第七章 蛋白质分选与膜泡运输
蛋白质分选与膜泡运输

COPⅠ有被小泡的形成 Ⅰ
LOGO
① 一种胞质溶胶中的 小分子GTP结合蛋白, GTP结合蛋白 小分子GTP结合蛋白,即 ARF,释放 释放GDP, GTP结 ARF,释放GDP, 同GTP结 形成ARF GTP复合物 ARF复合物, 合,形成ARF-GTP复合物, 并整合在高尔基体膜中。 并整合在高尔基体膜中。 COPⅠ同ARF以及高 ② COPⅠ同ARF以及高 尔基体膜蛋白的细胞质 部分结合; 部分结合; ③在脂酰CoA的帮助下 在脂酰CoA的帮助下 CoA 形成COPⅠ被膜小泡, COPⅠ被膜小泡 形成COPⅠ被膜小泡, 一旦COPⅠ COPⅠ小泡形成就 一旦COPⅠ小泡形成就 立即从供体膜释放出来, 立即从供体膜释放出来, COPⅠ包被去聚合 包被去聚合, COPⅠ包被去聚合, 并 与膜脱离, 与膜脱离, 这一过程是 由与ARF结合的GTP ARF结合的GTP水解 由与ARF结合的GTP水解 所触发。 所触发。
COPII有被小泡的组装与运输 COPII有被小泡的组装与运输
过程:
LOGO
• ER腔中可溶性货物蛋白 可溶性货物蛋白通过与选择性跨膜蛋白腔面一端 可溶性货物蛋白 结合而被募集 • 选择性跨膜蛋白胞质面一端的信号序列以及ER膜上整合 蛋白v-SNARE被COPII蛋白识别,形成有货物蛋白和 货物蛋白和v货物蛋白和 SNARE的有被小泡。 • 小分子GTP结合蛋白(Sar1蛋白,对包被组装和去组装起 调控作用)参与下脱被并定位到高尔基体的cis面。 • 暴露的v-SNARE与高尔基体cis面膜上的同类蛋白 t-SNARE配对,介导膜泡与靶膜融合,内含物进入高尔 基体。 • 被脱去的包被蛋白再循环利用。 11
网格蛋白有被小泡的形成 网格蛋白有被小泡的形成
a.配体同膜表面特异 受体结合, 网格蛋白 装配的亚基结合上去, 使膜凹陷成小窝状。 。 b.通过出芽的 方式形成小泡, 在发动蛋白 发动蛋白的 发动蛋白 作用下与质膜 割离。
细胞生物学 翟中和版 总结笔记第七章

Cell biology细胞生物学第七章真核细胞内膜系统、蛋白质分选与膜泡运输细胞内被膜区分类:细胞质基质、细胞内膜系统、有膜包被的细胞器第一节细胞质基质的含义和功能一、细胞质基质的含义(1)含义:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质主要含有:(1)与代谢有关的许多酶(2)与维持细胞形态和物质运输有关的细胞质骨架结构细胞质基质是一个高度有序的体系,细胞质骨架纤维贯穿在粘稠的蛋白质胶体中,多数的蛋白质直接或间接地与骨架结合,或与生物膜结合,从而完成特定的功能。
细胞质基质主要是由微管、微丝和中间丝等相互联系形成的结构体系,蛋白质和其他分子以凝聚或暂时的凝聚状态存在,与周围溶液的分子处于动态平衡。
差速离心获得的胞质溶胶的组分和细胞质基质溶液成分很大不同。
胞质溶胶中的多数蛋白质可能通过弱键结合在基质的骨架纤维上。
二、细胞质基质的功能(1)蛋白质分选和转运N端有信号序列的蛋白质合成之后转移到内质网上,通过膜泡运输的方式再转运到高尔基体。
其他蛋白质的合成都在细胞质基质完成,并根据自身信号转运到线粒体、叶绿体、细胞核中,也有些蛋白驻留在细胞质基质中。
(2)锚定细胞质骨架(3)蛋白的修饰、选择性降解1 蛋白质的修饰辅基、辅酶与蛋白的结合磷酸化和去磷酸化糖基化N端甲基化(防止水解)酰基化2 控制蛋白质寿命N端第一个氨基酸残基决定寿命细胞质基质能够识别N端不稳定的氨基酸信号将其降解,依赖于泛素降解途径3 降解变性和错误折叠的蛋白质4 修复变性和错误折叠的蛋白热休克蛋白的作用第二节细胞内膜系统及其功能细胞内膜系统是指在结构、功能乃至发生上相互关联、由膜包被的细胞器或细胞结构。
