沙雷氏Z4菌株Tn5转座突变株的群体淬灭活性筛选体系的建立及优化
【国家自然科学基金】_沙雷氏菌_基金支持热词逐年推荐_【万方软件创新助手】_20140801
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
科研热词 沙雷氏菌 表型分析 群体感应 生物表面活性剂 spsi突变 spli突变 黏质沙雷氏菌 降解 鉴定 菌株 茶尺蠖 肠道细菌 耐盐 绿色荧光蛋白 红色素 石油污染 盐渍化土壤 生物修复 生物信息学 普城沙雷氏菌 定殖 多样性 基因转化 变性梯度凝胶电泳 变形斑沙雷氏菌 内生细菌 产物性质分析 主控基因 一般胁迫反应 rpos 16s rrna序列
53 54 55 56 57 58 59
mtt法 k-b纸片法 dgge clip结构城 2-酮基-d-葡萄糖酸 16srrna基因 (nh4)2so4
1 1 1 1 1 1 1
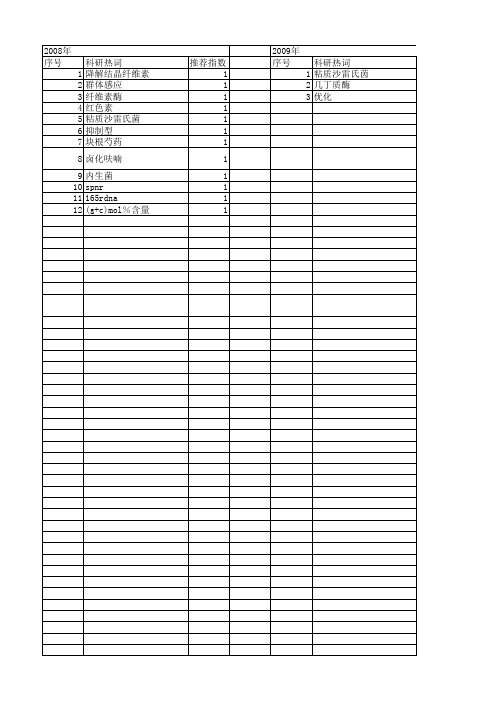
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35
2011年 科研热词 内生细菌 鉴定 重金属 粘质沙雷氏菌 筛选 沙雷氏菌 几丁质酶 黏质沙雷氏菌 铁载体 蕈菌 菌株筛选 苦豆子 脂肪酶 肠杆菌 耐盐植物 结构域 硝吡咯菌素 番茄灰霉病菌 生物防治 生物多样性 生物信息学 泳动运动 根际细菌 普城沙雷氏菌 小麦全蚀病 小rna 基因克隆 土壤微生物 克隆表达 促生 rsmb突变体 mrsa acc脱氨酶 16s-23s rdna间区 16s rdna 推荐指数 3 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
浙江省微生物生化与代谢工程重点实验室

研究条件
重点实验室经过近五年的积累和建设,已有近1000余万的设备投入,有关分子生物学、基因工程、蛋白质工 程、微生物发酵、细胞实验等设备比较齐全;实验室设备(包括部分平台仪器)配备了实时定量PCR仪、多功能 酶标仪、快速高效蛋白纯化工作站(AKTA)、化学发光(ECL)专用成像系统、超声破碎仪(ChIP实验专用)、 激光共聚焦扫描显微镜、扫描电子显微镜、DNA测序仪、Affymetrix芯片系统、凝胶图像系统、高效液相色谱仪、 同位素荧光仪芯片扫描仪、超速冷冻离心机、高效蛋白层析仪、细胞流式仪、荧光倒置显微镜、液相色谱-质谱联 用仪生物生物合成
本方向主要研究内容为(1)酶活性水平调控:合成酶定向进化;(2)催化效率调控:提高固定化微生物细胞通 透性;(3)合成代谢反应调控:利用反应与分离耦合技术,解除底物与产物抑制效应,对合成代谢过程进行调控; (4)针对L(+)-酒石酸、D(-)-酒石酸、L-苹果酸、L-半胱氨酸、异维生素C、D-阿拉伯醇、赤藓糖醇、没食子酸 丙酯等产品,建立催化反应与分离耦合的工业化生产技术。
研究方向
抗生素次级代谢调控
本研究方向全面开展链霉菌抗生素次生代谢调控研究,建立抗生素生产菌基因定向改造技术体系,从源头上 解决抗生素发酵水平问题。主要内容为(1)抗生素生物合成基因簇克隆和生物合成机制研究;(2)深入研究放 线菌次生代谢调控机制,包括途径专一调控因子、多效调控因子(A因子、σ因子、PI因子、严谨响应因子)、 全局调控因子(双组份信号传导体系),通过对染色体基因组目标基因敲除和整合重组,构建高产突变株;(3) 通过放线菌蛋白转运与分泌途径和针对前体物的初级代谢调控机制研究,对基因改造后的突变株进行发酵工艺优 化研究,采用多尺度组合调控抗生素生物合成水平以提高抗生素发酵产量。
利用转座子Tn5诱变冰核细菌获得无冰核活性菌株

利用 转 座子 的插 人 突 变 是 现 代分 子遗 传 学 基
122 受体菌抗利福平突变的筛选 ..
因操作的重要手段。Os 等研究结果表明了冰核 rr e 细菌的成冰核活性是由单个基因所决定的, 该基因 缺失可导致 冰核细 菌丧失冰核活性_2 lJ l。研究 以 云南省冰核细菌 的优势种类 丁香假单 胞菌 Pe- st t dm nsy/ a o oa rge和菠 萝 泛 菌 眦m ∞ ∞L 5为 s n ' ,J 重组基 因工程菌的受体菌 , 采用 T5 n 转座子诱变技 术, 以大肠杆菌 Ec rh i S7p J :T5 s ecl c l 1/Z2 :n 作 h i a o 5 为转座子诱变的供体菌 , 与野生型冰核细菌融合 , 大量筛选失去冰核活性突变株, 供进一步研究和应
维普资讯
第 1 卷 第 1 7 期
2O O 2正
J u a fY n a n utrlUnvri o r l 云南 n Ag c l学报 es y n o u n 农业 大学 a u i t
. 7 N 1 1 o
Ma .2 0 r O 2
m 利福平的 K E B培养基上 , 并涂布均 匀。每 个菌 设 置 3 重 复 。于 2 个 4℃的培 养 箱 中培养 4 , 8h后 测定供体菌对利福平敏感的最低浓度。
收稿 日期 : 0 1 l 4 20 一O 一0
按韦建福和张世斌的方法L 。 7 J 125 供体菌和受体菌接合试验 ( 座子诱变) .. 转
菌悬液 02m , 别 加入 到含 l ,O 3 , ,0u/ . E 分 O 2 ,O柏 5 g
普城沙雷氏菌silI及spsI基因突变株的构建

