关于特异性免疫应答的特点及其机制课件
合集下载
特异性免疫ppt课件模板

γδT细胞:识别抗原,参 与免疫应答和炎症反应
抗原分子表面特定的三维结构:通过抗原抗体结合特异性的机制进行识别
结合亲和力:结合时抗体的立体构象要能精确识别抗原表面的结构,才具备足够的亲和力进行结合。亲和力过高过低的抗原 抗体结合,亲和力过高会导致血液凝集反应,亲和力过低则无法形成肉眼可见的凝集反应。
定义:疫苗接种是特异性免疫诱导的一种主要方式,通过向人体注射含有抗原成分的疫苗,使人体免疫系统产 生针对该抗原的特异性免疫反应。:目的:疫苗接种的目的是使人体在免疫系统未被病原感染的情况下,提前 产生针对特定病原的免疫力,从而在病原感染时迅速并有效地抵抗病原,保护机体健康。
疫苗种类:疫苗包括灭活疫苗、减毒活疫苗、基因工程疫苗等,其中灭活疫苗是最常用的疫苗类型。:接种方 式:疫苗接种通常采用注射的方式进行,部分疫苗也可以通过口服或鼻吸的方式进行接种。
汇报人:XXX
01 02 03 04 05 06
定义:特异性免疫是指免疫系统针对特定病原的免疫反应,也称为适应性免疫。
特点:特异性免疫具有高度的特异性,只对特定的病原体或抗原产生免疫反应,而对其他病原体无效。
特异性免疫的记忆功能:当身体与特定病原体再次接触时,能够更快、更有效地产生免疫反应。:特异性免疫由T淋 巴细胞和B淋巴细胞等参与,通过释放特异的细胞因子或抗体来达到消除病原体的目的。
单击添加标题
发现新的免疫细胞类型:研究已经发现了一些新的免疫细胞类型,这些细胞具 有独特的生物学特性和功能,可以用来治疗某些疾病。
单击添加标题
揭示免疫反应的机制:科学家们已经更深入地了解了免疫反应的机制,这有助 于更好地理解免疫系统如何应对感染和疾病。
单击添加标题
改进疫苗设计:基于对免疫反应机制的理解,科学家们正在开发更加高效和安 全的疫苗,以预防和治疗各种疾病。
特异性免疫幻灯片课件

特异性免疫幻灯片课件
汇报人: 日期:
• 引言 • 特异性免疫基础知识 • 特异性免疫的细胞组成及其作用 • 特异性免疫的过程及其调控 • 特异性免疫在抗感染中的作用
• 特异性免疫在自身免疫性疾病和过 敏反应中的作用
• 特异性免疫在肿瘤免疫中的作用 • 总结与展望
01
引言
什么是特异性免疫
定义
特异性免疫是人体在接触外来物质或 抗原后,产生的一种针对该物质的特 殊防御机制。
化、增殖和分化。
效应T、B细胞的产生与功能
T细胞效应
通过释放穿孔素和颗粒酶等效应分子,诱导靶细胞凋亡。
B细胞效应
通过产生抗体发挥体液免疫功能,同时也可通过吞噬病原体 等发挥细胞免疫功能。
免疫记忆与再次免疫应答
免疫记忆
初次免疫应答产生的记忆细胞可在再次接触相同抗原时迅速、高效 地产生免疫应答。
再次免疫应答
02
特异性免疫具有高度特异性和记 忆性,能够快速并有效地对抗再 次入侵的病原微生物。
特异性免疫研究的前沿与挑战
深入探究特异性免疫的分子机制和细胞相互作用,揭示其在免疫应答和免疫耐受中 的作用。
研究特异性免疫在免疫相关疾病发生发展中的作用,为免疫治疗提供新的思路和方 法。
针对特异性免疫细胞的靶向治疗和疫苗设计是当前研究的热点,已有许多研究成果 应用于临床。
基于特异性免疫的免疫治疗前景
基于特异性免疫的免疫治疗已成 为一种重要的治疗策略,在肿瘤 、感染性疾病、自身免疫性疾病
等治疗中取得显著进展。
进一步挖掘特异性免疫细胞的调 控机制,提高其活化和扩增能力 ,对于提高治疗效果具有重要意
义。
针对特异性免疫细胞的靶向药物 和疫苗设计仍需不断改进和完善 ,以满足不同疾病的治疗需求。
汇报人: 日期:
• 引言 • 特异性免疫基础知识 • 特异性免疫的细胞组成及其作用 • 特异性免疫的过程及其调控 • 特异性免疫在抗感染中的作用
• 特异性免疫在自身免疫性疾病和过 敏反应中的作用
• 特异性免疫在肿瘤免疫中的作用 • 总结与展望
01
引言
什么是特异性免疫
定义
特异性免疫是人体在接触外来物质或 抗原后,产生的一种针对该物质的特 殊防御机制。