研究方法:电镜技术免疫标记和放射自显影离心技术和遗传突变体分析一、内质网的形态结构和功能内质网是由封闭的管状或扁平囊状膜系统及其包被的腔形成的互相沟通的三维网络结构。
(一)内质网的两种基本类型糙面内质网和光面内质网。
糙面内质网:扁囊状整齐附着有大量核糖体功能:合成分泌性蛋白和膜蛋白光面内质网:分支管状,小功能:脂质合成,出芽位点部分细胞合成固醇类激素糙面内质网有20多种和光面内质网不同的蛋白,说明有特殊装置隔开两种内质网的组分。
第七章真核细胞内膜系统、蛋白质分选与膜泡运输-测试题(满分:70)

第七章真核细胞内膜系统、蛋白质分选与膜泡运输- 测试题(满分:70)一、选择题(共25小题,1~20题每题1分,21~25题每题2分)1、下列关于信号肽,最正确的一项是()A. 是C端的一段氨基酸序列 C. 具有信号作用,但不被切除B. 是N端的一段氨基酸序列 D. 跨膜运输后要被切除2、细胞质基质中合成,到内质网上继续合成的蛋白的定位序列为()A. 信号肽 C. 转运肽B. 导肽 D. 信号斑3、参与蛋白质合成与运输的一组细胞器是()A. 核糖体、内质网、高尔基体 C. 细胞核、微管、内质网B. 线粒体、内质网、溶酶体 D. 细胞核、内质网、溶酶体4、台-萨氏病是一种与溶酶体有关的遗传缺陷病,主要是()缺乏而不能水解神经节苷脂GM2。
A. 磷酸二酯酶 C. β-氨基己糖酯酶AB. N-乙酰氨基转移酶 D. 腺苷酸环化酶5、指导蛋白质转运到线粒体上的氨基酸序列被称为()A. 导肽 C. 转运肽B. 信号肽 D. 新生肽6、溶酶体的H+ 浓度比细胞质基质中高()A. 5倍 C. 50倍B. 10倍 D. 100倍以上7、下面()不是在粗面内质网上合成的A. 抗体 C. 胶原蛋白B. 溶酶体膜蛋白 D. 核糖体蛋白8、下列细胞器中的膜蛋白在粗面内质网上合成的为()A. 叶绿体 C. 过氧化物酶体B. 线粒体 D. 溶酶体9、具运输和分拣内吞物质的细胞器是()A. 有被小体 C. 胞内体B. 滑面内质网 D. 溶酶体10、下列()是特化的内质网A. 肌质网 C. 乙醛酸循环体B. 脂质体 D. 残余小体11、真核细胞中下列()细胞器或细胞结构上不可能有核糖体存在A. 内质网 C. 细胞核膜B. 细胞质基质 D. 细胞质膜12、下列细胞器中,有极性的是()A. 溶酶体 C. 线粒体B. 微体 D. 高尔基体13、蛋白质的糖基化及其加工、修饰和寡糖链的合成是发生在高尔基体的()A. 顺面管网状结构 C. 反面管网状结构B. 中间膜囊 D. 反面囊泡14、合成后的磷脂被()转运至过氧化物酶体的膜上A. 磷脂转位因子 C. 膜泡B. 磷脂转换蛋白 D. 细胞骨架15、真核细胞中被称为异质性细胞器的是()A. 溶酶体 C. 内质网B. 核糖体 D. 高尔基体16、细胞中合成脂类的重要场所是 ( )A. 粗面内质网 C. 高尔基体B. 光面内质网 D. 胞质溶胶17、真核细胞内生物大分子运输的交通枢纽是()A. 溶酶体 C. 内质网B. 胞内体 D. 高尔基体18、真核细胞中生物大分子的合成基地是()A. 溶酶体 C. 内质网B. 胞内体 D. 高尔基体19、溶酶体的()借助M6P分选途径完成其分类、转运。
细胞学第7章复习题

第七章真核细胞内膜系统、蛋白质分选与膜泡运输1一.名词解释1.细胞内膜系统:位于细胞质内,在结构、功能及发生上密切相关的内膜包绕形成的细胞器或细胞结构,是真核细胞特有的结构。
