广西部分棘蛙类物种的分子系统学研究
【国家自然科学基金】_系统发育地位_基金支持热词逐年推荐_【万方软件创新助手】_20140803
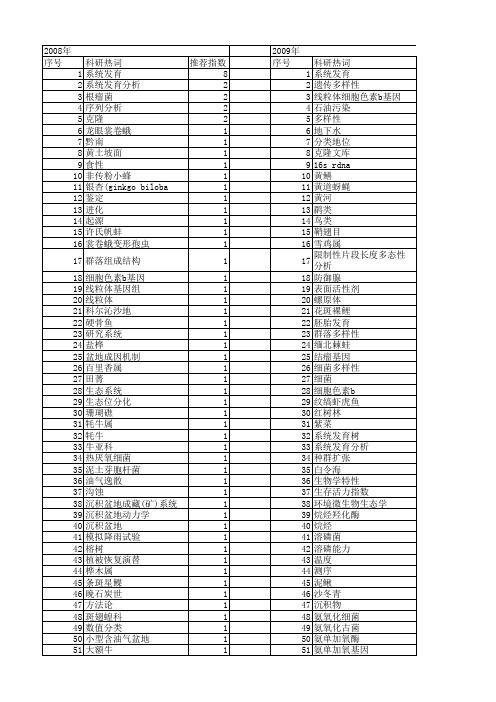
科研热词 系统发育 遗传多样性 线粒体细胞色素b基因 鹛类 鸟类 鞘翅目 雪鸡属 限制性片段长度多态性分析 防御腺 表面活性剂 螺原体 花斑裸鲤 胚胎发育 群落多样性 缅北棘蛙 结瘤基因 细菌多样性 细菌 细胞色素b 纹缟虾虎鱼 红树林 紫菜 系统发育树 系统发育分析 种群扩张 白令海 生物学特性 生存活力指数 环境微生物生态学 烷烃羟化酶 烷烃 溶磷菌 溶磷能力 温度 测序 泥鳅 沙冬青 沉积物 氨氧化细菌 氨氧化古菌 氨单加氧酶 氨单加氧基因 根瘤菌
柴达木裸裂尻鱼 杂交水稻种子 放线菌群落 拟步甲科 戈登氏菌 形态特征 序列分析 察隅棘蛙 大西洋 多系发生 固有细菌 叶状体 叶杆菌 刺甲族 分类 分子系统学 分子生态学 免培养方法 光照周期 仔鱼发育 于田盐池 三叶草 sphaerophysa salsula rrna基因 rflp rbcl基因 ph dna条形码 dna分类 18s 16srdna 16s rrna 16s rdna全序列 16s rdna pcr-rflp 12s rrna
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
动物学(下)试题

动物学(下)试题棘皮动物门一、填空题1. __________________________________ 棘皮动物特有的结构是—水管系統___________________________________ 和—管足_______ ,是次生体腔的一部分特化形成的管道。
二、名词解释1. 辐射对称:通过身体中轴,可以有许多个切面把身体分为两个相等的部分,具这样体型的动物适于漂流生活。
三、选择题1. 被囊动物指的是(C )。
A.柱头虫B.海鞘C.海盘车D.海胆四、判断题1. 棘皮动物体腔的一部分特化成“血窦”。
()五、简述题1. 棘皮动物门为什么是最高等的无脊椎动物?有哪些主要特征?半索动物门一、选择题1. 半索动物与下列哪些动物亲缘关系最近(D.棘皮动物名词解释A.头索动物B.尾索动物C.脊索动物1. 后口动物:在胚胎发育中原肠胚的胚孔形成肛门或封闭而另外形成口的动物。
2. 原口动物:在胚胎发育中由原肠胚的胚孔形成口的动物简答题1. 半索动物和什么动物亲缘关系最近?为什么?答:半索动物和棘皮动物的亲缘关系最近,它们可能是由一类共同的原始祖先分支进化而成。
理由是:①半索动物和棘皮动物都是后口动物。
②两者的中胚层都是由原肠凸出形成。
③柱头虫的幼体(柱头幼虫)与棘皮动物的幼体(例如短腕幼虫)形态结构非常相似。
④脊索动物肌肉中的磷肌酸含有肌酸,无脊椎动物肌肉中的磷肌酸含有精氨酸,而棘皮动物的海胆和半索动物的柱头虫肌肉中都同时含有肌酸和精氨酸。
这些证据都说明了这两类动物有较近的亲缘关系。
脊索动物门一、填空题1. 文昌鱼由于终生具有发达的_脊索___________ 、___神经管_________ 和___咽鳃裂 ___ 等脊索动物的重要特征,而被认为是典型脊索动物的简化缩影。
2. 脊索动物门的三大主要特征是_脊索___________ 、背神经管和咽鳃裂,另外还具有一些次要特征,如心脏_____________ 位,具___肛后尾。
广西师范大学学报:自然科学版 第26卷(2008年)目次

第 1 期
量 子相 干 网络 ………………………………………… …………………… ………………・ 毕 桥, 柳贵平 , 永 李
推 广 小世界 网络上 的 S n j zad舆论 模型
… … … … … … … … … … … … … … … … 一王 茹, 勖 蔡
供 应 链管 理 中基 于蚁 群 算法 的 多 Agn 合 作 … …… … … … … … … … … ………….赵 翔, et . 黄厚宽
基 于 Mut Ag n 的空 中交 通 协 同流量 管理 … ……………………………………一 王 飞, 肖 , 静 l— e t i 徐 豪 张
S R一 凝法 处 理奶 牛 场废水 C B 絮 ODc………… ……………………………………. 张 翔, . 万亚珍, 刘金盾, 等
扶 绥 黑 叶猴食 物 多样性 的初步 研 究 …………………………………………… ….李友邦, . 黄乘明, 黄中豪, 等
北 部湾 秋 、 季渔 业 生物 群落 结构 特征 的 变化 ………………… …………………一 乔延龙, 冬 林昭进, 邱永松 广 西 部分 棘蛙 类物 种 的分 子 系统 学研 究 ……… ……………………………………“ 高成伟, 张碧文, 秦新民
[ Mn H2 2C C O)( H。 2 H2 ・ H3 的合 成与 晶体 结构 Mn ( L)( H。O 4C OH)]・ O C OH
… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … … …
蒋春芳 , 黎
燕 , 福 沛 梁
基于分子学学角度探讨无尾目分类

XXXX大学作业题目:线粒体基因探讨无尾目的分类研究进展课程名称:动物分类学任课教师姓名:XXXXX研究生姓名:XXXXX学号:XXXXXXXX年级:2010级专业:动物学学院(部、所):生命科学学院任课教师评分:年月日线粒体基因探讨无尾目的分类研究进展摘要:运用PCR技术和相关生物软件,对大头蛙Limnonectes kuhlii和脆皮大头蛙Limnonectes fragilis的线粒体CO I基因全序列进行了测定与分析.结合GenBank上已公布的无尾目1O个科22个物种的同源序列,以爬行纲楔齿蜥Spkenodon punctatus和密河鳄Alligator m ss厶s 如,2s厶为外群,基于核苷酸序列(选取前两位密码子)和氨基酸序列两种数据形式,分别构建NJ树和MP 树.结果表明:系统树将无尾目1O个科分为两大支,一支为新蛙亚目Neobatrachia 的5科15个物种,一支为古蛙亚目Archaeobatrachia的5个科9个物种(其中包括锄足蟾科Pelobatidae和负子蟾科Pipidae),并推测这两个亚目均为单系群.古蛙亚目一支内的盘舌蟾科Discoglossidae与铃蟾科Bombinatoridae亲缘关系较近;新蛙亚目内蟾蜍科Bufonidae与雨蛙科Hylidae,蛙科Ranidae与树蛙科Rhacophoridae两两亲缘关系较近.蛙科中泽陆蛙Fejervarya limnocharis与大头蛙属3个种首先形成姐妹群,表明其较近的亲缘关系,同时推断大头蛙与福建大头蛙Limnonectes fujianensis亲缘关系近于脆皮大头蛙.关键词:大头蛙;脆皮大头蛙;CO I基因;无尾目;系统关系目前,两栖动物的分类和保护工作一直是国际野生动物保护的热点。
由于人们对两栖类资源的过度利用和捕杀,加之生境破坏,栖息地减少,造成野外种群数量逐渐减少,尤其是两栖类无尾目动物的保护现状更加严峻,例如:棘胸蛙已被列入濒危动物红皮书,而虎纹蛙在野外多年没有报道。
【国家自然科学基金】_16s rrna基因序列_期刊发文热词逐年推荐_20140731
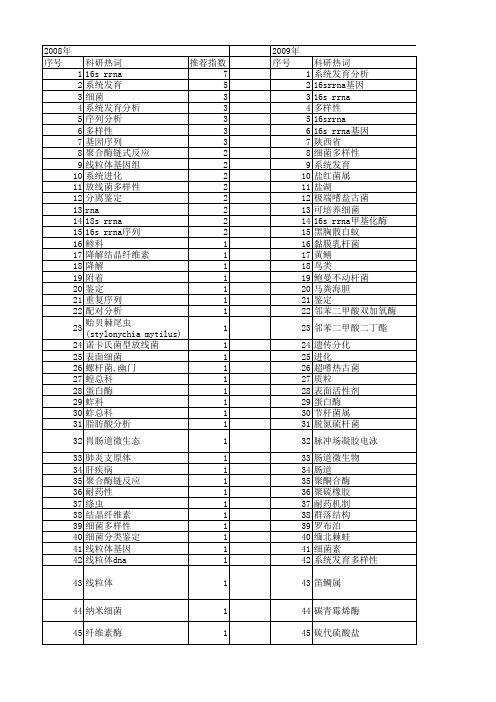
科研热词 系统发育分析 16srrna基因 16s rrna 多样性 16srrna 16s rrna基因 陕西省 细菌多样性 系统发育 盐红菌属 盐湖 极端嗜盐古菌 可培养细菌 16s rrna甲基化酶 黑胸散白蚁 黏膜乳杆菌 黄鳝 鸟类 鲍曼不动杆菌 马粪海胆 鉴定 邻苯二甲酸双加氧酶 邻苯二甲酸二丁酯 遗传分氮硫杆菌 脉冲场凝胶电泳 肠道微生物 肠道 聚酮合酶 聚硫橡胶 耐药机制 群落结构 罗布泊 缅北棘蛙 细菌素 系统发育多样性 笛鲷属 碳青霉烯酶 硫代硫酸盐 盐生硫杆菌属 瘤胃细菌 瘤胃 甲基转移酶类 生物降解 生物氧化 生物学特性
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106
硫化叶菌 1 53 瓯江彩鲤 硝尔库勒湖 1 54 猪链球菌 研究 1 55 焦磷酸测序 直翅目 1 56 烷烃羟化酶 盐矿 1 57 烷烃 病原菌 1 58 深海热液口 病原微生物 1 59 海葵 生物浸出 1 60 海洋混凝土工程 生物安全 1 61 海洋弧菌 猪肠道乳酸杆菌 1 62 浮游细菌 点突变 1 63 活性污泥 演替 1 64 泥鳅 海洋鱼类 1 65 油樟 海洋细菌 1 66 氨基糖苷钝化酶 浮游细菌 1 67 核苷酸序列 泥土芽胞杆菌 1 68 普鲁兰酶 泉古菌,多样 1 70 整合素类 气相色谱 1 71 放线菌群落 橙叶病 1 72 抑制性差减杂交 植原体 1 73 戈登氏菌 梭子蟹 1 74 微生物 核苷酸序列 1 75 形态特征 核糖体蛋白l4 1 76 引物 核糖体蛋白l22 1 77 废水处理 标本 1 78 序列比较 柑橘 1 79 序列变异 极端环境 1 80 序列分析 极端嗜热古菌 1 81 差异表达基因 条斑星鲽 1 82 屎肠球菌 晋南牛 1 83 小型兽类 新疆 1 84 察隅棘蛙 新异名 1 85 大西洋 支原体,肺炎 1 86 外膜蛋白k基因 排污口 1 87 复制 抗药性 1 88 基因克隆 快速鉴定 1 89 嗜盐古菌 微生物群落 1 90 嗜吞噬细胞无形体 微生物敏感性试验 1 91 厌氧生物降解 形态特征 1 92 动性球菌 形态 1 93 利玛原甲藻 小鼠 1 94 分离鉴定 小叶 1 95 分离 宏基因组dna 1 96 分子鉴定 大环内酯类 1 97 分子系统学 大尾柱虫(urostyla grandis) 1 98 分子杂交 塔里木盆地 1 99 内生菌 基因,rrna 1 100 共生古菌 城市污水 1 101 免培养方法 圆斑星鲽 1 102 体色 嗜酸氧化亚铁硫杆菌 1 103 仔猪 嗜盐放线菌 1 104 产芽孢细菌 厦门海域 1 105 五氯酚 厌氧氨氧化菌 1 106 于田盐池
现代生物学实验习题

现代生物学实验习题(一)每个实验按“实验报告纸”的格式完成一份实验报告(二)回答各部分实验各模块中的相关思考题。
思考题汇总:第一部分:生物化学实验第一模块:糖实验技术1、如何选择生化物质实验中的实验材料?有何标准?2、材料的破碎方法有哪些?3、碱性铜试剂法测定还原糖是直接滴定还是间接滴定?两种滴定方法各有何优点?4、在粗纤维的测定中哪些因素是影响测定结果的主要因素?5、在粗纤维的测定中为什么必须严格控制实验条件。
6、用本方法测定的结果为什么称为“粗纤维”?第二模块:酶学实验技术1、酶的特异性实验为什么要做试剂检查实验?省略该步骤可能有怎样的结果?2、酶作用的专一性有哪几种?3、试说明可逆抑制剂与不可逆抑制剂的作用特点。
4、三种可逆抑制剂的抑制程度与底物浓度有何关系?5、高温导致酶活性丧失的分子机理是什么?如何在酶的分离纯化中克服这种影响?6、还有哪些因素可以导致酶活性的降低?7、 pH值导致酶活性丧失的分子机理是什么?8、 pH值改变导致的活性丧失, pH值恢复后活性也能恢复吗?9、试说明米氏常数K m的物理意义和生物学意义。
