Ach受体信号通路
乙酰胆碱及受体的作用和与人类健康的关系

乙酰胆碱及受体的作用和人类健康王帅(09级辽宁大学生命科学院生物技术专业本科生291303118)[ 摘要] 乙酰胆碱( acetylcho line, ACh) 是一种经典的兴奋性神经递质, 通过结合特异受体, 在神经细胞之间或神经细胞与效应器细胞之间中起着信息传递作用。
ACh 及其受体存在于从细菌到人类、从神经细胞到其他多种非神经细胞中, 提示它是一类与系统发生相关的古老分子, 可能不仅仅具有作为生理性递质的传递功能。
多种人类疾病与ACh 及其受体相关, 尤其是近年来的研究发现ACh 及其受体在多种肿瘤中发挥自分泌和旁分泌因子作用, 参与细胞的生长调节, 甚至与肿瘤的发生发展相关。
因此, ACh 涉及到神经系统外非胆碱能传递的作用显得格外引人注目, 可能成为新的肿瘤治疗靶标。
[ 关键词] 受体;乙酰胆碱;人类健康;肿瘤1 乙酰胆碱及其受体简述乙酰胆碱( acetylcho line, ACh) 是一种经典的兴奋性神经递质, 包括外周神经如运动神经、自主神经系统的节前纤维和副交感神经节后纤维均合成和释放这种神经递质。
ACh 由胆碱( choline) 和乙酰辅酶A 合成, 由胆碱乙酰化酶( choline acety lase, ChAT ) 催化,随后进入囊泡贮存。
当动作电位沿神经到达神经末梢时, 触发神经末梢Ca2+ 通道开放,囊泡与突触前膜融合、破裂, ACh 释放入突触间隙或接头间隙, 作用于突触后膜或效应细胞膜的乙酰胆碱受体( acet ylcholine recepto rs, AChRs) 引起生理效应。
其中位于副交感神经节后纤维所支配的效应器细胞膜的胆碱受体对以毒蕈碱为代表的拟胆碱药较为敏感, 故称为毒蕈碱型胆碱受体( muscar inic acet ylcho line receptor s, mAChRs) ; 位于神经节细胞膜和骨骼肌细胞膜的胆碱受体对烟碱比较敏感, 故称为烟碱型胆碱受体( nicot inic acety lcholine r ecepto rs,nAChRs) 。
各种传导通路汇总

各种传导通路汇总在生物学和医学领域中,传导通路是指生物体内用于传递信号和信息的分子或细胞机制。
这些通路可以包括细胞内信号传导通路、神经传导通路、电传导通路和生化传导通路等。
以下是一些常见的传导通路的汇总:1.细胞内信号传导通路:-G蛋白偶联受体通路:G蛋白偶联受体通过与G蛋白结合,激活下游效应器分子,如磷脂酶C(PLC)、腺苷酸酰化酶(AC)等。
-酪氨酸激酶受体通路:酪氨酸激酶受体通过自身磷酸化,激活下游蛋白激酶,如丝裂原活化蛋白激酶(MAPK)等。
- 精氨酸磷酸化酶(PPA)通路:PPA通过磷酸化下游效应器蛋白,如磷酸酪氨酸酉激酶(Src)等。
2.神经传导通路:-突触传导通路:神经元通过突触间的化学信号传递,将信息从一个神经元传递到另一个神经元。
-神经递质受体通路:神经递质通过结合相应的受体,触发信号传导,如乙酰胆碱受体、谷氨酸受体等。
-非突触传导通路:神经元通过细胞间的直接电流传导或电化学信号传导,实现信息的传递。
3.电传导通路:-肌动电传导通路:心肌细胞通过特定通道中的离子流动,产生心脏的电活动,如心房肌、心室肌等。
-脑电传导通路:神经元在脑内产生的电活动通过特定通道传导,如脑电图中的α波、β波等。
4.生化传导通路:-糖代谢通路:包括糖酵解、糖异生、糖原合成等通路,用于维持细胞能量供应和新陈代谢的平衡。
-脂代谢通路:包括脂肪酸β氧化、胆固醇合成等通路,用于脂质的合成和分解。
-蛋白质合成通路:包括转录、转译和翻译等通路,用于合成特定的蛋白质。
除了以上提到的通路之外,还有很多其他种类的传导通路,如细胞凋亡通路、细胞周期调控通路、免疫传导通路等。
这些通路通过特定的分子相互作用和信号转导,实现了生物体内的各种生理和生物化学过程。
总结起来,传导通路是生物体内用于传递信号和信息的分子或细胞机制。
它们可以涉及细胞内信号传导、神经传导、电传导和生化传导等多个层面,通过特定的分子相互作用和信号转导,实现了生物体内的各种生理和生物化学过程。
信号通路关键蛋白质分子

信号通路关键蛋白质分子信号通路是一系列相互作用的生化反应,用于传递细胞内外的信息。
关键蛋白质分子在信号通路中发挥着至关重要的作用,它们调节着信号的传导和细胞的响应。
本文将介绍几种常见的信号通路关键蛋白质分子,包括G蛋白偶联受体、酪氨酸激酶、细胞内信号传导蛋白和转录因子。
G蛋白偶联受体是一类广泛存在于生物体内的膜受体,它们通过与G蛋白结合,介导细胞对外界信号的感知和响应。
G蛋白偶联受体分为三类:Gs、Gi和Gq。
Gs蛋白激活腺苷酸环化酶,产生第二信使cAMP,进而激活蛋白激酶A,调节细胞内的代谢和信号传导。
Gi 蛋白抑制腺苷酸环化酶活性,降低cAMP水平,起到负调节的作用。
Gq蛋白则激活磷脂酶C,产生第二信使二酰甘油和肌醇三磷酸,参与细胞内信号传导。
酪氨酸激酶是一类重要的信号传导蛋白,它们通过磷酸化反应调节多种细胞过程。
酪氨酸激酶分为受体型和非受体型。
受体型酪氨酸激酶包括表皮生长因子受体(EGFR)、血小板源性生长因子受体(PDGFR)和肿瘤坏死因子受体(TNFR),它们在细胞增殖、分化和存活等过程中发挥重要作用。
非受体型酪氨酸激酶主要包括SRC 家族激酶和JAK家族激酶,它们参与免疫应答、细胞凋亡和细胞迁移等生物学过程。
细胞内信号传导蛋白是信号通路中的另一类关键分子。
它们包括蛋白激酶C(PKC)、丝裂原激活蛋白激酶(MAPK)和磷脂酰肌醇3-激酶(PI3K)等。
PKC是一类丝氨酸/苏氨酸激酶,参与细胞增殖和分化等过程。
MAPK是一类丝氨酸/苏氨酸激酶,它们通过磷酸化反应调节细胞的生长、分化和凋亡等。
PI3K是一类磷脂酰肌醇激酶,它参与细胞的增殖、存活和迁移等过程。
转录因子是一类能够结合到DNA上调节基因转录的蛋白质。
它们在信号通路中起着重要的调节作用。
转录因子包括核转录因子(NF-κB)、激活蛋白-1(AP-1)和CREB等。
NF-κB参与细胞的免疫应答和炎症反应等过程。
AP-1是由c-Jun和c-Fos等蛋白质组成的转录因子复合物,参与细胞的增殖和凋亡等过程。
概述受体酪氨酸激酶介导的信号通路的组成、特点及其主要功能。