主要包括内质网、高尔基体和溶酶体等。
2.初级溶酶体:是指刚从高尔基体的边缘膨大分离出来,还未同消化物融合的潜伏状态,无活性。
3.次级溶酶体:是酶在进行或完全消化作用的溶酶体,内含水解酶和相应的底物。
根据自身消化物质得来源不同,分为自噬溶酶体和异噬溶酶体4. 细胞质基质:真核细胞的细胞质中除去可分辨的细胞器以外的胶状物质,主要含有与中间代谢有关的酶和维持细胞形态、胞内物质运输有关的细胞质骨架结构等。
二.填空题1.在内质网上合成的蛋白主要包括分泌蛋白、膜整合蛋白、细胞器驻留蛋白等。
2.蛋白质的糖基化修饰主要分为N-连接的糖基化修饰,指的是蛋白质上的天冬酰胺残基与N-乙酰葡萄糖胺直接连接,和O-连接的糖基化修饰,指的是蛋白质上的丝氨酸或苏氨酸残基与N-乙酰半乳糖胺直接连接。
3.肌细胞中的内质网异常发达,被称为肌质网。
4.原核细胞中核糖体一般结合在细胞质膜上,而真核细胞中则结合在粗面内质网上。
5.真核细胞中,光面内质网是合成脂类分子的细胞器。
6.内质网的标志酶是葡萄糖-6-磷酸酶。
7.细胞质中合成的蛋白质如果存在信号肽,将转移到内质网上继续合成。
如果该蛋白质上还存在停止转移序列,则该蛋白被定位到内质网膜上。
8.高尔基体的标志酶是糖基转移酶。
9.具有将蛋白进行修饰、分选并分泌到细胞外的细胞器是高尔基体。
10.被称为细胞内大分子运输交通枢纽的细胞器是高尔基体。
11.蛋白质的糖基化修饰中,N-连接的糖基化反应一般发生在内质网中,而O-连接的糖基化反应则发生在和高尔基体中。
12.蛋白质的水解加工过程一般发生在高尔基体中。
13.从结构上高尔基体主要由顺面膜囊、中间膜囊和反面膜囊和囊泡组成。
14.植物细胞中与溶酶体功能类似的结构是圆球体、中央液泡和糊粉粒。
蛋白质分选膜泡运输

蛋白质分选膜泡运输概述蛋白质在细胞内的运输是维持细胞结构和功能的关键过程之一。
膜泡运输是一种重要的转运机制,通过分选膜泡将特定的蛋白质从细胞的起始地点(如内质网)运输到目的地(如高尔基体或细胞膜)。
蛋白质分选膜泡运输是由细胞质中的膜泡蛋白和适应性分选机制共同完成的。
分选膜泡的形成分选膜泡的形成是通过蛋白质的包裹和膜融合过程来实现的。
在起始地点,适应性分选机制识别目标蛋白质,并结合膜泡蛋白将其包裹成膜泡。
膜泡蛋白包裹蛋白质的过程需要依赖GTP酶Dynamin的作用,通过促进膜泡的分离和裂解来完成分选膜泡的形成。
一旦形成,膜泡会通过蛋白质的动力蛋白(如动力蛋白)来进行运输。
分选膜泡的运输分选膜泡的运输过程可以分为两个步骤:膜泡的运动和目的地的识别与融合。
膜泡的运动膜泡的运动是由动力蛋白(如肌动蛋白和微管蛋白)驱动的。
动力蛋白通过与膜泡蛋白的互作用来促进膜泡的移动。
这些动力蛋白通过细胞骨架的重排或膜泡附近的转运蛋白来定向膜泡的运动方向。
膜泡的运动过程中,蛋白质的特异信息将指导膜泡的准确运输。
目的地的识别与融合膜泡到达目的地后,需要通过与目的地膜的识别与融合来释放蛋白质。
这一过程主要由SNARE(SNAP 受体)蛋白质介导。
SNARE蛋白质分布在膜泡和目的地膜上,它们通过相互作用来促进膜泡与目的地膜的融合。
通过SNARE蛋白质的特异性识别和配对,只有适当的膜泡与目的地才能发生融合,从而实现蛋白质的释放和细胞功能的维持。
蛋白质分选膜泡运输的调控蛋白质分选膜泡运输是一个非常精确和复杂的过程,需要多种分子机器的协同作用来进行调控。
分选膜泡蛋白的调控膜泡蛋白是分选膜泡形成的关键组成成分,其正确的表达和定位是蛋白质分选膜泡运输的前提条件。
细胞通过转运蛋白的合成、翻译后修饰和定向转运等机制来调控膜泡蛋白的生成和定位。