10、为什么说米氏常数K m是酶的一个特征常数而V m则不是?11、请把已做的酶定性实验改造为定量实验。
第三模块:蛋白质实验技术1、为什么用0.145 mol/L TCA来抽提细胞色素C?2、抽提细胞色素C时加入硫酸铵时要注意什么?为什么?3、在凝胶过滤层析纯化细胞色素C实验中要注意哪些重要环节?4、SDS-聚丙烯酰胺凝胶电泳分离蛋白质为什么要在样品中加少许溴酚兰和一定浓度的蔗糖溶液?蔗糖及溴酚兰的作用分别是什么?5、简述聚丙烯酰胺等电聚焦电泳的基本原理。
6、进行聚丙烯酰胺等电聚焦电泳时的注意事项有哪些?7、在酪蛋白制备时为什么要用醋酸缓冲液调pH到4.8?8、在酪蛋白制备时.乙醚、乙醇作用是什么?9、与其它测定蛋白质浓度的方法相比,考马斯亮蓝染色法测定有何优点?10、如何用纸层析法对氨基酸进行定性和定量的测定?11、纸层析和柱层析、薄层层析、高效液相层析各有什么特点?第四模块:其他生物物质的分析1、粗脂肪的定量测定─索氏提取法实验中得到的是粗脂肪, 若要制备单一组分的脂类成分, 可用什么方法进一步处理?2、粗脂肪的定量测定中样品制备时烘干为什么要避免过热?3、DNA含量测定的方法有哪些?各有何优缺点?4、简述二苯胺法测DNA的基本原理。
2018国自然-生命科学部及医学科学部名单

2018国自然-生命科学部及医学科学部名单三七自毒物质降解菌的特性和作用机制研究黄荣韶DEELA蛋白介导赤霉素调节山药块茎膨大的分子机理王爱勤广西巴马小型猪2型糖尿病易感性相关LncRNA的筛选及机制研究兰干球自我构念影响心理理论加工的ERP研究:基于中国-东盟国家的跨文化比较蒋钦广西长寿人群饮食、肠道菌群和代谢物特征凝练与数据平台初建李全阳卵形鲳鲹SIGIRR通过TLRs-MyD88信号通路对巨噬细胞炎症应答的影响与机制研究韦友传鸡传染性支气管炎病毒多表位嵌合病毒样颗粒的构建及其免疫应答机制研究磨美兰甘蔗尾青贮核心发酵菌群和代谢物演替规律及其对水牛瘤胃细菌多样性的影响邹彩霞基于定量磷酸化蛋白质组学在水牛精子发生过程的分子机制研究张明小分子化合物在水牛原始卵泡体外激活中的作用及其分子机制的研究杨素芳KDM6B和Xist双基因表达调控对牛体细胞核重编程中X 染色体重塑影响机理的研究韦精卫Nanos2维持水牛精原干细胞干性特征机制的研究杨小淦应用基因捕获高通量测序精准鉴定影响水牛产奶性状的关键基因研究刘庆友寄生蜂定位致瘿害虫的化学机制研究——以孟氏胯姬小蜂为例郑霞林过氧化氢与一氧化氮互作对桉树耐铝性的调控作用及其机理滕维超沿海纬度梯度分布红树科植物对温度变化的水力结构和功能响应蒋国凤短氟碳低聚物定向接枝纤维素的合成机理及其构效关系研究朱红祥具有生物活性的3-蒈烯衍生物的合成及光异构研究段文贵心材化过程中木质部薄壁细胞代谢活性与心材成分生成机制符韵林番茄抗细菌性髓部坏死病遗传规律的研究及分子标记筛选王先裕番茄伴生植物的筛选及混栽促生组合番茄根际及内生微生物群落解析杨尚东芒果FT基因在成花过程中的分子机制研究罗聪咪唑啉酮类除草剂在水生微宇宙系统中对映选择性环境行为和毒性效应谭辉华乙烯调控罗汉果性别的细胞学及分子遗传研究周琼甘蔗体细胞融合育种中杂合体细胞再生植株的分子机制李素丽microRNA调控铝诱导花生根尖细胞程序性死亡发生的分子机制何龙飞水稻可塑性基因RPL1调节水稻株高的分子机制研究张翠翠中国原始被子植物瘿螨总科区系与分布格局机制研究王国全珍贵乡土树种与桉树混交修复土壤质量的效应及机理研究温远光Landscape and population genomics of endangered&nb Kim Shan We 共生真菌对Euops属卷叶象甲幼虫发育的营养功能及分子机制研究李晓琼红树植物生物钟预测环境和潮汐变化能力对其光合的贡献及机制研究朱俊杰高巢被捕食压力下北热带石灰岩森林鸟类的繁殖行为与策略蒋爱伍bE2-bW1复合体调控甘蔗黑穗病菌丝状生长的机制陈保善十字花科黑腐病菌小RNA分子伴侣蛋白Hfq的作用机理研究唐东阶水稻条斑病菌III型效应子XopN致病机理研究黄胜一个新的双组份信号转导系统感受子基因抑制III型分泌系统的信号途径研究姜伟十字花科黑腐病菌全局性转录调控因子HpaR1与Clp共同调控致病生化因子表达之模式研陆光涛冯军功能化纳米复合材料hCEs SERS探针的构建及在血清与细胞中检测羧酸酯酶活性的越南槐根内生真菌生态功能及其与宿主药材品质相关性研究姚裕群基于甘蔗生产全程机械化的肥料氮利用与土壤氮迁移研究韦剑锋潘丽霞三联吡啶类金属配合物、靶点DNA与DNA拓扑异构酶结合方式及三元复合物结构基础的研松香基分子印迹膜的结构调控及定向分离三七活性成分三七素的研究黄钦基于表型性状和ISSR标记的苹婆遗传多样性分析及核心种质构建周婧用晶格Boltzmann方法研究功能性微气泡生成机制何冰一种双吡嗪基脲衍生物通过ERK/Slug/vimentin/BCRP信号轴逆转表阿霉素耐药的研究陈家念中国吟螽属修订(直翅目:螽斯科:蛩螽亚科)边迅猫儿山小鲵生活史不同阶段的生境选择研究黄华苑生境破碎化对白头叶猴肠道寄生虫的影响及其行为适应周岐海中亚热带石灰岩常绿落叶阔叶混交林植物功能性状分异及其对环境变化响应的研究姜勇两种青牛胆属植物中抑制小胶质细胞活化成分的发现及其初步作用机制研究廖海兵王任翔边缘鳞盖蕨复合体种 (Microlepia marginata complex) 的网状盐藻类胡萝卜素合成调控相关的转录因子基因的作用分析尚常花蛋白纳米疫苗 VP6-Ferritin 高效免疫的结构基础研究王丹喀斯特地貌生境隔离对洞穴鱼类物种形成和分布格局的作用机制杨剑不同施肥措施下甘蔗田土壤温室气体排放过程机理及其调控因素李卓亭火干扰对大兴安岭森林更新结构和地上生物量动态的影响蔡文华喀斯特不同生境类型树种水力特征及其权衡关系研究刘艳艳岩溶植物根系对水环境变化的形态可塑性响应及其水分再分配的生态效应研究黄玉清淡水刚毛藻目的系统分类学研究赵志娟海洋生物来源天然小分子生物碱抗HIV-1机制研究周波长链非编码RNA LINC00857介导非小细胞肺癌耐药的机制研究王丽惠微环境CAFs源性外泌体调控非小细胞肺癌EGFR-TKI耐药及其机制阳洁靶向耐药肺癌的高选择性共价Bmx抑制剂的设计、合成及其生物活性研究何林洪circRNA-miRNA-p38MAPK通路调控网络对缺血性脑卒中及其中医证候的影响与机制研究苏莉留兰香香蜂草苷对肝纤维化内质网应激--自噬通路的调控作用机制林兴基于NF-κB信号通路探索青蒿琥酯对伴糖尿病牙周炎炎症反应及成骨调控的效应和机制研究农晓琳氯化两面针碱基于miR-125b-2-3p/IER3调控轴抗HCC作用的机制研究党裔武多源信息下基于贝叶斯网络的综合性医院门诊量不确定性分析与预测黄代政miR-132-RB1/E2F1信号通路与肝癌发生发展的分子流行病学研究容敏华我国HIV-1肺储存库的准种遗传多样性和细胞嗜性研究宁传艺黄东萍基于壮族出生队列的孕早期环境雌激素污染物联合暴露与胎儿婴儿发育情况的关系研究TNFR-RIPK1/RIPK3信号通路介导铅致神经细胞程序性坏死的机制研究李少军PP2Ac甲基化调控脂质自噬在苯并芘促进非酒精性脂肪性肝病中的作用研究李习艺炎症小体信号通路诱导的细胞焦亡在锰致机体认知损害中的作用邹云锋自噬在职业锰暴露致心脏毒性中的作用及机制研究杨晓波慢性低水平镉暴露经由ICAM-1和VCAM-1糖基化修饰促进血管内皮炎症的研究钟秋安miR-3942-5p在结肠癌中的功能机制研究及其在泛癌中的诊断和预后评估戴盛明增强Ⅱ型光敏剂介导的光动力抗白色念珠菌作用与机制研究黄力毅ICOS/ICOSL通路调控Th9分化介导日本血吸虫病肝纤维化的分子机制战廷正环状RNAcirc_103595/miR-185-3p/PLK1信号轴对肝癌不全消融后残余癌增殖侵袭的调控杨红多模态MRI对2型糖尿病小型巴马猪胰腺病变的病理对照研究及临床应用龙莉玲miR-106b-5p启动子甲基化状态介导MAPK/ERK信号通路调控腰椎间盘退变机制的研究江华EB病毒编码潜伏膜蛋白LMP2A通过ITGα6β4/ACSL4路径抵抗鼻咽癌细胞铁死亡温文胜MiR-3162-5p通过激活Ras信号通路促进宫颈鳞癌细胞EMT 的作用及机制研究陈军莹潜在抑癌基因LAMC3和LAMA2调控细胞自噬参与卵巢癌耐药的机制研究尹富强新生抗原neoantigen介导溶瘤病毒治疗复发性卵巢癌的应用及机制研究王琪OY-TES-1/miR-326协同调控PD-L1促胶质瘤免疫逃逸的机制初探罗彬SPINK4调控结肠癌细胞铁死亡的作用机制研究胡榜利冯振博hsa_circRNA_104515竞争性结合miR-1303调控PLIN1在肝细胞癌发生发展中的作用及机制HMGA1介导lncRNA-LINC01615增强THBS2表达及其在肝癌转移中的作用与机制研究唐博刘博白介素24定点整合自体诱导多潜能干细胞来源的间充质干细胞对子宫内膜癌的生长抑制红桂木凝集素扩增移植诱导的记忆干性T细胞抗肝癌研究周素芳RBM38结合并促进LncRNA-GAS5稳定逆转肝癌索拉非尼耐药的机制研究叶甲舟何宝玉长链非编码RNALOC553103通过ROCK1-RhoA/ROCK 信号通路调控细胞骨架重构促进鼻咽癌转高糖诱导肝细胞外泌体中miR-132差异表达促进EMT介导的胰腺癌转移的分子机制秦雯陈洁CirR-387与let-7d-5p竞争性调节GALNT1经EGFR/E-cadherin通路调控肝细胞癌EMT的机制广西HBV/AFB1双暴露女性原发性肝细胞癌早期预警模型的建立及相关分子机制的研究韩创业RNF125通过泛素化修饰PD-L1促进肿瘤免疫的分子机制研究吴飞翔基于UPLC-MS/MS联合MRM的靶向与非靶向代谢组学方法研究蛇伤中毒标志物廖明王威线粒体ATP敏感性钾通道和与线粒体自噬对急性心肌梗死大鼠心肌细胞的影响与机制研究淫羊藿苷经Notch通路调控大鼠ASCs促进超长随意皮瓣存活的机制研究梁至洁抑制PD-L1+中性粒细胞对脓毒症治疗的价值探索张森ERK抑制剂经由线粒体动力学-自噬途径促进心肺复苏后神经细胞生存研究陈蒙华弱酸性富钙磷离子环境下单核细胞来源外泌体的miRNA表达变化及其靶细胞效应研究廖红兵周诺lncRNA和MicroRNA调控胚胎内皮祖细胞Notch、BMP、Wnt 信号通路参与牵张成骨血管发生Notch信号通路在树鼩耳蜗感觉前体细胞增殖和分化中的调控作用谢利红宋星慧基于mTOR通路探讨外泌体介导树突状细胞活化在系统性红斑狼疮中的作用机制研究&nbs雷玲系统性硬化症中IL-35诱导iTr35细胞分化导致Treg/ Th17细胞失衡及纤维化病变的孔德燕黑色素纳米颗粒通过上调Iduna的表达促进间充质干细胞在缺血条件下的存活并增强其治刘斯佳基于石墨相氮化碳纳米片的双基因协同输送系统用于小鼠胚胎神经干细胞定向诱导分化G蛋白偶联受体MC1R抑制α-Syn聚集及细胞间传递的分子机制研究肖友生秦超内皮细胞膜微粒通过LncRNA MIAT调控神经元Beclin-1和Caspase-3 加重脑缺认知相关遗传变异及其与环境的交互作用对长寿人群认知功能的影响彭均华LncRNA