概述受体酪氨酸激酶介导的信号通路的组成、特点及其主要功能。
1.引言1.1 概述受体酪氨酸激酶介导的信号通路是细胞内一种重要的信号传导机制,它通过调控各种生物化学反应来参与细胞内的多种生理过程。
该信号通路的组成主要包括受体酪氨酸激酶、底物和下游的信号分子等。
受体酪氨酸激酶是一类能够磷酸化酪氨酸残基的酶,它能够通过与外界的信号分子结合,诱导其自身的激活,进而引发一系列的生物效应。
受体酪氨酸激酶可以被分为两种类型:单体型和受体型。
单体型受体酪氨酸激酶主要包括一些细胞内酪氨酸激酶;受体型受体酪氨酸激酶则包括一些质膜上的受体酪氨酸激酶。
这些受体酪氨酸激酶在结构上存在一定的相似性,但在功能上却可能有所差异。
受体酪氨酸激酶介导的信号通路具有一些特点。
首先,它是一种高度调控的信号传导网络,可以根据不同的外界刺激改变其活性。
其次,受体酪氨酸激酶的激活往往能够启动多条平行的信号通路,从而实现更为复杂的细胞反应。
此外,该信号通路具有信号传导速度快、反应机制多样等特点。
受体酪氨酸激酶介导的信号通路在许多生理过程中发挥着重要的功能。
例如,它参与了细胞的生长、增殖和分化过程;调节了细胞的凋亡和存活等。
此外,该信号通路还与多种疾病的发生和发展密切相关,如癌症、炎症和神经系统疾病等。
综上所述,受体酪氨酸激酶介导的信号通路具有复杂的组成、多样的特点和重要的功能。
深入了解该信号通路的组成、特点及其主要功能,对于揭示细胞信号传导的机制,以及发展相关疾病的治疗策略具有重要意义。
文章结构部分应该介绍文章的组织结构和各个章节的内容概述。
以下是一种可能的写作方式:1.2 文章结构本文将分为三个主要部分进行论述,分别是引言、正文和结论。
引言部分将首先对受体酪氨酸激酶介导的信号通路进行概述,包括其在细胞内的重要性和作用机制。
接着,本部分将介绍文章的结构和各个章节的内容。
正文部分将详细探讨受体酪氨酸激酶的组成、特点和主要功能。
在2.1小节中,将介绍受体酪氨酸激酶的组成成分,包括受体酪氨酸激酶本身和相关的配体和受体。
心血管系统中NO信号传导通路的调控机制

心血管系统中NO信号传导通路的调控机制氧化氮(NO)是一种重要的信号分子,它能够影响多种生物学过程,其中包括心血管系统的调控。
在心血管系统中,NO信号传导通路的调控机制非常复杂,涉及多种分子和细胞类型的相互作用,有助于维持心血管系统的正常功能。
本文将探讨心血管系统中NO信号传导通路的调控机制。
1. NO的生成和作用NO是一种气体信号分子,由内皮细胞和其他细胞类型通过NO合成酶(NOS)进行合成。
在内皮细胞中,胆碱能诱导乙酰胆碱(ACh)受体的激活,从而使得内皮细胞产生NO。
NO能够通过多种机制调控心血管系统的功能,其中包括:(1)血管松弛作用。
NO能够通过激活可溶性鸟苷酸环化酶(sGC)上的环磷酸鸟苷酸(cGMP)的形成,从而导致平滑肌松弛,血管扩张。
(2)抗凝作用。
NO能够调节血小板活性和凝血酶的活性,从而减轻凝血风险。
(3)抗粘附作用。
NO能够减少粘附分子的表达,降低血细胞在血管内膜上的黏附。
2. NO的信号传导通路NO的作用部分是通过下游信号传导通路实现的。
NO能够促进sGC上的cGMP形成,cGMP进一步激活PDE(cGMP-磷酸酶)从而降解cGMP,降低cGMP的含量。
cGMP的作用可以通过多种方式发挥,其中包括:(1)激活蛋白激酶G(PKG)和蛋白激酶A(PKA)。
(2)激活cGMP-敏感的离子通道。
(3)调节Ca2+离子通道。
(4)调节内质网钙离子释放。
(5)激活磷酸脂酰肌醇3-激酶(PI3K)/蛋白激酶B(Akt)信号通路等。
这些信号传导通路的作用可以进一步调节多种生物过程,包括负反馈调节NO 的合成、药物对心血管系统的作用、炎症、氧化应激等。
3. NO信号传导通路的调控机制在心血管系统中,NO信号传导通路的调控机制非常复杂,涉及多种分子和细胞类型的相互作用。
有许多分子对NO信号传导通路产生调节作用,其中包括:(1)cGMP-磷酸酶。
cGMP-磷酸酶能够降解cGMP,从而调节其作用。
第4章 细胞信号传导-导论

1. PTK受体( protein tyrosine kinase receptor ) PTK受体
EGFR的二聚化和自磷酸化 EGFR的二聚化和自磷酸化
配体与受体结合 受体二聚化 受体自磷酸化 底物磷酸化 级联反应 信号入核
EGFR signaling network
•
Molecular Targeting in Radiotherapy: Epidermal Growth Factor Receptor Theodore D. Chung and William C. Broaddus,Mol. Interv. 2005;5:15-19.
一、胞外化学信号 (Primary message)
• 物理信号 • 化学信号 • 生物信号
胞外化学信号
能与受体呈特异性结合的生物活性分子称 能与受体呈特异性结合的生物活性分子称 受体呈特异性结合的生物 配体( 配体(ligand)。 )。
胞外化学信号可以分为2 胞外化学信号可以分为2类:
1.可溶性的 1.可溶性的 2.膜结合的 2.膜结合的
Cell-signal 专题
/news/report/ 2004/signal.htm
细胞信号(生命科学新视野3), 细胞信号(生命科学新视野3), 3) 多克希, 2007年11月 (美)多克希,科学出版社 , 2007年11月 细胞信号传导的分子基础与功能调控,姜勇等, 细胞信号传导的分子基础与功能调控,姜勇等, 科学出版社,2005年 科学出版社,2005年9月 细胞信号传导(第三版),孙大业主编, 细胞信号传导(第三版),孙大业主编, ),孙大业主编 科学出版社,2001年10月 科学出版社,2001年10月
内容提纲
一、胞外信号 二、受体 三、第二信使 四、蛋白激酶系统 五、连接分子 六、细胞信号转导的效应 七、信号传导异常与疾病
2021生物物理学-生物物理学(精选试题)

