生物化学--第六章 生物氧化(3-4节)
生物化学--生物氧化

脱电子 Fe2+
Fe3+ + e
生物氧化中的CO2的生成
绝大部分有机物生物氧化中的CO2生成是经 ? 中的脱羧作用产生的。
答案:三羧酸循环
其他一些CO2产生途径如: 糖异生
草酰乙酸 + GTP → PEP +GDP + CO2 氨基酸脱羧
NH2
脱羧酶
NH2
R C COOH
R C H + CO2
磷酸烯醇式丙酮 酸羧激酶
COCOOH
GTP
GDP
β-氧化脱羧:
CH2 CO~ P + CO2 COOH
CHOH-COOH CH-COOH CH2-COOH
异柠檬酸脱氢酶
CO-COOH CH2
NAD+
NADH+H+ CH2-COOH
+CO2
生物氧化中H2O的生成
真核生物线粒体内膜上的电子传递链作用下产生
化合物
磷酸烯醇式丙酮酸 氨基甲酰磷酸
kJ/mol -61.9 -51.4
△E0′
(kcal/mol) (-14.8) (-12.3)
1,3-二磷酸甘油酸 磷酸肌酸
ATP →ADP+Pi 乙酰辅酶A
ADP →AMP+Pi 焦磷酸
1-磷酸葡萄糖
-49.3 -43.1 -30.5 -31.5 -27.6 -27.6 -20.9
线粒体结构模式图
二、ATP
NH2
NN
O- OOPγ~- O
OP~β O O-
O Pα O-
O CH2
N O
N
OH OH AM P ADP
ATP
高能磷酸键与高能磷酸化合物
生物化学及分子生物学(人卫第九版)-06-03节生物氧化

本章小结
氧化过程:线粒体基质的NADH和FADH2通过电子传递链进行氧化,产生 CO2、H2O 磷酸化过程:是产生ATP的主要机制,电子传递链在氧化电子的过程中、 泵出质子储存能量、至膜间隙侧而产生跨膜质子电化学梯度,储存电子氧 化释放的能量,形成质子驱动力,促使质子回流至基质释能而产成ATP
一、体内能量状态可调节氧化磷酸化速率
氧化磷酸化是机体合成能量载体ATP的最主要的途径
机体根据能量需求调节氧化磷酸化速率,从而调节ATP的生成量 细胞内ADP的浓度以及ATP/ADP的比值感应机体能量状态的变化 耗能代谢反应活跃时,ATP分解为ADP和Pi的速率增加,使ATP/ADP的比值降 低、ADP的浓度增加,氧化磷酸化速率加快 ATP和ADP也同时调节糖酵解、柠檬酸循环途径,调节NADH和FADH2的生成
抗氧化
抗氧化体系清除ROS
超氧化物歧化酶(superoxide dismutase,SOD)
催化2分子·O2-分别进行氧化和还原,生成O2和H2O2
活性强,是人体防御超氧离子损伤的重要酶
哺乳动物细胞有3 种SOD 同工酶:
Cu/Zn-SOD:胞外、胞质
Mn-SOD:线粒体
过氧化氢酶(catalase)
二羧酸转运蛋白
α -酮戊二酸转运蛋白 天冬氨酸-谷氨酸转运蛋白 单羧酸转运蛋白 三羧酸转运蛋白 碱性氨基酸转运蛋白 肉碱转运蛋白
HPO42苹果酸 谷氨酸 丙酮酸 苹果酸 鸟氨酸 脂酰肉碱
苹果酸
α -酮戊二酸 天冬氨酸 OH柠檬酸 瓜氨酸 肉碱
胞浆NADH的跨膜转运
胞浆NADH需转运至线粒体基质进行氧化
为何生物氧化主要的能量 代谢产物是ATP?
ATP合酶的作用 ATP的作用
人民卫生出版社《生物化学》第六章 生物氧化

⊿Gº’ = -nF ⊿Eº'
n:传递电子数;F:法拉第常数
➢ 合成1摩尔ATP 需能量约30.5kJ
偶联部位
NADH~CoQ CoQ~Cytc Cyta-a3~O2
电位变化 (∆E0')
0.36V 0.21V 0.53V
自由能变化 (∆G0')
69.5KJ/mol 40.5KJ/mol 102.3KJ/mol
三、NADH和FADH2是呼吸链的电子供体
1、NADH氧化呼吸链 NADH →复合体Ⅰ→CoQ →复合体Ⅲ→Cyt c →复合体Ⅳ→O2
2、琥珀酸氧化呼吸链 琥珀酸 →复合体Ⅱ →CoQ →复合体Ⅲ→Cyt c →复合体Ⅳ→O2
呼吸链各组分的排列顺序的实验依据
➢ 标准氧化还原电位 ➢ 特异抑制剂阻断 ➢ 还原状态呼吸链缓慢给氧 ➢ 将呼吸链拆开和重组
生物氧化与体外氧化之不同点
生物氧化
➢ 反应环境温和,酶促反应逐步进 行,能量逐步释放,能量容易捕 获,ATP生成效率高。
体外氧化
➢ 能量突然释放。
➢ 通过加水脱氢反应使物质能间接 获得氧;脱下的氢与氧结合产生 H2O,有机酸脱羧产生CO2。
➢ 物质中的碳和氢直接氧 结合生成CO2和H2O 。
生物氧化的一般过程
胞液侧 4H+
2H+ 4H+ Cyt c
+
+++++ +
++
+
Q
Ⅰ
--
NADH+H+
NAD+
Ⅱ
-
延胡索酸
琥珀酸
Ⅳ
Ⅲ- - -
生物化学教案:第六章 生物氧化

一系列酶促反应逐步进行,能
量逐步释放有利于机体捕获
能量,提高 ATP 生成的效率
通过加水脱氢反应使物
物质中的碳和氢直接氧
质能间接获得氧,并增加脱氢 结合生成 CO2 和 H2O 。 的机会;脱下的氢与氧结合产
生 H2O , 有 机 酸 脱 羧 产 生 CO2。
二、生成 ATP 的氧化磷酸化体系 1、呼吸链
15 mins
教学主要内容
备注
高能磷酸键:水解时释放的能量大于 21KJ/mol 的磷酸酯键,常
表示为 P
高能磷酸化合物即含有高能磷酸键的化合物 5、线粒体内膜对各种物质进行选择性转运
25 mins
线粒体外膜通透性高,线粒体对物质通过的选择性主要依
赖于内膜中不同转运蛋白(transporter)对各种物质的转运。 胞液中 NADH 的氧化 转运机制主要有:
Cyt c1 ,
Cyt a
复 合 细胞色素 162
体Ⅳ
C 氧化酶
13 血红素 a, Cyt c(膜
a3,
间隙侧)
CuA, CuB
排列顺序:(1)NADH 氧化呼吸链 NADH →复合体Ⅰ→Q →复合体Ⅲ→Cyt c →复合体Ⅳ→O2
(2)琥珀酸氧化呼吸链 琥珀酸 →复合体Ⅱ →Q →复合体Ⅲ→Cyt c →复合体Ⅳ→O2
氧化磷酸化偶联机制是产生跨线粒体内膜的质子梯度
偶联机制:化学渗透假说
电子经呼吸链传递时,可将质子(H+)从线粒体内膜的基质侧
泵到内膜胞浆侧,产生膜内外质子电化学梯度储存能量。当质子顺
浓度梯度回流时驱动 ADP 与 Pi 生成 ATP。
质子顺梯度回流释放能量被 ATP 合酶利用催化 ATP 合成
生物化学--第六章 生物氧化(3-4节)