适应性分选机制的调控适应性分选机制是指细胞通过特定的受体和配体相互作用来识别目标蛋白质并将其包裹成膜泡的过程。
细胞生物学膜泡运输与蛋白质分选

高尔基复合体与细胞内的膜泡运输高尔基符合体在分泌蛋白和细胞质膜蛋白的形成过程中不仅起着加工的作用,同时还有形成囊泡,进行包装和运输的作用。
分泌型蛋白、多数细胞质膜的膜蛋白都是在糙面内质网上合成后,经高尔基体的加工与分装通过膜泡运输的方式输送到细胞表面;而细胞质外的大分子和颗粒性物质及细胞质膜的膜蛋白也会通过胞饮或吞噬作用,以膜泡运输的方式进入细胞内。
高尔基复合体不论在向外运输的膜泡转移中,还是在内吞形成的膜泡转移中都起着重要的作用,且高尔基复合体中的G蛋白对高尔基复合体膜泡运输具有调控作用。
在细胞内的膜泡运输中,至少有10种以上的运输小泡参与膜泡运输过程,有三类。
COPl 有被小泡、COPll又被小泡、成笼蛋白有被小泡,前二者主要起始于内质网和高尔基复合体的运输,而后者主要调节起始于高尔基复合体和质膜的运输。
膜泡运输的类型1.成笼蛋白有被小泡起源于高尔基复合体的反面网状结构,其衣被主要成分为成笼蛋白和衔接蛋白。
结构上外层为由成笼蛋白构成的蜂巢样网络结构,内壳则由衔接蛋白构成,覆盖在细胞质基质侧的膜泡表面。
成笼有被小泡主要是负责蛋白质从高尔基体反面网状结构向质膜、胞内体或溶酶体和植物液泡运输。
此外,也参与胞内体到溶酶体的运输以及受体介导的内吞途径。
2.COPll有被小泡COPll有被小泡主要参与从内质网到高尔基复合体的物质运输,由五种蛋白亚基构成。
COPll蛋白能识别并结合跨膜内质网蛋白质膜一段的信号序列;而跨膜内质网蛋白的腔面一端作为受体与内质网腔中的可溶性蛋白结合。
COPll有被小泡具有对转运物质的选择性并使之浓缩。
3.COPl有被小泡主要负责回收与转运内质网逃逸蛋白返回内质网,而且在“糙面内质网-高尔基复合体-分泌泡-细胞表面”的蛋白质转运过程中,还行使非选择性批量运输的功能。
以上三种小泡介导的膜泡运输,需依赖于多种GTP结合蛋白对膜交换的时空变化进行调控。
膜泡运输的定向为了确保膜运输的有序进行,运输小泡在正确识别其将要与之融合的靶膜是必须具有高度选择性。
细胞生物学 第七章 细胞内膜系统及蛋白质分选与泡膜运输

周围大小不等的囊泡——物质运输
高尔基体与细胞骨架关系密切; 高尔基的膜囊上存在微管的马达蛋白(
kinesin)和微丝的马达蛋白(myosin)。最近 还发现特异的血影蛋白(spectrin)网架 。它 们在维持高尔基体动态的空间结构以及复杂的 膜泡运输中起重要的作用。
❖ 残余小体(residual body),次级溶酶体未被消化 的残渣,又称后溶酶体。
用电镜细胞化学技术显示其中含有的酸性磷酸酶, M:线粒体,L:溶酶体(朴英杰)
动物细胞溶酶体系统示意图
溶酶体膜的特征: 嵌有质子泵,形成和维持溶酶体中酸性的内环境; 具有多种载体蛋白用于水解的产物向外转运; 膜蛋白高度糖基化,可能有利于防止自身膜蛋白的
❖ 二、内膜系统的结构与功能 ❖ (二)高尔基体( Golgi complex ) ❖ 2、功能
❖ (3)蛋白酶的水解和其他加工过程
蛋白质在高尔基体中酶解加工的几种类型
无生物活性的蛋白原(proprotein)高尔基体切除 N-端或两端的序列成熟的多肽。如胰岛素、胰高血糖 素及血清白蛋白等
蛋白质前体高尔基体水解同种有活性的多肽,如 神经肽等
一种分选途径。
❖ 二、内膜系统的结构与功能
❖ (二)高尔基体( Golgi complex ) ❖ 2、功能