化、增殖和分化。
效应T、B细胞的产生与功能
T细胞效应
通过释放穿孔素和颗粒酶等效应分子,诱导靶细胞凋亡。
B细胞效应
通过产生抗体发挥体液免疫功能,同时也可通过吞噬病原体 等发挥细胞免疫功能。
免疫记忆与再次免疫应答
免疫记忆
初次免疫应答产生的记忆细胞可在再次接触相同抗原时迅速、高效 地产生免疫应答。
再次免疫应答
02
特异性免疫具有高度特异性和记 忆性,能够快速并有效地对抗再 次入侵的病原微生物。
特异性免疫研究的前沿与挑战
深入探究特异性免疫的分子机制和细胞相互作用,揭示其在免疫应答和免疫耐受中 的作用。
研究特异性免疫在免疫相关疾病发生发展中的作用,为免疫治疗提供新的思路和方 法。
针对特异性免疫细胞的靶向治疗和疫苗设计是当前研究的热点,已有许多研究成果 应用于临床。
基于特异性免疫的免疫治疗前景
基于特异性免疫的免疫治疗已成 为一种重要的治疗策略,在肿瘤 、感染性疾病、自身免疫性疾病
等治疗中取得显著进展。
进一步挖掘特异性免疫细胞的调 控机制,提高其活化和扩增能力 ,对于提高治疗效果具有重要意
义。
针对特异性免疫细胞的靶向药物 和疫苗设计仍需不断改进和完善 ,以满足不同疾病的治疗需求。
高中生物特异性免疫精品课件

。其他组进行完善并可提出疑问。
请结合模型图用思维导图写出体液免疫的过程
抗原
特
异
巨噬细胞(处理)
性 结
抗原-MHC复合体
体
合
液
辅助性T细胞(特异性识别)
致敏
白细胞介素-2
B淋巴细胞
免 疫
克隆、分化
记忆B细胞
效应B细胞 (浆细胞)
分泌 抗体:与抗原特异性结合
思考: 若从欧洲人体内检测到天花抗体,说明什么? 感染过天花病毒的人为什么会终身免疫?
请用今天所学知识提出适当的措施来降低埃及法 老陵墓考古学家的死亡率
谢谢
一、抗体和抗原
1、抗 原 可使机体产生特异性免疫反应的物质。
2、抗 体 概念: 能与抗原发生特异性结合的具
有免疫功能的球蛋白。
抗体立体模式图
结构: Y形的,两臂上各有一个相同 的抗原结合位点
抗体 效应B淋巴细胞
抗体的产生需要哪些细胞参与呢?
资料一:给失去免疫功能的小鼠输入来自胸腺的淋巴 细胞(T细胞),小鼠不能产生体液中的抗体;如果 输入来自骨髓的淋巴细胞(B细胞),小鼠产生抗体 的功能只能部分恢复;如果同时输给小鼠T细胞产生 的淋巴因子,小鼠产生抗体的功能全部恢复。
(1)从中看出免疫反应中最早发现抗原的是哪一种 细胞?
(2)抗原、巨噬细胞、T细胞之间的顺序关系?
巨噬细胞处理、呈递抗 原
小组活动:体液免疫的过程
活动方案:前后四人一个小组。每组将得到一个信封
(每个信封内有标记了抗原,抗体,巨噬细胞,抗原-MHC复合体, B淋巴细胞,辅助性T细胞,效应B细胞,记忆B细胞的卡片)
记忆B细胞
效应B细胞
克隆、分化 (浆细胞)
抗体:与抗原特异性结合
请结合模型图用思维导图写出体液免疫的过程
抗原
特
异
巨噬细胞(处理)
性 结
抗原-MHC复合体
体
合
液
辅助性T细胞(特异性识别)
致敏
白细胞介素-2
B淋巴细胞
免 疫
克隆、分化
记忆B细胞
效应B细胞 (浆细胞)
分泌 抗体:与抗原特异性结合
思考: 若从欧洲人体内检测到天花抗体,说明什么? 感染过天花病毒的人为什么会终身免疫?
请用今天所学知识提出适当的措施来降低埃及法 老陵墓考古学家的死亡率
谢谢
一、抗体和抗原
1、抗 原 可使机体产生特异性免疫反应的物质。
2、抗 体 概念: 能与抗原发生特异性结合的具
有免疫功能的球蛋白。
抗体立体模式图
结构: Y形的,两臂上各有一个相同 的抗原结合位点
抗体 效应B淋巴细胞
抗体的产生需要哪些细胞参与呢?
资料一:给失去免疫功能的小鼠输入来自胸腺的淋巴 细胞(T细胞),小鼠不能产生体液中的抗体;如果 输入来自骨髓的淋巴细胞(B细胞),小鼠产生抗体 的功能只能部分恢复;如果同时输给小鼠T细胞产生 的淋巴因子,小鼠产生抗体的功能全部恢复。
(1)从中看出免疫反应中最早发现抗原的是哪一种 细胞?
(2)抗原、巨噬细胞、T细胞之间的顺序关系?
