《细胞生物学》核糖体与核酶自习报告
6核糖体与核酶共6页

1. 核糖体(riboso me)核糖体是细胞内一种核糖核蛋白颗粒(ribonucleoprotein particle), 其惟一功能是按照mRNA的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
按核糖体存在的部位可分为三种类型:细胞质核糖体、线粒体核糖体、叶绿体核糖体。
按存在的生物类型可分为两种类型:真核生物核糖体和原核生物核糖体。
原核细胞的核糖体较小, 沉降系数为70S,相对分子质量为2.5x103 kDa,由50S和30S两个亚基组成; 而真核细胞的核糖体体积较大, 沉降系数是80S,相对分子质量为3.9~4.5x103 kDa, 由60S和40S两个亚基组成。
在真核细胞中, 核糖体进行蛋白质合成时,既可以游离在细胞质中, 称为游离核糖体, 也可以附着在内质网的表面, 称为膜旁核糖体或附着核糖体。
真核细胞含有较多的核糖体, 每个细胞平均有106~107个, 而原核细胞中核糖体较少每个细胞平均只有15×102~18×103个。
典型的原核生物大肠杆菌核糖体是由50S大亚基和30S小亚基组成的。
在完整的核糖体中,rRNA约占2/3, 蛋白质约为1/3。
50S大亚基含有34种不同的蛋白质和两种RNA分子,相对分子质量大的rRNA的沉降系数为23S,相对分子质量小的rRNA为5S。
30S小亚基含有21种蛋白质和一个16S的rRNA分子。
真核细胞核糖体的沉降系数为80S,大亚基为60S,小亚基为40S。
在大亚基中,有大约49种蛋白质,另外有三种rRNA∶28S rRNA、5S rRNA 和5.8S rRNA。
小亚基含有大约33种蛋白质,一种18S的rRNA。
2. 基因扩增(gene a mp li fica tion)细胞内选择性复制DNA, 产生大量的拷贝。
如两栖类卵母细胞在发育的早期,rRNA基因的数量扩增到1000多倍。
基因扩增是通过形成几千个核进行的,每个核里含有几百拷贝的编码28S、18S和5.8S的rRNA基因,最后卵母细胞中的这些rRNA基因的拷贝数几乎达到50万个,而在相同生物的其它类型细胞中,这些rRNA基因的拷贝数只有几百个。
细胞生物学实验报告[1]
![细胞生物学实验报告[1]](https://img.taocdn.com/s3/m/af2e19e7763231126fdb11ca.png)
细胞生物学实验报告细胞生物学实验报告细胞生物学实验报告细胞生物学实验报告动物细胞融合【实验目的】⒈了解动物细胞融合的常用方法。
⒉学习化学融合的基本操作过程。
⒊观察动物细胞融合过程中细胞的行为和变化。
【实验原理】细胞融合是指在自发或者诱导条件下,两个或两个以上细胞合并为双核或者多核细胞的过程。
目前人们已经发现有很多方法可以诱导细胞融合,包括病毒诱导融合、化学诱导融合和电激诱导融合。
⒈病毒诱导融合仙台病毒、牛痘病毒、新城鸡瘟病毒和疱疹病毒等可以介导细胞的融合。
这类病毒的被膜中含有融合蛋白,可介导病毒和宿主细胞融合,同时也可介导细胞与细胞的融合。
用紫外线灭活后,这些病毒即可诱导细胞发生融合。
⒉化学诱导融合很多化学试剂能够诱导细胞融合,如聚乙二醇(PEG)、二甲基亚砜、山梨醇、甘油、溶血性卵磷脂、磷脂酰丝氨酸等。
这些物质能够改变细胞膜脂质分子的排列,在去除这些物质之后,细胞膜趋向于恢复原有的有序结构。
在恢复过程中相接触的细胞由于接口处脂质双分子层的相互亲和与表面张力,细胞膜融合,胞质流通,发生融合。
化学诱导方法,操作方便,诱导融合的概率比较高,效果稳定,适用于动、植物细胞,但对细胞具有一定的毒性。
PEG是被广泛使用的化学融合剂。
⒊电激诱导融合包括电诱导、激光诱导等。
其中,电诱导是先使细胞在电场中极化成为偶极子,沿电力线排布成串,再利用高强度、短时程的电脉冲击破细胞膜,细胞膜的脂质分子发生重排,由于表面张力的作用,两细胞发生融合。
电诱导方法具有融合过程易控制、融合概率高、无毒性、作用机制明确、可重复性高等优点。
【实验用品】⒈材料鸡血红细胞。
⒉试剂50%PEG、Hank’s溶液(pH7.4)、Alsever’s细胞保存液、0.85%氯化钠溶液。
⒊器材正置显微镜、离心机、离心管、载玻片、盖玻片、滴管、注射器等。
【实验步骤】⒈鸡血红细胞的制备和保存用无菌注射器先吸入Alsevers溶液1ml,再从鸡翼下静脉取血0.5~1ml,取出后放入刻度离心管中,再加入2~3mlAlsevers溶液,使总量为4~5ml,混匀并封口,置于4℃冰箱内。
细胞生物总结报告范文(3篇)

第1篇一、引言细胞生物学作为生物学的一个重要分支,研究细胞的形态、结构、功能及其生命活动规律。
随着科学技术的不断发展,细胞生物学在生命科学领域取得了举世瞩目的成就。
本报告旨在总结细胞生物学的研究成果,分析其发展趋势,为我国细胞生物学研究提供参考。
二、细胞生物学研究进展1. 细胞膜与信号转导细胞膜是细胞与外界环境进行物质交换和信息传递的重要界面。
近年来,研究人员在细胞膜的结构、功能及其调控机制方面取得了显著成果。
(1)细胞膜的结构:研究者通过冷冻电镜技术揭示了细胞膜的动态结构,发现细胞膜具有流动性,由磷脂双分子层和蛋白质组成。
(2)信号转导:细胞膜上的受体蛋白在接收外界信号后,通过一系列信号转导途径,将信号传递至细胞内部,调控细胞功能。
研究者成功解析了信号转导途径的关键分子,如G蛋白、磷酸化酶等。
2. 细胞骨架与细胞运动细胞骨架是维持细胞形态、参与细胞运动和细胞分裂的重要结构。
