子对翻译的影响 DNA复制时,引物酶催化一段RNA引物的合成,引物酶 由dnaG编码。rpsU-dnaG-rpoD组成一个转录单位,产生多 顺反子转录物。细胞内三个基因的终产物的浓度相差却很 大,rpsU产物浓度为4×104个/细胞,dnaG产物50个/细胞, rpoD产物2800个/细胞。菌体通过使用稀有密码子,使转 录为一条mRNA链的三个基因的表达产物量可以有很大差异。
第二节 原核基因表达的调控
ቤተ መጻሕፍቲ ባይዱ
图6-10 色氨酸操纵子的负调控
第二节 原核基因表达的调控
4.阿拉伯糖操纵子 阿拉伯糖与乳糖一样,可替代葡萄糖作为碳源物质被 菌体利用。大肠杆菌中,阿拉伯糖(Ara)代谢所需酶的 三个基因分别是:核酮糖激酶基因( araB)、L-Ara异构 酶 基 因 ( araA)、L- 核 酮 糖 - 5 - 磷 酸 差 向 异 构 酶 基 因 ( araD),组成一个基因簇,有共同的启动子 PBAD。与其 它操纵子不同的是,操纵序列位于 PBAD 上游,操纵序列左 端有另一方向转录的启动子 PC,负责调节基因araC的转录, 其产物AraC蛋白有两种活性形式,Pr 对 PBAD 的表达起阻遏 作用,Pi对PBAD的表达起激活作用(图6-11)。
最新7原核生物基因表达调控汇总

2020/8/13
6
指挥基因调控的信号
– 原核生物:营养状况、环境因素 – 真核生物:激素水平、发育阶段
2020/8/13
7
基因表达调控的时间性和空间性
2020/8/13
8
7.1.1 原核基因调控分类
原核生物的基因调控主要是转录调控,包 括负转录调控和正转录调控。
• 正调控(positive control): 在操纵子中,结构基因本 来不表达,可当调节蛋白(无辅基诱导蛋白)出现时,使 该结构基因进行表达。这样的调控叫正调控。
• 调节蛋白(regulatory protein):调节基因的表达产物, 叫调节蛋白。包括正调节蛋白,又叫激活蛋白 (activator);负调节蛋白,又叫阻遏蛋白(repressor)。
• 由于调节蛋白(RNA)能够自由地与其相应的结合位点 结 合 , 故 又 称 为 反 式 作 用 因 子 ( trans-acting element)。
7原核生物基因表达调控
7.1 原核基因表达调控总论
• 基因表达(gene expression) :指基因经过转 录、翻译,产生具有特异生物学功能的蛋白质分 子或RNA产物的过程。
• 对 这 个 过 程 的 调 节 称 为 基 因 表 达 调 控 ( gene regulation 或 gene control)
2020/8/13
3
调控型基因(regulated genes)
• 又称奢侈基因(luxury genes) 在不同组织 细胞中选择表达的基因。根据细胞生长、 发育的需要或环境因素的改变,其活性受 到调控。
2020/8/13
4
2020/8/13
5
基因表达调控主要表现在以下两个方面: • 转 录 水 平 上 的 调 控 (transcriptional
第11章 原核生物基因表达的调控