巨噬细胞处理、呈递抗 原
小组活动:体液免疫的过程
活动方案:前后四人一个小组。每组将得到一个信封
(每个信封内有标记了抗原,抗体,巨噬细胞,抗原-MHC复合体, B淋巴细胞,辅助性T细胞,效应B细胞,记忆B细胞的卡片)
记忆B细胞
效应B细胞
克隆、分化 (浆细胞)
抗体:与抗原特异性结合
医学免疫学特异性免疫应答的特点文稿演示

意义,是预防接种的免疫学基础。
T记忆细胞特征
Tm比Tn更易激活,所需抗原浓度较低; Tm对协同刺激信号的依赖性较Tn低; Tm分泌的CK更多,且对CK更敏感; Tn寿命短,Tm寿命长,表达更多的bcl-2,
CK(IL-15)维持其生存; 记忆CD8+T细胞的维持无需CD4+T细胞辅
助。
B记忆细胞特征
疫不成熟状态,有利于诱导免疫耐受。
免疫应答的耐受性
* 免疫耐受的概念及特性 * 免疫耐受的诱导条件 * 免疫耐受形成机制 * 研究免疫耐受的意义
自身免疫耐受的机制
* 中枢性免疫耐受 胚胎期免疫系统未成熟的T、B细
胞在中枢性免疫器官接触抗原所形 成的免疫耐受。
* 外周性免疫耐受 成熟的T、B细胞在外周淋巴器官
(一)抗原方面
3. 抗原免疫途径 静脉注射/口服>腹腔注射>皮下/肌肉注射
4. 其他因素 是否用佐剂等
(二)机体方面
* 免疫系统的成熟程度种属和品系(遗传)
大鼠、小鼠 > 兔、有蹄类、灵长类
* 机体生理状态 用免疫抑制剂破坏成熟淋巴细胞,造成类似新生期免
医学免疫学特异性免疫应答的特点文稿演示
主要内容
* 免疫应答的特异性 * 免疫应答的记忆性 * 免疫耐受性
特异性表现: 特异性本质:
BCR多样性的机制
* 组合多样性 * 连接多样性 * 体细胞高频突变(B细胞)
主要内容
* 免疫应答的特异性 * 免疫应答的记忆性 * 免疫耐受性
免疫应答的记忆性
免疫应答的耐受性
* 免疫耐受的概念及特性 * 免疫耐受的诱导条件 * 免疫耐受形成机制 * 研究免疫耐受的意义
胚胎期及新生期免疫耐受现象
T记忆细胞特征
Tm比Tn更易激活,所需抗原浓度较低; Tm对协同刺激信号的依赖性较Tn低; Tm分泌的CK更多,且对CK更敏感; Tn寿命短,Tm寿命长,表达更多的bcl-2,
CK(IL-15)维持其生存; 记忆CD8+T细胞的维持无需CD4+T细胞辅
助。
B记忆细胞特征
疫不成熟状态,有利于诱导免疫耐受。
免疫应答的耐受性
* 免疫耐受的概念及特性 * 免疫耐受的诱导条件 * 免疫耐受形成机制 * 研究免疫耐受的意义
自身免疫耐受的机制
* 中枢性免疫耐受 胚胎期免疫系统未成熟的T、B细
胞在中枢性免疫器官接触抗原所形 成的免疫耐受。
* 外周性免疫耐受 成熟的T、B细胞在外周淋巴器官
(一)抗原方面
3. 抗原免疫途径 静脉注射/口服>腹腔注射>皮下/肌肉注射
4. 其他因素 是否用佐剂等
(二)机体方面
* 免疫系统的成熟程度种属和品系(遗传)
大鼠、小鼠 > 兔、有蹄类、灵长类
* 机体生理状态 用免疫抑制剂破坏成熟淋巴细胞,造成类似新生期免
医学免疫学特异性免疫应答的特点文稿演示
主要内容
* 免疫应答的特异性 * 免疫应答的记忆性 * 免疫耐受性
特异性表现: 特异性本质:
BCR多样性的机制
* 组合多样性 * 连接多样性 * 体细胞高频突变(B细胞)
主要内容
* 免疫应答的特异性 * 免疫应答的记忆性 * 免疫耐受性
免疫应答的记忆性
免疫应答的耐受性
* 免疫耐受的概念及特性 * 免疫耐受的诱导条件 * 免疫耐受形成机制 * 研究免疫耐受的意义
胚胎期及新生期免疫耐受现象
特异性免疫应答的特点及其机制

②重组酶—特异性识别并切除RSS
等位排斥和同种型排斥 等位排斥
指B细胞中位于一对染色体上的轻链
或重链基因,其中只有一条染色体上的基因得到表 达。保证一个B细胞只表达一种轻链和一种重链。
同种型排斥 指κ轻链和λ轻链之间的排斥,
一个B细胞只能表达其中一种κ链或链。
免疫球蛋白类别转换
(class switching,isotype switching)