(1)高尔基体与细胞的分泌活动 ❖ 蛋白质的分选及其转运 ❖ 溶酶体酶的分选 (2)蛋白质糖基化及其修饰
(3)蛋白酶的水解和其他加工过程
蛋白质糖基化类型
特征
N-连接
O-连接
1. 合成部位
2. 合成方式
细胞生物学膜泡运输与蛋白质分选
高尔基复合体与细胞内的膜泡运输高尔基符合体在分泌蛋白和细胞质膜蛋白的形成过程中不仅起着加工的作用,同时还有形成囊泡,进行包装和运输的作用。
分泌型蛋白、多数细胞质膜的膜蛋白都是在糙面内质网上合成后,经高尔基体的加工与分装通过膜泡运输的方式输送到细胞表面;而细胞质外的大分子和颗粒性物质及细胞质膜的膜蛋白也会通过胞饮或吞噬作用,以膜泡运输的方式进入细胞内。
高尔基复合体不论在向外运输的膜泡转移中,还是在内吞形成的膜泡转移中都起着重要的作用,且高尔基复合体中的G蛋白对高尔基复合体膜泡运输具有调控作用。
在细胞内的膜泡运输中,至少有10种以上的运输小泡参与膜泡运输过程,有三类。
COPl 有被小泡、COPll又被小泡、成笼蛋白有被小泡,前二者主要起始于内质网和高尔基复合体的运输,而后者主要调节起始于高尔基复合体和质膜的运输。
膜泡运输的类型1.成笼蛋白有被小泡起源于高尔基复合体的反面网状结构,其衣被主要成分为成笼蛋白和衔接蛋白。
结构上外层为由成笼蛋白构成的蜂巢样网络结构,内壳则由衔接蛋白构成,覆盖在细胞质基质侧的膜泡表面。
成笼有被小泡主要是负责蛋白质从高尔基体反面网状结构向质膜、胞内体或溶酶体和植物液泡运输。
此外,也参与胞内体到溶酶体的运输以及受体介导的内吞途径。
2.COPll有被小泡COPll有被小泡主要参与从内质网到高尔基复合体的物质运输,由五种蛋白亚基构成。
COPll蛋白能识别并结合跨膜内质网蛋白质膜一段的信号序列;而跨膜内质网蛋白的腔面一端作为受体与内质网腔中的可溶性蛋白结合。
COPll有被小泡具有对转运物质的选择性并使之浓缩。
3.COPl有被小泡主要负责回收与转运内质网逃逸蛋白返回内质网,而且在“糙面内质网-高尔基复合体-分泌泡-细胞表面”的蛋白质转运过程中,还行使非选择性批量运输的功能。
以上三种小泡介导的膜泡运输,需依赖于多种GTP结合蛋白对膜交换的时空变化进行调控。
膜泡运输的定向为了确保膜运输的有序进行,运输小泡在正确识别其将要与之融合的靶膜是必须具有高度选择性。
细胞生物学 第7章 真核细胞内膜系统蛋白质分选与膜(共103张PPT)
一,其意义在于:大大增加了细胞内膜的表面积;为多种酶特
别是多酶体系提供了大面积的结合位点;酶系统的隔离与连接;
蛋白质、糖、脂肪的合成、加工和包装;运输分泌物;扩散屏
障及膜电位建立;离子梯度的维持等。
二、内质网的形态结构与功能
(一)、内质网的形态结构及化学组成
②赋予蛋白质传导信号的功能;
③某些蛋白只有在糖基化之后才能正确折叠。
糖基一般连接在4种氨基酸上,分为2种:
(O-linked glycosylation):与
Ser、Thr和Hyp的OH连接,连接的糖为半乳糖
或N-乙酰半乳糖胺,在高尔基体上进行。
(N-linked glycosylation):与
丝氨酸、苏氨酸、
羟赖氨酸、羟脯氨酸
一般1~4个糖残基,
但ABO血型抗原较长
N—乙酰半乳糖胺等
内质网上进行N-连接的糖基化。
糖分子首先被糖基转移酶转移到膜上的磷酸多萜醇
(dolichol phosphate)分子上,装配成寡糖链。
再被寡糖转移酶转到新合成肽链特定序列(Asn-X-Ser
或Asn-X-Thr)的天冬酰胺残基上。
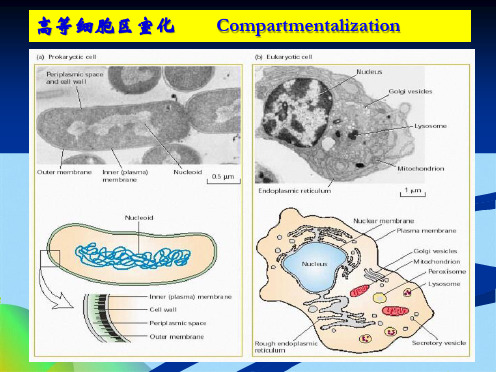
高等细胞区室化
Compartmentalization
质膜
内膜
生物膜