近年来,细胞骨架的研究取得了以下进展:(1)细胞骨架的组成:研究者通过荧光标记技术,揭示了细胞骨架的动态结构,包括微管、微丝和中间纤维。
(2)细胞骨架的调控:细胞骨架的组装、解聚和重构受到多种分子的调控,如肌动蛋白结合蛋白、微管相关蛋白等。
3. 细胞周期与细胞分裂细胞周期是细胞从出生到死亡的过程,细胞分裂是生物体生长发育的基础。
近年来,细胞周期与细胞分裂的研究取得了以下进展:(1)细胞周期调控:研究者通过研究细胞周期蛋白、周期素依赖性激酶等分子,揭示了细胞周期调控的机制。
(2)细胞分裂:研究者通过研究纺锤体、染色体分离等分子,揭示了细胞分裂的机制。
4. 遗传与基因表达遗传与基因表达是细胞生物学研究的重要内容。
近年来,以下研究成果具有重要意义:(1)基因编辑技术:CRISPR/Cas9基因编辑技术为研究基因功能提供了有力工具。
(2)转录组学:研究者通过转录组学技术,揭示了基因表达调控的复杂机制。
三、细胞生物学发展趋势1. 细胞器结构与功能研究细胞器是细胞内具有特定功能的亚细胞结构,如线粒体、内质网等。
细胞生物学实验报告_2

细胞生物学实验报告细胞凝集反应【实验目的】⒈了解细胞膜的表面结构。
⒉掌握凝集素促使细胞凝集的原理。
⒊学习研究细胞凝集反应的方法。
【实验原理】⒈细胞膜是双层脂镶嵌蛋白质结构, 脂和蛋白质又能和糖分子结合为细胞表面的分枝状糖外被。
目前认为: 细胞间的联系, 细胞的生长和分化, 免疫反应和肿瘤发生都和细胞表面的分枝状糖外被有关。
⒉凝集素(lectin)是一类含糖(少数例外)并能与糖专一结合的蛋白质, 它具有凝集细胞和刺激细胞分裂的作用。
凝集素使细胞凝集是由于它与细胞外被中的糖分子连接, 在细胞间形成“桥”的结果, 加入与凝集素互补的糖可以抑制细胞的凝集。
【实验用品】⒈材料土豆块茎、家兔。
⒉试剂⑴PBS缓冲液称取NaCl 7.2g、Na2HPO4 1.48g、KH2PO4 0.43g, 加蒸馏水, 定容至1L, 调pH至7.2。
⑵1%肝素溶液0.1g肝素溶解于10mL0.9%氯化钠溶液中。
⒊器材5mL注射器、酒精棉球、双凹片、普通光学显微镜、托盘天平、滴管、离心管。
【实验程序】⒈以无菌方法抽取兔耳缘静脉血液(5mL注射器先吸取3mL1%肝素溶液)1mL, 用0.9%氯化钠溶液洗5次, 每次3000r/min离心5min, 最后按沉淀的红细胞体积用0.9%氯化钠溶液配成1%红细胞悬液。
⒉称取去皮土豆块茎2g, 切成薄片, 加10mLPBS缓冲溶液, 浸泡2h, 浸出的粗提取液中含有可溶性土豆凝集素。
⒊用滴管吸取土豆凝集素和PBS缓冲液各一滴, 置于双凹片的左右两孔内, 再分别滴入一滴1%红细胞悬液, 振荡10min。
4.待红细胞液发生明显凝集反应后, 将双凹片置于显微镜下观察。
【实验结果】⒈结果细胞凝集反应实验现象⒉图像加土豆凝集素加PBS缓冲液【分析与讨论】⒈结果分析实验过程中可以观察到: 在双凹片上, 加有土豆凝集素的一侧, 震荡过程中, 红细胞液先变浑浊, 随后又变澄清, 同时发生凝集反应, 且凝集后的块状物聚集在液滴中央;而加有PBS缓冲液的一侧, 震荡过程中, 红细胞不发生明显变化, 静置后, 后细胞逐渐集中于液滴中间部位, 而四周表现为澄清。
细胞生物学实验报告(3篇)

细胞生物学实验报告(3篇)细胞生物学实验报告(精选3篇)细胞生物学实验报告篇1一、实验目的:1、掌握显示细胞中过氧化物酶反应的原理和方法。
2.了解细胞凋亡的生物学意义3、掌握凋亡细胞的形态学检测方法二、实验原理:1、细胞内的过氧化物酶能把许多胺类氧化为有色化合物,用联苯胺处理标本,细胞内的过氧化物酶能把联苯胺氧化为蓝色的联苯胺蓝,进而变为棕色产物,因而可以根据颜色反应来判定过氧化物酶的有无或多少。
中间产物蓝色联苯胺是不稳定的,无需酶的参加即可氧化为棕色化合物。
2、细胞凋亡是指细胞在生理或病理条件下,遵循自身的程序,自己结束其生命的过程。
它是一个主动的、高度有序的,基因控制的,一系列酶参与的过程。
3、凋亡细胞形态学特征是:体积变小,细胞质浓缩;细胞核发生染色质凝聚和聚集于核膜周围(边缘化);细胞膜有小泡状形成;晚期细胞膜内陷形成大小不同的凋亡小体;根据细胞凋亡形态学特征进行显微观察是检测细胞凋亡的一种直观、可靠的方法。
三、实验步骤:细胞中过氧化物酶的显示1、在载片上滴一滴PBS缓冲液;2、取骨髓细胞:用断颈法处死小鼠,立即剪取后肢,去除肌肉,剥出后肢股骨,剪开股骨一端,用牙签尖的一端插入剪开的小孔中,抠取少许骨髓细胞置滴有PBS的载片上;3、涂片:用另一玻片将骨髓细胞沿一个方向涂布推开,室温晾干;4、媒染:在涂片上滴0.5%硫酸铜液,以盖满涂片为宜,处理30秒-1分钟。
5、倾去硫酸铜液,直接滴入联苯胺混合液反应6分钟(以盖满涂片为宜)6、清水冲洗,番红复染2min。
7、镜检:清水冲洗,室温晾干,先低倍镜下观察,后换高倍镜下观察(油镜100_)细胞凋亡的形态学检测与观察吉姆萨染色:1、取细胞爬片置于小培养皿中(有细胞面朝上)2、生理盐水轻轻漂洗细胞3、95%乙醇固定5min4、PBS缓冲液洗2次5、吉姆萨染色液染色5min6、蒸馏水轻轻洗去染液7、普通光学显微镜下观察。
吖啶橙染色:1、取细胞爬片置于小培养皿中(有细胞面朝上)2、生理盐水轻轻漂洗细胞3、甲醇:冰醋酸(3:1)固定5min4、PBS缓冲液洗2次每次1min5、0.01%吖啶橙染色液在避光环境下染色5min6、蒸馏水轻轻洗去染液6、选用蓝光激发滤片在荧光显微镜下观察。
细胞生物学(翟中和版)——第9章 核糖体和核酶

?