Ø 葡萄糖代谢导致cAMP浓度下降; Ø cAMP可以活化乳糖操纵子的激活蛋白:
CRP: cAMP receptor protein(cAMP受体蛋白) CAP: catabolite gene activator protein
(代谢降解物活化蛋白)
Ø cAMP-CRP/CAP
乳糖操纵子的正调控
Ø 每个阻遏蛋白四聚体与两个 operator 结合; Ø 阻遏蛋白与Operator结合导 致DNA弯折,干扰mRNA的 合成。
p.286 图11-7
乳糖操纵子的正调控
当细菌在含有葡萄糖和乳 糖的培养基中生长时,通常 总是优先利用葡萄糖,而不 利用乳糖;只有当葡萄糖耗 尽后,细菌经过一段停滞期, 才能在乳糖的诱导下,合成 β-半乳糖苷酶等分解利用 乳糖的酶类,细菌才能利用 乳糖。
ttrrppRR
OOPPtrptrEpE trptDrpDtrpCtrpCtrpBtrpBtrpAtrpA
ttrrppRR
OOPPtrptrEpE trptDrpDtrpCtrpCtrpBtrpBtrpAtrpA
色氨酸操纵子的衰减作用
trpR
OP trpL trpE trpD trpC trpB trpA
5’
(1) 新合成的正链 RNA可以翻译A蛋白;
3’ (-) A
5’(+)
5’
但是很快形成二级结构,阻止A蛋白 的继续合成;
所以 A蛋白与C蛋白的量为1:180
Ø Rep的合成依赖于C蛋白的表达, 证据:C基因的codon6发生无义突 变:核糖体停留在该处,导致rep基 因RBS附近的二级结构无法打开, 则rep基因无法表达。
AraC既是阻遏蛋白, 又是激活蛋白;
原核生物基因表达调控

20
同位素示踪实验
把大肠杆菌细胞放在加有放射性35S标记的氨基酸,但没 有半乳糖诱导物的培养基中繁殖几代然后再将这些带有 放射活性的细菌转移到不含35S、无放射性的培养基中 随着培养基中诱导物的加入, β-半乳糖苷酶便开始合成。 分离β-半乳糖苷酶, 发现这种酶无35S标记说明酶的合 成不是由前体转化而来的, 而是加入诱导物后新合成的。
• Jacob和Monod认为诱导酶(他们当时称为适应酶)
现象是个基因调控问题, 可以用实验方法进行研究, 因此
选为突破口, 终于通过大量实验及分析, 于1961年建立
了该操纵子的控制模型。
-
21
酶的诱导
-
22
• 酶的诱导现象是生物进化过程中出现的一种合理、 经济地利用有限资源的本能。
• 酶诱导已证明是低等生物的普遍现象。
倒位片段
鼠伤寒沙门菌鞭毛素基- 因的调节
H1鞭毛素
10
鼠伤寒沙门氏菌(S.typhimrium)的相转变(phase variation)
-
11
2.σ 因子对原核生物转录起始的调控
σ因子:原核生物RNA聚合酶的一个亚基,是转录起 始所必需的因子,主要影响RNA聚合酶对转录起始 位点的正确识别,这种σ因子称σ70,此外还有分子量 不同,功能不同的其他σ因子 。
PO
操纵子可视为原核生物的转录单位,它可以逐个
地从原核生物基因组中分离出来,对其结构功
能加以研究。
-
15
3.乳糖操纵子
1) 乳糖操纵子的结构
启动子 操纵基因
调节蛋白
(阻遏蛋白)
-
结构基因
16
3个编码的结构基因
• Z编码β-半乳糖苷酶: 将乳糖水解成葡萄糖和半乳糖,还能 将乳糖转变为异构乳糖
分子生物学第七章原核生物基因表达调控

(三)、阻遏物 lac I 基因产物及功能
Lac 操纵子阻遏物 mRNA 是由弱启动子控制下组 成型合成的,该阻遏蛋白具有4个相同的亚基,每个亚 基均含347个氨基酸残基。
lacI 基因为组成型,通过启动子的上升突变体可获 得较多的阻遏蛋白;
阻遏物 2022/10/18
β-半乳糖苷酶 透过酶 转乙酰3酶2
2022/10/18
16
调节机理:
细胞中某一氨基酸或嘧啶的浓度发生改变
氨酰 – tRNA的浓度变化
核糖体在转录产物RNA上的结合位置不 同,使得RNA形成特定的二级结构 由RNA的二级结构判断基因能否继续转录
2022/10/18
17
3、降解物对基因活性的调节P252
葡萄糖效应或降解物抑制作用:细菌培养基中在 葡萄糖存在的情况下,即使加入乳糖、半乳糖等 诱导物,与其对应的操纵子也不会启动,这种现 象称为葡萄糖效应或降解物抑制作用。
这是通过阻止乳糖操纵子表达来完成的,这种 效应称为降解物抑制(catabolite repression)。
2022/10/18
35
(五)、cAMP与代谢物激活蛋白
葡萄糖
葡萄糖-6-磷酸
甘油 某些代谢产物抑制活性
腺苷酸环化酶
ATP
cAMP
编码
cAMP-CAP
Crp基因
代谢物激活蛋白 CAP
葡萄糖对其它糖的代谢抑制,是通过对 cAMP的抑制完成的。
2022/10/18
22
一、酶的诱导 ——
lac 体系受调控的证据
两种含硫的乳糖类似物:
异丙基巯基半乳糖苷
(IPTG)
巯甲基半乳糖苷(TMG)
E. coli 在不含乳糖的培养基生 长时,β-半乳糖苷酶含量极低;
原核生物基因表达调控概述