原核细胞 – 质膜构成的单一区室
真核细胞 – 内膜 – 各种细胞器
内膜系统出现的意义:
各自独立,各司其职
相互依存,协调同一
大大提高了细胞代谢反应的效率
细胞核
细
胞
细胞质
过氧化物酶体
线粒体
细胞骨架
翟中和细胞生物学第七章总结2(名词解释)

第七章真核细胞内膜系统、蛋白质分选与膜泡运输1.细胞质基质:在真核细胞的细胞质中,除去可分辨的细胞器以外的胶状物质,也称胞质溶胶,内含水、无机离子、酶以及可溶性大分子和代谢产物。
21、许多中间代谢过程在细胞质基质中进行。
包括糖酵解过程、磷酸戊糖途径、糖醛酸途径、糖原合成与分解以及蛋白质与脂肪酸的合成等。
2、细胞质骨架是细胞质基质的主要结构成分,与维持细胞形态、细胞运动、物质运输及能量传递有关,而且也是细胞质基质结构体系的组织者,为细胞质基质中其他成分和细胞器提供锚定位点。
3、与蛋白质的修饰及选择性降解有关。
①蛋白质的修饰,在细胞质中发生的蛋白质修饰的类型主要有:辅酶或辅基与酶的共价结合;磷酸化与去磷酸化,用以调节很多蛋白质的生物活性;糖基化作用;对某些蛋白质的N端进行甲基化修饰;酰基化。
②控制蛋白质的寿命。
③降解变性和错误折叠的蛋白质。
④帮助变性或错误折叠的蛋白质重新折叠,形成正确的分子构象。
这一功能主要靠热休克蛋白来完成。
3①辅酶或辅基与酶的共价结合。
②磷酸化与去磷酸化,用以调节很多蛋白质的生物活性。
③糖基化作用:糖基化主要发生在内质网和高尔基体中,在细胞质基质中发现的糖基化是指在哺乳动物的细胞中把N-乙酰葡糖胺分子加到蛋白质的丝氨酸残基的羟基上。
④对某些蛋白质的N端进行甲基化修饰:这种修饰的蛋白质,如很多细胞支架蛋白和组蛋白等,不易被细胞内的蛋白质水解酶水解,从而使蛋白质在细胞中维持较长的寿命。
⑤酰基化:最常见的一类酰基化修饰是内质网上合成的跨膜蛋白在通过内质网和高尔基体的转运过程中发生的,它由不同的酶来催化,把软脂酸链共价地连接在某些跨膜蛋白的暴露在细胞质基质中的结构域;另一类酰基化修饰发生在诸如src基因和ras基因这类癌基因的产物上,催化这一反应的酶可识别蛋白中的信号序列,将脂肪酸链共价地结合到蛋白质特定的位点上。
如src基因编码的酪氨酸蛋白激酶与豆蔻酸的共价结合。
酰基化与否并不影响酪氨酸蛋白激酶的活性,但只有酰基化的激酶才能转移并靠豆蔻酸链结合到细胞质膜上。
第七章真核细胞内膜系统、蛋白质分选与膜泡运输

◆扁平囊泡(sacuules)
◆小囊泡(vesicle) ◆大囊泡(vacuoles)
The Golgi Apparatus
培养的上皮细胞中高尔基体的分布 (高尔基体为红色,核为绿色)
高尔基体的形态结构
高尔基体的极性
◆高尔基内侧网络 (cis-Golgi network, CGN) 顺面、形成面 ◆中间潴泡 (medial cisternae) ◆高尔基外侧网络 (trans Golji network,TGN) 外侧面、成熟面
磷脂,磷脂酰胆碱含量较高,鞘磷脂含量较少,没有或很 少含胆固醇。
ER约有30多种膜结合蛋白,另有30多种位于内质网腔, 这些蛋白的分布具有异质性,如:葡糖-6-磷酸酶,普遍存
在于内质网,被认为是标志酶,核糖体结合糖蛋白
(ribophorin)只分布在RER,P450酶系只分布在SER。
内质网的模式图
解毒作用
2+ Ca 离子浓度的调节作用
磷脂的合成
磷脂交换蛋白的作用
解毒作用
◆光面内质网含有丰富的氧化酶系统(如细 胞色素P450、NADH细胞色素C还原酶等)能 使许多有害物质解毒 ,转化为易于排出的 物质。
粗面内质网的功能 --信号肽与蛋白质运输
核糖体循环
?