NR_120526结合转录因子ILF2/3调控血红蛋白F合成的机制研究何云燕血栓弹力图评估遗传性异常纤维蛋白原血症患者凝血功能的价值研究闫婕应燕萍抗阻运动下调miR-92a-3p介导内皮细胞HMOX1/MAPKs/NF-κB预防导管相关性血栓形成的机制研究GLP-1受体激动剂通过IRS-1/PI3K/Akt信号通路调节骨代谢何玉玲基于静息态功能磁共振的急性甲亢肌病大脑活动变化及其分子机制研究罗佐杰PDK1调控破骨细胞在骨质疏松症发病中的分子机制研究宗少晖lnc-XLOC作为ceRNA参与miR-155调控脊柱结核椎间盘破坏过程中的作用机制研究詹新立可控释CGRP的硼硅酸盐生物活性支架促进骨缺损修复的作用机制研究李兵新型一氧化氮响应型纳米缓释微粒对骨性关节炎作用的研究靳攀陆真慧PHLPP1调节TNF信号通路在眼镜蛇毒神经生长因子(NGF)成软骨诱导中的作用及其机制山楂叶总黄酮调控微环境促进脊髓损伤后神经再生的分子机制研究曾高峰氧化应激介导的Nrf2/mTOR/NF-κB通路对氯胺酮相关性膀胱炎的影响和机制以及褪黑素的保护作用研究米华NF-κB/miR-375/CTGF信号介导咖啡因所致子代骨发育异常的血管发生机制罗瀚文HOXA10通过上调E-cadherin改善子宫内膜容受性的作用及机制研究杨一华极化的巨噬细胞通过NLRP3炎症小体调控肝纤维化的机制罗薇膜联蛋白A1在抗肝纤维化中的作用机制范俊华粪菌移植对滤泡辅助性T细胞和滤泡调节性T细胞调控在炎症性肠病中的作用研究吕小平FGF21调控ATF6/eIF2α/CHOP信号通路介导内质网应激肝损伤的分子机制研究郭超黄荣杰miR-155通过SHIP-1/p53MAPK/STAT3介导Th17细胞分化在盐敏感性高血压左室肥厚中的作曾晓聪环状RNAmmu_circ_0000021介导miR-143-3p靶向调控NPY 在心肌缺血再灌注微循环障碍的邱诗林IRF8/PD-L1通路在烟草暴露树突状细胞调控T淋巴细胞介导肺部炎症中的作用及机制研究烟草烟雾暴露下Tim-3调控CD4+T淋巴细胞介导COPD/肺气肿发生的作用及机制研究段敏超何志义中性粒细胞胞外陷阱(NETs)通过上调单核细胞PI3K-δ/Akt通路活性增加烟草烟雾暴露诱发的糖皮质激素抵抗?张景鸿基于ROS-NLRP3炎性小体/细胞焦亡途径研究活性维生素D3干预中性粒细胞性哮喘的作用基于单细胞测序及限时饲喂研究骨髓生物钟及概日节律王弘非神经性ACh在胆碱能M3受体调控的尿路上皮稳态增殖中的作用及机制研究易贤林接头连接蛋白在成年小鼠脑内神经发生中的作用机制研究刘媛媛HIV-1感染通过激活pDCs引起ILC2s损耗及机制研究叶力基于“络病”理论探讨壮骨方通过H型血管和成骨偶联干预糖尿病性骨质疏松的机制李双蕾参附注射液对连续灌注联合控制性通气保存供肺保护作用的机制研究莫安胜ciRS-7/miR-7/mTOR/p70S6K信号通路介导神经元自噬导致大鼠复苏后脑损伤的机制及参邓海霞天然牛磺酸对肝硬化门静脉高压抑制作用量子力学研究邓鑫秦刚基于IFITM1/Wnt/β-catenin信号通路调控骨肉瘤细胞恶性增殖探讨抑瘤汤抗骨肉瘤的机制复方扶芳藤合剂对小鼠单个核细胞亚群活化与分化的影响研究肖健基于IL-31—TRPV1轴的蛇床子素抗AD慢性瘙痒作用机制研究伍冠一基于GLUT4靶点的壮药“勾瓢更”抗Ⅱ型糖尿病药效物质基础及作用机制研究卢汝梅基于影响TLR4-NLRP3炎症小体信号通路对总丹参酮治疗肺炎作用机制研究高红伟基于G蛋白偶联受体相关通路的隐丹参酮抗血小板聚集机制研究刘振杰基于肝脏糖异生AMPK信号通路阐释三叶苷纠正2型糖尿病糖代谢紊乱的分子机制赵立春邱华白花香莲解毒颗粒调控Treg细胞Furin/TGFβ1正反馈环路促进慢乙肝HBeAg血清学转换的机制研究从LncBANCR介导ERK/mTOR调控自噬探讨推拿对SNL大鼠的镇痛效应机制卢栋明基于线粒体自噬探讨青光安对青光眼模型RGCs保护作用的机制研究姚小磊基于“微生物-肠-脑轴”探讨安神定志灵调控脑神经递质及免疫状态的机制研究孙继超基于“肺-肠轴”探讨壮医三十六荡坎蛤散治疗儿童哮喘缓解期的作用机制李伟伟王兵基于STAT3/TGF-β1/Smads信号通路调控EMT探讨大黄灵仙颗粒干预肝内胆管结石形成机制研究朱闽前列消汤干预自身免疫性前列腺炎IL-6-JAK2-STAT3信号通路调控CD4+T细胞凋亡及分化基于Erbin调控NF-κB信号通路研究安肠汤治疗溃疡性结炎的抗炎作用机制邓松华大黄煎剂作用于肠道菌群调节LPS/TLR4通路对MHE神经保护作用的机制研究付蕾长链非编码RNA对缺血性中风中经络不同证候的影响及功能机制研究古联健脾益气方通过IL-6/STAT3炎症信号通路抑制肝癌细胞Vimentin过度表达的机制研究卓少元陈延强基于“脾为之卫”理论从肠道微生态研究历节胶囊治疗胶原诱导性关节炎小鼠的作用机庞学丰基于Notch调控PI3K/Akt/mTOR信号通路调节T细胞糖代谢探讨寒痹康汤治疗类风湿关节炎麦门冬汤通过SDF-1/CXCR4轴对肺纤维化大鼠BMSCs迁移及归巢的影响刘锐黄连解毒汤调控SHR内质网应激通路的分子机制研究岳桂华陈峭基于钙离子振荡介导下的内质网应激-自噬反应探讨“以俞调枢”法对Cajal间质细胞的孟立锋内质网应激通路IRE1α-XBP1诱导腹膜间皮细胞自噬在EMT中的发生机制及健脾益气方的干预效应吕建林基于肝细胞lncRNA-miRNA-mRNA调控网络和“核-线粒体”信号传递探讨解毒化瘀颗粒拮基于自噬—NLRP3炎症体途径探讨芪七连胶囊保护SHR血管内皮损伤的效应机制研究罗远基于Rho/Rock信号通路探讨温肺降浊方靶向VaD大鼠突触损伤修复的分子机制胡跃强唐友明基于miR-584调控的内质网应激-细胞自噬途径探讨安胃汤干预胃黏膜肠上皮化生机制研基于神经血管单元探讨“三焦次第治疗”对脑缺血损伤大鼠Wnt/β-catenin信号通路的调控机制唐农李晓红ARHGEF-7在慢性应激肝郁脾虚证大鼠模型海马、胃组织RhoA/Rac1/CDC42/PAKs细胞骨架夏猛基于外泌体的功能特点以及胶质—神经细胞的相互作用关系探讨加味逍遥散治疗肝郁脾禽呼肠孤病毒σC蛋白抗肿瘤作用及其分子机制的研究黄莉基于代谢组和蛋白组学对大鼠射精机理脑神经物质基础的研究覃喜军真菌促进濒危药用植物青天葵种子萌发作用的研究谭小明广西特有濒危地方品种“瑶蚕”的生物学性状与进化起源研究张桂征纺锤体组装检查点介导胚胎染色体嵌合自我校正机制研究桂宝恒miR-483-5p作为乙肝病毒双突变(1762T,1764A)株感染者肝癌高危标记物的前瞻性研究方钟燎羟基羧酸催化蒎烯水合/乙酯化反应机理研究孟中磊炭疽病胁迫香蕉中磷脂酸代谢关键酶基因表达对其生成转化的调控机制研究孙健葡萄转录因子VlNAC60应答高温胁迫的功能及调控机制郭荣荣光调控广西毛葡萄酚类物质代谢机理研究成果SlWRKY1调控番茄抗南方根结线虫防卫反应的分子机制研究陆秀红葡萄霜霉菌无毒基因AvrRpv1参与的菌株毒力变异分子机制研究尹玲新型抑菌活性物质DMTS作用于胶孢炭疽菌麦角固醇生物合成的机制唐利华普通野生稻抗褐飞虱基因Bph33(t)的克隆及育种利用张月雄王泽平甘蔗抗梢腐病氮代谢和系统获得性抗性途径关键组分γ-谷氨酰转移酶基因SoGGT1克隆及功能鉴定基于 CRISPR-Cas9 技术的广西地方花生优良品种油酸含量改良研究贺梁琼小粒野生稻抗白背飞虱新基因WBPH9的克隆与抗虫机理分析郭嗣斌一个广谱持久抗稻瘟病基因pi-DY的克隆和功能分析高利军水稻东兰墨米种皮花青素合成关键基因BP3的图位克隆及功能研究杨行海伯克氏固氮菌GXS16与甘蔗根系高效联合固氮的生理和分子基础研究李长宁粉垄栽培木薯的肥料养分利用效率机制及其环境影响效应申章佑蔗叶与蔗叶生物炭还田下其C、N协同归还研究刘昔辉糖厂滤泥还田对广西红壤蔗区土壤有机碳库和温室气体排放的影响朱晓晖粉垄耕作对甘蔗土壤碳固定和温室气体排放的影响及机制黄金生霉变甘蔗病原菌节菱孢菌株产毒能力评价及产毒调节机理研究莫磊兴线粒体自噬NIX和Pink1信号交互作用对索拉非尼治疗肝癌的影响及干预研究张国染料木黄酮通过TTTY18/ miR-95/ SGK1环路干预结直肠癌细胞增殖凋亡的作秦俭Atg5介导树突状细胞自噬在烟草烟雾暴露下肺部炎症中的作用及机制的研究邝良鉴澳洲坚果种质资源的分子遗传多样性研究蔡元保基于miRNA和蛋白质组的桑寄生顽拗性种子脱水敏感性分子机制研究潘丽梅三倍体罗汉果种子败育分子机理研究韦荣昌草果种子休眠解除关键蛋白的挖掘及基因功能研究潘春柳南方石漠化区植被修复的微地形效应及其适应性管理策略研究吕仕洪东兴金花茶和同域分布的近缘广布种长尾毛蕊茶生殖生态学特性比较研究韦霄基于稳定同位素技术研究喀斯特季节性雨林代表性植物水分利用策略黄甫昭广义石山苣苔属(苦苣苔科)系统发育重建和分类学修订温放自然选择和遗传漂变对喀斯特特有铁甲秋海棠复合群物种分化的影响董莉娜中国冷水花属(荨麻科)分类修订和系统学研究韦毅刚苦苣苔科吊石苣苔属的分类学研究许为斌中国典型喀斯特天坑土壤微生物分布格局及其驱动机制蒲高忠RSK4靶向PI3K/AKT通路调控ACTN4在乳腺癌化疗耐药中的机制研究杨华伟应用时间系列代谢组学研究肝细胞肝癌患者手术前后血清代谢物变化及潜在标志物识别黎远冬缺氧条件下miR-210-5p调控SPON2介导肝癌TAM细胞M2型极化参与耐药的机制研究罗小玲用于肝癌早期诊断的多通道纸基微流控生物传感器的研究陈真诚西番莲酸性多糖改善2型糖尿病糖脂代谢紊乱的机制研究李霞基于miR-378b-p110α-PI3K/Akt通路甲基阿魏酸改善糖脂代谢抗ALD的机制研究李丽异槲皮苷介导miRNA-29调控靶基因改善胰岛素抵抗的作用机制黄桂红多靶点复合杯芳烃金纳米粒在阿尔茨海默症的诊断治疗作用及分子机制研究莫靖欣三株西沙软珊瑚共附生真菌抗肿瘤活性成分发现及其作用机制研究李云秋基于TLC和iTRAQ技术进行地蚕中抗金黄色葡萄球菌环肽的发现和其作用机制研究梁成钦李清初基于miR-21调控NF-κB信号通路介导肾缺血再灌注诱导的凋亡和炎症探讨桂枝去桂加茯苓白术汤的肾保护作用机制的研周燕园基于PPARγ/TGF-β1/Samds通路探讨鼠李柠檬素改善多囊卵巢综合征(PCOS)大鼠卵巢纤维化的分子机制麝香酮和适配体级联修饰多烯紫杉醇脑靶向递送系统抗胶质瘤作用及机制的研究齐娜抑癌基因CMTM4在肝细胞癌中的作用及机制研究谭盛葵PP2A介导MYC相关锌指蛋白在肝细胞恶性转化中的作用机制研究朱小年发育相关印记基因表观遗传学改变介导孕期PEs暴露与妊娠结局吕元明IL-33/ST2通路调控CD8+T和免疫抑制相关细胞的功能抗软组织肉瘤的作用和机制陈火英阻断VEGF/VEGFR-2信号通路逆转TAMs极化在乳腺癌T细胞免疫中的作用及机制研究何少忠lncRNA Loc554202通过调控染色质状态促进脊索瘤发生和进展的机制研究夏学巍张学梅m6A甲基转移酶METTL3调控TXNIP对结直肠癌生物学特征的影响和机制研究KDM5B促进胃癌侵袭、转移的作用及机制研究王振冉PAIP1-mTOR正反馈环路促进肝癌发生郑剑锋卵巢癌产生的IgG在维持卵巢癌干细胞生物学行为中的作用及其机制研究廖沁园表观遗传介导IRF7相关lncRNA FTX抑制非小细胞肺癌发展的作用及机制研究杜振宗长链非编码RNA XLOC_012366在表观遗传学介导的肺癌转移抑制中的机制研究宋剑非Nrf2基因修饰的脂肪间充质干细胞对硬皮病的治疗作用及机制研究罗婧莹闫建国帕金森病中CDK5磷酸化依赖的C9orf72泛素化降解介导alpha-synuclein清除障碍和神经贺亮米诺环素通过Notch信号通路调控小胶质细胞极化在脊髓缺血后延迟性瘫痪中作用的研究王勇基于HMGB1-TLR4介导的NF-κB信号通路探讨毛蕊异黄酮在脑缺血再灌注损伤中的保护机制不同人胰淀素聚集程度通过TLR4/MyD88介导调节性T细胞免疫机制张晓溪细胞焦亡在糖尿病肾病足细胞损伤中的机制研究周素娴姚军无抗冻剂超快速冷冻兔睾丸/附睾精子对ICSI胚胎及子代印记基因。