2019/1/4
17
2019/1/4
18
化学渗透假说的内容★★
1.呼吸链中传氢体和电子传递体是间隔交替排列的,且
在线粒体内膜都有特定的位置,催化反应是定向的。
2.内膜对H+不能自由通过,泵出膜外侧的H+不能自由
返回膜内侧,造成电化学梯度
3.复合体Ⅰ、Ⅲ、Ⅳ都有质子泵的作用
4.ATP合酶存在于线粒体内膜上,H+梯度是ATP合成的 驱动力
2019/1/4 24
ATP
四 线粒体 ATP合酶(mitochondrial ATPase) 形成ATP的机理
F1
柄
F0
2019/1/4
25
2019/1/4
26
ATP合酶
Hale Waihona Puke ATP合成酶由 疏水的 F0(a1b2c1012) 和亲水的 F1(33)组 成. 质子穿过a时, 推动c环象水 车一样转动, 连带F1转动.
(二)质子梯度的形成
(三)线粒体 ATP合酶(mitochondrial ATPase)
(四)ATP合成的机制
2019/1/4
16
(一)能量偶联假说 1953年 Edward Slater 化学偶联假说
1964年 Paul Boyer 构象偶联假说 1961年 Peter Mitchell 化学渗透假说
2019/1/4
44
2019/1/4
45
七、有关氧化磷酸化物质的运输
• 胞液中的3-磷酸甘油醛,3-磷酸甘油或乳 酸脱氢,均可产生NADH。 • 这些NADH可经穿梭系统而进入线粒体氧 化磷酸化,产生H2O和ATP。
2019/1/4 46
-磷酸甘油穿梭示意图
呼吸链
6生物化学习题(答案)

6生物化学习题(答案)6 生物氧化一、名词解释1、生物氧化:生物细胞将糖、脂、蛋白质等燃料分子氧化分解,最终生成CO2和H2O并释放出能量的作用。
生物氧化包括:有机碳氧化变成CO2;底物氧化脱氢、氢及电子通过呼吸链传递、分子氧与传递的氢结成水;在有机物被氧化成CO2和H2O的同时,释放的能量使ADP转变成ATP。
2、呼吸链:有机物在生物体内氧化过程中所脱下的氢原子,经过一系列有严格排列顺序的传递体组成的传递体系进行传递,最终与氧结合生成水,这样的电子或氢原子的传递体系称为呼吸链或电子传递链。
电子在逐步的传递过程中释放出能量被用于合成ATP,以作为生物体的能量来源。
3、氧化磷酸化:在底物脱氢被氧化时,电子或氢原子在呼吸链上的传递过程中伴随ADP磷酸化生成ATP的作用,称为氧化磷酸化。
氧化磷酸化是生物体内的糖、脂肪、蛋白质氧化分解合成ATP的主要方式。
4、P/O:电子经过呼吸链的传递作用最终与氧结合生成水,在此过程中所释放的能量用于ADP磷酸化生成ATP。
经此过程消耗一个原子的氧所要消耗的无机磷酸的分子数(也是生成ATP的分子数)称为磷氧比值(P/O)。
如NADH的磷氧比值是3,FADH2的磷氧比值是2。
5、底物水平磷酸化:在底物被氧化的过程中,底物分子内部能量重新分布产生高能磷酸键(或高能硫酯键),由此高能键提供能量使ADP(或GDP)磷酸化生成ATP(或GTP)的过程称为底物水平磷酸化。
此过程与呼吸链的作用无关,以底物水平磷酸化方式只产生少量ATP。
6、能荷:能荷是细胞中高能磷酸状态的一种数量上的衡量,能荷大小可以说明生物体中ATP-ADP-AMP系统的能量状态。
1、真核细胞的呼吸链主要存在于线粒体内膜,而原核细胞的呼吸链存在于细胞质膜。
2、NADH呼吸链中氧化磷酸化的偶联部位是复合体Ⅰ、复合体Ⅲ、复合体Ⅳ。
3、在呼吸链中,氢或电子从电负性较大(氧化还原电位较低)的载体依次向电正性较大(氧化还原电位较高)的载体传递。
基础生物化学第6章

类别:黄素脱氢酶类(如NADH脱氢酶、琥珀酸脱氢酶)
需氧脱氢酶类(如L—氨基酸氧化酶)
加单氧酶(如赖氨酸羟化酶)
铁硫蛋白
特点:含有Fe和对酸不稳定的S原子,Fe和
S常以等摩尔量存在(Fe2S2, Fe4S4 ),构成 Fe—S中心,Fe与蛋白质分子中的4个Cys残 基的巯基与蛋白质相连结。 +e
NO2
• 离子载体
短杆菌肽等
• 解偶联蛋白
NADH 鱼藤酮 安密妥 FMN Fe-S
电 子 传 递 抑 制 剂
复合物 I
琥珀酸
FMN
Fe-S
CoQ
Cyt b
复合物 II
抗霉素A
复合物 III
Fe-S Cyt c1 Cyt c Cyt aa3
氰化物 CO
复合物 IV
O2
化学渗透假说示意图
+++++++++
电 子 传 递 链 中 各 中 间 体 的 顺 序
NADH
FMN
复合物 I
NADH 脱氢酶
Fe-S
琥珀酸等
FMN
Fe-S
CoQ
复合物 II
琥珀酸-辅酶Q 还原酶
Cyt b
复合物 III
Fe-S
辅酶Q-细胞色素 还原酶
Cyt c1
Cyt c
复合物 IV
Cyt aa3
细胞色素C 还原酶
O2
呼吸链中电子传递时自由能的下降
NADH呼吸链
还原型代 谢底物
MH2
NAD+
FMNH2
CoQ
2Fe2+
细胞色素
06生物化学习题与解析--生物氧化