二、ER的功能(RER)
第二节
细胞内膜系统及其功能
定义:指在结构、功能乃至发生上相互关联、由膜包被的 细胞器或细胞结构, 主要包括内质网、高尔基体、溶酶体、
胞内体和分泌泡等。
功能:区隔化;增加内表面积,提高代谢和调节能力。 从系统发生来看内膜系统起源于质膜的内陷和内共生。 从个体发生来看新细胞的内膜系统来源于原有内膜系统的 分裂,具有核外遗传的特性。
细胞生物学总结(复习重点)——7.内膜系统、蛋白质分选、膜泡运输

1、细胞质基质:真核细胞的细胞质中除去细胞器和内含物以外的、较为均质半透明的液态胶状物称为细胞质基质或胞质溶胶。
4、内膜系统:细胞内在结构、功能乃至发生上相关的、由膜围绕的细胞器或细胞结构的统称,主要包括内质网、高尔基体、溶酶体、胞内体、分泌泡等。
2、微粒体:为了研究ER的功能,常需要分离ER膜,用离心分离的方法将组织或细胞匀浆,经低速离心去除核及线粒体后,再经超速离心,破碎ER的片段又封合为许多小囊泡(直径约为100nm),这就是微粒体。
3、糙面内质网:细胞质内有一些形状大小略不相同的小管、小囊连接成网状,集中在胞质中,故称为内质网。
内质网膜的外表面附有核糖体颗粒,则为糙面内质网,为蛋白质合成的部位。
核糖体附着的膜系多为扁囊单位成分,普遍存在于分泌蛋白质的细胞中,其数量随细胞而异,越是分泌旺盛的细胞中越多。
5、分子伴侣:细胞中,这类蛋白能识别正在合成的多肽或部分折叠的多肽,并与多肽的一定部位相结合,帮助这些多肽的转移、折叠或组装,但其本身并不参与最终产物的形成。
6、溶酶体:溶酶体几乎存在于所有的动物细胞中,是由单层膜围绕、内含多种酸性水解酶类、形态不一、执行不同生理功能的囊泡状细胞器,主要功能是进行细胞内的消化作用,在维持细胞正常代谢活动及防御方面起重要作用。
7、残余小体:在正常情况下,被吞噬的物质在次级溶酶体内进行消化作用,消化完成,形成的小分子物质可通过膜上的载体蛋白转运至细胞质中,供细胞代谢用,不能消化的残渣仍留在溶酶体内,此时的溶酶体称为残余小体或三级溶酶体或后溶酶体。
残余小体有些可通过外排作用排出细胞,有些则积累在细胞内不被排出,如表皮细胞的老年斑、肝细胞的脂褐质。
8、蛋白质分选:细胞中绝大多数蛋白质均在细胞质基质中的核糖体上开始合成,随后或在细胞质基质中或转至糙面内质网上继续合成,然后,通过不同途径转运到细胞的特定部位并装配成结构与功能的复合体,参与细胞的生命活动的过程。
又称定向转运。
细胞生物学(8章真核细胞内的区域化)

Quality control of newly synthesized proteins---The role of N-linked glycosylation in ER protein folding
Functions of cytoplasmic matrix:
➢The Cytosol performs most of the cell’s intermediary metabolism. ➢The Cytosol is the site of protein synthesis and degradation or modification.