关键 词 : 沙雷氏菌; 群体感应 ; s p l I突变 ; s p s I突变; 表 型分析 中图分类号 : Q 7 8 文献标识码 : A 文章编号 : 2 0 9 5—1 7 3 6 ( 2 0 1 3 ) 0 1 — 0 0 0 5— 0 4
Co n s t r u c t i o n o f mut a n t s d e ic f i e n t i n s p/ /a n d s ps I i n Se r r a t i a pl y mu t h i c a
菌。 目前对 S .p l y mu t h i c a G 3的两个群体 感应 系统 s p l I / s p l R与 S p s R / S p s I 的 了解仍十分有 限。构建 了 2个 Ⅳ . 乙酰 基 高丝氨 酸 内酯信号合成酶 编码基 因 s p l I和 s p s l突变 菌株 及互补 菌株 , 并检 测 了其生物表 型。结果发 现 s p s I 、 s p l l
e t y o f p h y t o p a t h o g e n i c f u n g i d u e t o i t s a b i l i t y t o p r o d u c e s e v e r a l a n t i f u n g a l f a c t o r s ,a s w e l l a s p l a n t a u x i n i n d o l e 一 3 一 a c e t i c a c i d( I A A) . S 0 f a r , o u r k n o w l e d g e a b o u t t h e t w o Q u o r u m s e n s i n g( Q s )s y s t e m s( S p l I / S p l R a n d S p s R / S p s I )i n S . p l y mu t h i c a G 3 i s s t i l l v e r y l i m i t - e d .I n t h i s s t u d y , s t r a i n G 3 w a s u s e d a s t h e m o d e l o r g a n i s m, s ll i a n d s p s l mu t a n t s i n G 3 t h r o u g h g e n e — r e p l a c e me n t s t r a t e g y w e r e c o n - s t uc r t e d .T h e p h e n o t y p i c a n ly a s i s o f G 3 a n d i t s d e r i v a t i v e s r e v e a l e d t h a t mu t a t i o n o f s p s I a n d s ll i s l i g h t l y a f f e c t e d t h e b i o s y s t h e s i s o f b a c t e r i a l s i g n a l mo l e c u l e s N - a c y l h o m o s e r i n e l a c t o n e s( A H L s ) , H o w e v e r , t h e s e t o w m u t a n t s s i g n i f i c a n t l y d e c r e a s e d t h e p r o t e a s e a c t i v i t y i n c o m p a r i s i o n w i t h t h e w i l d t y p e G 3 .F u r t h e r mo r e , s w i m mi n g a s s a y o f t h e m u t a n t s a n d t h e c o mp l e me n t a y r s t r a i n s s h o w e d t h a t s ll i n e g —
粘质沙雷氏菌(Serratia marcescens)几丁质酶基因克隆的筛选及序列分析

粘质沙雷氏菌(Serratia marcescens)几丁质酶基因克隆的筛
选及序列分析
张表;赵晓瑜;乔环宇
【期刊名称】《河北大学学报(自然科学版)》
【年(卷),期】2003(023)002
【摘要】用改进方法提取粘质沙雷氏菌染色体DNA.通过PCR扩增,得到几丁质酶(chiA)基因,利用pUC19质粒构建了含有chiA基因的克隆载体 ,并转化E.coli DH5α.经测序分析,证明克隆片段与文献报道基本相同,仅在第437位碱基由C变为T.
【总页数】4页(P184-187)
【作者】张表;赵晓瑜;乔环宇
【作者单位】河北大学,生命科学学院,河北,保定,071002;河北大学,生命科学学院,河北,保定,071002;河北大学,生命科学学院,河北,保定,071002
【正文语种】中文
【中图分类】Q785
【相关文献】
1.粘质沙雷氏菌几丁质酶(ChiC)基因克隆及其生物信息学分析 [J], 魏巍;贺淹才;方柏山;刘爱花
2.粘质沙雷氏菌(Serratia marcescens)的研究Ⅰ-Serratia marcescens 9-2菌株分离、分类鉴定和形态特征 [J], 黄文芳
3.嗜甲基细菌Serratia Marcescens NH8的r筛选及其pqq基因的克隆 [J], 彭乔木;周璐璐;熊力;彭清忠;胡广
4.粘质沙雷氏菌(Serratia marcescens)研究Ⅱ--营养基质和不同色光对粘质沙雷氏菌生长的影响 [J], 黄文芳;诸瑜
5.粘质沙雷氏菌(Serratia marcescens)发酵培养基优化的研究 [J], 郝林华;陈靠山;牛德庆;张玉凤
因版权原因,仅展示原文概要,查看原文内容请购买。
【国家自然科学基金】_tn5转座子_基金支持热词逐年推荐_【万方软件创新助手】_20140801

2011年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
2011年 科研热词 转座子 胞外多糖 突变体库 磷酸乙酰转移酶 反刍月形单胞菌 副猪嗜血杆菌 减毒株 乙酸激酶 青枯雷尔氏菌 遗传改良 突变 短小芽孢杆菌 电击转化 生物防治 甜瓜采后病害 水稻条斑病菌 毒性 无致病力突变株 基因位点 反向pcr ttc培养基 tn5转座子 tn5插入突变体 2,4-二乙酰基间苯三酚 推荐指数 5 2 2 2 2 2 2 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
推荐指数 3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 推荐指数 野油菜黄单胞菌 1 调控 1 表达调控 1 致病相关基因 1 群体感应 1 细菌耐药 1 细菌素 1 电穿孔 1 生物防治 1 甘油-3-磷酸-酰基转移酶 1 大肠埃希菌 1 可转移因子 1 假单胞菌 1 tn5突变 1 ez-tn5 1 4-二乙酰基间苯三酚 1 2 1
2012年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14
科研热词 推荐指数 青枯雷尔氏菌 1 荧光假单胞菌(pseudomonas fluorescens) 1 致病性 1 致病力 1 群体感应 1 突变体 1 水稻基腐菌 1 无致病力突变株 1 弱化指数 1 tn5转座子 1 mvav 1 mvat 1 4-二乙酰基间苯三酚 1 2 1
2013年 序号 1 2 3 4
科研热词 耐盐基因 盐敏感突变株 甜菜碱 中度嗜盐菌
211105505_基于重组减毒沙门氏菌载体递送疫苗及药物的肿瘤免疫治疗研究新进展