生物物理学-生物物理学1、什么是生物物理学?2、生物物理学包含的分支学科(主要内容)有哪些?3、什么是生物物理技术?4、什么叫电磁波?5、什么叫波谱学(spectroscopy)和波谱技术?6、波谱学的物理基础是什么?7、波谱是如何产生的?8、波谱有哪些参数反映物质信息?9、波谱仪有哪些主要部件?10、简单分子的能级是如何构成的?11、紫外-可见吸收光谱是怎样形成的?12、紫外-可见吸收分光仪的基本构成是怎样的?13、紫外-可见吸收光谱术有什么基本应用?(只须回答四点即可)14、荧光是怎样产生的?15、荧光光谱的特点是什么?16、荧光光谱术的具体技术有哪些?17、荧光光谱术的基本应用有哪些?18、荧光光谱术和紫外-可见吸收光谱术有哪些异同?19、什么是红外吸收光谱?20、拉曼光谱是怎样形成的?21、红外与拉曼光谱的特点是什么?22、红外吸收光谱仪的构成是什么?23、红外吸收光谱术的特点是什么?24、红外与拉曼光谱术的基本应用有哪些?25、什么叫核磁共振?26、为什么同一种原子核共振吸收峰的频率并不相同?27、什么叫化学位移?28、什么叫谱线分裂?其产生原因是什么?29、什么是电子自旋共振技术?30、电子自旋共振和核磁共振的异同是什么?31、电子自旋共振波谱的特点是什么?32、电子自旋共振波谱的应用有哪些?33、组成细胞的小分子物质有哪些?有何作用?34、试述蛋白质的四级结构。
35、核酸分为哪两类?有何区别?36、试述DNA双螺旋结构的特点?有何功能?37、比较三种RNA的区别。
38、为什么说支原体是最小最简单的细胞?39、试比较原核细胞的异同,你能得出什么结论?40、细胞膜的主要成分是什么?有何功能?41、细胞膜的主要特征有哪些?有何生物学意义?42、试述被动运输的几种运输方式。
43、以Na+-K+泵为例,说明物质的主动运输过程。
44、膜受体的生物学特征是什么?有何意义?45、试述膜受体的类型及传导途径。
细胞的信号转导-学习笔记

cAMP
三磷酸肌醇(inositol triphosphate,IP)
二酰甘油(diacylglycerol,DG/DAG)
环-磷酸鸟苷(cyclic guanosine monophosphate,cGMP)
钙离子
花生四烯酸(arachidonic acid,AA)及其代谢产物
另外,cAMP还可直接作用于膜离子通道而产生信号转导作用,如直接门控超极化激活的环核苷酸门控阳离子通道(hyperpolarization-activated cyclic nucleotide-gated cation channel,HCN);cAMP还可通过cAMP直接激活的交换蛋白(exchange protein activated by cAMP,EPAC)激活Ras相关蛋白(Rap)介导的非cAMP-PKA通路,调节细胞的功能。
根据受体的氨基酸组成及跨膜区的特点,可将离子通道型受体分为三类受体超家族:
I型受体超家族:以烟碱型乙酰胆碱受体(nicotinic acetylcholine receptor,nAChR)为代表,常存在于神经元和神经肌肉接头处,有α2、β、γ、δ五个蛋白亚单位(四个亚基):
每一亚基的分子量为40~58kD,各含4个长度不同的穿膜区域,每个亚基的第二个跨膜区共同构成Na离子通道的内壁。乙酰胆碱(ACh)的结合位点位于α亚基的N末端区域,因此nAChR有两个ACh结合位点:
2、G蛋白效应器(Gprotein effector)
G蛋白效应器是指G蛋白直接作用的靶标,包括效应器酶、膜离子通道以及膜转运蛋白等。
效应器酶的作用是催化生成(或分解)第二信使物质,主要有:
腺苷酸环化酶(adenylyl cyclase,AC)
受体和信号转导通路的基本原理及其在疾病研究中的应用

受体和信号转导通路的基本原理及其在疾病研究中的应用随着科学技术的不断发展,对人体生物学、分子生物学和细胞生物学的研究逐渐深入,受体和信号转导通路逐渐成为研究的热点,因为它们在生命过程中发挥着重要的作用。
本文将重点介绍受体和信号转导通路的基本原理,并阐述其在疾病研究中的应用。
一、受体的基本功能和类型生物体中的受体是分子生物学研究的重要对象,它可以是蛋白质、酶、核酸或配体。
在细胞膜或细胞质中,受体对外界环境中的化学和物理刺激作出反应,引起细胞内信号传递,从而产生特定的生理、生化反应或调节细胞功能。
不同类型的受体有不同的信号传递途径,如离子通道、酶偶联型受体、G蛋白偶联型受体等。
其中,离子通道受体与在神经系统、神经肌肉接头等处超快速信号传递中扮演重要角色;酶偶联型受体主要介导细胞因子介导的生物学效应;而G蛋白偶联型受体则具有丰富、广泛的作用,涉及光信号转导、味觉和嗅觉感受、内分泌反应、细胞分裂和凋亡等各种生物学过程。
二、信号转导通路的基本原理细胞发生生理和病理变化时,通常是依赖于细胞内外发生的分子信号传递,信号传递的核心是信号转导通路。
信号转导通路是指外来信号(如激素、细胞因子、营养成分、神经递质等)通过激活细胞膜上的受体,从而引起一系列的分子信号传递步骤,最终导致细胞的生理反应。
信号转导通路步骤大致分为:受体激活、信号传递、下游反应和终末效应等四个步骤。
其中,受体和信号传递是信号转导的核心环节。
受体的激活是信号转导通路的第一步,它通常由外来的信号分子引起,通过细胞膜钙离子通道、酶偶联型受体、G蛋白偶联型受体等将外界信号传递到细胞内。
信号传递的过程涉及到多种信号分子、相互作用的蛋白及其调节因子等。
下游反应指对信号传递路径产生影响,如它可以是酶的激活、双向调节蛋白、转录因子激活和QCrh18等。
终末效应通常指生理反应,例如细胞分化、增殖、凋亡、迁移等。
三、受体和信号转导通路在疾病研究中的应用研究受体和信号转导通路在疾病研究中的应用,可以帮助人们更好地理解疾病发生机理以及寻找有效的治疗方式。
十八章细胞信号转导异常与疾病-精选文档