2015年广西科学研究与技术开发计划项目(课题)表(第三批).xls

2-3-7
2-3-8
20
樊兰兰 2015ED26 007
3 3-1 3-1-1
630 315 50 冯诚诚 2015DD24 012 廖乃雄 2015DD24 006 梁志坚 2015DD24 017 尹意求 2015DD24 002 李建强 2015DD24 034 陈桂芬 2015DD24 020 曹斌 2015DD24 040 彭慧婧 2015DD24 030 赵晓芳 2015DD24 039
2015年广西科学研究与技术开发计划项目(课题)表 (第三批)
序号 1 1-2 1-3 财政拨款 课题负 申报编号 (万元) 责人 国际科技创新成果引进与合 2015.011440 作研究开发 2017.12 航空用硬铝合金扁锭软启铸 广西南南铝加工有限公司 2015.0180 何克准 2015DD23 技术引进与合作开发 2017.12 033 水牛重要经济性状遗传解析 广西壮族自治区水牛研究所、广西华 2015.0180 梁贤威 2015DD23 关键技术的合作研究及其应 胥水牛繁育有限公司 2017.12 028 用 航空用铝合金板材残余应力 广西南南铝加工有限公司 2015.0180 赵解扬 2015DD23 相控阵超声波检测技术与合 2017.12 042 作开发 蔗渣木素催化转化制备对乙 广西科学院、复旦大学 2015.0180 黎演明 2015DD23 基苯酚的关键技术研究 2017.12 053 生物蛋白大分子核磁共振分 广西科学院 2015.0180 廖思明 2015DD23 析技术的引进与合作研究 2017.12 054 靶向肿瘤记忆性干T细胞疫 广西医科大学 2015.0160 周素芳 2015DD23 苗的研究 2017.12 039 木质纤维水解糖化技术的引 广西科学院 2015.0160 芦志龙 2015DD23 进与合作研究 2017.12 055 抗疟药制剂和原料药技术引 桂林南药股份有限公司 2015.0150 刘永宏 2015DD23 进创新与合作研发 2017.12 006 2.6米双轴定向特种薄膜生 桂林电器科学研究院有限公司 2015.0150 冯勇刚 2015DD23 产线研制 2017.12 011 基于高内涵筛选平台的京族 广西中医药大学 2015.0150 邓家刚 2015DD23 习用海洋药物抗癌新药发现 2017.12 036 关键技术合作研究 亚纳秒级ZnO:Ga闪烁单晶的 中国有色桂林矿产地质研究院有限公 2015.0150 左艳彬 2015DD23 水热法生长技术研究 司(国家特种矿物材料工程技术研究 2017.12 008 中心) 定性与定量核磁共振技术在 广西壮族自治区中国科学院广西植物 2015.0150 王磊 2015DD23 广西特色中草药的开发中应 研究所 2017.12 024 用 微藻处理发酵工业废水技术 南宁邦尔克生物技术有限责任公司、 2015.0150 蒙健宗 2015DD23 合作研究 广西大学 2017.12 041 整合型表达系统分泌表达碱 广西科学院、南宁市新科健生物技术 2015.0150 王青艳 2015DD23 性脂肪酶的研究及产品开发 有限责任公司 2017.12 049 葡萄重要病害快速检测及脱 广西特色作物研究院 2015.0140 娄兵海 2015DD23 毒技术合作研究 2017.12 005 船舶防锈颜料先进技术的引 广西新晶科技有限公司 2015.0130 李开成 2015DD23 进与合作研究开发 2017.12 038 广西富硒功能农产品研发关 广西壮族自治区农业科学院农业资源 2015.0130 刘永贤 2015DD23 键技术引进与创新 与环境研究所 2017.12 010 火龙果功能特性及加工新技 广西壮族自治区农业科学院农产品加 2015.0130 李丽 2015DD23 术的引进与合作研究 工研究所 2017.12 037 超低成本五分类血细胞分析 桂林优利特电子集团有限公司 2015.0130 唐雪辉 2015DD23 仪的研制 2017.12 001 耕地土壤综合肥力评价及研 广西壮族自治区农业科学院农业资源 2015.0130 苏天明 2015DD23 究技术引进与创新 与环境研究所 2017.12 013 基于眼动脑波控制技术的认 广西壮族自治区人民医院 2015.0130 秦岭 2015DD23 知评价训练平台的研发 2017.12 045 国际科技合作与技术转移服 2015.01780 务平台建设 2016.12 项目(课题)名称 承担单位 起止年月
我国林蛙属物种的分类学研究综述

我国林蛙属物种的分类学研究综述作者:崔光旭来源:《安徽农业科学》2018年第14期摘要林蛙(Wood frog)是一类具有背侧褶、非繁殖期生活在山林中的棕色蛙类,是两栖类的重要组成部分,主要分布于亚欧大陆的暖温带地区和北美洲西部。
从形态分类学、细胞分类学、分子系统学3个方面对我国林蛙属物种的分类学研究进展进行了综述,以期为林蛙属的进一步研究提供资料。
关键词林蛙;形态分类学;细胞分类学;分子系统学中图分类号 S966.3+5 文献标识码A 文章编号 0517-6611(2018)14-0027-02Research Progress on the Taxonomy of Rana Genus in ChinaCUI Guangxu (School of Life Sciences, Ludong University, Yantai,Shandong 264039)Abstract Wood frog is a kind of brown frogs with dorsolateral fold, which is an important part of amphibians.They live in mountain forest during the nonreproductive period.The warm temperate region of Eurasia and the western part of North America are their mainly distribution area.From three aspects of morphological taxonomy, cytotaxonomy and molecular systematics, this paper reviewed the research progress on the taxonomy of Rana genus in China, in order to provide materials for further study of Rana genus.Key words Wood frog;Morphological taxonomy;Cytotaxonomy;Molecular systematics林蛙(Wood frog)是一类具有背侧褶、明显鼓膜、非繁殖期主要生活于山林中的棕色蛙类,因此也称为棕蛙(Brown frog)。
2018年第一批广西自然科学基金项目

92 93 94 95 96 97 98 99
Ni纳米晶金属蠕变行为的晶体相场模拟与微观机理研究 乙酰基-L-肉碱对不同繁殖季节水牛卵子体外成熟的影响及其调控类固醇激素合成 的研究 在液化天然气温区实现高温超导输电的Tl-2223薄膜及其带材的制备与性能研究 广西和越南北部边境地区HIV传播性耐药及其跨境传播特点 槐树微卫星(SSR)引物开发及槐米芦丁含量地理变异格局与空间遗传结构的关联 分析 不同PRRSV毒株感染诱导免疫细胞氧化应激及其组蛋白乙酰化修饰的研究 U-2OS细胞特异性DNA适配体筛选及应用于小鼠骨肉瘤模型靶向治疗研究 Nur77调控脂多糖介导人肺泡上皮细胞ECM稳态的信号转导机制
10 10 10 9 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10
10 10 10 9 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10 10
广西民族大学 广西大学 广西大学 广西师范学院 广西壮族自治区人民医院 广西中医药大学 广西医科大学 广西医科大学 贺州学院 广西医科大学 广西师范大学 桂林理工大学 广西中医药大学 广西医科大学 广西医科大学 广西壮族自治区肿瘤防治研究所 广西大学 广西壮族自治区农业科学院甘蔗研 究所 桂林医学院 广西医科大学 广西医科大学 柳州市工人医院 桂林电子科技大大学 广西壮族自治区兽医研究所 广西壮族自治区农业科学院葡萄与 葡萄酒研究所 桂林理工大学 桂林理工大学 桂林电子科技大学 桂林电子科技大学 玉林师范学院 广西科学院 广西师范大学 广西师范大学 广西师范大学 广西中医药大学 广西医科大学 广西医科大学 桂林理工大学 广西大学 广西壮族自治区林业科学研究院 桂林理工大学 桂林电子科技大学 广西师范大学 广西师范大学 广西医科大学 广西医科大学
广西姑婆山及邻近地区大型真菌物种多样性

广西姑婆山及邻近地区大型真菌物种多样性作者:张艳成张强牟光福刘演赵博来源:《广西植物》2023年第12期摘要:姑婆山緊邻南岭生物多样性保护优先区,孕育了丰富的生物资源。
为了解该区域大型真菌物种多样性,该文采用随机踏查法对广西姑婆山自治区级自然保护区(以下简称姑婆山)及邻近地区大型真菌资源进行调查采集,并结合形态学和分子生物学方法进行物种鉴定。
结果表明:(1)共发现348种大型真菌,隶属于2门6纲17目69科175属,其中包含食用菌40种、药用菌42种、毒菌7种。
(2)对该区的物种组成分析表明,优势科为蘑菇科(Agaricaceae)、牛肝菌科(Boletaceae)、粉褶蕈科(Entolomataceae)和锈革菌科(Hymenochaetaceae)等9科;优势属为鹅膏属(Amanita)、靴耳属(Crepidotus)、毛皮伞属(Crinipellis)和粉褶蕈属(Entoloma)等11属。
(3)共发现2个中国特有属,即脆孔菌属(Fragiliporia)和臧氏牛肝菌属(Zangia);11个中国特有种,即厚集毛菌(Coltricia crassa)、魏氏集毛菌(C. weii)、丛生粉褶蕈(Entoloma caespitosum)、极细粉褶蕈(E. praegracile)、近薄囊粉褶蕈(E. subtenuicystidiatum)、薄管层卧孔菌(Fomitiporia tenuitubus)、香榧嗜蓝孢孔菌(F. torreyae)、脆孔菌(Fragiliporia fragilis)、南岭多年卧孔菌(Perenniporia nanlingensis)、亚白灰孢多年卧孔菌(P. subtephropora)以及脑形银耳(Tremella cerebriformis)。
(4)该区域大型真菌科级区系地理成分可分为世界广布成分(66.67%)、泛热带成分(17.39%)以及北温带成分(15.94%);属级区系地理成分可分为7类,主要以世界广布成分为主(61.71%),其次为泛热带成分(20.57%)和北温带成分(13.71%)。