Vk
Germline
Jk
Ck
Rearranged 1° transcript
Spliced mRNA
Ig重链V区基因重排
Cm
VH 1-123
DH1-27
JH 1-9
Ig重链C区基因重排
Cm Primary transcript RNA AAAAA
Each domain of the H chain is encoded by a separate exon
类别转换
抗原激活B细胞后,膜上表达
和分泌的Ig类别会从IgM转换成IgG、IgA、
IgE等其它类别或亚类的Ig,即Ig的V区不 变C区发生转换,这种现象称类别转换。
膜型Ig和分泌型Ig重链基因重排
发生时间:在转录加工中造成 由两个外显子决定(SC分泌型、MC膜型)
在转录时,若是转录到SC,则为分泌型Ig。
C基因组成
BCR、TCR基因的重排
重排发生在TB细胞在中枢免疫器官发育过程中 BCR和TCR胚系基因中隔开的V、(D)、J基因片 段通过基因片段的重排,形成V(D)J连接重排后 的DNA。
转录初级RNA,剪接形成mRNA(含C基因)
翻译为重链( β、δ链)或轻链( α、γ链)
特异性免疫ppt课件

系统性红斑狼疮
总结词
系统性红斑狼疮是一种典型的自身免疫性疾 病,特异性免疫在疾病中发挥关键作用。
详细描述
系统性红斑狼疮的发病机制复杂,可能与遗 传、环境、感染等因素有关。特异性免疫细 胞如T淋巴细胞和B淋巴细胞在疾病中发挥 重要作用。这些细胞可针对自身抗原产生抗 体,导致全身性炎症和组织损伤。
自身免疫性甲状腺炎
白细胞介素-2
由T细胞产生,具有刺激T细胞 的生长和分化作用。
白细胞介素-3
由造血细胞产生,具有刺激造 血细胞的作用。
白细胞介素-4
由Th2细胞产生,具有刺激B细 胞的作用。
MHC分子与T细胞受体
MHC分子
主要组织相容性复合体,在抗原 提呈中发挥重要作用。
T细胞受体
T细胞表面的一种糖蛋白,可以识 别抗原-MHC分子复合物并激活T 细胞。
特异性免疫系统在抵抗疟疾感染 中具有关键作用,识别疟原虫抗 原并产生相应的抗体和细胞因子
,抑制寄生虫的生长和繁殖。
阿米巴病
阿米巴病感染时,特异性免疫系 统通过产生相应的抗体和细胞因 子,加速寄生虫的清除和减轻病
情。
弓形虫感染
特异性免疫系统能够识别弓形虫 抗原,触发有效的免疫应答,控
制感染的扩散。
05
感染性疾病的免疫治疗定义
感染性疾病的免疫治疗是指通过激活 患者自身的免疫系统来抵抗病原微生 物感染的治疗方法。
感染性疾病的免疫治疗分类
根据作用机制不同,感染性疾病的免 疫治疗可分为抗病毒免疫治疗、抗细 菌免疫治疗等。
抗病毒免疫治疗
抗病毒免疫治疗可增强机体免疫力, 促进机体产生病毒抗体,从而抵抗病 毒感染。
特异性免疫的历史与发展
发现与发展
免疫学第十三章-特异性免疫应答的特点及其机制
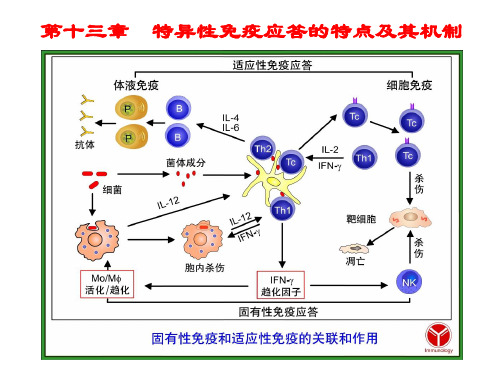
特点:免疫耐受具有免疫特异性,即只对特定的抗原 不应答,对不引起耐受的抗原,仍能进行良好的免疫应 答;一般情况下。不影响适应性免疫应答的整体功能。
二、 免疫耐受的诱导条件和形成机制 胚胎发育期形成的免疫耐受原则上可长期持续,不会轻
易打破。而后天形成的耐受,可持续一段时间,可随诱导因 素的消失而解除。
1. 记忆性T细胞的特征 ① Tm(memory T cell)比Tn( naïve T cell)更易激活,所需
抗原浓度较低; ② Tm对协同刺激信号的依赖性较Tn低; ③ Tm分泌的CK更多,且对CK更敏感; ④ Tn寿命短,Tm寿命长, CK(IL-15)维持其生存; ⑤ 记忆CD8+T细胞的维持无需CD4+T细胞辅助。
中枢耐受
(2)外周耐受
① 克隆清除和免疫忽视
• 克隆清除是指T细胞克隆的TCR对组织特异自身抗原有 高亲和力,且浓度高时,经APC提呈,可致此类T细胞 克隆清除(delection)。