19
20
¡与mRNA的结合位点 ¡与新掺入的氨酰-tRNA的结合位点——氨酰基位点,又称A位
点 ¡与延伸中的肽酰-tRNA的结合位点——肽酰基位点,又称P位
点 ¡肽酰转移后与即将释放的tRNA的结合位点——E位点(exit site) ¡与肽酰tRNA从A位点转移到P位点有关的转移酶
(即延伸因子EF-G)的结合位点 ¡肽酰转移酶的催化位点 ¡与蛋白质合成有关的其它起始因子、延伸因子和
细胞质 80S
60S(大亚基)
28S
(真核生物)
40S(小亚基) 18S,5.8S,5S
细胞质 70S
50S(大亚基)
23S
(原核生物)
30S(小亚基)
16S,5S
线粒体 55-60S 45S(大亚基)
16S
(哺乳动物)
35S(小亚基)
12S
线粒体 75S
53S(大亚基)
21S
(酵母)
35S(小亚基)
¡多聚核糖体的生物学意义细胞内 Nhomakorabea种多肽的合成,不论其分子量的大小
或是mRNA的长短如何,单位时间内所合成的
多肽分子数目都大体相等。
以多聚核糖体的形式进行多肽合成,对mRNA
的利用及对其浓度的调控更为经济和有效。
26
27
RNA在生命起源 中的地位及其演 化过程
28
生命是自我复制的体系
三种生物大分子,只有RNA既具有信息载体 功能又具有酶的催化功能。因此,推测RNA 可能是生命起源中最早的生物大分子。
CHAPTER 9
核糖体
Ribosome
1
OUTLINE
• Ribosome structure
细胞生物学(第五版)-第10章-核糖体精选全文完整版

二、蛋白质的合成
又称蛋白质的翻译,是细胞中最复杂、最精确的生 命活动之一。蛋白质合成需要各种携带氨基酸的 tRNA、核糖体、mRNA、多种蛋白质因子、阳离子 及GTP等的参与
蛋白质合成分为三步:
起始(Initiation)包括核糖体与mRNA 结合,形 成起始复合物,其中含有第一个氨酰-tRNA。
仅发现在哺乳动物成熟的红细胞 等极个别高度分化的细胞内没有 核糖体,线粒体和叶绿体中也含 有核糖体。 核糖体是细胞最基本的不可缺少 的结构。
核糖体是一种不规则颗粒状的结构,其主 要成分是RNA和蛋白质,直径约25 nm 核糖体蛋白分子主要分布在核糖体表面, 核糖体RNA(rRNA)位于内部,二者靠共价 键结合在一起。
甲基转移酶催化形成的。
30S小亚基与mRNA的结合需要 起始因子(initiation factor,IF)的 帮助。 这些起始因子仅位于30 S亚基上。 一旦30 S亚基与50 S亚基结合形 成70S核糖体后便释放。 起始因子的主要作用:帮助形成 起始复合物。 原核细胞有3种起始因子: IF1、IF2和IF3。
主要包括4个步骤: 1、氨酰-tRNA进入核糖体A位点的选择 2、肽键的形成 3、转位(translocation) 4、脱氨酰-tRNA的释放。
1.氨酰-tRNA在核糖体A位点的入位
起始的tRNAiMet占据P位点, 核糖体接受第2个氨酰-tRNA进 入A位点,这就是肽链延伸的 第一步。 为了有效地结合A位点,第二 个氨酰-tRNA必须与有GTP的 延伸因子(elongation factor, EF)EF-Tu结合形成复合物氨酰 -tRNA·EF-Tu·GTP。
三、核糖体蛋白质与rRNA的功能
核糖体上具有一系列与蛋白质合成有关的结合位 点与催化位点
06核糖体与核酶解析

第六章.核糖体与核酶核糖体(r i b o s o me),是细胞内一种核糖核蛋白颗粒(r i b o n u c l e o p r o t e i n p a r t i c l e),其惟一功能是按照mR N A的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
核糖体最早是Al b e r t C l a u d e于20世纪30年代后期发现的,其后又证明了其蛋白质合成功能。
随着分子生物学的发展,核糖体概念的涵意有了进一步的发展。
细胞内除了从事蛋白质合成的核糖体外,还有许多其它功能的核糖核蛋白体颗粒,通常是一些小分子的R N A同蛋白质组成的颗粒,它们参与R N A的加工、R N A的编辑、基因表达的调控等。
发现核糖体及核糖体功能鉴定的两个关键技术是什么?(答案)答:核糖体最早是Al b e r t C l a u d e于1930s后期用暗视野显微镜观察细胞的匀浆物时发现的,当时称为微体(M i c r o s o me s),直到1950s中期,Ge o r g e P a l a d e在电子显微镜下观察到这种颗粒的存在。
当时G e o r g e P a l a d e和他的同事研究了多种生物的细胞,发现细胞质中有类似的颗粒存在,尤其在进行蛋白质合成的细胞中特别多。
后来P h i l i p S i ek e v i t z用亚细胞组份分离技术分离了这种颗粒,并发现这些颗粒总是伴随内质网微粒体一起沉积。
化学分析揭示,这种微粒富含核苷酸,随之命名为r i b o so me,主要成分是核糖体R N A(r R N A),约占60%、蛋白质(r蛋白质)约占40%。