中国十三省农田池塘蛙类的衰减

中国十三省农田池塘蛙类的衰减戴建华;马佳月;徐玲琳;魏永强;顾易凡;周开亚【摘要】Field trips for frog specimens were made in Fujian, Jiangxi, Zhejiang, Jiangsu, Hunan, Hubei, Anhui,Henan, Hebei, Shandong, Shanxi, Liaoning and Jilin provinces between the year of 2004 and 2009. The declines of farmland and pond frogs in these areas were recorded during the field works. Threats to frogs include loss and degradation of habitat, pollution, overfishing, and effects of alien invasive species. Measures for conservation of farmland and pond frogs were recommended.%2004~2009年,研究者在福建、江西、浙江、江苏、湖南、湖北、安徽、河南、河北、山东、山西、辽宁、吉林等地进行蛙类标本采集.在野外工作中记录了这些地区农田池塘蛙类衰减的情况.导致蛙类衰减的主要因素包括:栖息地丧失与退化、污染、过度捕捉和外来入侵种的影响.本文还提出了保护农田池塘蛙类的几项措施.【期刊名称】《南京师大学报(自然科学版)》【年(卷),期】2011(034)001【总页数】6页(P80-85)【关键词】中国;农田池塘蛙类;衰减【作者】戴建华;马佳月;徐玲琳;魏永强;顾易凡;周开亚【作者单位】南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046;南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046;南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046;南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046;南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046;南京师范大学生命科学学院,江苏省生物多样性及生物技术重点实验室,江苏南京210046【正文语种】中文【中图分类】Q9两栖类是对环境变化非常敏感的动物,可以作为环境变化的“指示种”.两栖类的严重衰减引起了人们的广泛关注[1-6].有研究表明,近几十年内,大约有近 100种蛙类灭绝,约 1/3的种类处于受威胁和濒危状态.世界自然保护联盟 (I UCN)和世界动物园及水族馆协会 (WAZA)共同宣布,将 2008年定为“世界蛙年”,以保护日益减少的以蛙类为主的两栖动物多样性.中国蛙类资源也遭受了非常严重的破坏.据《中国物种红色名录》,中国广布的 32种蛙科动物中有 11种生存受到威胁,5种处在近危、6种为易危状态.在中国窄布的91种蛙科动物中 17种被评估为近危,31种被评估为易危,4种被评估为濒危,3种被评估为极危[7].中国两栖动物中极危、濒危和易危种类达到40%,而全球的平均值是 32.5%[8,9].可见,中国两栖动物受胁程度非常严峻,两栖类的生存令人担忧. 2004~2009年,研究者赴福建、江西、浙江、江苏、湖南、湖北、安徽、河南、河北、山东、山西、辽宁、吉林等地采集蛙类标本,并记录了这些地区农田池塘蛙类衰减的情况.导致蛙类种群数量下降最主要、最直接的原因是栖息地的丧失与退化.近几十年来,由于城市及农村基础建设速度的爆炸性增长,城镇住宅区的扩张和农业、工业、旅游业的发展,导致森林、河流、湖泊、池塘、沟渠、沼泽、草地等适合蛙类生存的环境被破坏.一些地区的小面积农田也使得蛙类栖息地碎片化、分布范围缩小、种群遗传多样性降低、蛙的种群数量衰减,甚至出现局部灭绝[8,10].湖南省宜章县是湖北侧褶蛙 (Pelophylax hubeiensis)分布的最南界[11],以前湖北侧褶蛙在该县分布非常广泛 (沈猷慧教授个人交流).2006年 6月,研究者在宜章采集标本时发现当地环境变化巨大,县城原有大面积的适合黑斑侧褶蛙 (Pelophylax nigrom aculata)及湖北侧褶蛙生活的荷塘已被毁坏殆尽.沈猷慧教授当年采集湖北侧褶蛙的地方已经被建成城区.在县城附近以及该县一些农村地区,虽然发现了很多适合湖北侧褶蛙生存的环境,但是在 4天时间里均未发现湖北侧褶蛙 (表 1).据河南师范大学瞿文元教授介绍,在河南省西部栾川、灵宝、卢氏等地均有金线侧褶蛙分布.2006年 8月,研究者在瞿教授介绍的采集地灵宝市朱阳乡调查,那里的环境已被破坏,河床鹅卵石暴露,水土流失,没有发现金线侧褶蛙.在附近水塘、水库、小河寻找,除黑斑侧褶蛙外未发现金线侧褶蛙.根据瞿教授的指导,研究者在栾川县很多地方寻找金线侧褶蛙,均没有发现.瞿教授以前采集到金线侧褶蛙的五里川乡老婆沟,环境已完全被破坏,除还有很少的黑斑侧褶蛙,未发现金线侧褶蛙.蛙类在浙江省嵊泗县的现状也许能反应栖息地破坏对蛙类生存的影响情况.2005年10月,研究者在嵊泗本岛采集标本,在很少的几个池塘中能发现蟾蜍 (B ufo gargarizans),黑斑侧褶蛙数量极少.研究者几乎找遍了整个嵊泗岛,仅在菜园镇一条很小的菜地沟里发现了十几只包括当年出生的小蛙在内的金线侧褶蛙.以前这里是一片水稻田,现在被开发成了一片住宅区,仅有的这块菜地旁仍然在修建房屋,金线侧褶蛙在嵊泗岛面临灭绝的危险.蛙类种群数量急剧减少与日益严重的环境污染密切相关,污染物质包括工业废水、生活废水及农药等.蛙类需要水域及陆地环境来完成其生活史,而大部分的化学污染物最终都要排入水域,所以化学污染物对蛙类的毒害作用是直接的.对蛙类发育的影响日益严重的有农药 (除草剂、杀虫剂等)和重金属离子.不仅残留的除草剂和杀虫剂会对蛙蟾类造成毒害[12,13],重金属污染对蛙蟾类造成的毒害也极为严重.很多实验证明,铜、镉、铅、汞、铬、锌等重金属对蛙蟾类的毒害作用在形态、器官、组织、细胞和分子水平上均有表现[14-21],其危害甚至从胚胎时期就开始了[22].这些污染物质既造成蛙类的适宜栖息地减少,还可以通过食物间接影响蝌蚪的发育,导致发育过程中出现畸形.在野外发现的农药有井冈霉素、比双灵、噻嗪酮、杀虫单、螟虱一次净、吡虫灵、扑虱灵、杀虫双、毒敌等,这些农药在很多地方被大量使用甚至滥用,对环境造成了很大的危害.由于废水、农药等污染,福建省农田池塘蛙类的数量急剧减少.2004年 7月,研究者在福建省进行调查.在福安市赛岐镇几个样点,除少数沼水蛙 (Hylarana guentheri)外,没有发现黑斑侧褶蛙及福建侧褶蛙(Pelophylax fukienensis);在福州市闽侯县荆溪镇,由于水体富营养化及污染严重,池塘及荷塘水体发黑,除在河边发现少量沼水蛙外,没有发现黑斑侧褶蛙及福建侧褶蛙;在漳州龙海市程溪镇林下村大片水稻田中,由于农药化肥的大量使用,除发现少量泽陆蛙 (Fejervarya m ultistriata)外,未发现黑斑侧褶蛙和福建侧褶蛙;在武夷山市星村镇稻田、水沟及池塘,除沼水蛙外,2个晚上只发现 1只福建侧褶蛙,没有发现黑斑侧褶蛙;在太宁县朱口镇,只发现很少沼水蛙及黑斑侧褶蛙,没有发现福建侧褶蛙.2006年 4月研究者在福建省福清市南岭镇五龙村调查时发现,由于大量圈养生猪,猪场排出的粪便严重污染下游水体,河水发黑变臭,致使鱼虾大量死亡,当地以往常见的黑斑侧褶蛙及福建侧褶蛙已经很难寻觅.据文献记载,在山西省的运城、临汾、太原等地均有金线侧褶蛙分布.2005、2006年研究者先后 2年在清徐县、曲沃县、运城市的盐湖区、永济市、侯马市、太原市等地(未去历山保护区)寻找金线侧褶蛙.但这些地方环境污染非常严重,一些河流、池塘水体发黑发臭.虽然一些地方黑斑侧褶蛙数量还比较多,但是在很多原本适合金线侧褶蛙生存的栖息地,由于水体的严重污染,一只金线侧褶蛙也没有发现.山西省太原市晋祠镇是山西省传统种植水稻的极少地区之一.2006年 8月 8日晚,研究者在晋祠镇附近的稻田及田间水沟里寻找了近 5 h,竟然没有发现一只黑斑侧褶蛙和金线侧褶蛙 (后在一个水塘里发现少量的黑斑侧褶蛙).由于蛙类的几近灭绝,当地蝗虫数量惊人.用普通捕虫网做了捕捉统计:用捕虫网贴着水稻快速扫半圈,然后统计网中蝗虫数量.10次一共捕捉 389只,这是在很多蝗虫在扫网接近时受惊吓飞走的情况下的捕捉结果,当地蝗虫的密度由此可想而知了.据访问当地农民得知,十几年前当地蛙类数量非常多,夏季夜晚真正是“稻花香里说丰年,听取蛙声一片”.可近些年当地蛙类资源极度衰竭,稻田及田间水沟里几乎看不到了.湖北省鄂州市汀祖镇一个村,20年前蛙类非常多.由于一个选矿厂多年排放的废水的污染,还有农药的滥用以及滥捕,如今这里蛙类几乎灭绝.研究者于 2004、2006和 2009年的 7月 3次赴江西赣县采集标本,由于水体污染、农药滥用等原因,在一些原先适合蛙类生存的荷塘、鱼塘、稻田、水沟等生境,一只金线侧褶蛙也没有发现.由于政策法规的不够完善,蛙类中的经济物种被过度捕捉的情况非常严重,主要是黑斑侧褶蛙、金线侧褶蛙、湖北侧褶蛙、福建侧褶蛙、棘胸蛙 (Paa spinosa)、沼水蛙、林蛙 (Rana dybowskii),蟾蜍也大量被捕捉用于实验、制药及食用.很多地方甚至捕捉国家二级重点保护动物虎纹蛙 (Haplobatrachus chinensis).蛙类的贩卖在中国很多地方非常猖獗.福建省很多地方农贸市场有蛙类出售,如邵武市、武夷山市、福清市、福鼎市、南靖县、连城县、沙县等地的农贸市场.这些市场除卖黑斑侧褶蛙、沼水蛙外,还卖虎纹蛙和棘胸蛙.2006年 4月 20日,研究者在柘荣县某农贸市场见到一位农民一次出卖 25 kg棘胸蛙,获利2 000多元.江西省捕捉出售蛙类的情况非常严重,几乎所有的市、县农贸市场均有蛙类出售,一些地方有人定时、定点收购蛙类及蛇类等野生动物.在黎川、赣州、宁都、萍乡、南康、抚州、崇义、万载、万高、奉新、余干、修水、武宁、铜鼓、铅山、上饶、樟树、遂川、吉安、湖口、瑞金、泰和等地的农贸市场都发现有蛙类出售.很多市场还公开出售国家二级保护动物虎纹蛙,如上犹、会昌、于都、赣州、瑞金、泰和、遂川等地农贸市场.2006年 7月 4日早晨,研究者在泰和县某农贸市场一个摊位发现丢弃的死亡的虎纹蛙 30多只.湖南省和湖北省几乎所有的市县农贸市场均有蛙类出售,除出售黑斑侧褶蛙及湖北侧褶蛙外,有的还出售虎纹蛙.安徽省的情况也很严重,几乎所有的市、县农贸市场均有蛙类出售.2004年 10月,在蚌埠市某镇一个十几年来一直从事贩卖野生动物的窝点,研究者发现至少有 100多个装满了蛙和蟾蜍的泡沫冰盒,两辆卡车准备将这些蛙和蟾蜍运到上海.据当地人介绍,该窝点长期向附近农民收购蛙类,甚至在冬季也捕捉冬眠中的蛙蟾类出售.