• 若T细胞克隆的TCR对组织特异自身抗原的亲和力低, 且浓度很低时,经APC提呈,不足以活化相应的T细胞, 这种自身应答T细胞克隆与相应组织抗原并存,在正常 情况下,不引起自身免疫病的发生,称为免疫忽视 (immunological ignorance)。
抗原剂量与免疫耐受
T细胞和B细胞耐受比较
• T细胞耐受易于诱导,所需抗原剂量低,耐受持 续时间长(数月-数年)
• B细胞耐受,所需抗原剂量较大,持续时间短 (数周) 抗原性质
• 蛋白单体易诱导免疫耐受,可溶性抗原较颗粒性 抗原诱导免疫耐受。 抗原免疫途径
• 口腔、静脉注射解聚的抗原容易诱导免疫耐受
免疫耐受实验的诱导
(3)免疫耐受诱导条件 ① 抗原因素
二、 免疫耐受的诱导条件和形成机制 胚胎发育期形成的免疫耐受原则上可长期持续,不会轻
易打破。而后天形成的耐受,可持续一段时间,可随诱导因 素的消失而解除。
1. 记忆性T细胞的特征 ① Tm(memory T cell)比Tn( naïve T cell)更易激活,所需
抗原浓度较低; ② Tm对协同刺激信号的依赖性较Tn低; ③ Tm分泌的CK更多,且对CK更敏感; ④ Tn寿命短,Tm寿命长, CK(IL-15)维持其生存; ⑤ 记忆CD8+T细胞的维持无需CD4+T细胞辅助。
中枢耐受
(2)外周耐受
① 克隆清除和免疫忽视
• 克隆清除是指T细胞克隆的TCR对组织特异自身抗原有 高亲和力,且浓度高时,经APC提呈,可致此类T细胞 克隆清除(delection)。
• 若T细胞克隆的TCR对组织特异自身抗原的亲和力低, 且浓度很低时,经APC提呈,不足以活化相应的T细胞, 这种自身应答T细胞克隆与相应组织抗原并存,在正常 情况下,不引起自身免疫病的发生,称为免疫忽视 (immunological ignorance)。
抗原剂量与免疫耐受
T细胞和B细胞耐受比较
• T细胞耐受易于诱导,所需抗原剂量低,耐受持 续时间长(数月-数年)
• B细胞耐受,所需抗原剂量较大,持续时间短 (数周) 抗原性质
• 蛋白单体易诱导免疫耐受,可溶性抗原较颗粒性 抗原诱导免疫耐受。 抗原免疫途径
• 口腔、静脉注射解聚的抗原容易诱导免疫耐受
免疫耐受实验的诱导
(3)免疫耐受诱导条件 ① 抗原因素
第4章第2节特异性免疫-2024-2025学年高二生物选择性必修1(配人教版)配套课件

生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
2.记忆细胞与二次免疫反应的特点 (1)记忆细胞的特点:可以在抗原消失后存活几年甚至十几年。 (2)二次免疫反应:当相同抗原再次入侵时,记忆细胞能迅速增殖分化, 分化后快速产生大量抗体,迅速、高效地产生免疫反应。
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
1.下图是体液免疫的部分过程模式图,下列相关叙述正确的是( D )
A.图中物质 Ⅰ 和物质 Ⅱ 都能与病原体特异性结合 B.病原体被细胞①特异性结合并处理后会暴露出它特有的抗原 C.与细胞③相比,细胞④含特有的与分泌物质 Ⅱ 相关的细胞器 D.细胞③活化除了需要病原体和细胞②两个信号的刺激外,还需要 物质 Ⅰ 的作用
提示 B细胞和细胞毒性T细胞的活化离不开辅助性T细胞的辅助,可 见辅助性T细胞在体液免疫和细胞免疫中都起着关键的作用。
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
1.体液免疫与细胞免疫的比较
项目
体液免疫
细胞免疫
B 细胞、抗原呈递细胞、 靶细胞、细胞毒性 T 细胞、抗原
参与细胞 辅助性 T 细胞、浆细胞、 呈递细胞、辅助性 T 细胞、记忆
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
3.结合体液免疫过程思考:体液免疫和细胞免疫针对某种病原体时 的独特作用是什么?