中国免疫学杂志2023 年第 39 卷基于重组减毒沙门氏菌载体递送疫苗及药物的肿瘤免疫治疗研究新进展李琦 陈多多 王弘瑞 胡迪 杨明 孙丽媛 (北华大学医学技术学院,吉林132013)中图分类号 R392.11 文献标志码 A 文章编号 1000-484X (2023)03-0670-05[摘要] 沙门氏菌介导的肿瘤免疫治疗方法已经成为抗肿瘤研究领域的热点之一。
首先,沙门氏菌作为兼性厌氧菌,能够特异性地定植在肿瘤区域,直接杀伤肿瘤细胞或诱导机体产生免疫反应,介导包括巨噬细胞、中性粒细胞和T 细胞等在内的免疫细胞浸润从而发挥抗肿瘤效应。
其次,沙门氏菌具有高度遗传修饰性,可被改造为毒力减低或精准靶向的重组沙门氏菌。
此外,重组沙门氏菌还可作为疫苗、细胞毒性药物、RNAi 和纳米颗粒递送的有效平台,在治疗的同时还可预防肿瘤的复发和转移。
如重组沙门氏菌载体活疫苗裂解系统,能够加强肿瘤抗原呈递,促进T 细胞发挥杀伤效应。
本文介绍了减毒沙门氏菌载体的抗肿瘤免疫特性和肿瘤靶向性,并对以重组减毒沙门氏菌为递送系统进行肿瘤免疫治疗的研究进展进行综述。
[关键词] 沙门氏菌;肿瘤;疫苗;细胞毒性药物;RNAiAdvances in vaccine and drug delivery for tumor immunotherapy based on recombinant attenuated Salmonella vectorLI Qi , CHEN Duoduo , WANG Hongrui , HU Di , YANG Ming , SUN Liyuan. School of Medical Technology , Beihua University , Jilin 132013, China[Abstract ] Salmonella -mediated immunotherapeutic approaches for tumors have become one of hot spots in the field of antitu⁃mor research. First , Salmonella , as a parthenogenic anaerobic bacterium , is able to specifically colonize the tumor area. It can also directly kill tumor cells and induce immune responses in body , mediating the infiltration of immune cells such as macrophages , neu⁃trophils and T cells to exert anti -tumor effects. Second , based on the highly genetically modified features , it can be modified to become virulence -reduced or precisely targeted recombinant Salmonella. In addition , recombinant Salmonella could serve as an efficient plat⁃form for the delivery of cancer vaccine , cytotoxic agents , RNAi and nanoparticles , which showed a strong inhibitory effect on growth of cancer , and exhibited a prevention of tumor recurrence and metastasis. For example , a vaccine lysis system based on recombinantattenuated Salmonella vector , which can enhance tumor antigen presentation and promote antitumor effects of T -cell. This reviewdescribes the antitumor immune properties and tumor targeting of Salmonella ,and also elaborate the progress of research on tumor immunotherapy using recombinant attenuated Salmonella as a delivery system.[Key words ] Salmonella ;Tumor ;Vaccines ;Cytotoxic agents ;RNAi传统的肿瘤疗法有手术、化疗和放疗等,但存在靶点选择性差、药物渗透性差和患者不良反应性等缺点[1]。
鸡白痢沙门氏菌Mini-Tn5转座子插入突变

进行筛选,获得大量在不同位点插入突变的突变体,从中筛选鸡白痢沙门氏菌某功能缺陷型突变株。通过对突变
株的基本特征以及PCR鉴定后,再进行插入基因定位。结果表明,Km和Cm抗性基因已成功转座至鸡白痢沙门
氏菌基因组上;基因定位显示突变株均有且只有1个插入位点,插入位点的位置不尽相同。这为研究鸡白痢沙门
氏菌功能基因和筛选特定突变株提供了必要的基础。
失,并对其某种功能进行有针对性的筛选(如肠内黏 附、细胞内生存等),最后通过基因定位鉴定突变株 插入的功能失活基因,从而获得诸多与此功能相关 的突变株[3]。因此突变株的获得对研究细菌的感染 与致病的分子机制十分有用[4-5]。为此,本研究利用 自杀性质粒pUT 携带的mini-Tn5转座子对鸡白痢 沙门氏菌进行转座突变[6-8],构建并鉴定突变株,以
表1 试验用的细菌和质粒 Table 1 Bacteria and plasmids in the research
应条件:94℃5min;94℃5个循环;72℃10min。PCR产物预期大小约
为1.0kb。回收PCR产物连接至pUC18载体中,
转化至E.coli DH5α,获得重组质粒,命名为 pUC-
关键词:鸡白痢沙门氏菌;基因转座;突变;基因定位
中图分类号:S852.61
文献标识码:A
文章编号:0366-6964(2008)05-0621-06
Insertion Mutagenesis in Salmonella pullorum by Mini-Tn5 Transposon
GENG Shi-zhong,JIAO Xin-an*,CHEN Xiao-iuan, PAN Zhi-ming,ZHANG Hui,CHEN Xiang
Cm,经鉴定后送宝生物工程(大连)有限公司测序。
应用转座子Tn5对生防菌P-936进行遗传改良的研究

Ba t ru n P 一9 6 b a s o o Tn ce il 3 y Tr n p s n 5
L a g—mi ,HU IXin n ANG R i o g ,HU J u —rn A u—l g ,L O Re i U n—h a ,L U Pn n u I ig—a n
关键词 : 座子; 防菌; 转 生 遗传 改 良 中图 分 类 号 :4 6 1 文 献 标识 码 : 文 章 编 号 :0 1 8 8 (0 2 0 00 ¥7 .9 A 10 — 5 12 1 )4— 12一o 3
S ud e n Ge e i mpr v m e to o o i a n r l t is o n tc I o e n fBi l g c lCo t o
体茵 , 通过结合转移技术, 将转座子 T 5从供体 菌中引入 到受体菌 P一 3 n 96的基 因组 D A 中, N 共获得 3 1 0 1个 插入 突变体 。 I 在 离体测定中, 4个突变体对稻瘟病 菌的拮抗作 用明显增强, 1 有 有 个突变体 完全丧失 了拮抗作 用; 突变体对番茄青枯病 2个 菌的拮抗作 用明显增强。通过盆栽和 田间小 区试验 , 突变体 T一10 27在连 续 3次防 治后 , 对稻 瘟病 的防治效果达到 6 .7 , 19 % 而野生菌株 P一 3 96的防治效果为 5 .5 , 15 % 表现 了一定的增效作用。
2 gi tr ueuo ninC u t i agi rv c , i in300 , h a .A r u ua B ra f i a ony nJ nx Poi e J xa 340 C i ) c l l Jx i n n n
Ab tac s r t:Usng awi i de—s e tu a tg n si c ei m e do n ∞ r c珊 P 一9 6 a c e e ,a d Esh rc a c l p cr m n a o itcba tru Psu mo as e 3 sa c ptr n c e ihi oi¥1 7
放线菌SL_5链霉素耐药突变菌株的分离_活性筛选及发酵优化研究