病毒性感染性疾病与G蛋白偶联受体的关系
HIV感染中:
趋化因子受体CXCR4(属G蛋白偶联受体) 是HIV进入CD4+细胞的辅受体;
CCR5是HIV进入巨噬细胞的辅受体
当其缺失突变对HIV感染具有抵抗。
多毛症,性早熟,低肾素性高血 压
分类
自身免疫性受体病
累及的受体
主要临床特征
重症肌无力 自身免疫性甲状腺病
艾迪生病 II型糖尿病 继发性受体异常
Ach受体
刺激性TSH受体 抑制性TSH受体
活动后肌无力
甲亢和甲状腺肿大 甲状腺功能减退
ACTH受体
色素沉着,乏力,血压低 继发性受体异常
胰岛素受体
高血糖,血浆胰岛素正常或升高
R:Arg;W:Trp;Q:Gln
四、G蛋白偶联受体与感染性疾病
信号传导过程) 霍乱(cholera):
实质是细胞内cAMP含量急剧升高所致。 霍乱毒素cholera toxin CT:A、B亚基组成
A亚基为CT的毒性部分,A、B结合无毒性作用,A 亚基释放才能发挥作用;
⑤受体再循环障碍 基因突变使内吞的受体不能在酸性pH下与LDL解离,
受体在细胞内降解。
因LDL受体数量减少或功能异常,其对血浆LDL的清除能 力降低,患者出生后血浆LDL含量即高于正常,发生动脉粥 样硬化的危险也显著升高。患者多于40~50岁发生冠心病。
(二)自身免疫性受体病
因体内产生抗受体的自身抗体而引起的疾病,可因刺激性 抗体引起细胞对配体的反应性增强,或因阻断性抗体干扰配 体与受体的结合,导致细胞的反应性降低。
07-3-细胞内受体介导的信号转导

12.3 细胞内受体介导的信号传递同学们好!前面我介绍细胞信号转导系统的构成要素时,已经讲过,受体可分为细胞膜受体和细胞内受体。
细胞内受体包括细胞质受体和核内受体,由于大多数细胞质受体在活化之后会转移到细胞核,因此,我们通常把细胞内受体统称为核受体。
还有少数非核受体的细胞质受体,如一氧化氮(NO)受体。
细胞内受体接收一些亲脂性信号分子,这些信号分子可直接通透细胞膜,无需特别的跨膜信号转导机制,就可到细胞内找到并结合相应的胞内受体,传递信号,引起细胞效应。
下面我们就着重讲一下核受体介导的信号通路和NO介导的信号通路。
一、核受体信号通路人细胞中已发现48个核受体。
核受体的配体包括雌激素和孕激素等类固醇激素,以及视黄醇和甲状腺素等脂溶性信号分子。
核受体可区分为I型核受体和II型核受体,它们介导的信号通路有一定差异。
1、I型核受体信号通路。
在这一信号通路中,I型核受体通常在胞质中与分子伴侣热休克蛋白90(简称HSP90)结合处于失活状态。
当雄激素、雌激素和孕激素等激素进入细胞,与受体结合,引起HSP90与受体解离,受体分子将形成二聚体,暴露核定位信号,即可迁移到细胞核内。
入核后,激素-受体复合物借助DNA结合结构域(DBD),与DNA中的激素响应元件(HRE)发生特异性结合,并招募转录共激活因子,激活目的基因的表达。
2、II型核受体信号通路。
在这一信号通路中,II型核受体已经定位在核内,并与DNA 中的特定HRE结合,只不过还结合有转录共抑制因子,使之处于失活状态。
当视黄醇、维甲酸、维生素D和甲状腺素等脂溶性配体扩散入核与受体结合形成复合体后,将招募转录共激活因子替换转录共抑制因子,从而激活目的基因的表达。
类固醇激素诱导的基因活化通常分为两个阶段:1)快速的初级反应阶段,直接激活少数特定基因转录;2)延迟的次级反应阶段,初级反应阶段的基因表达产物再激活更多基因表达,是对初级反应的持续放大。
核受体信号通路在细胞的增殖、分化等生命活动中发挥着重要调控作用。
《基础医学各论3》资料:神经元的信号传递

影响突触传递的因素
• 影响突触前末梢释放递质的因素 * 递质释放量与进入末梢内的Ca2+成正相关 [Ca2+]o↑或[Mg2+]o ↓ → 递质释放量↑ 到达末梢的AP频率和幅度↑→Ca2+内流↑→递质释放↑ * 突触前受体 (presynaptic receptor) 的调制作用 * 影响突触囊泡着位 (docking) 的有关蛋白
5. 环状 (recurrent circuit) 联系 可构成正、负反馈 及时终止或产生后放
中枢兴奋传播的特征
1. 单向传播 (one-way conduction) 意义在于限定信息沿指定路线运行
2. 中枢延搁 (central delay) 3. 总和 (summation) 与阻塞 (occlusion) 4. 兴奋节律的改变 (change of excitatory rhythm) 5. 后发放 (after discharge) 和反馈 (feedback)
• 经典突触的微细结构
* 由突触前膜、突触间隙、突触后膜组成 * 突触前膜内结构
微丝、微管、线粒体 突触囊泡(三类)
(synaptic vesicles) 活化区 (active zone) * 突触间隙 (cleft) 20~40nm宽 * 突触后膜上结构 受体 (receptors) 通道 (channels)
传导方向
双向
单向
时间延搁
无
有
电位变化
全或无
总和、节律改变
后发放
无
有
完整性
要求
要求
疲劳
相对不易
相对容易
信号通路讲解

信号通路讲解信号通路(signal pathway)是指当细胞里要发生某种反应时,信号从细胞外到细胞内传递的一种信息,细胞要根据这种信息来做出反应的现象。
这些细胞外的分子信号(称为配体,ligand)包括激素、生长因子、细胞因子、神经递质以及其它小分子化合物等。
信号通路实际上是一种酶促反应通路,通过它,细胞外的分子信号可以经过细胞膜传入细胞内并发挥效应。
在这个过程中,配体特异性地结合到细胞膜或细胞内的受体(receptor)上,然后这些受体将信号传递到细胞内。
在细胞内,信号通过一系列的传递和放大过程,最终引起细胞内部的某些变化,如基因表达的改变、细胞代谢的改变等。
信号通路主要包括受体、蛋白激酶、转录因子等成员。
受体是信号通路的起点,能够识别并结合配体,从而启动信号传递过程。
蛋白激酶则是一类磷酸转移酶,通过将ATP的磷酸基转移到特定的蛋白上,从而改变这些蛋白的构象和活性,进一步传递信号。
转录因子则是对基因转录有调节作用的蛋白,能够识别和结合DNA,从而调控基因的表达。
信号通路在细胞的生命活动中起着至关重要的作用。
它们能够响应外界的刺激,将信号传递到细胞内,引起细胞内部的相应变化,从而实现对细胞功能的调控。
同时,信号通路也是药物研发的重要靶点,通过对信号通路的调控,可以实现对疾病的治疗和预防。
常见的信号通路包括Notch信号通路、AMPK信号通路等。
Notch信号通路是一种通过膜蛋白作为配体和受体,介导两个细胞相互靠近接触之后的活化效应的信号通路。
而AMPK信号通路则是一种能够响应细胞压力、能量摄取改变等刺激的信号通路,通过激活AMPK上游激酶来传递信号。
总的来说,信号通路是一种复杂的细胞内信号传递系统,通过它,细胞能够感知外界环境的变化,并做出相应的反应,从而维持细胞的正常生理功能。
八大类细胞信号通路