浙江省、江苏省也有很多市县农贸市场出售蛙类,甚至在管理较严的南京市也有一些农贸市场偷卖蛙类.2007年,洪泽湖地区公安机关曾截获一卡车非法捕捉的黑斑侧褶蛙和金线侧褶蛙.2010年 8月 2日晚,江苏省沭阳县公安局执法人员查获 5位农民捕获的金线侧褶蛙和黑斑侧褶蛙共 800多只.人们对野生蛙类的滥捕导致虫害猖獗,于是又不得不大量施用农药,这又造成生态环境破坏和污染的进一步加剧,导致蛙类的进一步减少.外来入侵物种可使原有生态系统的生物种类和数量发生显著改变,从而导致生态系统结构的变化.外来入侵物种与本地物种之间的生态位竞争(生活空间、食物等竞争)会影响当地物种的生长和繁殖,导致本地物种种群数量的减少甚至灭绝[23].研究者这几年在福建、江西等地调查的过程中,发现外来入侵种福寿螺在野外泛滥的状况触目惊心.从福寿螺在中国南方地区的泛滥情况看,人们对牛蛙的担心也许并不是多余的.牛蛙是我们非常熟悉的外来入侵物种,已经在一些地方发现了野外种群.2005年 10月,研究者在浙江省舟山市诸岛采集标本时在岱东、衢山、高亭、虾峙岛、湖泥岛等地的稻田、水沟、池塘里寻找了数日均未发现金线侧褶蛙,但是发现了逃逸的已经适应野外生存的牛蛙.除和当地的黑斑侧褶蛙、金线侧褶蛙抢占生存空间外,由于个体大、生长快,牛蛙蝌蚪在竞争中往往占据优势.有报道证实,牛蛙蝌蚪与其他蛙类蝌蚪间具栖息地竞争和食物竞争,甚至可以减少其他蛙类蝌蚪的取食活动以及影响它们变态时的身体大小和变态比例,降低它们的存活率,从而改变当地蛙的栖息地利用格局[24].研究证明,牛蛙的入侵已导致世界许多地区蛙类和蛇类种群数量严重下降、分布区缩小,甚至局部灭绝,极大地破坏了当地生态系统的结构[1,3,4,6,25].总体来看,农林公安部门在保护蛙类资源方面做了一些工作,但由于老百姓,包括一些执法部门保护蛙类的意识不强,所以成效普遍不显著.如前面提到的很多农贸市场里公开出售蛙类,一些市场管理人员只要收取了管理费,并不干涉人们买卖蛙类,即使是国家重点保护动物虎纹蛙亦公开出售.在很多地方甚至有专门在农贸市场出售蛙类的菜贩子,如江西省赣州市某农贸市场有几个摊位长期出售蛙类,每个摊位一天要卖掉很多蛙.值得庆幸的是,一些地方执法部门已经认识到保护野生动物的重要性,也制定了一些相关法规,加大了对非法捕捉野生动物的打击力度.多年来,受江苏省林业厅的委托,研究者对本省各地非法捕捉的蛙类进行司法鉴定,发现对蛙类的捕捉现象仍然严重.这几年江苏省加重了对非法捕捉蛙类的打击,研究者在进行司法鉴定时也了解到了一些案例.比较南方一些省份,北方贩卖蛙类的情况要少得多,这与当地的饮食习惯有关,也得力于有关部门对非法捕捉的打击.如辽宁省凌海市阎家镇,前些年有人大肆捕捉黑斑侧褶蛙,向广东等地贩卖蛙肉.近些年当地执法部门加大了对野生动物的保护,农、林、公安等部门对非法捕捉蛙类从重处罚,捕捉蛙类的情况少了很多,蛙类资源有了较大的恢复.但是,同样是在东北地区,由于林蛙的特殊营养作用,长期的大量捕捉导致资源几近枯竭.虽然蛙类种群数量减少的原因各种各样,但是现存的两栖动物所面临的主要威胁是人类活动对栖息地的破坏以及过度捕捉.无论是从物种多样性还是独特性来看,中国都是全球两栖动物保护的一个优先区域.在保护蛙类资源方面,迫切需要做好以下工作:(1)控制、消除各种理化污染源,保护和改善适合蛙类生存的环境蛙类的生长和繁殖需要湿润的陆地环境和合适的水体环境,生态环境的破坏是蛙类生存的最大威胁.为了保护蛙类资源,我们必须保护好蛙类的栖息地.在栖息地碎片化的地区,应尽量保留、改造出一些绿地、池塘、水沟及湿地等适合蛙类生存的环境.此外,还应治理被污染的环境,在农业、林业生产中选用高效、低毒、少量、分解快、少残毒甚至无残毒的农药,减少对土壤和水体的污染,保护和恢复蛙类的栖息地. (2)加强公民保护野生动物意识,加强法律、法规教育,提高蛙类的保护等级广泛宣传野生动物保护的各种法律法规,提高全民野生动物保护意识.这是保护野生动物,包括蛙类的最根本、最有效的措施.虽然很多省市将黑斑侧褶蛙、金线侧褶蛙、福建侧褶蛙、湖北侧褶蛙、棘胸蛙、棘腹蛙、林蛙等可食用蛙列为重点保护动物,但除虎纹蛙以外,其他的蛙类均没有被列为国家级保护动物,使得执法部门在处罚非法捕捉行为时仍然觉得缺乏法律支持,这不利于蛙类资源的保护,所以建议提高这些受胁蛙类的保护等级.(3)开展人工养殖,加强蛙类资源的综合利用和保护在短时期内改变人们食用蛙类的习惯非常困难,因此,对经济价值较高的蛙类进行商业化养殖是解决供需矛盾的有效途径.有几种蛙类的人工养殖已非常成功,如中国林蛙、牛蛙以及虎纹蛙.对棘胸蛙、虎纹蛙、中国林蛙、牛蛙等这些人们需求较大的蛙类进行人工繁殖,扩大养殖种群后再上市,既能满足人们食用的需要,又有望避免对野生种群的捕捉,从而保护野生蛙类资源.【相关文献】[1] Alford R A,Richards SJ.Global amphibian declines:a problem in applied ecology[J].Annu Rev Ecol Syst,1999,30:133-165.[2] Alford R A,Dixon PM,Pechmann J H.Ecology.Global amphibian populationdeclines[J].Nature,2001,412:499-500.[3] Blaustein A R,Kiesecker J plexity in conservation:lessons from the global decline of amphibian populations[J].E-cologyLetters,2002,5:597-608.[4] Collins J P,StorferA.Global amphibian declines:sorting the hypotheses[J].Diversity and Distribution,2003,9:89-98.[5] KatsL B,Ferrer R P.Alien predators and amphibian declines:review of t wo decades of science and the transition to conservation[J].Diversity and Distribution,2003,9:99-110. [6] Kiesecker J M,Blaustein A R,Belden L plex causes of amphibiandeclines[J].Nature,2001,410:681-684.[7] 江建平.中国蛙科动物的生存状况[J].大自然,2009(4):28-31.[8] 谢锋,刘惠宁,Stuart SN,等.中国两栖动物保护需求总述[J].中国科学:C辑生命科学,2006,36(6):570-581.[9] 于凤兰,陆宇燕.中国特有两栖类受胁现状分析[J].四川动物,2006,25(2):323-325.[10] 龚大洁,吴洪超,侯峰,等.甘肃康县棘腹蛙种群资源调查与分析[J].长江流域资源与环境,2009,18(12):1 162-1 165.[11] 费梁,叶昌媛,江建平,等.中国两栖动物检索及图解[M].成都:四川科学技术出版社,2005.[12] 潘道一.除草剂对泽蛙蝌蚪的毒性[J].动物学杂志,1990,25(1):32-34.[13] 耿德贵,张大生,程伟.四种除草剂对中华大蟾蜍蝌蚪红细胞微核及核异常的影响[J].动物学杂志,2000,35(1):12-16.[14] 杨再福.铜和汞对蝌蚪联合毒性的影响[J].农业环境保护,2001,20(5):370-371.[15] 董爱华,贾秀英,马小梅.镉对蟾蜍的 4种器官乳酸脱氢酶同工酶的影响[J].动物学杂志,2003,38(6):24-27.[16] 贾秀英,董爱华.镉、铅对蟾蜍精巢毒作用的酶学研究[J].生态学报,2004,24(10):2 329-2 333.[17] 贾秀英,董爱华,杨亚琴.铜、锌和三唑磷对泽蛙蝌蚪的毒性研究[J].环境科学研究,2005,18(5):26-29.[18] 耿宝荣,姚丹,纪冰,等.重金属铬离子对两种无尾两栖类蝌蚪的急性毒性影响 [M].两栖爬行动物学研究,第 10辑,长春:吉林人民出版社,2005:133-139.[19] 赵东芹,宋刚,张迎梅,等.Pb2+、Cd2+、Cu2+生物积累以及对花背蟾蜍蝌蚪发育的影响 [M]//两栖爬行动物学研究,第 10辑.长春:吉林人民出版社,2005:203-211.[20] 卢祥云,张燕萍,吴海东,等.汞离子和铜离子对中华大蟾蜍蝌蚪联合毒性研究[J].四川动物,2006,25(2):379-381.[21] 蔡玲玲,汪美贞,贾秀英.Cu2+Cd2+及其复合污染对中华大蟾蜍蝌蚪 DNA的影响[J].农业环境科学学报,2007,26(4):1 361-1 364.[22] Herkovits J,Perez-Coll C,Herkovits F D.Ecotoxicological studies of environmentalsamples from BuenosAires area using a standardized amphibian embryo toxicitytest(AMPH ITOX)[J].Environ Pollut,2002,116(1):177-183.[23] 汪松,谢彼德,解焱主编.保护中国的生物多样性 (二)[M].北京:中国环境科学出版社,2001.[24] KieseckerJ M,BlausteinA R,Miller C L.Potentialmechanism underlying the displacementof native red-legged frogs by introduced bullfrogs[J].Ecology,2001,82(7):1 964-1 970.[25] 武正军,王彦平,李义明.浙江东部牛蛙的自然种群及潜在危害[J].生物多样性,2004,12(4):441-446.。
蛙携带虹彩病毒的分子流行病学研究