第36卷 第12期西北农林科技大学学报(自然科学版)Vol.36No.12 2008年12月Journal of Northwest A&F University(Nat.Sci.Ed.)Dec.2008放线菌SL25链霉素耐药突变菌株的分离、活性筛选及发酵优化研究3沈 玲,郭正彦,吴文君(西北农林科技大学农药研究所,陕西杨凌712100)[摘 要] 【目的】为新型抗生素的开发研究提供有效途径。
【方法】对放线菌SL25在链霉素不同浓度下的耐药突变菌株进行分离和活性筛选,并采用单因素和正交试验对活性菌株进行发酵优化。
【结果】从无活性放线菌SL2 5的耐药菌株中得到1株活性突变菌株str214,室内生测结果表明,其发酵液对蜡状芽孢杆菌(B acill us cereus)的抑菌圈直径为17.5mm。
通过单因素和正交试验,确定该菌株的最佳摇瓶发酵培养基配方为:小米10g/L,葡萄糖20 g/L,酵母粉2g/L,蛋白胨7g/L,混合V B1.5mg/L,NaCl2.5g/L,CaCO32.0g/L。
最佳培养条件为:初始p H为8.0,装液量70mL(250mL三角瓶),180r/min培养7d。
【结论】通过引入链霉素耐药性突变获得抑菌活性物质,是创制新型抗生素的一条可行之路。
[关键词] 放线菌;链霉素耐药突变菌株;分离;发酵优化[中图分类号] S859.79+6[文献标识码] A[文章编号] 167129387(2008)1220205207St udies on t he isolation,activity screening and t he fermentation optimization of streptomycin2resistance mutant s of actinomycete SL25SH EN Ling,GUO Zheng2yan,WU Wen2jun(I nstit ute of Pestici de,N ort hwest A&F Universit y,S haanx i,Yangling712100,China)Abstract:【Objective】The st udies were carried out to provide an efficient way of discovering novel an2 tibiotics.【Met hod】Mutant s were isolated at different st reptomycin concent rations f rom wild st rain SL25 and under bioactivity screening,t hen t he fermentation medium optimization of t he active mutant was st ud2 ied by t he single2factor and ort hogonal test.【Result】St rain st r214was obtained f rom t he st reptomycin2re2 sistance mutant s of inactive st rain SL25,and t he result of bioassay i n vit ro showed t hat t he fermentation brot h of st r214exhibited obvio us inhibition circle of17.5mm against(B acill us cereus).The result s of sin2 gle2factor and ort hogonal test showed t hat t he optimum fermentation of strain st r214for producing t he mo st effective fermentation p roduct were Millet10g/L,Glucose20g/L,Yeast ext ract2g/L,Peptone7 g/L,Compound vitamins B1.5mg/L,NaCl2.5g/L,CaCO32.0g/L.The optimum conditions were initial p H of8.0,media volume70mL,shaking at180r/min for7days.【Conclusion】It is a feasible way for t he discovery of new antibiotics to find active substance by inducing streptomycin2resistance mutation.K ey w ords:actino mycete;st reptomycin2resistance mutant;isolation;fermentation optimization 在医药行业及农业的病虫草害综合治理中,抗生素一直发挥着重要作用。
纳他霉素高产突变菌株高通量筛选方法的建立

纳他霉素高产突变菌株高通量筛选方法的建立
刘陈梦;王雁龙;孙瑞杰;吴康;贾士儒;谭之磊
【期刊名称】《食品研究与开发》
【年(卷),期】2024(45)1
【摘要】为了提高纳他霉素(natamycin)高产菌株诱变后的筛选效率,建立一种24孔深孔板/酶标仪发酵初筛和摇瓶发酵复筛的高通量筛选检测方法,得到在24孔深
孔板发酵结果与摇瓶发酵有很好的一致性,证实了酶标仪检测可以替代高效液相色
谱法检测用于突变菌株的快速高通量筛选。
以褐黄孢链霉菌(Streptomyces gilvosporeus TUST01)作为出发菌株,通过多轮的紫外诱变、常压室温等离子体(atmospheric and room temperature plasma,ARTP)以及硫酸二乙酯复合诱变、孔板初筛与摇瓶复筛,从887株突变株中筛选出遗传性稳定的高产菌株S. gilvosporeus Y-4-75,其摇瓶纳他霉素产量(5.95 g/L)与生物量(29.19 g/L)较出发菌株分别提高了31.93%和15.19%。
【总页数】7页(P147-153)
【作者】刘陈梦;王雁龙;孙瑞杰;吴康;贾士儒;谭之磊
【作者单位】天津科技大学生物工程学院
【正文语种】中文
【中图分类】TS2
【相关文献】
1.青霉素高产菌株高通量检测和筛选方法的研究
2.从氯霉素抗性突变株筛选赤霉素高产菌株
3.链霉素抗性突变理性筛选新霉素高产菌株
4.链霉素抗性突变--纳他霉素高产菌株的选育研究
5.链霉素抗性突变理性筛选链霉素高产菌株
因版权原因,仅展示原文概要,查看原文内容请购买。
利用Tn5转座子构建杆状病毒AcMNPV随机突变体的初步研究