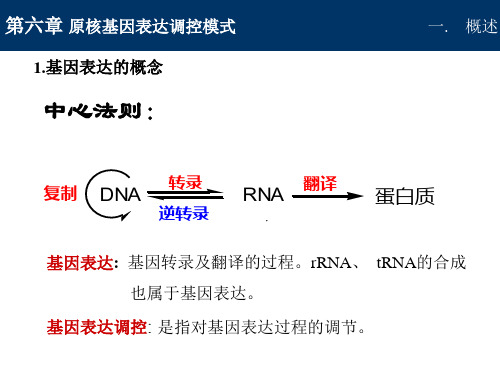
原核⽣物基因表达调控概述原核⽣物基因表达调控概述基因表达调控是⽣物体内基因表达调节控制机制,使细胞中基因表达的过程在时间,空间上处于有序状态,并对环境条件的变化做出适当的反应复杂过程。
1.基因表达调控意义在⽣命活动中并不是所有的基因都同时表达,代谢过程中所需各种酶和蛋⽩质基因以及构成细胞化学成分的各种编码基因,正常情况下是经常表达的,⽽与⽣物发育过程有关的基因则需在特定的时空才表达,还有许多基因被暂时的或永久的关闭⽽不来表达。
2.原核基因表达调控特点原核⽣物基因表达调控存在于转录和翻译的起始、延伸和终⽌的每⼀步骤中。
这种调控多以操纵⼦为单位进⾏,将功能相关的基因组织在⼀起,同时开启或关闭基因表达即经济⼜有效,保证其⽣命活动的需要。
调控主要发⽣在转录⽔平,有正、负调控两种机制在转录⽔平上对基因表达的调控决定于DNA的结构,RNA 聚合酶的功能、蛋⽩质因⼦及其他⼩分⼦配基的相互作⽤。
细菌的转录和翻译过程⼏乎在同⼀时间内相互偶联。
细胞要控制各种蛋⽩质在不同时期的表达⽔平,有两条途径:(1)细胞控制从其DNA模板上转录其特异的mRNA的速度,这是⼀条经济的途径,可减少从mRNA合成蛋⽩质的⼩分⼦物质消耗,这是⽣物长期进化过程中⾃然选择的结果,这种控制称为转录⽔平调控。
(2)在mRNA合成后,控制从mRNA翻译肽链速度,包括⼀些与翻译有关的酶及其复合体分⼦缔合的装配速度等过程。
这种蛋⽩质合成及其基因表达的控制称为翻译⽔平的调控。
⼆.原核⽣物表达调控的概念(1)细菌细胞对营养的适应细菌必须能够⼴泛适应变化的环境条件。
这些条件包括营养、⽔分、溶液浓度、温度,pH等。
⽽这些条件须通过细胞内的各种⽣化反应途径,为细胞⽣长的繁荣提供能量和构建细胞组分所需的⼩分⼦化合物。
(2)顺式作⽤元件和反式作⽤元件基因活性的调节主要通过反式作⽤因⼦与顺式作⽤元件的相互作⽤⽽实现。
反式作⽤因⼦的编码基因与其识别或结合的靶核苷酸序列在同⼀个DNA分⼦上。