06⽣物化学习题与解析--⽣物氧化⽣物氧化⼀、选择题A 型题1 .下列对呼吸链的叙述不正确的是A .复合体Ⅲ和Ⅳ为两条呼吸链所共有B .呼吸链中复合体Ⅰ、Ⅲ、Ⅳ有质⼦泵功能C .递氢体也必然递电⼦D .除 Cytaa 3 外,其余细胞⾊素都是单纯的递电⼦体E . Cyta 和 Cyta 3 结合较紧密2 .⼈体内⽣成 ATP 的主要途径是A .三羧酸循环B .氧化C .氧化磷酸化D .底物⽔平磷酸化E .糖酵解3 .呼吸链存在的部位是A .胞浆B .线粒体内膜C .线粒体内D .线粒体外膜E .细胞膜4 .细胞⾊素 C 氧化酶含有下列哪种⾦属元素A .鋅B .镁C .钙D .酮E .钼5 .下列哪种酶中含有硒元素A .乳酸脱氢酶B .⾕胱⽢肽过氧化物酶C .细胞⾊素 C 氧化酶D .过氧化氢酶A .两条呼吸链排列在线粒体外膜上B .两条呼吸链都含有复合体ⅡC .解偶联后,呼吸链就不能传递电⼦了D .通过呼吸链传递 1 个氢原⼦都可能⽣成 2.5 分⼦ ATPE .两条呼吸链的汇合点是辅酶 Q7 .能直接与氧结合的细胞⾊素类是A . CytbB . CytcC . Cytc 1D . Cytaa 3E . CytP 4508 .在线粒体内 NADH 进⾏氧化磷酸化的 P/O ⽐值为A. 1B. 1.5 C . 2.5 D. 4 E. 59 .电⼦按下列各式传递时能偶联磷酸化的是A .Cytc→Cytaa 3B .CoQ→CytbC .Cytaa 3 →1/2O 2D .琥珀酸→ FAD E. 以上都不是10 .关于化学渗透假说叙述错误的是A .必须把线粒体内膜外侧的 H + 通过呼吸链泵到内膜来B .需在线粒体内膜两侧形成电位差C .质⼦泵的作⽤在于存储能量D .由英国学者 Mitchell 提出E . H + 顺浓度梯度由膜外回流时驱动 ATP 的⽣成11 下列代谢物经过⼀种酶催化后脱下的 2H 不能经过 NADH 呼吸链氧化的是A . CH 3 CH 2 CH 2 CO~SCoAB .异柠檬酸C .α- 酮戊⼆酸D . HOOC-CHOH-CH 2 -COOHE . CH 3 -CO-COOH12 .影响氧化磷酸化的激素是A .胰岛素D .胰⾼⾎糖素E .肾上腺⽪质激素13 . NADH 和 NADPH 中含有共同的维⽣素是A . VitB 1 B . VitB 2C . VitPPD . VitB 12E . VitB 614 .体内能量存储的主要形式是A . ATPB . CTPC . ADPD .肌酸E .磷酸激酸15 .下列化合物中哪⼀个不是⾼能化合物A .⼄酰 CoAB .琥珀酰 CoAC . AMPD .磷酸激酸E .磷酸烯醇式丙酮酸16 .体内 CO 2 来⾃A .碳原⼦被氧原⼦氧化B .呼吸链的氧化还原过程C .有机酸脱羧D .脂肪分解E .糖原分解17 .苹果酸穿梭系统需要下列哪种氨基酸参与A . GlnB . AspC . AlaD . LysE . Val18 .肌⾁中能量的主要存储形式是C . UTPD . CTPE .磷酸肌酸19 .氰化物中毒是由于它抑制了A . CytbB . CytcC . CytP 450D . Cytaa 3E . Fe-S20 .下述各酶催化的反应与 H 2 O 2 有关,但例外的是A .⾕胱⽢肽过氧化物B .触酶C . SOD D .黄嘌呤氧化酶E .混合功能氧化酶(⼆)B 型选择题A. Vit-PPB. Vit-B 12 C . Fe-S D. ⾎红素 E. 苯醌结构1. CoQ 分⼦中含有2. NAD + 分⼦中含有A. 核醇B. 铁硫蛋⽩C. 苯醌结构D. 铁卟啉类E. 异咯嗪环3. CoQ 能传递氢是因为分⼦中含有4. FAD 传递氢其分⼦中的功能部分是A. F 1B. F 0 C . α- 亚基 D. OSCP E. β- 亚基5. 能与寡酶素结合的是6. 质⼦通道是A. NAD + /NADHB. NADP + /NADPHE. CoQ/ CoQH 27. 物质氧化时,⽣成 ATP 数的依据是8. 调节氧化磷酸化运转速率的主要因素是A. CH 3 -CO-S~ CoAB. PEPC. CPD. GTPE. 1,3- ⼆磷酸⽢油酸9. 上述化合物不含⾼能磷酸键的是10. 属于磷酸酐的物质是11. 属于混合酸酐的物质是(三) X 型题1. ⽣物氧化的特点是A. 反应条件温和B. 有酶参加的酶促反应C. 能量逐步释放D. 不需要氧E. 在细胞内进⾏2. 脱氢( 2H )进⼊琥珀酸氧化呼吸链的物质是A. 琥珀酸B. β- 羟丁酸C. 线粒体内的α- 磷酸⽢油D. HOOC-CH 2 -CH 2 -COOH3. 以 NAD + 为辅酶的脱氢酶有A. α- 磷酸⽢油醛脱氢酶B. 异柠檬酸脱氢酶C. 琥珀酸脱氢酶D. 苹果酸脱氢酶E. 脂酰 CoA 脱氢酶4. 琥珀酸氧化呼吸链和 NADH 氧化呼吸链的共同组成部分是A. NADHB. 琥珀酸C. CoQ5. 下列含有⾼能键的物质有A. ATPB. AMPC. ⼄酰 CoAD. 磷酸肌酸E. 琥珀酰 CoA6. 氧化磷酸化偶联部位有A. NADH→CoQB. CoQ→Cyt b , cC. Cy t c→Cyt aa 3D. Cyt aa 3 → O 2E. FAD→CoQ7. 琥珀酸氧化呼吸链中氢原⼦或电⼦的传递顺序为A. 琥珀酸→FADB. FMN→CoQ→CytC. FAD→CoQD. b→c 1 →c→aa 3E. FAD→Cyt b8. 下列每组内有两种物质,都能抑制呼吸链同⼀个传递步骤的是A. 粉蝶霉素 A 和鱼藤酮B. BAL 和寡霉素C. DNP 和 COD. H 2 S 和 KCNE. 异戊巴⽐妥和 CO9. 脱氢需经过α- 磷酸⽢油穿梭系统的物质有A. 琥珀酸B. CH 3 -CHOH-COOHC. 3- 磷酸⽢油醛D. 柠檬酸E. 丙酮酸10. 线粒体内可以进⾏的代谢是A. 三羧酸循环B. 氧化磷酸化E. 酮体的合成11. ⽣物氧化中 CO 2 的⽣成⽅式有A. α- 单纯脱羧B. α- 氧化脱羧C. β- 单纯脱羧D. β- 氧化脱羧E. 以上都是12. 体内清除 H 2 O 2 的酶有A. 过氧化氢酶B. SODC. 过氧化物酶D. 加双氧酶E. 单加氧酶⼆、是⾮题1.NAD + 在呼吸链中传递两个氢原⼦。
第六章 生物氧化