核糖体的蛋白质合成功能是通过放射性标记实验发现的。
将细胞与放射性标记的氨基酸短暂接触后进行匀浆,然后分级分离,发现在微粒体部分有大量新合成的放射性标记的蛋白质。
后将微粒体部分进一步分离,得到核糖体和膜微粒,这一实验结果表明核糖体与蛋白质合成有关。
细胞生物学核糖体的结构及功能

文档收集于互联网,已重新整理排版.word版本可编辑,有帮助欢迎下载支持.第十一章核糖体一、核糖体的结构及功能核糖体是体积较小的无膜包围的细胞器,在光镜下看不到。
1958年才把这种含有大量RNA的能合成蛋白质的关键装置定名为核糖核蛋白体ribosome,简称为核糖体。
(一)核糖体的一般性质1、存在与分布核糖体存在一切生物的细胞中,包括真核细胞和原核细胞。
这是有别于其它细胞器的特点。
在真核细胞中,有些核糖体是游离分布在细胞质基质中,也有许多是附着在rER膜及核膜外表。
此外,还有核糖体是分布在线粒体和叶绿体的基质中。
在原核细胞内,大量核糖体游离在细胞质中,也有的附着在质膜内侧面。
细菌的核糖体占总重量的25—30%2、形态和大小一般直径为25—30nm,由大、小两亚单位构成,通常是以大亚单位附在内质网膜或核膜外表。
当进行蛋白质合成时,小亚单位先接触mRNA才与大亚单位结合,而合成完毕后又自行解离分开。
另外,多个核糖体还可由mRNA串联成多聚核糖体polyribosome(=polysome),每个多聚核糖体往往由5-6个核糖体串成,但也有多至50个以上的(例如肌细胞中合成肌球蛋白的多聚核糖体是由60—80个串联而成)。
3.数量和分类细胞中的核糖体数量多少不一。
一般来说,增殖速度快的细胞中偏多,分泌蛋白质的分泌细胞中也较多。
例如分泌胆汁的肝细胞中为6X 106个,大肠杆菌中为1500—15000个。
在不同类型生物细胞之中,核糖体大小及组分都有一定差1文档来源为:从网络收集整理.word版本可编辑.文档收集于互联网,已重新整理排版.word版本可编辑,有帮助欢迎下载支持.异。
一般可分为两大类:80S型和705型。
大亚单位60S真核生物核糖体80S v小亚单位40S大亚单位50S原核生物核糖体70S一小亚单位30S(“S”是沉降系数的衡量单位。
大、小亚单位组成核糖体, 并非由其两者的S值直接相加,这是因为S值变化其实是与颗粒的体积及形状相关的。
细胞学核糖体和核酶

人细胞核糖体的合成与装配
21
7.3 核糖体的功能
◆核糖体的功能位点 ◆蛋白质合成的基本过程
22
核糖体的功能位点
23
核糖体的功能位点
大亚基: A位点:氨酰tRNA 位点(Aminoacyl-tRNA site)
P位点:肽酰tRNA 位点(Peptidyl-tRNA site)
E位点(Exit site):脱氨酰tRNA离开核糖体时的 临时停留点 小亚基: 与mRNA结合位点
29
第一个氨酰tRNA进入核糖体的P位点
原核生物: 携带甲酰甲硫氨酸的tRNA通过反密码子与
mRNA中的AUG的识别进入核糖体。起始tRNA与 GTP、IF2结合形成GTP-IF2-tRNAfmet复合物,复 合物与mRNA的AUG结合后,结合小亚基的IF3释 放。
30
完整复合物的装配
起始tRNA与AUG结合后,大亚基与GTPIF2-tRNAfmet复合物结合,形成核糖体-mRNA 起始复合物。
细胞有两种主要类型的核糖体:
◆原核细胞的核糖体: 沉降系数为70S,分子量为2.5×106,由50S和30S 两个亚基组成。
◆真核细胞(细胞质)的核糖体
沉降系数是80S,分子量为4.8×106,由60S和40S 两个亚基组成。
32
各种来源的核糖体亚基组成
来源 完整核糖体 核糖体亚基 核糖体RNA
细胞质 80S 60S(大亚基) 28S,5.8S,5S
(高等植物)
45S(小亚基) 18S
叶绿体 70S 50S(大亚基) 23S,5S
30S(小亚基) 16S 4
核糖体的化学组成
5
7.2 Ribosome Biogenesis(核糖体的生 物发生 )
细胞生物学中的核糖体功能研究

细胞生物学中的核糖体功能研究从生命诞生的那一刻起,细胞就成为了生命的基本单位。
而核糖体作为细胞内最为重要的基因功能执行器,扮演着维持生命的重要角色。
近年来,众多临床实践表明,核糖体在多种疾病的发生和进展中起到重要的调节作用,如何更深入地研究核糖体的生物学功能,已经成为细胞生物学研究的热点之一。
核糖体是细胞内的一种大分子复合物,由蛋白质和RNA两大部分组成。
在细胞内,核糖体起到重要的蛋白质合成和组装的作用,可谓是细胞的工厂。
具体而言,在核糖体的作用下,RNA将DNA的信息转录成为蛋白质,进而参与到细胞的各种功能过程中。
如此可知,核糖体是细胞内的一个非常重要的组成部分,其功能对于生命的维持至关重要。
在研究核糖体的功能过程中,一个重要的方向就是研究它是如何完成蛋白质合成的。
从最早的大小互补实验开始,到后来的X光晶体学结构解析,越来越多的研究显示,核糖体是一个非常复杂的大分子复合物,其蛋白质和RNA的互作关系十分复杂。