提示 体液免疫中产生的抗体,能消灭细胞外液中的病原体;消灭侵 入细胞内的病原体,要靠细胞免疫将靶细胞裂解,使病原体释放出来。
4.结合体液免疫过程思考:有人说,辅助性T细胞在免疫调节过程 中起着关键的调控作用。你认同这一观点吗?请说出你的理由。
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
2.记忆细胞与二次免疫反应的特点 (1)记忆细胞的特点:可以在抗原消失后存活几年甚至十几年。 (2)二次免疫反应:当相同抗原再次入侵时,记忆细胞能迅速增殖分化, 分化后快速产生大量抗体,迅速、高效地产生免疫反应。
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
1.下图是体液免疫的部分过程模式图,下列相关叙述正确的是( D )
A.图中物质 Ⅰ 和物质 Ⅱ 都能与病原体特异性结合 B.病原体被细胞①特异性结合并处理后会暴露出它特有的抗原 C.与细胞③相比,细胞④含特有的与分泌物质 Ⅱ 相关的细胞器 D.细胞③活化除了需要病原体和细胞②两个信号的刺激外,还需要 物质 Ⅰ 的作用
提示 B细胞和细胞毒性T细胞的活化离不开辅助性T细胞的辅助,可 见辅助性T细胞在体液免疫和细胞免疫中都起着关键的作用。
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
1.体液免疫与细胞免疫的比较
项目
体液免疫
细胞免疫
B 细胞、抗原呈递细胞、 靶细胞、细胞毒性 T 细胞、抗原
参与细胞 辅助性 T 细胞、浆细胞、 呈递细胞、辅助性 T 细胞、记忆
生物学 选择性必修1 稳态与调节 (配人教版)
第4章 免疫调节
3.结合体液免疫过程思考:体液免疫和细胞免疫针对某种病原体时 的独特作用是什么?
提示 体液免疫中产生的抗体,能消灭细胞外液中的病原体;消灭侵 入细胞内的病原体,要靠细胞免疫将靶细胞裂解,使病原体释放出来。
4.结合体液免疫过程思考:有人说,辅助性T细胞在免疫调节过程 中起着关键的调控作用。你认同这一观点吗?请说出你的理由。
生物学 选择性必修1 稳态与调节 (配人教版)
2022-2023学年 人教版 选择性必修一 特异性免疫 课件 (61张)

3.二次免疫反应 记忆细胞可以在体内存活几年到几十年,如果再次接触相同的抗原, 它们就会立即增殖分化为___细__胞__毒__性__T_细__胞____,____迅__速__、__高__效_____地 产生免疫反应。
微思考 细胞毒性T细胞怎样才能将靶细胞裂解? ______________________________________。 【答案】细胞毒性T细胞受到抗原刺激后,在辅助性T细胞分泌的 细胞因子的作用下,才能使靶细胞裂解
课堂 ·重难探究
体液免疫
[知识归纳] 1.体液免疫的作用对象 细胞外的病原体和毒素。 2.抗体 (1)产生的细胞:浆细胞。浆细胞的产生需要两个信号的刺激,还需 要细胞因子的作用。
(2)化学成分:蛋白质(免疫球蛋白)。 (3)特点:每一种抗体的结合位点只能与一种抗原匹配。 (4)功能:使病毒一类的抗原失去进入寄主细胞的能力;使一些细菌 产生的毒素被中和而失效;使一些抗原凝聚沉淀进而被免疫细胞吞噬消 化。
(2)细胞毒性T细胞分裂分化。 细胞毒性T细胞分裂并分化,形成_新__的__细__胞__毒__性__T_细__胞__和__记__忆__T_细__胞__。 辅助性T细胞活化后分泌的___细__胞__因__子____能加速这一过程。 (3)新形成的细胞毒性T细胞裂解靶细胞。 新形成的细胞毒性T细胞在体液中循环,它们可以_识__别__并_接__触__、__裂__解_ 被同样_病__原__体__感__染___的靶细胞。靶细胞裂解后,病原体失去寄生的基础。
[对点精练]
1.如图表示人体通过体液免疫消灭破伤风杆菌外毒素的过程,下
列叙述错误的是
()
A.细胞2、细胞3均起源于造血干细胞 B.细胞1可特异性识别抗原
C.②③过程与细胞膜上的蛋白质有关 D.细胞5属于保留分裂能力的细胞 【答案】B 【解析】细胞2是辅助性T细胞,细胞3是B细胞,两者均起源于造 血干细胞,A正确;细胞1为抗原呈递细胞,不能特异性识别抗原,B错 误;②和③过程是细胞之间的识别和传递,和细胞膜上的糖蛋白有关, C正确;细胞5是记忆B细胞,相同抗原再次入侵时,记忆B细胞能很快 分裂、分化产生浆细胞,D正确。
微思考 细胞毒性T细胞怎样才能将靶细胞裂解? ______________________________________。 【答案】细胞毒性T细胞受到抗原刺激后,在辅助性T细胞分泌的 细胞因子的作用下,才能使靶细胞裂解