第六章生物氧化Biological Oxidation一、授课章节及主要内容:第六章生物氧化二、授课对象:临床医学、预防、法医(五年制)、临床医学(七年制)三、授课学时本章共4节课时(每个课时为45分钟)。
讲授安排如下:第一次课(2学时):第一节生成A TP的氧化体系——氧化磷酸化偶联部位第二次课(2学时):影响氧化磷酸化的偶联机理——第二节其他氧化体系四、教学目的与要求生物氧化、呼吸链和氧化磷酸化的定义; ATP生成的方式;氧化磷酸化的过程。
五、重点与难点重点:1.主要是生成ATP的氧化体系;2.呼吸链电子传递的过程;3.ATP生成的方式;4.A TP的利用和储存形式;5.胞浆NADH+H+的氧化。
难点:氧化磷酸化的偶联机理六、教学方法及授课大致安排以面授为主,适当结合临床提问启发。
每次课预留5分钟小结本次课掌握内容及预留复习题,全章结束后小结本章内容。
七、主要外文专业词汇biological oxidation (生物氧化) electron transfer chain (电子传递链)respiratory chain (呼吸链) NAD+ (氧化型烟酰胺腺嘌呤二核苷酸)oxidative phosphorylation (氧化磷酸化) α-glycerophosphate shuttle (α-磷酸甘油穿梭)uncoupler (解偶联剂) CoQ (辅酶Q)malate-asparate shuttle (苹果酸-天冬氨酸穿梭) superoxide dismutase(SOD) (超氧物歧化酶) catalase (过氧化氢酶) FMN (黄素腺嘌呤单核苷酸)mixed-function oxidase (混合功能氧化酶) creatine phosphate (磷酸肌酸)ATP synthase (ATP合酶) FAD (黄素腺嘌呤二核苷酸)chemiosmotic hypothesis (化学渗透假说) peroxidase (过氧化物酶)cytochrome (细胞色素) NADP+ (氧化型烟酰胺腺嘌呤二核苷酸磷酸)八、思考题1.何为生物氧化?有何特点?2.试述呼吸链的定义,体内有哪两条呼吸链?3.试写出两条呼吸链组分的排列次序和ATP的生成部位。
第六章 代谢与生物氧化
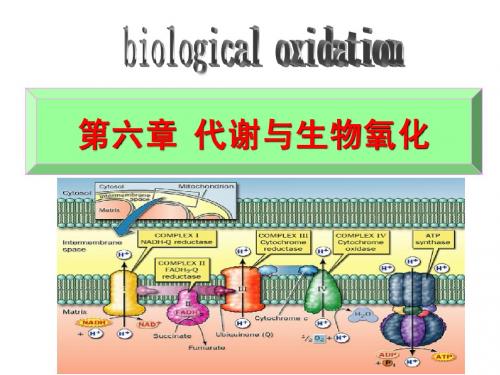
一、新陈代谢
第 六 章 代 谢 与 生 物 氧 化 4. 代谢
——完成某一代谢过程的一组相互衔接的酶 完成某一代谢过程的一组相互衔接的酶 促反应。 促反应。 特点: 特点:
没有完全可逆的代谢途径; 没有完全可逆的代谢途径; 的代谢途径 代谢途径形式是多样 形式是多样的 代谢途径形式是多样的; 代谢途径有确定的细胞定位 确定的细胞定位; 代谢途径有确定的细胞定位; 代谢途径是相互沟通的; 代谢途径是相互沟通 相互沟通的 能量关联; 代谢途径之间有能量关联 代谢途径之间有能量关联; 代谢途径的流量可调控 可调控。 代谢途径的流量可调控。
能
在高能化合物分子中, 在高能化合物分子中 , 被水解断裂时释放出大量 自由能的活泼共价键。 高能键常用符号“ 表示 表示。 自由能的活泼共价键。 高能键常用符号“ ~ ”表示。
“高能键”≠“键能高” 高能键” 高能键 键能高”
茶学与生物系-生物化学
代谢中的能量物质
第 六 章
根据分子结构和高能键的特征,高能化合物可分为: 根据分子结构和高能键的特征,高能化合物可分为: 焦磷酸化合物: (1) 焦磷酸化合物:如ATP
(C~S)型 型
茶学与生物系-生物化学
二、生物氧化
第 六 章 代 谢 与 生 物 氧 化 1.定义 定义
糖类、脂肪、 糖类、脂肪、蛋白质等有机物质在细胞中进行脱 加氧等氧化分解生成CO2和H2O,并释放出能量 氢、加氧等氧化分解生成 , 的过程称为生物氧化 生物氧化(biological oxidation)。 的过程称为生物氧化 。 其实质是需氧细胞在呼吸代谢过程中所进行的一 系列氧化还原反应过程,故又可称细胞呼吸 细胞呼吸。 系列氧化还原反应过程,故又可称细胞呼吸。
• 新陈代谢 一 新陈代谢一 物质 和 能量 转变
生物化学第六章生物氧化

(还原剂) (氧化剂)
可写成 A2+ B3+
A3+
B2+
2019/11/23
生物化学教研室
9
第三节 生成ATP的氧化体系
一、呼吸链的概念
代谢物脱下的成对氢原子(2H)通过多种酶和辅酶所 催化的连锁反应逐步传递,最终与氧结合生成水。由 于此过程与细胞呼吸有关,所以将传递链称为呼吸链, 也叫电子传递呼吸链。
氧化酶,而其它均为不需氧脱氢酶。其中a与 a3很难分开,常写为aa3。
在微粒体中主要为细胞色素b5、p450。p450作用 与aa3类似 。
2019/11/23
生物化学教研室
19
细胞色素的结构
2019/11/23
生物化学教研室
20
呼吸链复合体
人线粒体呼吸链通过上述5大类成分形成4个复合体。
2019/11/23
P/O比值:每消耗1摩尔原子氧所消耗的无机磷 原子的摩尔数。
2019/11/23
生物化学教研室
39
2、氧化磷酸化的偶联机制
内模胞浆侧
化 学 渗 透 学 说内膜基侧2019/11/23
生物化学教研室
40
ATP合酶(复合体Ⅴ)
由F1和F0组成。 F1 在线粒体内膜基质 侧形成颗粒状突起, 催化ATP的生成。 F膜0镶中嵌。在当线H+粒顺体浓内度 梯度经回流时,γ 亚基发生旋转,3个 β 亚基构象变化, 由紧密结合型变为 开放型,释放ATP。
根据呼吸链各组分的标准氧化还原电位测定(电位越 低越容易失去电子)、利用呼吸链特异性的阻断剂测 定其氧化和还原状态的吸收光谱及离体线粒体各组分 的氧化顺序等实验,确定了呼吸链各组分的排列顺序, 并发现体内存在两条主要的呼吸链。
《生物化学原理》张洪渊主编 课后习题及答案(二)