近年来,随着高通量技术的广泛应用,如同步辐射衍射技术,大大推动了对核糖体结构及其功能过程的研究进展。
除了研究核糖体执行蛋白质合成的过程,一些研究还涉及了核糖体的调节作用。
众所周知,在细胞内,生命的各种功能是由多种多样的分子共同实现的,核糖体的调节作用就是其中重要的一个部分。
研究表明,在细胞内,多种分子参与到核糖体的调节当中,如微RNA、长链非编码RNA等。
这些分子通过与核糖体的不同区域之间的相互作用,调节核糖体的活性,从而影响细胞内的蛋白质合成和组装过程。
这一方面的研究,为我们深入理解核糖体在生物学功能执行中的作用,提供了更深入和全面的认识。
不得不说,研究核糖体在生物学过程中的功能,既有其重要性,也面临诸多的挑战。
与传统的生物学研究不同,在研究核糖体的生物学功能中,需要综合运用到多种方法,如同步辐射衍射技术、核磁共振技术、分子生物学技术等。
且这些技术需要研究者具备一定的专业知识和技能,对研究者的要求也较高。
细胞生物学实验报告

细胞生物学实验报告引言细胞是生命的基本单位,它们组成了所有生物体,从微观的细菌到宏观的植物和动物。
了解细胞的结构和功能对于理解生命系统的运作机制至关重要。
本实验旨在通过观察和研究细胞的各个方面,探索细胞的奥秘。
材料与方法本实验使用了显微镜、荧光显微镜、细胞培养皿、培养基和染色剂等实验材料。
首先,我们在细胞培养皿中放入培养基,并加入细胞样本。
接着,我们使用显微镜观察细胞的结构,并通过染色剂标记特定的细胞结构。
最后,我们使用荧光显微镜观察细胞内的荧光分子。
结果与讨论1. 细胞的形态和结构通过显微镜观察,我们发现细胞具有不同的形态和结构。
植物细胞通常具有细胞壁和叶绿体,而动物细胞则没有细胞壁,但有细胞膜和细胞器如线粒体和高尔基体。
这些不同的结构为细胞的功能提供了支持。
2. 细胞的运动和分裂我们观察到细胞在培养皿中具有一定的运动能力。
细胞可以通过细胞膜的伸缩和细胞骨架的变形实现不同的移动方式,如原生动物的鞭毛运动和细胞质流动。
此外,我们还观察到细胞分裂的过程,这是细胞增殖和生长的基本方式。
3. 细胞的代谢活动通过观察荧光显微镜下的细胞样本,我们可以看到许多荧光分子在细胞内的活动。
这些分子参与了细胞的代谢过程,如蛋白质合成、物质转运和能量代谢。
荧光分子的存在使我们能够更直观地了解细胞的生理活动。
4. 细胞的信号传导在细胞培养条件下,我们还观察到细胞之间的相互作用和信号传导。
细胞可以通过细胞外信号分子的识别和细胞膜上的受体进行相互沟通。
这种信号传导机制对于细胞内外环境的调节和细胞功能的协调起着重要作用。
结论通过本实验,我们深入了解了细胞的形态、结构、运动、代谢和信号传导等方面。
细胞生物学是研究生命的基本单位的科学,其对于解答许多生物学问题具有重要意义。
今后的研究中,我们可以进一步探索细胞的分子机制和调控网络,以深化对细胞生物学的理解,并应用于生物医学和生物工程领域。
致谢感谢实验室的老师和同学们对本实验的支持和帮助,使我们能够顺利完成实验,并获得有价值的数据和观察结果。
细胞生物学实验报告_2
细胞生物学实验报告叶绿体的分离、纯化与荧光观察【实验目的】⒈通过植物细胞叶绿体的分离与纯化, 了解细胞器分离与纯化的原理和方法。
⒉熟悉荧光显微镜的使用方法, 观察叶绿体的自发荧光和间接荧光。
【实验原理】⒈叶绿体是植物细胞所特有的能量转换细胞器, 叶绿体是植物细胞所特有的能量转换细胞器, 光合作用就是在叶绿体中进行。
由于具有这一重要功能, 所以它一直是细胞生物学、遗传学和分子生物学的重要研究对象。
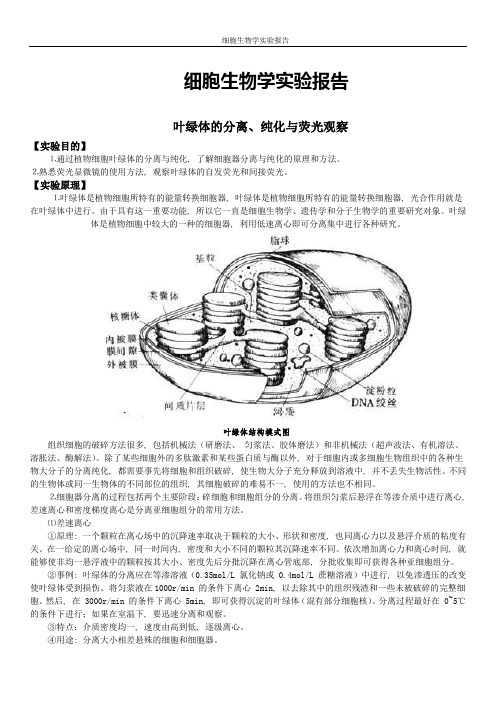
叶绿体是植物细胞中较大的一种的细胞器, 利用低速离心即可分离集中进行各种研究。
叶绿体结构模式图组织细胞的破碎方法很多, 包括机械法(研磨法、匀浆法、胶体磨法)和非机械法(超声波法、有机溶法、溶胀法、酶解法)。
除了某些细胞外的多肽激素和某些蛋白质与酶以外, 对于细胞内或多细胞生物组织中的各种生物大分子的分离纯化, 都需要事先将细胞和组织破碎, 使生物大分子充分释放到溶液中, 并不丢失生物活性。
不同的生物体或同一生物体的不同部位的组织, 其细胞破碎的难易不一, 使用的方法也不相同。
⒉细胞器分离的过程包括两个主要阶段:碎细胞和细胞组分的分离。
将组织匀浆后悬浮在等渗介质中进行离心, 差速离心和密度梯度离心是分离亚细胞组分的常用方法。
⑴差速离心①原理: 一个颗粒在离心场中的沉降速率取决于颗粒的大小、形状和密度, 也同离心力以及悬浮介质的粘度有关。
在一给定的离心场中, 同一时间内, 密度和大小不同的颗粒其沉降速率不同。
依次增加离心力和离心时间, 就能够使非均一悬浮液中的颗粒按其大小、密度先后分批沉降在离心管底部, 分批收集即可获得各种亚细胞组分。
②事例: 叶绿体的分离应在等渗溶液(0.35mol/L 氯化钠或 0.4mol/L 蔗糖溶液)中进行, 以免渗透压的改变使叶绿体受到损伤。