蛙携带虹彩病毒的分子流行病学研究邵军辉;张利峰;王树云;李聪;罗杨志;陈孝煊;袁军法【摘要】The multiplex real-time PCR assay was used to investigate the prevalence of iridoviruses in farmed or wild frog. Results indicated that the average positive rate was 8.90%,in which the average positive rate in farmed and wild frog was 4.57% and 16.82%,respectively. The most significant prevalence of iridoviruses was found among the farmed Quasipaa spinosa and Polypedates megacephalus,while the species of Rana catesbeiana and Rana nigromaculata rarely. Sequences alignement of major capsid protein suggested that there were no obvious host and geographic specificity among these detected Rana iri-doviruses,it indicated that the outburst of iridoviruses in farm produced negative effects on wild frogs,disease monitoring,dis-ease prevention and control measures were proposed to introduce into the artificial domestication propagation system of wild animal.%运用多重实时PCR对多地的养殖和野生蛙类携带虹彩病毒的本底展开调查,并进行病毒分型.结果表明,检测的样品平均阳性率为8.90%,养殖蛙的平均阳性率为4.57%,野生蛙的平均阳性率为16.82%.从蛙种类来看,养殖牛蛙和黑斑蛙带毒率较低,而养殖的棘胸蛙和野外捕捉的斑腿树蛙带毒率较高.不同蛙携带的蛙病毒的分子序列比较结果显示,不同来源的蛙病毒高度保守,缺乏明显的宿主与地域特异性,表明人工繁育场暴发虹彩病毒可能对其他野生蛙类资源产生了负面影响,建议在野生动物人工驯养繁殖体系中引入病害监测和疫病防控措施.【期刊名称】《湖北农业科学》【年(卷),期】2017(056)009【总页数】4页(P1702-1705)【关键词】蛙;虹彩病毒;多重实时PCR;流行病学【作者】邵军辉;张利峰;王树云;李聪;罗杨志;陈孝煊;袁军法【作者单位】华中农业大学水产学院,武汉 430070;北京出入境检验检疫局检验检疫技术中心,北京 100026;北京出入境检验检疫局检验检疫技术中心,北京 100026;华中农业大学水产学院,武汉 430070;华中农业大学水产学院,武汉 430070;华中农业大学水产学院,武汉 430070;华中农业大学水产学院,武汉 430070【正文语种】中文【中图分类】Q959.5+3近50年来,全球范围内多种两栖动物种群显著衰退,一些种类已经灭绝,部分物种濒危,原因包括气候变化、栖息地减少、环境污染以及疾病传播等[1,2]。
自治区级实验教学示范中心

自治区级实验教学示范中心申请书学校名称:广西师范学院中心名称:地理实验教学中心中心网址:http:/syz x/中心联系电话:(0771)联系人姓名:胡宝清联系手机:中心通讯地址:175号申报日期:2007年6月申报类型:示范中心广西壮族自治区教育厅制目录1、实验教学中心总体情况.................................... 错误!未定义书签。
2、实验教学 (6)3.实验队伍 (10)4.体制与管理 (12)5. 设备与环境 (13)6. 特色 (14)7. 实验教学效果与成果 (15)8.自我评价及发展规划 (18)9.学校意见 (17)附件1 近两年承担的自治区级(省部级)以上科研项目一览表附件2 厅局级以上获奖成果(含教学成果奖)一览表附件3 代表性论文论著(专利)一览表附件4:主要实验课程附件5:主要仪器设备清单1、实验教学中心总体情况中心人员简表2、实验教学3.实验队伍4.体制与管理5. 设备与环境6. 特色7. 实验教学效果与成果8.自我评价及发展规划9.学校意见附件1 近两年承担的自治区级(省部级)以上科研项目一览表附件2 厅局级以上获奖成果(含教学成果奖)一览表附件3 代表性论文论著(专利)一览表此表按照重要性选择论文,总数要求不超过30篇附件4:主要实验课程一、《地图学》实验教学大纲二、《自然地理》实验教学大纲四、《遥感概论》实验教学大纲五、《C语言》实验教学大纲六、《测量学》实验教学大纲七、《计算机网络》实验教学大纲八、《面向对象程序设计》实验教学大纲(段炼)九、《数据库技术》实验教学大纲十、《GIS设计与应用》实验教学大纲十一、《遥感软件应用》实验教学大纲十二、《GIS应用》实验教学大纲十三、《计算机辅助设计》实验教学大纲十四、《计算机制图》实验教学大纲(邓兴礼)十六、《GPS原理与应用》实验教学大纲十七、《数字图像处理》实验教学大纲十八、《网络GIS》实验教学大纲十九、《软件工程》实验教学大纲二十、《网页设计》实验教学大纲二十一、《土地规划学》实验教学大纲二十二、《地球概论》实验教学大纲附件5:主要仪器设备清单。
广西北部湾海洋弧菌多样性研究进展

广西北部湾海洋弧菌多样性研究进展覃欣怡;赵华显;杨恕;黄炯清;廖能健;李晓丽;姜宫凌侠;李楠【期刊名称】《现代食品科技》【年(卷),期】2024(40)1【摘要】海洋弧菌在生物地球化学循环中发挥重要作用,但部分致病性海洋弧菌可危害人类健康和造成水产养殖业重大经济损失。
近年来,广西北部湾海洋弧菌多样性研究取得了众多科研成果。
然而,关于广西北部湾海洋弧菌多样性系统总结的报道仍较为缺乏。
为了更好地了解广西北部湾海洋弧菌物种组成、群落分布特点及其驱动机制,该文从广西北部湾海洋弧菌群落结构特征、环境驱动因子、群落构建机制等方面对广西北部湾海洋弧菌多样性研究进行综述。
结果表明,广西北部湾海洋弧菌多样性在不同海域和季节存在显著差异,其主要驱动因子为总溶解氮、溶解性无机氮、总溶解磷等营养盐。
广西北部湾海洋弧菌群落构建过程由随机过程主导。
此外,该研究对广西北部湾海洋弧菌多样性研究前景进行了展望,以期为致病性海洋弧菌防控提供新思路,并为海洋弧菌生态功能多样性研究及生物资源高值化利用提供理论参考。
【总页数】7页(P325-331)【作者】覃欣怡;赵华显;杨恕;黄炯清;廖能健;李晓丽;姜宫凌侠;李楠【作者单位】南宁师范大学北部湾环境演变与资源利用教育部重点实验室;鲁东大学农学院;桂林理工大学环境科学与工程学院;广东海洋大学近海海洋变化与灾害预警实验室、资源与环境重点实验室【正文语种】中文【中图分类】S94【相关文献】1.北部湾(广西海域)海洋微生物多样性研究现状与对策2.北部湾海洋微生物物种多样性与化学多样性研究进展3.广西北部湾局部海域海洋沉积物细菌多样性及生物活性评估4.北部湾广西海域海洋真菌多样性及其生物活性研究因版权原因,仅展示原文概要,查看原文内容请购买。
实验类型及其具体内容
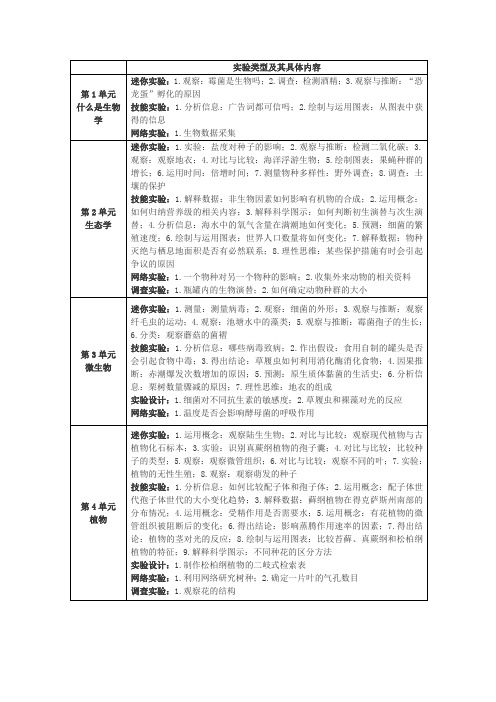
实验类型及其具体内容第1单元什么是生物学迷你实验:1.观察:霉菌是生物吗;2.调查:检测酒精;3.观察与推断:“恐龙蛋”孵化的原因技能实验:1.分析信息:广告词都可信吗;2.绘制与运用图表:从图表中获得的信息网络实验:1.生物数据采集第2单元生态学迷你实验:1.实验:盐度对种子的影响;2.观察与推断:检测二氧化碳;3.观察:观察地衣;4.对比与比较:海洋浮游生物;5.绘制图表:果蝇种群的增长;6.运用时间:倍增时间;7.测量物种多样性:野外调查;8.调查:土壤的保护技能实验:1.解释数据:非生物因素如何影响有机物的合成;2.运用概念:如何归纳营养级的相关内容;3.解释科学图示:如何判断初生演替与次生演替;4.分析信息:海水中的氧气含量在满潮地如何变化;5.预测:细菌的繁殖速度;6.绘制与运用图表:世界人口数量将如何变化;7.解释数据:物种灭绝与栖息地面积是否有必然联系;8.理性思维:某些保护措施有时会引起争议的原因网络实验:1.一个物种对另一个物种的影响;2.收集外来动物的相关资料调查实验:1.瓶罐内的生物演替;2.如何确定动物种群的大小第3单元微生物迷你实验:1.测量:测量病毒;2.观察:细菌的外形;3.观察与推断:观察纤毛虫的运动;4.观察:池塘水中的藻类;5.观察与推断:霉菌孢子的生长;6.分类:观察蘑菇的菌褶技能实验:1.分析信息:哪些病毒致病;2.作出假设:食用自制的罐头是否会引起食物中毒;3.得出结论:草履虫如何利用消化酶消化食物;4.因果推断:赤潮爆发次数增加的原因;5.预测:原生质体黏菌的生活史;6.分析信息:栗树数量骤减的原因;7.理性思维:地衣的组成实验设计:1.细菌对不同抗生素的敏感度;2.草履虫和裸藻对光的反应网络实验:1.温度是否会影响酵母菌的呼吸作用第4单元植物迷你实验:1.运用概念:观察陆生生物;2.对比与比较:观察现代植物与古植物化石标本;3.实验:识别真蕨纲植物的孢子囊;4.对比与比较:比较种子的类型;5.观察:观察微管组织;6.对比与比较:观察不同的叶;7.实验:植物的无性生殖;8.观察:观察萌发的种子技能实验:1.分析信息:如何比较配子体和孢子体;2.运用概念:配子体世代孢子体世代的大小变化趋势;3.解释数据:藓纲植物在得克萨斯州南部的分布情况;4.运用概念:受精作用是否需要水;5.运用概念:有花植物的微管组织被阻断后的变化;6.得出结论:影响蒸腾作用速率的因素;7.得出结论:植物的茎对光的反应;8.绘制与运用图表:比较苔藓、真蕨纲和松柏纲植物的特征;9.解释科学图示:不同种花的区分方法实验设计:1.制作松柏纲植物的二岐式检索表网络实验:1.利用网络研究树种;2.确定一片叶的气孔数目调查实验:1.观察花的结构第5单元分子与细胞迷你实验:1.实验:测定pH;2.运用概念:研究扩散的速度;3.测量:测量显微镜下的物体;4.实验:观察细胞器;5.建构模型:制作细胞膜模型;6.对比与比较:观察中心体;7.实验:分离色素;8.建构模型:用同位素来研究光合作用;9.预测:苹果汁发酵了吗技能实验:1.解释科学图示:原子核的研究发现;2.运用数学工具:冰能浮在水面的原因;3.因果推断:细胞膜是否为选择性屏障;4.解释数据:哪一细胞器指导细胞的生命活动;5.得出结论:当细胞体积增加时,其表面积的变化情况;6.观察与推断:细胞周期长短的差异;7.解释数据:癌症发病率的差异;8.因果推断:脂肪能贮存能量的原因;9.获取信息:有氧呼吸与无氧呼吸实验设计:1.温度对酶促反应的影响网络实验:1.研究光合作用的影响因素调查实验:1.观察和比较不同类型的细胞;2.洋葱根尖细胞的有丝分裂过程第6单元遗传学迷你实验:1.观察与推断:测算花粉粒的数目;2.建构模型:模拟染色体交叉;3.预测:转录和翻译;4.绘制图表:基因突变与蛋白质;5.分析信息:家系图;6.观察与推断:探索眼睛颜色的遗传模式;7.运用概念:将限制性内切酶与内切位点配对;8.运用数学工具:人类基因组数据信息技能实验:1.分析信息:孟德尔实验的数据分析;2.解释科学图示:同源染色体的辨别;3.解释数据:DNA的化学分析结果;4.构建模型:编码一种氨基酸所需的碱基数;5.绘制与运用图表:导致镰刀型细胞贫血征得突变类型6.运用概念:什么是几率;7.预测:兔毛颜色遗传的模式;8.得出结论:杜氏肌营养不良症的遗传模式;9.实验设计:何时需要测交;10.理性思维:如何确定基因是否转移成功;11.运用概念:如何通过DNA指纹进行身份鉴定实施设计:1.细胞质遗传网络实验:1.如何确定植物的表现型和基因型调查实验:1.RNA的转录;2.模拟DNA重组技术第7单元生命的演化迷你实验:1.观察与推断:海洋生物化石;2.整理数据:时间轴;3.建构模型:伪装的适应性特征;4.收集数据:种群的变异;5.推断:人类大拇指的作用;6.分析信息:人类与其他灵长类动物的蛋白质比较;7.分类:使用二岐式检索表;8.分类:利用演化树表示生物的亲缘关系技能实验:1.理性思维:蕨类植物是否曾在北极生长;2.解释数据:时钟能否用来模拟地球历史;3.解释数据:自然选择的观察方法;4.