参考答案 (一)名词解释
1.代谢物分子中的氢原子在脱氢酶作用下激活脱落后,经过一系列传递体的传递,最终 将电子交给被氧化酶激活的氧而生成水的全部体系,称为呼吸链或电子传递链。
2.伴随着呼吸链电子传递过程发生的 ATP 的合成称为氧化磷酸化。氧化磷酸化是生物体 内的糖、脂肪、蛋白质氧化分解,并合成 ATP 的主要方式。
从中获得多少能量(多少 ATP)?(单位用 mol)
3.1mol 乳酸完全氧化可生成多少摩尔 ATP?每生成 1molATP 若能储能 30.54kJ 计算,其储能
效率为多少?如果 2mol 乳酸转化成 1mol 葡萄糖,需要消耗多少 ATP?(单位用 mol) 4.用 14C 标记葡萄糖的第三碳原子,将这种 14C 标记的葡萄糖在无氧条件下与肝匀浆保温,那
www.khd课后a答w案.网com
一、课后习题
第六章 生物氧化
1.用对或不对回答下列问题。如果不对,请说明原因。
(1)不需氧脱氢酶就是在整个代谢途径中并不需要氧参加的生物氧化酶类。
(2)需氧黄酶和氧化酶都是以氧为直接受体。
(3)ATP 是所有生物共有的能量储存物质。 (4)无论线粒体内或外,NADH+H+用于能量生成,均可产生 2.5 个 ATP。
1.(1)有机物在生物体内氧化过程中所脱下的氢原子,经过一系列有严格排列顺序的传 递体组成的传递体系进行传递,最终与氧结合生成水,这样的电子或氢原子的传递体系称为 呼吸链或电子传递链。(2)线粒体生物氧化体系中,两类典型的呼吸链都由五类组分组成, 并按一定的顺序定位于线粒体内膜。NADH 呼吸链由 NADH 还原酶(复合体Ⅰ)、泛醌、细胞
(3)乙醛+延胡索酸→乙酸+琥珀酸
(4)丙酮酸+β-羟丁酸→乳酸+乙酰乙酸
生物化学试题及答案

生物化学试题及答案 IMB standardization office【IMB 5AB- IMBK 08- IMB 2C】物化学试题及答案(6)第六章生物氧化【测试题】一、名词解释1.生物氧化2.呼吸链3.氧化磷酸化4. P/O比值5.解偶联剂6.高能化合物7.细胞色素8.混合功能氧化酶二、填空题9.琥珀酸呼吸链的组成成分有____、____、____、____、____。
10.在NADH 氧化呼吸链中,氧化磷酸化偶联部位分别是____、____、____,此三处释放的能量均超过____KJ。
11.胞液中的NADH+H+通过____和____两种穿梭机制进入线粒体,并可进入____氧化呼吸链或___呼吸链,可分别产生____分子ATP或____分子ATP。
12.ATP生成的主要方式有____和____。
13.体内可消除过氧化氢的酶有____、____和____。
14.胞液中α-磷酸甘油脱氢酶的辅酶是____,线粒体中α-磷酸甘油脱氢酶的辅基是____。
15.铁硫簇主要有____和____两种组成形式,通过其中的铁原子与铁硫蛋白中的____相连接。
16.呼吸链中未参与形成复合体的两种游离成分是____和____。
17.FMN或FAD作为递氢体,其发挥功能的结构是____。
18.参与呼吸链构成的细胞色素有____、____、____、____、____、____。
19.呼吸链中含有铜原子的细胞色素是____。
20.构成呼吸链的四种复合体中,具有质子泵作用的是____、____、____。
21.ATP合酶由____和____两部分组成,具有质子通道功能的是____,____具有催化生成ATP的作用。
22.呼吸链抑制剂中,____、____、____可与复合体Ⅰ结合,____、____可抑制复合体Ⅲ,可抑制细胞色素c氧化酶的物质有____、____、____。
23.因辅基不同,存在于胞液中SOD为____,存在于线粒体中的 SOD为____,两者均可消除体内产生的____。
生物氧化与氧化磷酸化

例如,物质 A 转变为物质 B 的反应:
A
B
△G = GB-GA 当△G 为正值时,反应是吸能的,不能自发进行,必须从外界获得能量才能被动进行,
192
但其逆反应则是自发的;当△G 是负值时,反应是放能的,能自发进行,自发反应进行的
推动力与自由能的降低成正比。一个物质所含的自由能越少就越稳定。由此可见△G 值的
(oxidation-reduction potentials)相对地表示各种化合物对电子亲和力的大小。
生物体内任何的氧化还原物质连在一起,都可以有氧化-还原电位产生。生物体内许多
重要的生化物质氧化-还原体系的氧化-还原电位已经测出,其数据见表 6-1。
因为氧化还原电位较高的体系,其氧化能力较强;反之,氧化还原电位较低的体系,
可以看出,△G 不但取决于反应物和产物的化学结构,还取决于它们的浓度,因为浓 度决定反应的方向。
化学反应的自由能随环境温度和物质浓度(活度)而改变,在比较自由能变化时,必 须在标准状况下进行测定,即 25℃,溶液中溶质的标准状态为单位摩尔浓度,若为气体, 则为 101.325kPa,所测得的值称为标准自由能变化,用△G 表示。
本章首先介绍生物氧化的基本概念、特点及方式,然后侧重讨论各类有机物(糖、蛋 白质、脂肪等)在细胞内进行生物氧化所经历的一段共同的终端氧化过程中,代谢中间物 脱氢生成的还原型辅酶(NADH 和 FADH2)如何经电子传递链(呼吸链)的电子传递被分 子氧氧化,电子传递过程如何与 ADP 磷酸化生成 ATP 的过程相偶联。
在化学反应中,自由能和化学反应平衡常数 Keq 之间有如下的关系△G =-RTlnKeq。 在生物体内参与反应的物质浓度都很低,往往不是在标准状况,所测得的自由能变化并不 是标准自由能变化,用△G 表示。△G 与标准自由能变化△G 之间有一定的关系,可用公 式表示△G=△G +RTlnKeq。
生物化学 第六章 生物氧化