将匀浆液在1000r/min 的条件下离心 2min, 以去除其中的组织残渣和一些未被破碎的完整细胞。
然后, 在 3000r/min 的条件下离心 5min, 即可获得沉淀的叶绿体(混有部分细胞核)。
第06章 核糖体和核酶

第六章核糖体和核酶章节提要唐浩能生命科学大学院生命科学大类 13335155核糖体是细胞内一种核糖核蛋白颗粒,其唯一功能是按照mRNA的指令将氨基酸合成蛋白质多肽链。
所以又被称为蛋白质合成的机器。
核糖体由60%的核糖体RNA(rRNA)和40%的核糖体蛋白质组成。
本章内容包括:核糖体的形态结构、核糖体的生物发生、核糖体的功能——蛋白质的合成、反义RNA与核酶。
一、核糖体的形态结构核糖体由三种类型:细胞质核糖体、线粒体核糖体和叶绿体核糖体。
根据在不同生物体内的大小和组成的不同又可以分为真核生物核糖体和原核生物核糖体。
核糖体都是由两个大小不同的亚基组成。
核糖体的化学组成为核糖体RNA 和核糖体蛋白质。
原核生物核糖体沉降系数为70S,由50S大亚基(含33种不同的蛋白质以及23S和5S两种rRNA)和30S小亚基(含21种蛋白质以及16S rRNA)组成;真核生物核糖体沉降系数为80S,由60S大亚基(含大约49种蛋白质以及28S、5S和5.8S rRNA)和40S小亚基(含大约33种蛋白质以及18S rRNA)组成。
二、核糖体的生物发生核糖体的生物发生包括蛋白质和rRNA的合成、核糖体亚基的组装等。
因为细胞需要大量的rRNA,因此在进化过程中形成了一种机制:增加编码rRNA基因的拷贝数。
真核生物的18S、5.8S和28SrRNA基因首先转录成一个45S前rRNA,能够转录该前rRNA的DNA区域称为一个转录单位,意指他们有一个共同的转录起点和终点。
接着45SrRNA转变成32SrRNA,然后变成28S和18S rRNA。
真核生物合成前rRNA之后需要进行加工。
细菌等原核生物的rRNA基因的转录首先是形成16S-23S-5S前体RNA。
然后进行剪切分开。
核糖体的装配是自发进行的,但是其组装部位,在真核生物是在细胞核的核仁部位,在原核生物是在细胞质中。
三、核糖体的功能——蛋白质的合成在原核生物中有4种与RNA分子结合的位点,其中一个是与mRNA结合的位点,另外三个是tRNA结合的位点。
细胞生物学 章节提要 第六章 核糖体与核酶

细胞生物学章节提要第六章核糖体与核酶细胞生物学章节提要第六章核糖体与核酶第06章总结核糖体与核酶研究方法:足迹、放射性标记(核糖体发现)、原核核糖体重组实验和合成反义RNA干扰。
核糖体(ribsome)是细胞内一种核糖蛋白颗粒(ribonucleoproteinparticle),唯一功能是翻译mrna的指令,将氨基酸合成蛋白质多肽链。
核糖体包括线粒体核糖体、叶绿体核糖体、细胞质核糖体。
真核细胞和原核细胞的核糖体组成不同,由均有两个不同大小的亚基构成。
它有rrna和核糖体蛋白质(ribosomalprotein),原核细胞由50s和30s 两个亚基构成70s的核糖体,真核细胞由60s和40s两个亚基构成80s的核糖体。
核糖体生物发生包括蛋白质和核糖核酸的合成、核糖体亚基的组装等。
在进化过程中,通过增加染色体上rRNA基因的拷贝数和基因扩增,编码rRNA的基因数量增加。
但第一个含有rRNA基因的DNA是如何形成的尚不清楚。
真核18S、5.8S和28s rRNA基因形成一个转录单元,这是转录成45s的前提。
原核生物的16S、23S和5S rRNA基因构成一个转录单元。
与成熟rRNA相比,真核生物的前rRNA含有大量甲基化核苷和大量假尿苷。
原核生物的rRNA基因也是多拷贝的(在真核生物中重复率相对较低),需要转录成前体,然后加工成rRNA。
此外,在真核生物中编码5srrna的基因位于不同的染色体上,而细菌5srrna基因与其他两个rRNA结合形成一个转录单元。
在组装部位,真核生物在核仁中,原核生物在细胞质中。
核糖体的唯一功能是蛋白质合成(proteinsynthetize)。
核糖体中与trna结合位点有a位点(asite)、p位点(psite)、e位点(esite)。
原核生物通过sd序列识别结合,真核生物通mrna5’端甲基化帽子结构识别。
蛋白质合成主要的过程为:链的起始、链的延伸、链的终止。
大约90%以上的真核生物的起始密码子是aug。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
核糖体与核酶引言:1.核糖体(ribosome)是细胞内的一种核糖蛋白颗粒,其唯一的功能是按照mRNA的指令将氨基酸合成蛋白质多肽链,所以核糖体是细胞内蛋白质合成的分子机器。
6.1 核糖体的形态结构1.核酶是具有催化活性的反义RNA6.1.1 核糖体的类型和化学组成6.1.1.1 核糖体的类型和大小1.核糖体有种类型:细胞质核糖体、线粒体核糖体、叶绿体核糖体2.核糖体分为:真核生物核糖体和原核生物核糖体3.核糖体由大小两个不同的亚基组成,在不进行蛋白质合成时是分开的,各自游离在细胞质中,在进行蛋白质合成时结合在一起4.在真核细胞中,核糖体在进行蛋白质合成时:1.游离在细胞质中称游离核糖体2.附着在内质网的表面,称膜旁核糖体或附着核糖体。