课堂 ·重难探究
体液免疫
[知识归纳] 1.体液免疫的作用对象 细胞外的病原体和毒素。 2.抗体 (1)产生的细胞:浆细胞。浆细胞的产生需要两个信号的刺激,还需 要细胞因子的作用。
(2)化学成分:蛋白质(免疫球蛋白)。 (3)特点:每一种抗体的结合位点只能与一种抗原匹配。 (4)功能:使病毒一类的抗原失去进入寄主细胞的能力;使一些细菌 产生的毒素被中和而失效;使一些抗原凝聚沉淀进而被免疫细胞吞噬消 化。
(2)细胞毒性T细胞分裂分化。 细胞毒性T细胞分裂并分化,形成_新__的__细__胞__毒__性__T_细__胞__和__记__忆__T_细__胞__。 辅助性T细胞活化后分泌的___细__胞__因__子____能加速这一过程。 (3)新形成的细胞毒性T细胞裂解靶细胞。 新形成的细胞毒性T细胞在体液中循环,它们可以_识__别__并_接__触__、__裂__解_ 被同样_病__原__体__感__染___的靶细胞。靶细胞裂解后,病原体失去寄生的基础。
[对点精练]
1.如图表示人体通过体液免疫消灭破伤风杆菌外毒素的过程,下
列叙述错误的是
()
A.细胞2、细胞3均起源于造血干细胞 B.细胞1可特异性识别抗原
C.②③过程与细胞膜上的蛋白质有关 D.细胞5属于保留分裂能力的细胞 【答案】B 【解析】细胞2是辅助性T细胞,细胞3是B细胞,两者均起源于造 血干细胞,A正确;细胞1为抗原呈递细胞,不能特异性识别抗原,B错 误;②和③过程是细胞之间的识别和传递,和细胞膜上的糖蛋白有关, C正确;细胞5是记忆B细胞,相同抗原再次入侵时,记忆B细胞能很快 分裂、分化产生浆细胞,D正确。
4.2 特异性免疫-课件

一.免疫系统是如何识别病原体的?
每种淋巴都能识别各种不同类 型的抗原吗?
克隆选择学说:
1.一个淋巴细胞只针对一种特异性抗原。
2.与抗原对应的淋巴细胞选择性增殖。
。
这种机制有什么优点呢?
体液免疫的研究历程
19世纪末,科学家贝林发现血清中存在抗 毒素,他将这种物质命名为“抗体”。他因此 获1901年诺贝尔生理学或医学奖。
2.体液免疫能够消灭所有侵入体内的新冠病毒吗? 不能。有些新冠病毒寄生在细胞内,体液免疫无法消灭它们。 需另一种特异性免疫机制消灭细胞内部的新冠病毒等病原
体——细胞免疫。
体液免疫
小结
免疫细胞通过受体 识别抗原
体液免疫 研究历程
体液免疫过程
免疫系统对病 原体的识别
特异性 免疫
体液免疫
细胞免疫的研究历程
1890年,德国学者埃利希提出原始的体 液免疫学说,认为血清中存在的抗菌物质在抗 感染免疫中起决定作用。因此,埃利希荣获 1908年诺贝尔生理学或医学奖。
体液免疫:主要依靠抗体“作战”的特 异性免疫方式。
体液免疫的研究历程
1948年,法格雷乌斯证明抗体是受抗原刺激后,由淋巴细胞转化成的 浆细胞(效应B细胞)产生的。
触靶细胞来“作战”的特异性免疫方式。
细胞免疫过程动画
网络资料
一.绘制细胞免疫的流程图
①
病原体(如新冠病毒)
③
② 细胞因子
④
⑥ ⑤
1.细胞毒性T细胞的作用
具有效应功能的细胞毒性T细胞能 识别并裂解被同样病原体感染的靶细胞或 癌细胞等。
细胞毒性T细胞
靶细胞
穿孔
靶细胞
细胞毒性T细胞
细胞免疫
免疫学发展史
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
记忆性细胞的产生
记忆性B细胞的产生 记忆性B细胞在生发中心形成 部分活化的B细胞停留在终末分化期,并
失去分泌IgG的能力而成为记忆性B细胞 记忆性B细胞的产生机制与抗凋亡因子的
持续表达、抗原持续刺激和多克隆刺激剂 的作用有关
免疫记忆的维持
抗原的作用 特异性抗原的持续刺激 低水平抗原的周期性刺激 抗原抗体复合物的作用
记忆性T细胞
中枢性记忆性T细胞 具有较强TCR信号和较低的共刺激信号活 化阈,产生细胞因子的量多
效应性记忆性T细胞 具有直接胞毒活性,可产生大量的细胞因 子和穿孔素
记忆性细胞的产生
记忆性T细胞的产生 中枢性记忆性T细胞,来源于刚刚活化的初
始T细胞,因IL-2水平较低或存在高水平 IL-15而分化为记忆性T细胞 效应性记忆性T细胞,来源于部分效应性T 细胞,因难以启动AICD而存活分化为记 忆性T细胞
一个B细胞只能表达其中一种κ链或链。
免疫球蛋白类别转换
(class switching,isotype switching)
类别转换抗原激活B细胞后,膜上表达和分 泌的Ig类别会从IgM转换成IgG、IgA、 IgE等其它类别或亚类的Ig,即Ig的V区不 变C区发生转换,这种现象称类别转换。
膜型Ig和分泌型Ig重链基因重排
▪翻译为重链( β、δ链)或轻链( α、γ链)