电子传递链(呼吸链)
琥珀酸 复 合 体 Ⅰ
2H
复合体Ⅱ FAD.H2 (Fe-S)
2H 2H 2e
2H NAD+
复 合 体 琥珀酸氧化呼吸链 Ⅳ
2e
FMN (Fe-S)
Q10
2H+
Cytb Cytc1 2e (Fe-S) 复合体Ⅲ H2O
Cytc
2e
aa3
2e
NADH氧化呼吸链
O2-
1 2 O2
第三节 ATP的生成
(二)呼吸链成分的排列
由以下实验确定 ① 标准氧化还原电位 ② 拆开和重组 ③ 特异抑制剂阻断 ④ 还原状态呼吸链缓慢给氧
呼吸链中各种氧化还原对的标准氧化还原电位 氧化还原对 NAD+/NADH+H+ FMN/ FMNH2 FAD/ FADH2 Cyt b Fe3+/Fe2+ Q10/Q10H2 Cyt c1 Fe3+/ Fe2+ Cyt c Fe3+/Fe2+ Cyt a Fe3+ / Fe2+ Cyt a3 Fe3+ / Fe2+ 1/2 O2/ H2O Eº (V) ' -0.32 -0.30 -0.06 0.04(或0.10) 0.07 0.22 0.25 0.29 0.55 0.82
故又称混合功能氧化酶(mixed-function oxidase) 或羟化酶(hydroxylase)。 上述反应需要细胞色素P450 (Cyt P450)参与。
微粒体氧代谢的意义
参与体内正常物质代谢,如羟化、合成等
参与体内生理活性物质的灭活及药物、毒
物解毒转化和代谢清除反应、保护机体
生物化学第六章ATP

• 细胞色素可存在于线粒体内膜,也可存在于
微粒体。
• 存在于线粒体内膜的细胞色素有Cyt aa3, Cyt b(b560,b562,b566),Cyt c,Cyt c1;
Cyt c是呼吸链唯一水溶性球状蛋白, 不包含在复合体中,可将获得的电子 传递到复合体Ⅳ。
磷酸化,生成ATP,又称为偶联磷酸化。
呼 吸 链
AH2
2H(2H++2e)
1 2 O2
H2O
氧化 偶 联
A ADP+Pi
能量 ATP 磷酸化
(一)氧化磷酸化的偶联部位
根据P/O比值
自由能变化: ⊿Gº '=-nF⊿Eº '
1. P/O 比值 指氧化磷酸化过程中,每消耗1摩尔O 的同时所消耗磷原子的摩尔数。 ADP+H3PO4 ATP
NADH+H+
NAD+
FMN
FMNH2
还原型Fe-S 氧化型Fe-S
Q
QH2
复合体Ⅰ的功能
3. 复合体Ⅱ(琥珀酸-泛醌还原酶)
复合体Ⅱ含三羧酸循环中的琥珀酸脱氢酶, 功能是将电子从琥珀酸传递到泛醌。
电子传递:
琥珀酸→ FAD;Fe-S1; Fe-S2 ; Fe-S3
→CoQ
3.复合体Ⅲ(泛醌-细胞色素C还原酶)
组成
递氢体和电子传递体(2H 2H+ + 2e)
递氢体或递电子体通常以复合体的形式存在
于线粒体内膜上。
线粒体
基质侧
胞液侧 膜间隙
(一)呼吸链的组成
第六章 生物氧化

E0‘(V)
-0.32 -0.219 -0.219
氧化还原对
Cyt c1 Fe3+ /Fe2+ Cyt c Fe3+ /Fe2+ Cyt a Fe3+ /Fe2+
E0‘(V)
0.22 0.254 0.29
Cyt bL(bH) Fe3+/Fe2+
Q10 /Q10H2
0.05(0.10)
0.06
Cyt a3 Fe3+ /Fe2+
FMN(FAD)的结构:
CH2OPO32H H H C C C CH2 H3C N N O OH OH OH
N H3C N
异 咯 嗪
O
异咯嗪环的作用:
FMN/FAD
FMNH /FADH
FMNH 2/FADH
2
(氧化型)
(还原型)
铁硫蛋白 铁硫蛋白(Fe-S)共有
9种同工蛋白;分子中
含有由半胱氨酸残基硫
目录
泛醌从复合体Ⅰ、Ⅱ募集还原当量和电子并穿 梭传递到复合体Ⅲ。
电子传递过程:CoQH2→(Cyt bL→Cyt bH)
→Fe-S →Cytc1→Cytc
目录
细胞色素类:
这是一类以铁卟啉为辅基的酶。在生物氧 化反应中,其铁离子可为+2价亚铁离子,也可 为+3价高铁离子,通过这种转变而传递电子。
R=H: NAD+;
R=H2PO3: NADP+
目录
NAD+(NADP+)和NADH(NADPH)相互转变
氧化还原反应时变化发生在五价氮和三价氮之间。
目录
FMN结构中含核黄素,发挥功能的部位是异 咯嗪环,氧化还原反应时不稳定中间产物是 FMNH· 。在可逆的氧化还原反应中显示3种分子状 态,属于单、双电子传递体。
第6章--生物氧化习题