文章编号:042727104(2005)0420498205收稿日期:2005205209基金项目:国家自然科学基金资助项目(30070031)作者简介:李 惠(1979—),男,硕士研究生,通讯联系人钟 江教授.E 2mail :jzhong @.利用T n5转座子构建杆状病毒AcMNPV 随机突变体的初步研究李 惠,赵明磊,尹 隽,钟 江(复旦大学生命科学学院微生物学与微生物工程系,上海 200433)摘 要:以杆状病毒模式种AcMNPV 为研究对象,应用基于Tn5转座子的随机转座的方法,构建杆状病毒突变体库.将果蝇hsp70启动子后接绿色荧光蛋白基因后插入Tn5转座子,构建了可以在昆虫细胞中表达,易于跟踪的转座载体.利用体外转座系统将转座子随机插入AcMNPV 基因组,并用转座反应液转染S f21细胞,得到了表达绿色荧光蛋白的病毒突变体库.进一步纯化了两株病毒B9F 和Li6A ,进行了转座子插入位点的分析,确定两株病毒中,转座子分别插入了94K 基因和p10基因.该方法将为杆状病毒功能基因组研究提供重要的手段.关键词:AcMNPV ;Tn5转座子;转座酶;突变体中图分类号:Q 939.4 文献标识码:A 杆状病毒A utographa calif ornica multicapsid nucleopolyhedrovirus (AcMN PV )是一种应用广泛的真核表达载体1,2和具有潜力的生物杀虫剂3,4,近年来的研究显示它也有可能发展成为一种有效的哺乳动物细胞基因转导和基因治疗的载体5.AcMN PV 基因组长133894bp 6,共有154个潜在的开放阅读框.目前共有约70个基因的功能得到不同程度的研究和鉴定.同时,尚有大量基因的功能有待深入研究.高效地获得各种突变体是分析研究基因功能的重要环节.Tn5是一种细菌的转座子,两端分别带有转座元件IS50R 和IS50L ,可在Tn5转座酶催化下随机插入、整合到目标DNA 序列中,同时带入两端转座元件以内的DNA 序列.本研究构建了带有昆虫细胞表达元件和绿色荧光蛋白基因的转座载体,通过体外转座反应和细胞转染得到了带有随机插入突变的AcMN 2PV 突变库.在此基础上分离并分析了2个突变病毒株.1 材料和方法1.1 细胞、病毒和质粒草地贪夜蛾(S podoptera f rugierda )Sf21细胞系,E.coli DH5α,AcMN PV 1A 株,质粒p HZ402,pAcDZ1均为本实验室保存,转座子构建质粒pMod2购自Epicentre.Sf21细胞用TNM 2FH 培养液加10%胎牛血清培养.细菌、质粒、基因操作参照文献7.1.2 带有昆虫细胞表达框架的T n5转座子的构建以pAcDZ1为模板通过PCR 扩增果蝇hsp70启动子,引物分别为5’2ACA TGCA TGCTA 2G AA TCCCAAAACAAACTG 23’和5’2GG AA TTCTA TTCA G A GTTCTCTTCTTG 23’,反应条件为预变性95℃5min ;然后95℃1min ,56℃30s ,72℃1min 共30个循环;最后72℃延伸10min.扩增得到长度约为500bp 的产物,经电泳、胶回收试剂盒(V 2gene )纯化,并克隆到T 载体(Ta KaRa )上,得到p T 2hsp70p ,序列分析验证其正确.用Nco Ⅰ和S ac Ⅰ双酶切p T 2hsp70p ,分离2.7kb 片段;用同样的酶双酶切p HZ402,胶回收分离800bp 的gf p 基因片段.两片段连接得到p T 2hsp70p 2gfp.以B am H Ⅰ和S ac Ⅰ双酶第44卷 第4期2005年8月复旦学报(自然科学版)Journal of Fudan University (Natural Science )Vol.44No.4Aug.2005切p T 2hsp70p 2gfp ,回收1.3kb 片段,连接到用同样的酶双酶切处理的转座子构建质粒pMod2上,得到pMod 2hsp70p 2gfp (见图1).1.3 AcMNPV D NA 的制备Sf21细胞悬浮培养至约1×106个时(参照文献8),按MO I =1pfu/cell 接种AcMN PV ,48h 后,细胞悬液以5000r/min 离心15min ,取上清液.上清液再以17000r/min 4℃离心30min ,用450μL 无菌水悬浮沉淀.加入50μL 10%的SDS ,56℃30min ;加入蛋白酶K 至100μg/mL ,再次56℃30min.用酚/氯仿法抽提病毒基因组DNA ,乙醇沉淀DNA.以50μL 无菌水溶解DNA ,4℃放置待用.图1 pMod 2hsp70p 2gfp 质粒及转座子示意图Fig.1 Structure of pMod 2hsp70p 2gfp1.4 体外转座以pMod 2hsp70p 2gfp 为模板进行PCR ,引物分别为FPP (5’2A TTCA GGCTGCGCAACTGT 23’)和RPP (5’2GTCA GTG A GCG A GG AA GCGG AA G 23’)(Epicentre ,图1),得到线性转座子DNA 片段.反应条件为:94℃预变性5min ;然后,94℃30s ,55℃30s ,72℃1min 共30个循环;最后72℃延伸10min.PCR 产物纯化后用于进行体外转座反应.转座反应参照Epicentre EZ ∶∶TN 转座酶说明书进行,10μL 反应液包括2μL AcMN PV DNA ,6μL 线性转座子序列,1μL 10×buffer 和1μL EZ ∶∶TN 转座酶.37℃反应过夜.1.5 细胞转染5×105个Sf21细胞27℃静置培养2h ,使细胞贴壁.取5μL 上述转座反应液,用Cellfectin (Invitro 2gen )转染细胞,方法参见产品说明书.转染后细胞27℃培养,Nikon HFX 倒置荧光显微镜观察.转染后72h ,收集细胞的培养上清液,作为重组病毒库的原代病毒液,4℃保存备用.1.6 突变体病毒株的分离通过空斑试验进行.六孔细胞培养板中每孔加入5×105个Sf21细胞,27℃静置2h 后,吸去上清液,加入适当稀释度的病毒液1mL ,27℃感染3h 后,吸去病毒液,同时将3%低熔点琼脂糖融解,与两倍体积的细胞培养液混合,加入到各孔中,每孔2mL.静置培养1h ,待琼脂糖凝固后,每孔再加入1mL 培养液.27℃培养3~5d ,至空斑出现.挑取空斑,悬浮于培养液中.如此重复空斑试验数轮,得到纯化的病毒株.图2 PCR 确定转座子的插入位点示意图Fig.2 Primers for the determination of transposon interstion site via PCR1.7 转座子插入位点分析沿病毒基因组每隔3kb 左右设计一个引物,共44个引物(Ac1244,图2,表1),同时根据转座子序列设计向上游和向下游的引物(Mod 2up 和Mod 2down ,图2,表1).分别以Mod 2up 、Mod 2down 与Ac1244组成引物对,共88对,进行PCR 反应.Mod 2up 或Mod 2down 可与转座子插入位点附近的一个引物产生PCR 反应产物(图2).与野生型病毒对照,并通过对PCR 产物的序列分析,确定转座子的插入位点.2 结 果2.1 携带昆虫细胞表达元件的转座子的构建PCR 扩增hsp70启动子,并接到绿色荧光蛋白基因(gf p )上游,然后再将该表达框插入转座子构994 第4期李 惠等:利用Tn5转座子构建杆状病毒AcMNPV 随机突变体的初步研究建质粒pMod2,得到带有昆虫细胞表达框架的Tn5转座子的质粒pMod 2hsp70p 2gfp (图1).将质粒转染细胞后,荧光显微镜观察可见部分细胞产生微弱绿色荧光(图未显示),表明该hsp70启动子/gfp 表达框架可以在昆虫细胞中表达.表1 用于确定转座子位点的PCR 引物的序列和在病毒基因组上的位置Tab.1 The nucleotide sequences and positions in AcMNPV genome of primers used for the determination oftransposon instertion site via PCR名称序列(5’———3’)起点位名称序列(5’———3’)起点位Ac 21tgtggaccgcagaacagata 627Ac 224cgaatatggacctaacaacc 71362Ac 22aaggctctgacgcatttcta 3545Ac 225tgtggtaatagtggcgttgg 74761Ac 23cacaacggaaggtcgtctgc 6445Ac 226ccaacaaccgagttagagta 77805Ac 24atggattgcgagtatttgcg 9338Ac 227cacggcaatacctatcatct 80922Ac 25tacgcaaggcggactacaat 12520Ac 228cgttactttccaacacccag 84065Ac 26gacgcaacacgactacactg 15473Ac 229gcaaacgacgaccgcataat 87366Ac 27cggcatcaacgagccaactt 18365Ac 230ctgaatagcgatgctgatgc 90449Ac 28ctcctccgaaggtccgtcta 21327Ac 231gcttactgtgcctgtatcaa 93342Ac 29ccagttcaacaatccctctt 24462Ac 232ttgcgagaccgtcaacataa 96487Ac 210ctccgtctggatttactgcc 27451Ac 233ctcggtgttcccgtatcgtc 99366Ac 211cgatgacctcgtggtatgga 30866Ac 234caagggcaacaaatagacgc 102229Ac 212gagaatagccgtcgccacaa 33991Ac 235gcatcaatctcccaagcaaa 105343Ac 213ccaccactaccaacaacaac 36884Ac 236cccttctttgtagatgctgt 108545Ac 214acccttcttggaacacgaca 39922Ac 237agactcgttacccgacttga 111566Ac 215aatctgccgtccagcataaa 42864Ac 238tccgagacataccacaaagc 114765Ac 216gcagaaagcgatagtgaaag 45756Ac 239tggctcataactaaactcgc 117940Ac 217agcctgctgtcgtgaatacc 48985Ac 240gcggcacataataatcgtcg 120913Ac 218aaccgctgtcgtaatcttgg 52227Ac 241cgcaagatgatggctttcct 124001Ac 219tgacgcacaacatcaactac 55542Ac 242gttcgccattagggcagtat 127099Ac 220cggctcaccgctactttctc 58834Ac 243aaaactgccgtcgtcaatac 130185Ac 221cgtttagggattctatggtg 62188Ac 244gaacggagcgtgattagtgt 133066Ac 222tcgtcgtgttgtcatagccc 65122Mod 2down acgactacgcactagccaaca NA Ac 223cgacctttccacctatcacg68114Mod 2uptcggcatggacgagctgtacNA图3 PCR 检测病毒突变株中的hsp70启动子Fig.3 PCR detection of the hs p70promoter in the mutant virus 1.DNA 分子质量标准(λDNA Hi n d Ⅲ/Eco R Ⅰ);2.B9F 突变病毒株DNA 为模板;3.Li6A 突变病毒株DNA 为模板;4.野生型AcMNPV DNA 为模板.2.2 AcMNPV 突变体库的构建PCR 扩增pMod 2hsp70p 2gfp 中转座子序列,得1.4kb 的线性化片断.将病毒基因组DNA 和该转座子片段混合,在EZ ∶∶TN 转座酶的催化下,进行体外转座.转座反应液直接用于转染Sf21细胞.一天后在荧光显微镜下观察到绿色荧光.由此得到随机插入突变病毒库.对原代病毒液进行效价测定,达2×106pfu/mL ,其中产生绿色荧光的约占20%.将该原代病毒再次感染Sf21细胞,仍然观察到细胞发出荧光,证明转座子已经成功插入病毒基因组.2.3 突变病毒株的纯化空斑试验和多孔板稀释法从突变病毒库中分离得到了2株突变病毒株,分别为B9F 和Li6A.抽提病毒基因组DNA ,并以之为模板,用扩增hsp70启动子的引物进行PCR 检测,可以从突变病毒株的DNA 中扩增得到hsp70启动子片断,以野生型病毒DNA 为模板则不能得到该片断(图3).证明突变病毒株中带有插入的转座子.2.4 转座子插入位点的确定沿AcMN PV 基因组顺时针方向每隔3kb 左右设计1个寡聚核苷酸(Ac12Ac44,表1),分别以它们和根据转座子内部序列设计的寡聚核苷酸Mod 2up 和Mod 2down 作为引物,以突变病毒株的基因组DNA 为模板进行PCR ,对阳性的PCR 产物进行序列分析,从而确定了两株病毒中转座子的插入位点,结果如图4所示.这两个突变05 复旦学报(自然科学版)第44卷 株中,转座子分别插入并破坏了AcMN PV 的94K 基因和p10基因.B9F 突变株中,转座子插入位点位于94K 基因编码区,置换了基因组中114585至115089bp 之间的区域(图4A ),而Li6A 突变株中,转座子的插入位点位于AcMN PV 的p10基因编码区的119057bp 位点(图4B ).表明这两个基因都是病毒在体外细胞中复制非必需的位点.图4 B9F 和Li6A 突变株转座子插入位点示意图Fig.4 Transposon insertion sites of the mutant AcMNPV B9F and Li6A3 讨 论杆状病毒基因组学和基因功能的研究已经取得了很大的进展,对数十种病毒的基因组进行了全序列的分析,对其中的相当部分基因,特别是对于那些在病毒感染和复制中具有关键作用的基因进行了转录表达和功能的分析9.但由于杆状病毒的基因组庞大,其中一半左右的病毒基因的功能尚未有深入研究.获得大量的突变体病毒是研究病毒基因功能的重要途径之一.随机位点插入转座子在帮助人们分离大量基因功能破坏的突变方面发挥了重要的作用,已经被用在研究一些微生物乃至高等生物的基因组中10~13.杆状病毒的基因组是双链DNA ,具有感染性,DNA 转染细胞后就能开始复制,产生子代病毒.因此,杆状病毒非常适合用体外随机转座的方法获得大量的突变病毒,但目前国内外尚未有这方面研究的报道.本研究利用转座子的方法获得了一个随机突变AcMN PV 库,初步从中分离了两个突变病毒株,B9F 和Li6A ,确定了这两个病毒分离株中转座子插入位点分别为病毒的94K 基因和p10基因.AcMN PV 94K 基因是一种早期表达的基因,其基因功能还不清楚,对病毒感染细胞非必需14.p10基因则是一个在病毒感染极晚期高水平表达的基因,与病毒感染的细胞中纤维状结构有关,但对于病毒在体外培养细胞中的感染与复制也是非必需的15.这表明可以利用该方法确定病毒的非必需基因.由于破坏必需基因功能的突变会导致无法获得纯的突变株,因此尚不能以本文报道的方法进行研究,需要设计其他的研究手段,如通过改变培养条件,增加异源辅助病毒共同感染,或借助病毒全基因组的BAC (细菌人工染色体)克隆先在细菌中获得必需基因突变的病毒基因组的方法.这些研究还在进行中.利用转座子随机突变技术研究杆状病毒基因功能的工作可以帮助我们更加深入的认识病毒复制的机制,认识病毒与细胞及宿主昆虫之间的相互作用,从而为更好地利用这一病毒提供理论基础.如何更加高效地获得病毒分离株,并确定转座子的插入位点,还需要通过进一步的研究加以解决.参考文献:1 Possee R D.Baculoviruses as ex pression vectorsJ .Curr Opin Biotechnol ,1997,8:569272.2 Vialard J E ,Arif B M ,Richardson C D.Introduction to the molecular biolo gy of baculovirusesJ .Methds MolBiol ,1995,39:1224.3 Cunningham J C.Baculovirus as microbial insecticides M .In “Novel Approached to Integrated Pest Manage 2ment ”(R Reuveni Ed.).Boca Raton :CRC Press ,1995.2612292.4 Black B C ,Brennan L A ,Dierks P M ,et al .Commercialization of baculoviral insecticides M .In “The Bac 2uloviruses ”(L ler ,Ed ).New Y ork :Plenum Press ,1997.3412387.5 Ghosh S ,Parvez M K ,Banerjee K ,et al .Baculovirus as mammalian cell ex pression vector for gene therapy :an105 第4期李 惠等:利用Tn5转座子构建杆状病毒AcMNPV 随机突变体的初步研究emerging strategyJ .Mol Ther ,2002,6:5211.6 Ayers M D ,Howard S C ,Kuzio J ,et al .The complete DNA sequence of A utographa calif ornica nuclear poly 2hedrosis virusJ .V irology ,1994,202:5862605.7 Ausubel F M ,Brent R ,K ingstonR E ,et al .Short proptocols in molecular biology M .3rd ed.New Y ork :JohnWiley &S on ,Inc.,1995.(in Chinese )8 朱媛娥,徐慧斌,赵志安,等.昆虫杆状病毒表达系统表达人载脂蛋白A 2I J .生物工程学报,2003,19:6922697.9 Herniou E A ,Olszewski J A ,Cory J S ,et al .The genome sequence and evolution of baculovirusesJ .A nn RevEntomol ,2003,48:2112234.10 Yu D ,Silva M C ,Shenk T.Functional ma p of human cytomegalovirus AD169defined by global mutational anal 2ysisJ .PNA S ,2003,100:123962401.11 Carlson C M ,Dupuy A J ,Fritz S ,et al .Transposon mutagenesis of the mouse germlineJ .Genetics ,2003,165:243256.12 Otero J ,Jacobs W R J r ,G lickman M S.E fficient allelic exchan ge and transposon mutagenesis in Mycobacteriumavium by specialized transductionJ .A ppl Environ Microbiol ,2003,69:503925044.13 Castano I ,K aur R ,Pan S ,et al .Tn72based genome 2wide random insertional mutagenesis of Candida glabrataJ .Genome Res ,2003,13:905215.14 Friesen P D ,Miller L K.Divergent transcription of early 352and 942kilodalton protein genes encoded by theHin dIII K genome fragment of the baculovirus A utographa calif ornica nuclear polyhedrosis virusJ .J V irol ,1987,61:226422272.15 Vlak J M ,K linkenberg F A ,Zaal KJ ,et al .Functional studies on the p10gene of A utographa Calif ornica nu 2clear polyhedrosis virus using a recombinant expressing a p102beta 2galactosidase fusion gene J .J Gen V irol ,1988,69:765276.Preliminary Studies on the Construction of T n 5T ransposonB ased on R andom Insertional Mutants of B aculovirusLI Hui ,ZH AO Ming 2lei ,YI N J uan ,ZHONG Jiang(Depart ment of Microbiology and Microbial Engineering ,School of L if e Sciences ,Fudan U niversity ,S hanghai 200433,China )Abstract :AcMNPV ,the type species of baculovirus ,was used to construct a transposon based on mutant library using a Tn5random transposon vector.The green fluorescence protein gene led by Drosophila hsp70promoter was inserted into the Tn5transposon to construct a transposon that could be easily tracked in insect cell easily.In vit ro transposition was carried out using the transposon and AcMNPV genomic DNA to allow the random insertion of transposon into the virus genome.The transposed genome was then used to transfect insect cells S f21,and a library of mutant viruses capaple of ex 2pressing green fluorescence protein was obtained.Two mutant viruses ,B9F and Li6A ,was isolated ,and the transposon in 2sertion sites was determined within the coding region of 94K gene and p10gene ,respectively.This technology will be very useful in the functional genomics study of baculoviruses.K eyw ords :AcMNPV ;Tn5transposon ;transposase ;mutant205 复旦学报(自然科学版)第44卷 。
粘质沙雷氏菌合成灵菌红素新型调控因子的挖掘及其功能分析