八大类细胞信号通路八大类细胞信号通路是指细胞内外因子通过特定的信号传递机制,调控细胞内各种生物学过程的一种细胞信号通路。
这八大类细胞信号通路包括:细胞外基质信号通路、离子通道信号通路、G蛋白偶联受体信号通路、酪氨酸激酶受体信号通路、细胞内钙信号通路、细胞周期调控通路、细胞凋亡信号通路和细胞核转录因子信号通路。
一、细胞外基质信号通路细胞外基质信号通路是指细胞外基质分子通过与细胞表面受体结合,激活细胞内信号传导分子,最终调控细胞增殖、迁移和分化等生物学过程的通路。
其中,整合素受体信号通路是最重要的一类细胞外基质信号通路,它通过整合素受体激活下游信号分子,参与细胞间相互作用和细胞与基质之间的相互作用。
二、离子通道信号通路离子通道信号通路是指离子通道蛋白介导的离子流动通过调节细胞膜电位和细胞内离子浓度,从而影响细胞的生理功能的通路。
钠通道信号通路、钾通道信号通路和钙离子通道信号通路是离子通道信号通路的三个主要类型。
其中,钠通道信号通路参与了神经传导、心肌收缩等生理过程,钾通道信号通路参与了细胞膜电位的调节,钙离子通道信号通路参与了细胞内钙离子浓度的调节。
三、G蛋白偶联受体信号通路G蛋白偶联受体信号通路是指G蛋白偶联受体激活下游信号分子,最终调控细胞内多种生物学过程的通路。
G蛋白偶联受体通常包括G蛋白偶联受体本身、G蛋白和效应器等组成。
这一信号通路参与了多种细胞功能的调节,如细胞分化、细胞增殖和细胞凋亡等。
四、酪氨酸激酶受体信号通路酪氨酸激酶受体信号通路是指酪氨酸激酶受体激活下游信号分子,最终调控细胞内多种生物学过程的通路。
酪氨酸激酶受体包括单个膜通道的酪氨酸激酶受体和多个膜通道的酪氨酸激酶受体两类。
这一信号通路参与了细胞的增殖、分化和凋亡等生物学过程。
五、细胞内钙信号通路细胞内钙信号通路是指细胞内钙离子浓度的变化通过调控钙结合蛋白和钙离子通道等组分,最终调控细胞内多种生物学过程的通路。
细胞内钙信号通路参与了细胞的分化、增殖、凋亡和细胞骨架的重组等生物学过程。
生长因子受体介导的常见信号通路

生长因子受体介导的常见信号通路1. EGFR/EGFR通路:EGFR(表皮生长因子受体)通路是一种重要的信号传导通路,它能够控制细胞生长、分化、移动和增殖等。
EGFR通路的激活是通过表皮生长因子(EGF)与EGFR结合,而EGF的表达受多种因素的影响,包括位点修饰、转录因子介导的调控等。
聚合的EGFR与多种特定蛋白结合,形成跨膜复合体,从而刺激通路中下游信号分子的活化,影响最终细胞行为。
2. MAPK/ERK通路:MAPK/ERK通路是一种常见的信号通路,它可以调节细胞内多种生物过程,如细胞呼吸、增殖、迁移和凋亡等。
MAPK/ERK通路的激活取决于多种因素,其中介导信号从上游接收到下游传导到细胞核的主要元素是Raf/MEK/ERK三重复合蛋白。
Raf来自激活EGFR受体的EGF-MAPK途径,而MEK和ERK则是传导信号至细胞核的重要流程。
3. PI3K/AKT通路:PI3K/AKT信号通路是一种重要的生物过程,它以PI3K/AKT介导信号从上游接收到核内传导信号的形式参与调控细胞的生长、分化和凋亡等事件。
PI3K/AKT效应的激活与IGF-1受体等上游元件的结合有关,PI3K将激活的磷酸残基转移到质子素AKT蛋白上,以此来增强上游信号的传导。
AKT的活性可调控内含子的转录活性,从而调节有效生长、血管形成和细胞抗凋亡等。
4. JAK/STAT通路:JAK/STAT信号通路是一种重要的细胞信号传导机制,它以Janus激酶(JAK)/转录因子STAT(介导信号介质转录因子)介导信号从上游接收到核内传导信号的形式参与调控细胞活动。
JAK受体在存在引发活性时会迅速介导 Janus 激酶启动介导信号介质转录因子(STAT)的活性,STAT可以调控细胞内的许多机制,JAK/STAT信号通路可以非常有效地通过调节细胞形态、增殖、凋亡和角质形成来调节细胞行为。
5. Wnt/β-catenin通路:Wnt/β-catenin信号通路是一种重要的细胞信号传导机制,它可以参与调控细胞增殖、分化、迁移和细胞死亡等,以此调节正常的细胞生长。
细胞的跨膜信号转导功能

跨膜信号转导主要涉及到:胞外信号的识别与结合、信号转导、胞内效应等三 个环节。
跨膜信号转导方式大体有以下三类: ① 离子通道介导的信号转导 ② G蛋白偶联受体介导的信号转导 ③ 酶偶联受体介导的信号转导
一、离子通道介导的信号转导
离子通道大体有:化学、电压、机械性门控通道 如:
化学性胞外信号(ACh)
ACh + 受体=复合体
终板膜变构=离子通道开放
Na+内流
终板膜电位
骨骼肌收缩
二、G蛋白偶联受体介导的信号转导 (一) cAMP信号通路
神经递质、激素等(第一信使)
结合G蛋白偶联受体
激活G蛋白
兴奋性G蛋白(GS)
激活腺苷酸环化酶(AC的蛋白激酶A
细胞内生物效应
(二) 磷脂酰肌醇信号通路
激素(第一信使)
结合G蛋白偶联受体
激活G蛋白
兴奋性G蛋白(GS)
激活磷脂酶C(PLC)
(第二信使)
PIP2
IP3 和 DG
内质网 释放Ca2+
激活 蛋白激酶
C
细胞内生物效应
三、酶偶联受体介 导的信号转导
受体本身具有酶的活性,又称受 体酪氨酸激酶。
生物中ach总结