第六章生物氧化习题一、名词解释1.生物氧化:有机物质在生物体活细胞内氧化分解,同时释放能量的过程。
2 氧化磷酸化:指底物脱下的2H经过电子传递链传递到分子氧形成水的过程中释放出能量与ADP磷酸化生成 ATP的过程相偶联生成ATP的方式。
3 底物水平磷酸化:某些底物分子中含有高能磷酸键,可转移至ADP生成ATP的过程。
4呼吸链:代谢物上的氢原子被脱氢酶激活脱落后,经过一系列的传递体,最后传递给被激活的氧分子而生成水的全部体系称呼吸链。
5 高能化合物:在生物体内随水解反应或基团转移反应可放出大量自由能的化合物成为高能化合物。
6 磷氧比:指每消耗1mol氧原子所产生的ATP的物质的量。
7 电子传递抑制剂:能够阻断电子传递链中某一部位电子传递的物质称为电子传递抑制剂。
8 解偶联剂:具有解偶联作用的化合物称为解偶联剂。
9 氧化磷酸化抑制剂:是指直接作用于线粒体F0F1-ATP酶复合体中的F1组分而抑制ATP合成的一类化合物。
10 F0F1-ATP合酶:位于线粒体内膜基质一边,由F0和F1构成的复合体。
是一种ATP驱动的质子运输体,当质子顺电化学梯度流动时催化ATP的合成;当没有氢离子梯度通过质子通道F0时,F1的作用是催化ATP的水解。
二、选择题1.生物氧化的底物是:〔 D 〕A、无机离子B、蛋白质C、核酸D、小分子有机物2.除了哪一种化合物外,以下化合物都含有高能键?〔 D 〕A、磷酸烯醇式丙酮酸B、磷酸肌酸C、ADPD、G-6-PE、1,3-二磷酸甘油酸3.以下哪一种氧化复原体系的氧化复原电位最大?〔 C 〕A、延胡羧酸→丙酮酸B、CoQ(氧化型) →CoQ(复原型)C、Cyta Fe2+→Cyta Fe3+D、Cytb Fe3+→Cytb Fe2+E、NAD+→NADH4.呼吸链的电子传递体中,不是蛋白质而是脂质的组分是:〔 D 〕A、NAD+B、FMNC、FE、SD、CoQE、Cyt5.2,4-二硝基苯酚抑制细胞的功能,可能是由于阻断以下哪一种生化作用而引起? 〔 E 〕A、NADH脱氢酶的作用B、电子传递过程C、氧化磷酸化D、三羧酸循环E、电子传递与氧化磷酸化的偶联过程6.能使线粒体电了传递与氧化磷酸化解偶联的试剂是:〔 A 〕A、2,4-二硝基苯酚B、寡霉素C、一氧化碳D、氰化物7.呼吸链的各细胞色素在电子传递中的排列顺序是:〔 D 〕A、c1→b→c→aa3→O2B、c→c1→b→aa3→O2C、c1→c→b→aa3→O2D、b→c1→c→aa3→O28.在呼吸链中,将复合物I、复合物II与细胞色素系统连接起来的物质是什么?〔 C 〕A、FMNB、Fe·S蛋白C、CoQD、Cytb9.下述那种物质专一的抑制F0因子?〔 C 〕A、鱼藤酮B、抗霉素AC、寡霉素D、氰化物10.下述分子哪种不属于高能磷酸化合物:〔 C 〕A、ADPB、磷酸烯醇式丙酮酸C、乙酰COAD、磷酸肌酸11.细胞色素c是——:〔 C 〕A、一种小分子的有机色素分子B、是一种无机色素分子C、是一种结合蛋白质D、是一种多肽链12.以下哪种物质抑制呼吸链的电子由NADH向辅酶Q的传递:〔 B 〕A、抗霉素AB、鱼藤酮C、一氧化碳D、硫化氢13.以下哪个部位不是偶联部位:〔 B 〕A、FMN→CoQB、NADH→FMNC、b→cD、a1a3→O214.ATP的合成部位是:〔 B 〕A、OSCPB、F1因子C、F0因子D、任意部位15.目前公认的氧化磷酸化理论是:〔 C 〕A、化学偶联假说B、构象偶联假说C、化学渗透假说D、中间产物学说16.以下代谢物中氧化时脱下的电子进入FADH2电子传递链的是:〔 D 〕A、丙酮酸B、苹果酸C、异柠檬酸D、琥珀酸17.以下呼吸链组分中氧化复原电位最高的是:〔 C 〕A、FMNB、CytbC、CytcD、Cytc118.ATP含有几个高能键:〔 B 〕A、1个B、2个C、3个D、4个19.在使用解偶联剂时,线粒体内膜:〔 B 〕A、膜电势升高B、膜电势降低C、膜电势不变D、两侧pH升高20.线粒体电子传递链各组分:〔 C 〕A、均存在于酶复合体中B、只能进行电子传递C、氧化复原电势一定存在差异D、即能进行电子传递,也能进行氢的传递二、填空题1.生物氧化是有机分子在细胞中氧化分解,同时产生可利用的能量的过程。
第六章生物氧化(本科第七版)