6.1.1.2 核糖体的化学组成1.核糖体的大小两个亚基都是由核糖体RNA(rRNA)和核糖体蛋白质组成。
6.1.2核糖体的蛋白质与rRNA6.1.2.1 核糖体蛋白1. E.coli核糖体21个小亚基,为S1~S21,大亚基的核糖体蛋白命名为L1~L336.1.2.2 核糖体rRNA1.30S核糖体亚基的形态主要是由16S rRNA决定的6.1.3细菌核糖体的结构模型1.S4、S5、S8、S12等4个蛋白定位在核糖体的小亚基上,并且是背向大亚基。
2.小亚基中确定了与信使RNA(mRNA)和转移RNA(tRNA)结合位点3.催化肽键形成的位点位于大亚基,和GTP水解的功能区6.2核糖体的生物发生1.在细胞内,核糖体是自我装配的。
2.核糖体的生物发生包括蛋白质和rRNA的合成、核糖体亚基的组装等。
6.2.1 核糖体rRNA基因的转录与加工1.编码核糖体的基因分为两类:一类是编码蛋白质的基因,另一类是rRNA基因6.2.1.1 编码rRNA基因的过量扩增1.细胞为了满足大量需求的rRNA,在进化的过程中形成了一种机制:增加编码rRNA基因的拷贝数。
2.增加拷贝数有两种方法:1.在染色体上增加rRNA基因的拷贝数2.通过基因扩增6.2.1.2 真核生物18S、5.8S、28S rRNA和5S rRNA基因1.在真核生物的染色体中,18S、5.8S、28S rRNA和5S rRNA基因是串联在一起的,每个基因被间隔区隔开,5S rRNA基因位于不同的染色体上。
2.在间期核中,所有这五条染色体rRNA基因区域,转录时聚集在一起,形成一个核仁。
6.2.1.3 真核生物18S、5.8S、28S rRNA和5S rRNA基因转录与加工1.真核生物参加rRNA基因转录的酶是RNA聚合酶I。
6.2.1.4 真核生物前rRNA的修饰1.前rRNA有两个独特的特点:1.含有大量的甲基化的核苷2.具有很多假鸟苷2.甲基化的主要部位在核糖体第二位的羟基上,甲基化可能对加工起引导作用3.前rRNA的修饰可能有利于rRNA的正确折叠一集与别的分子正确的相互作用6.2.1.5 5SrRNA基因的转录与加工1.5S rRNA基因是由RNA聚合酶III转录的,原初转录的5`端与成熟的5S rRNA的5`端完全相同2.RNA聚合酶III通常是与位于转录部分内的启动子结合,而不是与转录其实位点上游的启动子结合。
6.2.1.6 原核生物rRNA基因及转录1.原核生物与真核生物rRNA基因在组织结构上的差异:1.在基因的重复次数上,真核生物的rRNA基因重复的次数相当高,而原核生物的rRNA基因的重复次数却很少。
2.在真核生物中编码5S rRNA的基因位于不同的染色体上,而细菌的5S rRNA基因与另外两种rRNA基因组成一个转录单位,它们在染色体上的排列顺序是16S-23S-5S。
2. 通过核酸酶III(RNaseIII)突变体的研究证明原核生物的3钟rRNA位于一个原初转录物中。
3. 在细菌中核酸酶III不是参与前rRNA加工的惟一的酶,还需要其他一些酶核酸酶参加,如果缺少其他一些酶,就不能得到正确大小的rRNA。
6.2.1 核糖体rRNA基因的转录与加工1.真核生物核糖体亚基的装配地点,原核生物核糖体亚基的装配则在细胞质中。
6.2.2.1 核糖体亚基的自我装配1.Nomura实验表明:核糖体是一种自组装的结构,即没有样板货亲体结构所组成的结构。
但是在组装的过程中,某些蛋白质必须首先结合到rRNA上,其他蛋白才能组装上去。
即组装有先后顺序。
2.小亚基在装配的起始,一些蛋白能独立地与16S rRNA结合,装配的中间物为其他蛋白的结合提供了架构。
中间体必须经过构型的变化才能与其他蛋白结合。
3.30S亚基上的蛋白分为三类:1.开始装配所必需的蛋白质,它们先和rRNA特定区段结合,形成核糖体亚基的核心。
2.结合后可与后一批蛋白提供结合位点3.非30S亚基形成所必需,但却是显示其活性所必须的。
6.2.2.2 原核生物核糖体重组实验1. 核糖体重组:指将不同源的核糖体大小亚基货不同亚基的rRNA或蛋白质重新组合,形成杂合的核糖体后研究其功能。
6.2.2.3 真核生物核糖体在核仁中装配1. 组成核糖体的18S、5.8S、28S rRNA在核仁中边转录边参与核糖体亚基的装配,5S rRNA却是在细胞核中转录后送到核仁中参与核糖体亚基的装配。
6.3核糖体的功能——蛋白质的合成1. 核糖体的功能是进行蛋白质合成,在核糖体上合成的只是蛋白质的一集结构。
2. 合成方向是5’到3’6.3.1 核糖体的功能位点1. 核糖体功能位点中有4种与RNA分子结合的位点,其中一个是与mRNA结合的位点,另3个是与tRNA结合的位点。
2. A位点:即氨酰基位点,是与新掺入的氨酰基tRNA结合的位点,又叫受位。
主要位于大亚基,是接受tRNA的部位。