生物中ACH总结ACH (acetylcholine) 是一种重要的生物分子在生物体内起着关键的作用。
它是一种神经递质化学物质,被发现存在于多种生物中,包括动物和植物。
ACH 在神经传递过程中发挥着重要的作用,影响了广泛的生理过程。
本文将对生物中ACH 的功能和重要性进行总结。
ACH在神经传递中的重要作用1.神经冲动传导:ACH在神经突触之间充当一个重要的信号分子。
当神经冲动到达突触前的终末分支时,ACH被释放到突触间隙中。
它和细胞膜上的ACH受体结合,并启动神经冲动的传导。
这一传导过程有助于在神经系统内传递信息。
2.运动控制:ACH也在运动控制中发挥着重要作用。
它通过神经肌肉接头传递神经冲动到肌肉细胞,从而控制肌肉运动。
ACH的释放和结合与肌肉收缩和松弛紧密相关。
3.认知和记忆:在大脑中,ACH在认知和记忆方面发挥着重要作用。
低水平的ACH与认知功能下降和记忆问题有关。
研究表明,ACH在大脑内部的释放和活性调节对于学习和记忆的形成和巩固至关重要。
ACH的疾病和药物作用1.肌无力:肌无力是一种影响肌肉控制的疾病,与ACH的缺乏或受损有关。
这种疾病中免疫系统攻击了产生ACH的细胞,导致ACH释放减少。
治疗肌无力的方法之一是使用乙酰胆碱酯酶抑制剂来增加ACH的浓度。
2.阿尔茨海默病:阿尔茨海默病是一种神经退行性疾病,与ACH的降解和减少有关。
这种疾病中,大脑中的ACH水平下降。
药物治疗通过抑制乙酰胆碱酯酶的作用,从而延长ACH在神经突触的活性,并改善症状。
3.药物作用:一些药物可以通过影响ACH的活性来产生作用。
例如,抗胆碱药物抑制ACH受体,用于治疗胃肠道疾病和呼吸道问题。
其他药物可以通过增加ACH的释放或增强ACH受体的敏感性来增加神经传递效果。
ACH和植物中的作用除了在动物中的重要功能外,ACH在植物中也起着一定的作用。
1.生长和发育:研究发现,ACH在植物的生长和发育过程中起着重要的调节作用。
免疫应答的细胞信号转导通路

免疫应答的细胞信号转导通路免疫应答是机体的重要保护机制之一,它涉及细胞信号转导通路的复杂过程。
细胞信号转导通路从外部信号到内部物质的影响,涉及多种信号通路和分子。
本文将介绍与免疫应答相关的细胞信号转导通路,包括T细胞受体、B细胞受体、Toll样受体、细胞因子受体等。
1. T细胞受体信号通路T细胞受体是T细胞表面上的一种重要受体,它在免疫应答中扮演着重要的角色。
T细胞受体的激活将启动细胞信号转导通路,从而引发T细胞的免疫应答。
T细胞受体信号通路主要包括四个步骤:酪氨酸激酶(Lck)的激活、ζ链的磷酸化、耦联受体PCKζ的磷酸化和后继的信号转导。
其中,Lck的激活是通过CD4和CD8的协同作用来实现的。
2. B细胞受体信号通路B细胞受体是B淋巴细胞上的重要受体,它能够识别特定的抗原,并启动相应的免疫应答。
B细胞受体的激活将启动细胞信号转导通路,从而引发B细胞的免疫应答。
B细胞受体信号通路主要包括四个步骤:B细胞受体抗原内化、蛋白酪氨酸激酶的激活、血小板衍生生长因子β的激活、后继的信号转导。
其中,蛋白酪氨酸激酶的激活是由CD19、CD21和CD81三种协同作用来实现的。
3. Toll样受体信号通路Toll样受体是一类免疫系统中的重要受体,它能够识别细菌、病毒等外来入侵物质,并启动相应的免疫应答。
Toll样受体的激活将启动细胞信号转导通路,从而引发免疫应答。
Toll样受体信号通路主要包括四个步骤:Toll样受体联合配体的结合、介导TRAF6的招募、启动下游信号转导、后继的信号转导。
其中,介导TRAF6的招募是通过MyD88、IRAK和TRAF6三种协同作用来实现的。
4. 细胞因子受体信号通路细胞因子是细胞间的一类重要信号传递分子,它们通过细胞因子受体在细胞表面上识别特定的信号,并启动相应的细胞信号转导通路。
细胞因子受体信号通路主要包括四个步骤:细胞因子与受体的结合、启动下游信号转导、后继的信号转导。
其中,启动下游信号转导的过程中,JAK家族的激酶和STAT家族的转录因子是非常重要的分子。
关于AChE,信号途径,ALDH与病理性造血的进展