糖为N-乙酰葡糖胺。 • O-连接的糖基化:与Ser、Thr和Hyp的OH连接,
连接的糖为半乳糖或N-乙酰半乳糖胺,在高尔基 体上进行。
Protein glycosylation in RER
• 内质网上进行N-连接的糖基化。糖的供体为核苷 糖,如GDP-甘露糖、UDP-N-乙酰葡糖胺。
• 糖分子首先被糖基转移酶转移到膜上的磷酸多萜 醇分子上,装配成寡糖链。
蛋白质转入内质网合成的过程:
• 信号肽与SRP结合→肽链延伸终止→SRP与受体结 合→SRP脱离信号肽→肽链在内质网上继续合成 →信号肽切除→肽链延伸至终止。
• 这种肽链边合成边向内质网腔转移的方式,称为 co-translation。
Insertion of Transmembrane protein into the ER membrane
(1) the cytosol (2) the endomembrane system:
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
7
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
8
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
9
(一)信号假说
1975年Blobel和sabatini等提出了信号假说( signal hypothesis),即分泌性蛋白N端作为序列信号 肽(signal sequence或signal peptide),指导分泌 性蛋白到内质网膜上合成,在蛋白合成结束前信号肽被 切除。信号识别颗粒(SRP)和内质网膜上的信号识别颗 粒的受体(停泊蛋白,DP)等因子协助完成这一过程。
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
4
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
5
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
6
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
19
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
20
跨膜蛋白的类型及其取向
开启易位子
初生肽链
peptidase
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
10
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
11
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
12
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
13
易位子
链接子因子 新生蛋白
• 英文名称为translocon,或translocator 或translocation channel。易位子是与新生蛋白或多肽跨膜转运相关的多种蛋白的复 合物,其成员有Sec61、SRP、SRPR、TRAM及信胞生精物品课学件(第4版)© 高等教育出版社 2011
翟中和 王喜忠 丁明孝 主编
细胞生物学(第4版)
第7章 第二部分 蛋白质分选与膜泡运输
Copyright © 高精等品教课件育出版社 2011
1
细胞的社会联系
• 多细胞生物体内没有细胞是“孤立”的 • 通过细胞通讯、黏着、连接以及细胞与胞外基质的相互作用
构成复杂的细胞社会 • 决定细胞的行为和命运,包括结构与功能分化、位置以及生
死抉择,维持机体的稳态平衡
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
2
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
3
• Randy Schekman发现了囊泡传输所需的一 组基因;James Rothman阐明了囊泡是如 何与目标融合并传递的蛋白质机器; Thomas Südhof则揭示了信号是如何引导 囊泡精确释放被运输物的。
25
蛋白质分选的途径:
1、翻译后转运途径:在细胞质基质游离核糖体上完
成多肽链的合成,然后转运至膜围绕的细胞器,如线粒
体、叶绿体、过氧化物酶体及细胞核,或者成为细胞质
基质的可溶性驻留蛋白和支架蛋白
2、共转运翻译途径:蛋白质合成在游离核糖体
上起始之后由信号肽引导转移至糙面内质网,然后新生
肽边合成边转入糙面内质网中,再经高尔基体加工包装
18
(二)信号肽与共翻译
肽链边合成边转移至内质网腔中的方式称为共转移。 跨膜蛋白的起始转移序列和终止转移序列
开 始 转移序列:蛋白质氨基末端的信号序列除作为信号被SRP 识别外,还具有起始穿膜转移的作用。 内 在 停止转移 锚 定 序列:肽链中还有某些序列与内质网膜具 有很强的亲合力而结合在脂双层之中,能阻止肽链继续进入 内质网腔,使其成为跨膜蛋白质,称为停止转移序列。 内在信号信号序列: 跨膜蛋白的类型及其取向 起始转移序列和终止转移序列的数目决定多肽跨膜次数
14
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
15
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
16
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
17
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
21
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
22
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
23
• 信号序列(signal sequence):存在于蛋白质一级结构上 的线性序列,通常由15-60个氨基酸残基组成,有些信号序 列在完成蛋白质的定向转移后被信号肽酶( signal peptidase)切除;通常信号序列对所引导的蛋白质没有特 异性要求。
• 信号斑(signal patch):存在于完成折叠的蛋白质中,构 成信号斑的信号序列之间可以不相邻,折叠在一起构成蛋白 质分选的信号。
• 每一种信号翟中序和 王列喜决忠 丁定明特孝 主殊编 的细胞蛋生精物白品课学质件(第转4版运)©方高等向教。育出版社 2011
24
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
并有其膜泡运送至溶酶体、细胞质膜或分泌到胞外。
翟中和 王喜忠 丁明孝 主编 细胞生精物品课学件(第4版)© 高等教育出版社 2011
26
蛋白质分选的类型:
1、蛋白质的跨膜转运:在细胞质基质中合成的 蛋白质转运到内质网、线粒体、质体和过氧化物酶 体等细胞器;
2、膜泡运输:通过不同类型的转运小泡从糙面 内质网合成部位转运至高尔基体,进而分选转运至 细胞的不同部位;