运用数学工具:幼年和成年灵长类动物的差异;5.运用概念:尼安德塔人与现代人类的相似程度;6.得出结论:红狼是否属于一个独立的物种;7.运用图表:六界中每界所含的物种数网络实验:1.自然选择与等位基因频率调查实验:1.测定岩石的年龄;2.三种灵长类动物的颅骨比较;3.制作甲虫的二岐式检索表第8单元无脊椎动物迷你实验:1.观察与推断:观察动物的特征;2.观察与推断:观察醋虫;3.观察:观察水螅摄食;4.观察:观察旋毛虫的幼虫;5.对比与比较:辨认贝壳的种类;6.解释科学图示:蚯蚓的解剖结构;7.对比与比较:龙虾的特征;8.对比与比较:比较不同的变态类型;9.观察与推断:观察叉棘的结构;10.观察:观察文昌鱼技能实验:1.解释科学图示:第一次细胞分裂对青蛙发育的重要性;2.分类:对称型与动物的其他性状的关系;3.运用概念:海生海绵的种类比淡水海绵多的原因;4.解释数据:限制珊瑚物种数量的原因;5.预测:地球上最早出现的扁形动物;6.解释数据:线性动物能否消灭植物的象鼻虫害;7.观察与推断:淡水蚌如何繁殖;8.分类:环节动物的特征;9.运用数学工具:地球上节肢动物的种类;10.实验设计:促使海参向水中排放配子的因素;11.解释科学图示:观察无脊椎脊索动物的切片实验设计:1.蚯蚓对外界环境变化作出反应;2.盐度对丰年虫卵孵化的影响网络实验:1.斑马鱼的发育调查实验:1.观察涡虫的再生;2.比较不同的棘皮动物第9单元脊椎动物迷你实验:1.观察与推断:鳃的结构与功能;2.对比与比较:比较蛙和蝌蚪的适应性结构;3.对比与比较:比较羽毛;4.对比与比较:喂鸟;5.对比与比较:牙齿的解剖结构;6.观察:哺乳动物的骨骼;7.实验:测试等足类动物对光的反应;8.实验:拼图游戏技能实验:1.理性思维:侧线系统的重要性;2.分析信息:地球上哪些地区的鸟濒临灭绝;3.分析信息:哪种动物的消化道最长;4.实验设计:冬眠属于先天行为,还是学习行为;5.解释数据:鸟类如何学习鸣叫实验设计:1.哪种蛋的形状最佳网络实验:1.研究育种犬的适应性调查实验:1.制作两栖动物的二岐式检索表;2.蜗牛的行为第10单元人体生理迷你实验:1.对比与比较:观察指纹;2.解释说明:检查肌肉收缩;3.解释数据:评价一碗汤的营养价值;4.观察:比较甲状腺和甲状旁腺组织;5.实验:注意力分散与反应时间;6.分析信息:药品标签;7.实验:测定脉搏;8.实验:检测仿尿液样品中的葡萄糖;9.观察与推断:观察精子、卵细胞和早期的胚胎发育;10.绘制图表:绘制胚胎长度表;11.实验:检测疾病的传播;12.观察与推断:区分血细胞技能实验:1.因果推断:人体内热量集聚时的反应;2.绘制与运用图表:男性与女性骨密度的差异;3.对比与比较:皮肤细胞、骨细胞及肌肉细胞的差异;4.因果推断:胆囊切除后对消化过程的影响;5.运用数学工具:BMI值;6.分析数据:运动对胰岛素分泌和胰高血糖素水平的影响;7.分析说明:巨大的声响对听力是否有害;8.构建模型:不同的药物对突触中神经递质水平的影响;9.解释数据:如何比较吸入和呼出的空气成分;10.分析信息:如何分析心电图;11.运用概念:为什么月经周期不是正好28天;12.解释数据:如何减少先天缺陷;13.实验设计:单纯疱疹病毒的传播过程;14.分析信息:疫苗的作用实验设计:1.疲劳是否会影响比赛成绩;2.药物对水蚤心率的影响网络实验:1.新兴疾病和再兴疾病调查实验:1.淀粉酶对早餐中谷类食品的作用;2.测量呼吸速率;3.根据hCG 水平来判断是否怀孕。
棘腹蛙营养成分及其影响因素分析

棘腹蛙营养成分及其影响因素分析打开文本图片集摘要:本文通过对比四川雅安、湘西、贵州三地棘腹蛙中16种氨基酸的含量,比较呈味氨基酸和必需氨基酸的相对含量,以及野生棘腹蛙与养殖棘腹蛙营养成分的差异,分析影响其肉质的可能因素。
结果表明,生态养殖棘腹蛙不仅可以保证其丰富的营养成分和鲜美程度,还有利于更好地保护生态环境。
关键词:棘腹蛙;氨基酸;生态养殖棘腹蛙(RanaboulengeriGuenther)属于动物界脊索动物门两栖纲无尾目蛙科[1],俗称石蛙,因其叫声为“bangbang”又被称为梆梆鱼,在四川、贵州、江西、甘肃和湖南等省区都有发现[2-7]。
棘腹蛙是国家二级重点保护动物,由于气候变暖、栖息地被破坏,棘腹蛙的种群数量严重下降,棘腹蛙现已经被《中国濒危物种红皮书》列为“易危”物种[8]。
棘腹蛙对生境要求高,一般生活在海拔700m左右,是狭生态幅的动物。
棘腹蛙个体肥硕,弹跳能力强,多生活在缓流中或者溪边的静水洼中。
棘腹蛙四肢和颌部都有深色纹理,瞳孔呈菱形;雄性棘腹蛙前肢发达,背面由长疣和圆疣排列成行,性成熟后胸部布满黑色棘突;雌性棘腹蛙的前肢不发达,背面无长疣粒,胸部没有棘突,腹面白而光滑。
棘腹蛙主要食害虫和一些花果,对生态环境有益。
棘腹蛙是处于食物链中间环节的两栖类动物,既是捕食者又是被捕食者,可以检测和平衡生态系统[9]。
一、棘腹蛙的营养价值和药用价值棘腹蛙可誉为“蛙王”,其体大味美、肉质细腻;用其煮汤色雪白,味道鲜美,营养价值高,实属一道美味佳肴。
在氨基酸成分上,温安祥等[10-11]已经与甲鱼、田鸡进行了对比试验。
棘腹蛙与滋补上品甲鱼、田鸡相比也毫不逊色,棘腹蛙与甲鱼、北京田鸡(田鸡学名黑斑侧褶蛙)、广东田鸡的氨基酸含量相比稍微少一些,但是其氨基酸总量、必需氨基酸和鲜味氨基酸含量都高于北京田鸡,所含的必需氨基酸也比甲鱼多。
棘腹蛙与其他普通肉类的水分、蛋白质和一些重要元素等相比有很大的差异。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
收 稿 日期 :0 70 —6 2 0 —91
基 金 项 目 : 西 博 士 学 位 授 权 点 学 科 建 设 专 项基 金 资 助 项 目( Y2 0 Z 3 ; 西 科 技 攻 关 项 目( 科 改 0 2 0 92 广 XK 0 6 D0 ) 广 桂 2 8 1— ) 通 讯 联 系 人 : 新 民 (9 6 )男 , 西 灵 川 人 , 西 师 范 大 学 教 授 , 士 。E malx i@malo .x u e u c 秦 1 5一 , 广 广 博 — i:mqn i x g n .d .n b
V o1 .26 No.1
2O O8年 3月
M a . 08 r 20
广西 部分 棘 蛙 类 物 种 的分 子 系 统 学研 究
高成伟 , 碧 文 , 新民 张 秦
( 广西师范大学 生命科学学院 , 广西 桂林 5 1 0 ) 4 0 4
摘
要: 以广 西 棘 蛙 类 为研 究 对 象 , 用 P R 方 法 扩 增 线 粒 体 1Sr NA 基 因 和 C t 应 C 6 R y 因 的 DN 片 段 , b基 A 进
行D NA 序 列 测 定 。 以虎 纹 蛙 和 弹 琴 水蛙 为 外 群 , 邻 接 法 ( ) 最 大 简 约 法 ( ) 建 3 棘 蛙 的 系统 发 育 用 NJ和 MP 构 种 关 系 。 结 果 表 明 : 研 究 的棘 蛙 类 可 分 为 棘 胸 蛙 居 群 、 腹 蛙 居 群 和 棘 侧 蛙 居群 3 , 形 态 分 类 结 果相 同 。 所 棘 支 与 关 键 词 : 子 生 物 学 ; 蛙 属 ;6 R 分 棘 1Sr NA 基 因 C t y 因 ; b基 系统 发 育 中 图分 类 号 : 5 . 3 Q9 9 5 文献标识码 : A 文 章 编 号 :0 16 0 (0 8 0— 1 50 1 0— 6 0 2 0 ) 100 —5
1 Sr 6 RNA 和 C t 基 因片段扩 增 引物 的设 计 分别 参考 文献 [ ,] y b 67。
1 Sr RNA 引物 : L 6
H
5_ G C TT C C TG TAC AAAAAC r C AT一 3
5一 CCG G GCT GA A CT CA G A TCA CGT ~ 3
棘 蛙属 于两 栖纲 Amp iin 无 尾 目 An r 、 科 R nd e棘蛙 属 Pa 。 o ln e 根据 棘蛙 物种 的外 hba 、 u a蛙 a ia a B ue g r 形 和骨骼 特 征将其 列 为蛙属 中的蛙 亚属 ; 承 钊等 [对 中国 已知 棘蛙 物 种 的形 态进 行 了描述 , 将其 归 刘 2 ] 并
维普资讯
第2卷 6
第 1 期
广西 师范 大 学学 报 : 自然 科学 版
J un l f a g i r l iest : trl c n eE io o r a o n x ma Unv ri Naua S i c dt n Gu No y e i
维普资讯
16 0
广西师范大学学报 : 自然 科 学 版
ห้องสมุดไป่ตู้
第 2 卷 6
1 2 方 法 .
1 2 1 总 DNA 的提 取 ..
采 用 S S 蛋 白酶 K 提 取法 提取 总 DNA。 D /
1 2 2 1 Sr . . 6 RNA 和 C tb 因 的 P R扩 增 y 基 C
Ct y b引物 :
L1 8 7 5_ AT T AGC GATGAAAC TC G一 4 4 , AC C C AT T G 3
H1 3 52 0 5一 TA CTG A G A A G CCCCCTCA A A TTCA T TG 一 3
1 Sr NA 基 因片段 的 P R 扩增反 应 采用 5 L体积 , 括 : 品 DNA 模 板 1 L、 6 R C 0 包 样 引物 各 1 L( 0 1
无水 乙醇浸 泡保 存 外 , 其余 的 均为 新鲜 蛙腿 部肌 肉 , 置于 一7 0℃保 存 。实验 所用 样 品见表 1 。
表 1 供 试 材 料 名 称 及 来 源
Tab S c e .1 pe i snamesa o a iis o x r m e t m a e il nd l c lte f e pe i n t ra
探讨 广西 棘蛙类 种 间和 种 内的分 类地 位 和亲 缘关 系 , 为棘 蛙类 的分 类 、 化 等提 供分 子 依据 。 进
1 材 料 与方 法
1 1 实验 材料 .
本研 究 所用材 料 中 , 个 种来 自广西 棘蛙属 , 个 种 为外 群 , 5种 蛙 3 个 样 品 。 3 2 共 6 除云 南省 的样 品是用
为真 蛙类 Ra a 法 国学者 D b i[把它 从蛙 属 中分 离 出来 , n; u os3 独立 为一属 P a 该 属 动物 雄性 都具 有不 同的 a。 皮肤 棘而 成为 一个 自然 类 群 。 目前 , 蛙分 为 2 种 , 要分 布 于亚 洲东 南部 亚热 带和 热带 地 区 , 布 区 棘 9个 主 分 东起 我 国沿海 地 区 , 至阿 富 汗 、 西 巴基 斯 坦 , 南至越 南 、 缅甸 , 至喜 马拉 雅 山和秦 岭 。 国共 有 2 北 我 1个种 , 占 全 部 种 的 7 . 1 , 中特 有 种 7个 , 布 在秦 岭 以南 的各 省 区 ( 2 4 其 分 西起 藏 滇 , 至 闽浙 , 至 桂粤 ) 但 海 南 东 南 , 和 台湾没 有分 布 。该属 系统 关 系研 究对 研究 蛙科 两栖 动物 的进 化 和地 理分 布等 都有 着 一定 的科研 价值 。 关于 棘蛙 属 的分 类 和物 种命 名 问题 一 直都 存 在争 议 [ ]本 研 究对 广 西 棘蛙 类 进行 分 子 系 统学 研 究 , 4,