究提供了新的思路。
揭示了X因子的功能
03
研究表明,X因子可以影响灵菌红素的合成途径中的多个关键
酶的活性,从而调控灵菌红素的产量和纯度。
研究不足与展望
实验条件限制
本研究的实验条件仅限于实验室环境,对于X因子的功能和作用机制仍需进一步深入研究。
缺乏对比研究
本研究未与其他已知的灵菌红素调控因子进行对比研究,未来可以开展此类对比研究以更全面地了解X因子的独特性和优势 。
抑制灵菌红素的合成
新型调控因子可能降低灵菌红素的合成量,对灵菌红素 的产量产生负面影响。
对粘质沙雷氏菌生长的影响
促进粘质沙雷氏菌的生长
新型调控因子可以加快粘质沙雷氏菌的生长 速度,提高细菌的数量。
抑制粘质沙雷氏菌的生长
新型调控因子可能会降低粘质沙雷氏菌的生 长速度,甚至导致细菌数量的减少。
对其他生物学特性的影响
灵菌红素的合成受到多个调控因子的影响,这些因子包括编码酶的基因、转录因子以及小分子调节物 等。
灵菌红素合成调控因子的种类和作用
编码酶的基因
这些基因编码一系列参与灵菌红素生物合成的酶。通过 对这些基因的表达调控,可以影响灵菌红素的产量。
转录因子
转录因子是调控基因表达的重要因子,它们可以结合到 目标基因的启动子区域,促进或抑制该基因的表达。
粘质沙雷氏菌合成灵菌红素 新型调控因子的挖掘及其功
能分析 2023-11-02
目 录
• 引言 • 粘质沙雷氏菌概述 • 灵菌红素合成调控机制研究 • 新型调控因子的挖掘 • 新型调控因子的功能分析 • 结论与展望 • 参考文献
01
引言
研究背景与意义
01
02
03
粘质沙雷氏菌是一种重要的工业微生 物,具有合成多种生物活性物质的潜 力,如灵菌红素。
一种沙雷氏菌菌株及其应用[发明专利]
![一种沙雷氏菌菌株及其应用[发明专利]](https://img.taocdn.com/s3/m/07310cf9d05abe23482fb4daa58da0116c171f82.png)
专利名称:一种沙雷氏菌菌株及其应用
专利类型:发明专利
发明人:成新跃,邹曼凌,谈延祥,鲁峻如,孙欣申请号:CN202110381544.7
申请日:20210409
公开号:CN113088467B
公开日:
20220624
专利内容由知识产权出版社提供
摘要:本发明公开了一种沙雷氏菌菌株及其应用,该菌株的分类命名为粘质沙雷氏菌Serratiamarcescens,保藏于中国微生物菌种保藏管理委员会普通微生物中心,保藏地址为北京市朝阳区北辰西路1号院3号,中国科学院微生物研究所,保藏日期为2021.03.09,保藏号为CGMCCNO.21883。
菌株从染病的马铃薯块茎蛾幼虫体内提取。
所述菌株的菌落为圆形,表面光滑,能产生红色素。
本发明的菌株用于制备生物杀虫剂,防治马铃薯块茎蛾等鳞翅目害虫。
申请人:北京师范大学
地址:100875 北京市海淀区新街口外大街19号
国籍:CN
代理机构:北京东正专利代理事务所(普通合伙)
代理人:李梦福
更多信息请下载全文后查看。
副猪嗜血杆菌EZ-Tn5转座子插入突变体库的构建及减毒株的筛选