关于AChE,信号途径,ALDH与病理性造血的进展前言白血病(Leukaemia)是一种造血组织的恶性肿瘤,其特征为体内各组织和器官白细胞浸润,白血病细胞有质和量的异常。
临床上出现不同程度的贫血、出血、发热及肝、脾、淋巴结肿大。
在对白血病的研究方面,除了临床实践之外,基础研究在当今显得更为突出。
从各种途径入手,寻找相关因子与致病作用的关联。
本篇介绍关于乙酰胆碱与C-终端连接蛋白作用于T细胞减少症;NOTCH路径调节血细胞凋亡以及ALDH的致病因素方面的内容以及相关的最新进展。
AChE/C-终端连接蛋白相互作用修改Ikaros功能,引起T淋巴细胞减少症血液的改变,因多种压力刺激引起,伴随初级的AChE 3'接合AChE-S和AChE-R.为寻找确定的AChE-S交互作用于造血的路径,我们使用了一个酵母双混筛.信使核酸形成的辅抑制物C-核绑定蛋白(CtBP)被确定为AChE-S C终点的蛋白助手.在白血病的K562细胞中,AChE-S显示了细胞核定位和与CtBP的物理交互作用.此外,辅助转染AChE-S减少了CtBP对造血转录因子(Ikaros)的辅助压抑效应.在转基因小鼠中,过度表达的人类AChE-S mRNA引起了Ikaros选择性的骨髓上调,同时抑制FOG,另一个CtBP的信使核酸形成助手.转基因骨髓细胞表现了一个为生成起源群体而相对提高的电位,在调控下,当外周血表现出红血球计数增长,而相反的血小板,粒细胞和T淋巴细胞则降低.AChE的3'端选择性接合,在AChE-S/CtBP交互作用中通讯改变,从而在造血的调控和免疫功能电位调节中活动积极,支持关于免疫故障在削弱结合点选择之下的报告.哺乳动物的造血作用包括细胞增殖,血型选择和成熟,有些包括切除结合选择,改变了血细胞起源中的多样性前mRNAs的产物mRNA.这使得复合物能够被需要去保证造血和对改变的生物环境快速反应.结果的更改在选择性接合的环节决定了绑定蛋白,细胞内定位,生物化学活动和许多蛋白的稳定性.一个在外周刺激反应中能选择性结合的造血活性基因的例子,就是AChE,负责水解ACh.主要的AChE mRNA抄本连接外显子(E)4到E6,引起AChE-R,"突触的"对碘氧基苯甲醚和螺旋结构的两性分子C终点的生成. 另一个副本,AChE-R,表达于胚胎或者肿瘤细胞,由连续的信使核酸在基因内区I4形成.多重压力刺激下,AChE-R和AChE-S的比率在脑和血细胞中都相似的增加.AChE-R,通过他的C-终端缩氨酸ARP,与细胞内的蛋白RACK1和他的蛋白激酶C(PKC)助手结合. 在造血细胞中,这能诱导骨髓组织生成. 同时,用转基因小鼠实验,明确的得出了AChE-R提高了淋巴细胞的增殖和血小板计数. 然而,仍然遗留了细胞核是否有能力在压力的干扰下自行获取AChE-R,并足够去诱导骨髓组织生成和血小板增多的问题,或者是否AChE-R的功能丢失也间接涉及.在培养的巨幼细胞中,AChE存在于核上,并且在巨核细胞生成过程中受到抑制.此外,AChE的基因表达在各种类型的细胞的凋亡过程中增多,并且以微小的干涉来抑制AChE基因可以阻止细胞凋亡的传播. 为寻找参与这些现象的精确的AChE变量,我们使用了ASP67,一种高度演化的保存缩氨酸,来自人类的AChE-S终端,作为A YEAST TWO-HYDBRID SCREEN的诱导剂.为证明结合点选择过程,这个过程决定ACHE-S/ACHE-R的比率,对普通造血的影响,我们使用了TgS转基因小鼠来证明细胞核定位了AChE-S变体. 我们确定了关键的辅助抑制物C-终端绑定蛋白(CtBP)为AChE-S的助手,CtBP与AChE-S之间的作用是一项影响了CtBP对造血转录因子Ikaros协助压抑作用的交互作用. 相应的,TgS小鼠的骨髓被发现可以成为高级别的Ikaros的解释,伴随着调控,它们的外周血表现出有联系的红细胞增多,血小板减少和T淋巴细胞减少症. 这些观察结果证明,AChE-S作为一个多系血液因子,通过CtBP,调节多种造血细胞转录因子的转录活动.AchE长久以来都被怀疑在普通和癌基因造血中扮演重要角色。
Ach受体信号通路
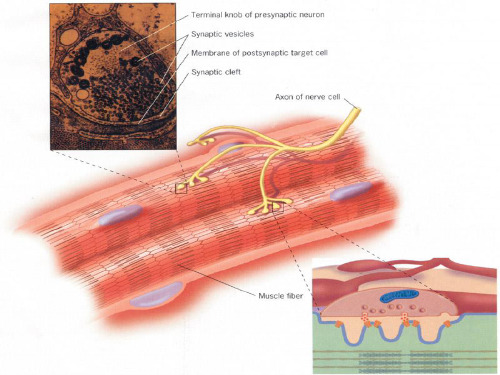
受体
parasympathetic nerve 副交感神经 synapses n.突触 atropine n.阿托品
postsynaptic neurons n.突触后神经元 bungarotoxin n.金环蛇毒素 venom n.蛇毒 homogenize vt.使均质 blender n. 搅拌机 centrifuging n.离心过滤 pellet n. 小球 nonionic detergent非离子型洗涤剂 hydrophobic adj.疏水性的 hydrophilic adj.亲水的, 吸水的
目前公认的导致AChR在突触后膜高 密度聚集途径
Agrin/ MuSK信号转导通路 Neuregulin/ErbB信号转导通路
Agrin/ MuSK信号转导通路
集聚蛋白(Agrin)主要由运动神经元合成, 经神经末梢释放并沉积于基底层,触发 AChR聚集,而不增加AChR的数量。
肌肉的受体酪氨酸激酶(muscle-specific receptor tyrosine kinase, MuSK)是Agrin的 受体,选择性在骨骼肌表达,与AChR共同 聚集于成年神经肌肉接头的突触后膜。
受Agrin的影响,MuSK被快速磷酸化,介 导神经肌肉接头nAChR的磷酸化。
此过程还必须依赖于突触后膜ACh受体缔合 蛋白(acetylcholine receptor-associated protein at synapse,Rapsyn)的存在。 Rapsyn主要在肌肉组织合成,是一种细胞 骨架蛋白。它在体内与神经肌肉接头 nAChR以等摩尔数存在,共同定位 。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
in the vicinity of adv.在邻近 phosphorylation n.磷酸化 kinase n.激酶 polymerization n.聚合 sepharose beads n.琼脂糖凝胶珠 affinity column n.亲和层析柱 flaxedil n.三乙碘化三苯 elute vt.洗脱
将准备好的富含nAChR的细胞膜涂抹在一 个支持网络上,并使它能够流到装有能使 细胞膜冻结的液氮乙烷的容器中。在 nAChR溶液到达冷冻池前约5毫秒(msec) 时,向网格喷射ACh溶液,ACh就与受体结 合并导致其构象变化而打开通道。
通过比较nAChR通道 开放与关闭的电镜照 片,Unwin发现ACh 的结合触发了受体亚 基细胞膜外结构域的 构象发生了改变,并 且传播到整个受体蛋 白,导致围绕在孔道 周围的α-helix构象改 变。(Figure6)孔道 的封闭式由于α-helix 处结的形成,这一点 可从Figure6b的白、 蓝条比较看出。
acetylcholine (ACh) n.乙酰胆碱 ray n.鳐 potential prey 潜在的食物 acetylcholinesterase n.乙酰胆碱酯酶 motor nerves n.运动神经 acetylcholine receptor (nAChR) n.乙酰胆碱
1980s U.J. McMahan:agrin诱导聚 集nAChR的机制
在19世纪80年代,斯坦福大学的U.J. McMahan和他的同事们在EO中发现了一种 蛋白,当把这种蛋细胞膜的聚 集。并且还发现它在EO的突触间隙处有很高 的浓度。