Ig轻链基因的重排
Vk
Jk
Ck
Germline
Rearranged 1° transcript
Spliced mRNA
Ig重链V区基因重排
VH 1-123
DH1-27
JH 1-9
Cm
Ig重链C区基因重排
Primary transcript RNA
Cm AAAAA
关于特异性免疫应答的特点及 其机制
特异性免疫应答的特点
➢特异性(specificity) ➢获得性(adaptiveness) ➢排他性(discrimination) ➢多样性(diversity) ➢记忆性(memory) ➢转移性(transferability) ➢耐受性 (immunotolerance)
存在着分隔的、一定数量的胚系基因片段,在淋
巴细胞分化成熟过程中,这些基因片段发生不同
的重排和组合。
BCR胚系基因结构
V区基因和C区基因
➢重链V区基因由三种胚系基因片段:V、D、
J拼接而成
➢轻链V区基因由V、J两个基因片段拼接成
的
➢V区基因的下游是编码C区的C基因
TCR胚系基因结构
➢β、δ链相当于Ig重链基因,由V、D、
免疫应答的特异性
➢特异性表现在:
应答的启动、应答的对象和应答的结局
➢特异性的本质是:
TCR和BCR特异性识别抗原,而TCR和BCR 的多样性是保证机体对种类繁多的抗原产生 特异性应答的基础
TCR和BCR的多样性及其分子基 础
机体为何能产生数量巨大、识别抗原特异性各异 的TCR和BCR? 其遗传学基础是T、B淋巴细胞在发育早期阶段
免疫耐受与免疫抑制的比较
免疫耐受
免疫抑制
产生原因
细胞系消失或不活化 免疫活性细胞发育缺 Ts细胞的抑制作用 损或增殖分化障碍
产生条件 特异性
可先天或后天获得, 特别是免疫功能未损或人为产生, 如X-射线,免疫抑制 药物,抗淋巴细胞血 清的作用
无
持续性
长期,一时性或终生
胚胎期嵌合体形成中的耐受
Owen于1945年首先报 道了在胚胎期接触同 种异型Ag所致的免疫 耐受现象
红细胞嵌合体
免疫耐受(immune tolerance)
指机体免疫系统在接触某种抗原后产生 的特异性免疫无反应状态 其本质是对抗原特异应答的T与B细胞, 在抗原刺激下,不能被激活产生特异免疫 效应细胞,从而不能执行正免疫应答效应 的现象。
pAm
Cm3
Cm4
Membrane coding
sequence
基因的重排的机制
①重组及重组信号序列 ②重组酶—特异性识别并切除RSS
等位排斥和同种型排斥
等位排斥 指B细胞中位于一对染色体上的轻链
或重链基因,其中只有一条染色体上的基因得到表 达。保证一个B细胞只表达一种轻链和一种重链。
同种型排斥 指κ轻链和λ轻链之间的排斥,
发生时间:在转录加工中造成 由两个外显子决定(SC分泌型、MC膜型) 在转录时,若是转录到SC,则为分泌型Ig。 若包括MC,而将SC剪切掉,则为膜型Ig。
二、抗原识别受体多样性产生的机制
组合造成的多样性 (BCR/TCR) 连接造成的多样性 (BCR/TCR) 体细胞高频突变造成的多样性 (BCR)
J、C基因组成
➢α、γ链相当于Ig轻链基因,由V、J、
C基因组成
BCR、TCR基因的重排
▪重排发生在TB细胞在中枢免疫器官发育过程中 ▪BCR和TCR胚系基因中隔开的V、(D)、J基因 片段通过基因片段的重排,形成V(D)J连接重排 后的DNA。 ▪转录初级RNA,剪接形成mRNA(含C基因)
免疫记忆的维持
T细胞激活信号强度的作用 低强度信号诱导中枢性记忆性T细胞形成 强信号促进效应性T细胞形成 在应答晚期,信号强度的降低可使部分效应
细胞转变为效应性记忆细胞 过强信号刺激可导致效应性T细胞发生AICD
免疫记忆的维持
细胞因子的作用 IL-15 IL-4
免疫耐受
一、胚胎期及新生期接触抗原所致免疫耐受
TCR多样性机制与BCR的区别
TCR无体细胞高频突变 TCR的N-核苷酸插入多于BCR
免疫应答的记忆性
可传递记忆-指围产期个体抵抗病原微生 物的保护性记忆
获得性记忆-个体通过感染或接种而再次 接触相同抗原,可产生较初次应答更快、 更强的应答
免疫记忆细胞的生物学特征
记忆性T、B细胞 是指对特异性抗原有记忆能力、寿命较长 的淋巴细胞。当再次遇到相同抗原后,可 迅速活化、增殖、分化为效应细胞。
一时性
Each domain of the H chain is encoded by a
separate exon
Cm1 h Cm2
Secretion coding
sequence
Polyadenylation site (secreted)
pAs
Polyadenylation site (membrane)