副猪嗜血杆菌EZ-Tn5转座子插入突变体库的构建及减毒株
的筛选
贺云霞; 徐慧; 叶飞; 孙慧玲; 王宏俊; 龚玉梅; 张莉; 黄秀芬; 张培君
【期刊名称】《《农业科学与技术(英文版)》》
【年(卷),期】2011(012)002
【摘要】[目的]获得副猪嗜血杆菌减毒株.[方法]应用转座子技术构建转座子插入突变体库,卡那抗性筛选阳性菌株,PCR扩增卡那特异片段去除假阳性,小鼠感染试验检测突变株毒力,并对获得的减毒株进行生物学特性检测。
[结果]所获得减毒突变菌株具有与野毒株相似的增殖能力,传代后毒力稳定,遗传学特性稳定。
[结论]该研究结果为进一步探讨HPS毒力因子、致病机制奠定了基础。
【总页数】6页(P295-300)
【作者】贺云霞; 徐慧; 叶飞; 孙慧玲; 王宏俊; 龚玉梅; 张莉; 黄秀芬; 张培君
【作者单位】北京市农林科学院畜牧兽医研究所北京100097; 首都师范大学生命科学学院北京100048
【正文语种】中文
因版权原因,仅展示原文概要,查看原文内容请购买。
转座子Tn5诱变对三叶草根瘤菌早期侵染和结瘤的影响