cation n.阳离子 postulate n.假定 conformational n.构象的 inward flux 内流 depolarization n.去极化 sodium n.钠 potassium n.钾 mutagenesis n.诱变 neurotransmitter n.神经递质
目前公认的导致AChR在突触后膜高 密度聚集途径
Agrin/ MuSK信号转导通路 Neuregulin/ErbB信号转导通路
Agrin/ MuSK信号转导通路
集聚蛋白(Agrin)主要由运动神经元合成, 经神经末梢释放并沉积于基底层,触发 AChR聚集,而不增加AChR的数量。
肌肉的受体酪氨酸激酶(muscle-specific receptor tyrosine kinase, MuSK)是Agrin的 受体,选择性在骨骼肌表达,与AChR共同 聚集于成年神经肌肉接头的突触后膜。
信号的传导路径由于Tyr的磷酸化而被激活, 并常会导致细胞骨架元件的重排,特别是 肌动蛋白纤维。细胞骨架经常是连在肌动 蛋白骨架(AS)上的,AS在整合膜蛋白的 分配上扮演着一个关键角色。
最近的研究显示,细胞表面信号的产生是 在agrin和它的受体结合导致AF去极化之后, 这被认为是nAChR聚集的直接原因。如果 AF的形成被能与actin单体结合并阻止单体 间联合形成多聚体的药物阻断,agrin诱导 nAChRs的聚集就会被阻止。
上述3种分子密切相连,神经轴突分泌Agrin 后,在Rapsyn的参与下MuSK被磷酸化, 使nAChR磷酸化,由此诱导AChR在终板膜 聚集。
Neuregulin/ErbB信号转导通路
神经调节蛋白Neuregulin(NRG)由神经元 产生,自突触前膜释放入突触间隙并固定在 神经肌肉接头。
受Agrin的影响,MuSK被快速磷酸化,介 导神经肌肉接头nAChR的磷酸化。
此过程还必须依赖于突触后膜ACh受体缔合 蛋白(acetylcholine receptor-associated protein at synapse,Rapsyn)的存在。 Rapsyn主要在肌肉组织合成,是一种细胞 骨架蛋白。它在体内与神经肌肉接头 nAChR以等摩尔数存在,共同定位 。
Agrin诱导聚集nAChR的机制,对我们来说,仍然 知之甚少,但这一过程中的许多步骤已被界定。 将agrin加入到未成熟的肌细胞后,发生的事件之 一是nAChR β亚基的一个或多个酪氨酸(Tyr)残 基的磷酸化。
这个磷酸化作用发生在出现nAChR聚集的迹象之 前很长一段时间。
Tyr的磷酸化在细胞膜信号传导的过程中常是一个 关键步骤。如果nAChR的Tyr磷酸化被加入的Tyr 激酶失活物阻断,nAChR将不再聚集,这表明 agrin诱导nAChR的Tyr磷酸化在聚集作用中是一 个不可缺少的步骤。
ethane n.乙烷 spray vt.喷射 grid n. 格子, 栅格 peripheral adj.外围的 agrin n.集聚蛋白,集聚素[由运动神经分泌
并可诱 导肌纤维的乙酰胆碱酯酶和乙酰 胆碱受体发生聚集] axon n.轴突 a cascade of events n.级联放大反应
专业词汇
pharmacist n.配药者, 药剂师 elucidate vt. 阐明, 说明 curare n.箭毒马鞍子,箭毒(箭毒马鞍子的毒
素,南美印地安人用以浸制毒箭) toxic adj.有毒的, 中毒的 antagonistic adj.颉颃的 vagus n.迷走神经 Vagusstoff n.迷走神经释放物
孔道的门控开关被认为是在跨膜
通道的中央附近,在这里,5个
α-helix屈向中心,形成一个结。
在这个模型中,
5个α-helix的
Leu(亮氨酸)
残基组成了一个紧缩环,限制
离子的跨膜流动。当ACh分子
与两个α亚基膜外结构域口袋
结构的一个位点结合后,门控
开关被打开。
为了研究通道开放过程中nAChR的变化, Unwin做了下面的实验。
nAChR更精确的模型已经被英格兰医学研 究所的Nigel Unwin和他的同事们所描绘。 经过对EO冷冻膜电镜照片的数学分析, Unwin将nAChR的组成描述为5个亚基围绕 着一个中央通道。(Figure5)
离子通道是一个很窄的直径只有
7~8Å的孔道,孔道被5个亚基
的α-helix片断组成的墙所包围。
受体
parasympathetic nerve 副交感神经 synapses n.突触 atropine n.阿托品
postsynaptic neurons n.突触后神经元 bungarotoxin n.金环蛇毒素 venom n.蛇毒 homogenize vt.使均质 blender n. 搅拌机 centrifuging n.离心过滤 pellet n. 小球 nonionic detergent非离子型洗涤剂 hydrophobic adj.疏水性的 hydrophilic adj.亲水的, 吸水的
1987年,他们纯化了这种蛋白,并将它命名 为集聚蛋白(agrin)。
McMahan提出agrin是由运动神经合成,延 轴突运输到终端神经节,并在此被释放到 神经肌肉接头处的细胞外空间。这些agrin 分子而后与邻近肌细胞质膜上的特异性 agrin受体结合,触发一系列级联放大反应, 使nAChR分子(以及突触后膜的其他组成 部分)聚集到活化的agrin受体附近,并达 到最大浓度。
phospholipid n.磷脂 disruptive adj.使破裂的, 分裂性的 carboxyl n.羧基 resemblance n.相似之处 affinity chromatography n.亲合色谱法(层
析)
pentamer n.五聚物 electrophoresis n.电泳, 现象 polyacrylamide gel n.聚丙烯酰胺凝胶
其他多肽如降钙素基因相关肽(calcitonin gen-related peptide)作为一种神经肽,存在 于脊椎动物的运动神经元,推测与突触后 受体的积聚有关。
更深入的关于神经肌肉接头(NMJ)形成 的问题,由一项对鼠的agrin基因敲除实验 解决。即通过遗传工程的方法使鼠的运动 神经不能产生agrin。Agrin缺陷鼠在母体中 发育正常,但在妊娠期却死亡。
这些鼠在死前没有任何肌肉运动的迹象出 现,并且在肌肉组织的检查中,发现缺少 正常的神经肌肉突触。
Agrin作为突触间隙细胞外物质的一部分, 帮助维持后突触膜的结构,并保证新合成 的nAChRs在突触位点的富集。
多种研究都支持这一假说,包括利用抗体 与agrin结合的特异性所进行的研究。
如果使鸡胚胎脊髓的一段,携带抗agrin的 抗体,此抗体就会特异性地标记运动神经 的细胞质,这表明agrin分子来源于此。抗 体当然也存在于神经肌肉接头处的突触间 隙。若将此抗体加入到鸡的运动神经和正 在生长的肌细胞的混合培养物中,所培养 的两类细胞接触位点处nAChR的正常聚集 将会被阻断。
纯合子缺乏NRG的小鼠由于心脏发育的缺陷 而在神经肌肉接头形成前夭折,杂合子缺失 NRG的小鼠显示在神经肌肉接头的AChR减 少50%,神经肌肉间神经递质传递也存在缺 陷。可见NRG具有AChR诱导活性,在发育 和成熟肌肉细胞中促使AChR基因表达。
ErbB受体在神经肌肉接头突触后膜聚集, 与NRG结合后,激活ErbB受体酪氨酸激酶, 经过接头Shc和Grb2将信号传递给下游 Ras/Raf/MEK/ERK激酶信号传导途径。导 致突触下细胞核特异性转录,以保持AChR 在成熟肌肉接头的高密度。
疑问
我们知道膜外周的部件可限制膜整合蛋白的 移动。在nAChR上也发现了一个典型的例子。 在胚胎早期,肌肉无神经支配时,nAChR均 匀分布在肌肉表面。而成熟神经肌肉接头处 nAChR则高密度聚集。
运动神经末梢下的突触区小于肌肉细胞表面 积的0.1﹪,却集中了约90﹪的nAChR。这是 为什么呢?