FADH2或FMNH2
2)铁硫蛋白(铁硫中心)及其辅基铁硫簇(Fe-S):在 生物氧化中起递电子的作用。
5、复合体Ⅲ
复合体Ⅲ又叫泛醌-细胞色素C还原酶,细胞色素 b-c1复合体,含有细胞色素b(b562, b566)、细胞 色素c1和一种可移动的铁硫蛋白(Rieske protein)。
电子传递过程:CoQH2→(Cyt b566(L)→Cyt
NADH氧化呼吸链
FADH2氧化呼吸链
琥珀酸
FAD (Fe-S) NADH FMN (Fe-S) CoQ Cyt b→Cyt c1→Cyt c Cyt aa3 O2
二、 ATP的生成
底物水平磷酸化
ATP生成方式 氧化磷酸化
底物水平磷酸化(substrate level phosphorylation ): 底 物脱氢使分子内部能量重新分布,生成的高能键, 直接转给ADP磷酸化生成ATP的过程。
F
+
-
Ⅰ
NAD+
Ⅱ
NADH+H+
延胡索酸 琥珀酸
-
Ⅲ -
- 1/2O2+2H+
- H2 O
Ⅳ
0
F1
基质侧
ADP+Pi ATP H+
2、ATP合酶(ATP synthase):复合体Ⅴ
F0(疏水部分):由a1b2c9~12亚基组成,是镶嵌
在线粒体内膜中的质子通道
ATP合酶
F1(亲水部分):由33亚基组成,其功能是
能量,当质子顺浓度梯度回流时驱动ADP与Pi生成ATP。
简 单 示 意 图
线 粒 体 内 外 膜 间 隙
线粒体内膜 线粒体基质
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
氧化途 主要存 主要承 径 在的组 担酶 织
-磷酸 骨骼肌、 α-磷酸
甘油穿 神经细 甘油脱
梭
胞
氢酶
苹果酸 肝、心 苹果酸 穿梭 肌组织 脱氢酶
胞液中 主要承 担酶的
辅基 NAD+
NAD+
线粒体 内主要 承担酶 的辅基
FAD
被完全 氧化时 经过的 呼吸链
琥珀酸氧 化呼吸链
完全氧 化时产 生的 ATP量
下的氢和电子沿呼吸链传递过程
中,逐步释放能量使ADP氧化生
成ATP。这种氧化与磷酸化紧密
偶联的过程称氧化磷酸化作用。
17.11.2020
h
5
AH2 2H
A
NADH++H (或 FADH2)
H2O
NAD+ (或 FAD)
电子传递链
氧化过程
氧
1/2 O2
化
磷
释放能量
酸 化
ADP + Pi ATP合成酶 ATP 磷酸化过程
第三节 底物水平磷酸化
一ATP的生成方式
底物水平磷酸化 氧化磷酸化
17.11.2020
h
1
(一)底物水平磷酸化
1.定义:由底物分子因脱氢或脱
水而使分子内部能量重新分配产生
的高能磷酸键(或高能硫酯键),
在激酶作用下将高能键上的键能直
接转移给ADP(或 GDP)而生成
ATP(或 GTP)的反应。
17.11.2020
Fe-S Cyt c1
Cyt c
Cyt aa3 复合物 IV
O2
呼吸链抑制剂的阻断位点
抗霉素A 二巯基丙醇
17.11.2020
鱼藤酮 粉蝶霉素A 异戊巴比妥 h
CO、CN-、 N3-及H2 S
38
17.11.2020
h
39
(四) 解偶联剂
不阻断电子传递,但拆散氧化和磷酸化的 偶联作用。
P/O很低,甚至为零
17.11.2020
h
19
化学渗透假说的支持证据
①电子传递能形成H+浓度梯度 ②线粒体内膜对H+、OH-、Cl-、K+等不能透过 ③ ATP合成需要有完整的内膜 ④破坏H+浓度梯度使得磷酸化不能进行 ⑤膜表面能储存大量质子,也能迅速转移质子
17.11.2020
h
20
化学渗透假说示意图
H
+
H
+
H
H
17.11.2020
h
36
(三) 呼吸链抑制剂
粉蝶霉素A
鱼藤酮 安密妥
琥珀酸
作用:阻断电子传递
FMN
Fe-S
复合物 II
抗霉素A
NAD FP Q b c aa3
NAD FP Q b c aa3
抗霉素 A的 抑制部位
呼吸链的比拟图解
氰化物 CO
NADH
FMN 复合物 物 III
17.11.2020
h
40
2,4一二硝基酚(DNP)
使电子传递和ATP两个过程分离,失掉它们的 紧密联系,只抑制ATP的形成过程,但不抑制 电子传递过程
17.11.2020
h
41
解偶联作用机制
H+
H+
H+
解偶联蛋白
H+
17.11.2020
热
H+
ADP+ ATP+H
P
2
h
i
42O
氧化磷酸化抑制剂
17.11.2020
h
6
2、呼吸链成分的排列次序
标准氧化还原电位及自由能变化 复合体体外拆开与重组 特异抑制剂阻断 还原状态呼吸链缓慢给氧
17.11.2020
h
8
17.11.2020
h
9
呼吸链电子传递过程中,哪些区段 放出的能量能实现ADP的磷酸化?
17.11.2020
h
10
二、氧化磷酸化偶联部位 确定
(二)质子梯度的形成 (三)线粒体 ATP合酶(mitochondrial ATPase) (四)ATP合成的机制
17.11.2020
h
16
(一)能量偶联假说 1953年 Edward Slater 化学偶联假说 1964年 Paul Boyer 构象偶联假说 1961年 Peter Mitchell 化学渗透假说
直接干扰 ATP的生成 过程
阻止质子从 F0质子通道 回流。
寡霉素
17.11.2020
h
43
(五) 离子载体抑制剂 能与某些离子结合并作为载体使之穿越膜 它们能与除H+以外的所有一价阳离子结合
17.11.2020
h
44
17.11.2020
h
45
七、有关氧化磷酸化物质的运输
• 胞液中的3-磷酸甘油醛,3-磷酸甘油或乳 酸脱氢,均可产生NADH。
h
33
六、氧化磷酸化的影响因素 (一) 激素 甲状腺素 (二) ADP/ATP (三)抑制剂的作用 (四) 线粒体DNA突变
17.11.2020
h
34
(一) 激素 甲状腺素 促进细胞膜上的Na+-K + -ATPase 的生成,
促进ATP的分解
Na+-K+ ATP酶活性 ATP分解 ADP/ATP 氧化磷酸化
质子穿过a时,
推动c环象水 车一样转动,连 带F1转动.
17.11.2020
h
27
17.11.2020
h
28
五 ATP合成的机制
旋转催化机制
定子
转子
ba
17.11.2020
c
h
29
17.11.2020
h
30
17.11.2020
h
31
17.11.2020
h
32
旋转催化具体过程
17.11.2020
2ATP
NAD+ NADH氧 3ATP
化呼吸链
17.11.2020
h
52
17.11.2020
h
17
17.11.2020
h
18
化学渗透假说的内容★★
1.呼吸链中传氢体和电子传递体是间隔交替排列的,且 在线粒体内膜都有特定的位置,催化反应是定向的。
2.内膜对H+不能自由通过,泵出膜外侧的H+不能自由 返回膜内侧,造成电化学梯度
3.复合体Ⅰ、Ⅲ、Ⅳ都有质子泵的作用
4.ATP合酶存在于线粒体内膜上,H+梯度是ATP合成的 驱动力
• 这些NADH可经穿梭系统而进入线粒体氧
化磷酸化,产生H2O和ATP。
17.11.2020
h
46
-磷酸甘油穿梭示意图
NADH 油-脱磷氢酸酶甘+H+
NA D
+
呼吸链
FAD H
-磷酸甘
2
油脱氢酶
FA D
17.11.2020
h
47
α-磷酸甘油穿梭
NADH
(细胞液)
磷酸二羟丙酮
NAD+
3-磷酸甘油
磷酸二羟丙酮
17.11.2020
h
35
(二) ADP/ATP
定义式:能荷=
[ATP]+0.5[ADP] —————————
[ATP]+[ADP]+[AMP]
意义: 能荷由ATP 、 ADP和AMP的相对数量决定, 数值在0~1之间,反映细胞能量水平。
能荷对代谢的调节可通过ATP 、 ADP和AMP作为代 谢中某些酶分子的别构效应物进行变构调节来实现。
Ⅱ
谷氨酸 草酰乙酸 NADH+H+
谷草转氨酶
谷草转氨酶
天冬氨酸 -酮戊二酸 Ⅲ -酮戊二酸 天冬氨酸
呼吸链
17.11.2020
Ⅳ
h
50
(Ⅰ、 Ⅱ、 Ⅲ、 Ⅳ为膜上的转运载体)
线粒体外NADH两种氧化途径的比 较
穿梭物质 进入线粒 体前后转变 成的物质 进入呼吸链 生成ATP数 存在组织
α-磷酸甘油穿梭 α-磷酸甘油 磷酸二羟丙酮
NAD+/FAD H2
琥珀酸氧化呼吸链 1.5/2
某些肌肉、神经组织
苹果酸-天冬氨酸穿梭 苹果酸、 谷氨酸 天冬氨酸、α-酮戊二酸
NAD+/NADH+ H+ NADH 氧化呼吸链
2.5/3 肝脏和心肌组织
相同点
17.11.2020
将胞浆中NADH的还原当量转送到线粒体
内
h
51
NADH两种氧化途径的比较
氧化过程
物质氧化
高能电子 氧
energy
质子动力势 energy
ADP+Pi
ATP
17.11.2020
磷酸化过程
h
24
四 线粒体 ATP合酶(mitochondrial ATPase)
形成ATP的机理
F1
17.11.2020
h
柄
F0
25
17.11.2020
h
26
ATP合酶
ATP合成酶
由疏水的 F0(a1b2c1012) 和亲水的 F1(33)组 成.
h
2
2.举例:糖酵解过程的底物磷酸化:
CHO
dehydrogenase
O
CO~ P
CHOH +NAD+ + Pi
CHOH
CH2O P
CH2O P
+NADH +H+
O
CO~ P
CHOH