转座子Tn5诱变对三叶草根瘤菌早期侵染和结瘤的影响黄世贞【期刊名称】《生物工程学报》【年(卷),期】1990(6)3【摘要】对三叶草根瘤菌Rhizobium trifolii野生型菌株ANU843及其转座子Tn5诱导的突变株nod258和nod261的侵染和结瘤能力进行了比较。
Ⅲ区结瘤基因nodFE的突变株(nod258)能在地三叶草(Subterranean clover)上正常侵染和结瘤,但瘤数比ANU843所诱导的略有减少。
而Ⅱ区结瘤基因nodIJ的突变抹(nod261)却表现侵染力减弱,诱导无效瘤和瘤数大大减少。
显微镜观察和超微结构研究表明菌株ANU843在接种后24h侵染已经从植物根毛开始,48h侵染线(infection thread)发展到表皮细胞并开始分支,接种后72h侵染线进一步发展,深入皮层细胞,这时皮层细胞已经大量分裂增生,形成瘤的分生组织。
成熟的瘤细胞内充满了类菌体。
而nod261突变株侵染植物比ANU843推迟了24h,侵染线的发展受阻碍,接种后72h侵染线仍然停留在根毛细胞中。
nod261突变株所诱导的瘤细胞内没有或仅有个别类菌体,是无效瘤。
这表明结瘤基因nodIJ与侵染线的正常形成和发展密切相关。
【总页数】6页(P212-217)【作者】黄世贞【作者单位】无【正文语种】中文【中图分类】S188【相关文献】1.利用转座子Tn5诱变冰核细菌获得无冰核活性菌株 [J], 韦建福;张世珖;马永翠2.导入三叶草根瘤菌寄主范围基因对豌豆根瘤菌侵染白三叶草的影响 [J],3.转座子Tn5对葡萄根癌病生防菌MI15的诱变 [J], 董江丽;张鹤龄;朱景奇4.百脉根根瘤菌MAFF303099 Tn5转座子插入突变体库的构建及筛选 [J], 周向珍;朱辉5.以转座子Tn5作弗氏中华根瘤菌的可识别生态学标记的研究──Tn5的水平转移及其对R.fredii Tn5突变株运动的影响 [J], 蔺继尚;崔明学;靳素英;李明祺;李宁;赵巍因版权原因,仅展示原文概要,查看原文内容请购买。
洋葱伯克霍尔德菌T1828和ZWL15-Tn5转座子插入突变体库的构建及相关突变体的筛选初探

洋葱伯克霍尔德菌T1828和ZWL15-Tn5转座子插入突变体库的构建及相关突变体的筛选初探刘好桔;钟义军;饶志强;葛岚;吴晓玉【期刊名称】《中国农学通报》【年(卷),期】2014(30)9【摘要】采用双亲结合法将含有Tn5转座子的质粒pRLl063a导入洋葱伯克霍尔德菌T1828和ZWL15中,在含有卡那霉素和链霉素的抗性平板上筛选抗性接合子。
通过随机插入诱变后,共获得300个突变株。
PCR检测随机挑选的6株突变株,结果显示为阳性;且6株突变株发酵后高效液相色谱检测不能合成冠毒素。
研究结果表明,6株突变株均已被Tn5插入染色体基因组且丧失冠毒素合成的能力。
本研究为今后转座子插入位点侧翼序列的克隆与测序,并进行功能回复试验提供了基础。
【总页数】6页(P242-247)【关键词】洋葱伯克霍尔德菌;冠毒素;Tn5转座子;突变体库【作者】刘好桔;钟义军;饶志强;葛岚;吴晓玉【作者单位】江西农业大学生物科学与工程学院;南昌市发酵应用技术重点实验室;江西省红壤研究所;国家红壤改良工程技术研究中心【正文语种】中文【中图分类】S182【相关文献】1.大丽轮枝菌菌核型菌株T-DNA插入突变体库的构建及微菌核发育异常突变体的筛选 [J], 梁曼;邓晟;张昕;林玲2.热稳定性伯克霍尔德菌脂肪酶A突变体的筛选 [J], 刘艳如;邱黎清;黄建忠;赵丙春;王作镇;朱晓兰;高媛媛;舒正玉3.稻瘟病菌T-DNA插入突变体库构建及致病相关突变体筛选 [J], 贺春萍;林春花;廖奇亨;李锐;郑服丛4.水稻基腐细菌Tn5插入突变体库的构建及其致病相关突变体的筛选 [J], 谭秀明;陈雪风;林敏怡;刘琼光5.百脉根根瘤菌MAFF303099 Tn5转座子插入突变体库的构建及筛选 [J], 周向珍;朱辉因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
沙雷氏Z4菌株Tn5转座突变株的群体淬灭活性筛选体系的
建立及优化
杨惠婷;刘栋;赵一竹;杨瑞雪;葛世玫
【期刊名称】《温州大学学报:自然科学版》
【年(卷),期】2022(43)2
【摘要】利用Tn5转座子插入突变技术构建沙雷氏Z4菌株突变体库,研究不同因素对转座突变的影响,在此基础上初步探究了突变株的群体淬灭能力.结果表明,采用双亲本接合法,选择对数生长期的沙雷氏Z4菌株与大肠杆菌S17-1(pRL1063a)菌液1∶1混合,30℃培养3 h后,涂布于含50μg/mL卡那霉素和25μg/mL四环素的LB固体培养基上,长出的突变株数量最多;随机选择突变株进行群体淬灭能力测试,结果筛选到了3株群体淬灭能力明显与沙雷氏Z4野生型有差异的突变株.本研究采用的Tn5转座突变技术为研究群体淬灭菌的基因功能以及生理代谢提供了有效的分子遗传工具,也为研究群体淬灭提供了新的途径.
【总页数】10页(P44-53)
【作者】杨惠婷;刘栋;赵一竹;杨瑞雪;葛世玫
【作者单位】温州大学生命与环境科学学院
【正文语种】中文
【中图分类】X172
【相关文献】
1.青枯雷尔氏菌TN5转座子无致病力突变株插入位点的鉴定与分析
2.高效离子交换色谱法分析青枯雷尔氏菌Tn5转座子突变菌株的异质性
3.从芽胞杆菌dhs-330转座突变库筛选获得高产表面活性剂突变株
4.青枯雷尔氏菌Tn5转座子无致病力突变株构建及其生物学特性
5.以转座子Tn5作弗氏中华根瘤菌的可识别生态学标记的研究──Tn5的水平转移及其对R.fredii Tn5突变株运动的影响
因版权原因,仅展示原文概要,查看原文内容请购买。