3. P位点,即肽酰tRNA位点,又叫供位,主要位于大亚基,是肽基tRNA移交肽链后肽酰tRNA所占据的位置,即子延伸中与tRNA结合位点4. E位点:是脱氨酰tRNA离开P位点到完全从核糖体释放出来的一个中间停靠位点。
6.3.2 蛋白质合成的基本过程1.在核糖体上合成多肽链,分为3步:链的起始、链的延伸、链的终止。
6.3.2.1 蛋白质合成的起始1.蛋白质合成起始涉及mRNA、起始tRNA和核糖体小亚基之间相互用,最后装配成完整的核糖体。
2.起始过程分三步:1.30S亚基与mRNA的结合:mRNA不能与完整的何烨结合,但是能够同独立存在的30S核糖体小亚基结合。
核糖体小亚基与mRNA的结合还需要起始因子。
2.第一个氨酰-tRNA进入核糖体。
3.完整的起始复合物的装配。
6.3.2.2 多肽链的延伸1.蛋白质的延伸:1.氨酰tRNA进入核糖体的A位点2.肽键的形成3.转位4.脱氨酰tRNA释放在整个过程中需要GTP和延长因子的参与。
2.氨酰tRNA进入核糖体的A位点:由起始tRNA占据P位点,核糖体开始接受第二个氨酰-tRNA进入A位点,为延伸的第一步3.肽键的形成:由A位点的氨酰-tRNA上氨基酸的氨基与P位点氨酰t-RNA上氨基酸的羧基间形成肽键4.转位5.脱氨酰tRNA释放6.在蛋白质合成的延伸反应,每一次循环至少水解两分子的GTP6.3.2.3 终止1.蛋白质合成的终止是指核糖体沿着mRNA移动,如果进入A位点的是终止密码子,由于没有与之匹配的反密码子,而终止蛋白质的合成。
2.终止密码子:UAA、UAG、UGA3.蛋白质合成的终止需要释放因子(RF)的存在。
6.3.3多聚核糖体1.在蛋白质合成过程中,同一条mRNA分子能够同多个核糖体结合,同时合成若干条蛋白质多肽链,结合在同一条mRNA上的核糖体就称为多聚核糖体。
2.多聚核糖体达达提高蛋白质合成的速率,也减轻了细胞核进行基因转录和加工的压力。
6.3.4蛋白质合成抑制剂6.3.4.1 抗生素1.抗生素是主要的蛋白质合成抑制剂2.链霉素主要是抑制起始tRNA和非起始tRNA与核糖体的结合,导致肽链合成的提前终止。
6.3.4.2 嘌呤霉素1.嘌呤霉素是一种蛋白质合成抑制剂,它具有与tRNA分子末端类似的结构,能够同氨基酸结合,代替氨酰tRNA同核糖体A位点结合,6.3.5 细胞中蛋白质的寿命与讲解6.3.5.1 蛋白质的寿命信号1.细胞内新生的蛋白质通常有3种特别的信号:加工信号、分选定位信号、寿命信号。
每一种蛋白质都有寿命特征,称为半寿期。
2.蛋白质N端与半寿期的关系,称为N端规则6.3.5.2 蛋白酶体对蛋白质的降解1.执行降解蛋白质的任务的是多蛋白的复合体,称为蛋白酶体。
2.蛋白酶体既存在于细胞核中,又存在于胞质溶胶中。
3.蛋白酶体主要讲解两种类型的蛋白质:一类是错误折叠的蛋白质,另一类是需要进行数量调节的蛋白质。
4.蛋白酶体对蛋白质的降解通过遍在蛋白质介导,所以又称为遍在蛋质降解途径。
5.蛋白酶体对蛋白质的降解作用分为两个过程:1.对被降解的蛋白质进行标记由遍在蛋白质完成。
2.蛋白酶解作用,由蛋白酶体催化。
6.4反义RNA与核酶6.4.1 小分子RNA与反义RNA6.4.1.1 小分子RNA的类型1.主要有两种小分子RNA:1.snRNA,存在于细胞核中2.scRNA,存在于细胞质中。
2.由于snRNA富含尿嘧啶核苷,分类时把它分为U1、U26.4.1.2反义RNA与蛋白质合成的抑制1.反义RNA是指与mRNA互补的RNA分子,也包括与其他RNA互补的RNA分子。
2.由于核糖体不能翻译双链的RNA,所以反义RNA与mRNA互补结合即抑制了该mRNA的翻译。
3.细胞中反义RNA的来源有两种途径:1.反向转录的产物 2.来源是不同的基因产物4.原核细胞中发现的反义RNA,根据作用分为3类:1. I类反义RNA:这类RNA直接作用于其靶mRNA的SD序列和(或)编码区,引起翻译的直接抑制(1A类);或与靶mRNA结合后引起该链RNA分子对RNaseIII的敏感性增加,使其降解(1B类)2. II类反义RNA:与mRNA上的SD序列的上游非编码区结合,从而抑制靶mRNA的翻译功能3.III类反义RNA:可直接抑制靶mRNA的转录6.4.1.3 反应snRNA在前rRNA加工中的作用1.细胞核中的snNA通常与特定的蛋白质包裹在一起形成颗粒结构,称小核糖核蛋白2.SnRNA与前rRNA通过互补形成的RNA-RNA双链部分可作为前rRNA进行加工修饰的标志3.根据反义snRNA的功能和序列的相似性,分为:1.C/D框snRNA,这类RNA的5’有C/D框结构,它们的功能是决定何种核苷的核糖甲基化。
2.H/HAC框snRNA,这类反义snRNA的3’有HAC序列,它们的作用是决定哪些尿苷被转变成假尿苷。
6.4.2 核酶1.核酶:化学本质是核糖核酸,却具有酶的催化功能6.4.2.1 核酶大发现1.在大多数真核生物的rRNA加工中,从前rRNA中切除的序列都是间隔区而不是内含子,所谓间隔区就是位于编码基因间的序列而不是位于基因内的序列。